Introduction
Pulmonary arterial hypertension (PAH) is a
progressive disease associated with the excessive proliferation of
pulmonary vascular smooth muscle cells (pVSMCs) and pulmonary
vascular endothelial cells (pVECs), deposition of the extracellular
matrix (ECM) and ECM remodeling, which results in a persistent
increase in pulmonary arterial pressure with vascular remodeling
and organ fibrosis.
It has been previously reported that the excessive
proliferation of pVSMCs and pVECs is mediated by various signaling
molecules, including protein kinase B (Akt) and transforming growth
factor (TGF)β (1). The
Akt/mechanistic target of rapamycin (mTOR) signaling pathway is
associated with the differentiation of myofibroblasts and
extracellular remodeling, which are critical for organ fibrosis.
mTOR signaling promotes cell proliferation and is essential for
hypoxia-mediated pVSMC or pVEMC proliferation and angiogenesis
(2). TGFβ may induce pulmonary
fibrosis (3) and activate the
phosphoinositide 3-kinase (PI3K)/Akt signaling pathway, which
enhances fibroblast proliferation and the production of collagen
(4). Lu et al (4) previously demonstrated that the
inhibition of PI3K abrogated the effects of bleomycin, a fibrosis
inducer that induces rapid lung fibrosis and activation of the
PI3K/Akt signaling pathway, on fibroblast proliferation and
collagen production.
Phosphoinositide-dependent protein kinase-1 (PDK1)
serves a key role in the activation of the AGC subfamily of protein
kinases, including Akt (5). A
previous study revealed that PDK1-dependent AGC kinase activation
is a requirement of Akt oncogenic activity (6). In addition, PDK1-dependent metabolic
reprogramming dictates the metastatic potential of breast cancer
cells (7). PDK1 is associated with
the proliferation of cancer and basilar artery smooth muscle cells
by Akt-mediated signals (8–10). Yu et al (11) recently revealed that tanshinone IIA
induced the inactivation of PDK1 as well as the development of
hypertension. Previous studies indicated that PDK1 is a
hypoxia-responsive protein, which serves an important role in
stress responses (12,13). PDK1 deficiency results in heart
failure and increased hypoxia sensitivity in cardiac muscle
(14). However, to the best of our
knowledge, the association between PDK1 and PAH has not yet been
determined.
The present study aimed to investigate the effect of
PDK1 on the development of PAH. A mouse model of hypoxia-induced
PAH was generated using PDK1flox/+: Tie2-Cre mice
or control mice. The activation of the PDK1/Akt signaling pathway
was detected by western blot; histological and hemodynamic analyses
were also performed. The association between the PDK1-mediated Akt
signaling pathway and hypoxia-induced PAH progression was
evaluated.
Materials and methods
Animals and treatment
A total of 25 C57/BL6 male mice (age, 8 weeks;
weight, 20–30 g) were purchased from Model Animal Research Center,
Nanjing University (Nanjing, China) and maintained under
stress-free conditions at 23±2°C with a humidity of 70% and a 12-h
light/dark cycle with food and water provided ad libitum.
The PAH model was induced by hypoxia exposure in hypobaric
chambers. The experimental animals (C57/BL6 mice,
PDK1flox/+ mice and PDK1flox/+: Tie2-Cre
mice; n=15, respectively) were exposed to chronic hypobaric hypoxia
(0.5 atm, 10% O2, 50% humidity and 5% CO2 at
23±2°C) in chambers for 21 days and were let out three times per
week as previously described (15).
The control animals (C57/BL6 mice, PDK1flox/+ mice and
PDK1flox/+: Tie2-Cre mice; n=10, respectively) were
exposed to normobaric chambers (room air) over the same 21-day
period. Mice were anesthetized and received surgery following 21
days hypoxia exposure and pulmonary artery tissues were obtained
following euthanasia by cervical dislocation and prepared for
western blot or reverse transcription-quantitative polymerase chain
reaction (RT-qPCR) analysis. The Institutional Animal Care and Use
Committee of Shanghai Fifth People's Hospital approved the present
study.
Generation of PDK1 partial knockout
mice in endothelial cells
As the PDK1flox/flox: Tie2-Cre
mice (complete PDK1 knockout in endothelial cells) succumbed during
the embryonic period, the PDK1flox/+: Tie2-Cre
(PDK1 partial knockout in endothelial cells) mice were generated by
hybridization of PDK1flox/flox mice and
Tie2-Cre mice (with Cre-expressing endothelial cells), which
were provided by Model Animal Research Center, Nanjing University.
The PDK1 genotyping was performed by PCR using DNA extracted from
the tails of mice 7 days after birth. Different groups of mice,
including C57BL6 mice, PDK1flox/+ mice (n=25) and
PDK1flox/+: Tie2-Cre mice (n=25) were selected
for further study.
Semi-quantitative PCR
Genomic DNA was extracted from individual samples
(200 µl) using the DNeasy tissue kit (cat. no. 69506; Qiagen, Inc.,
Valencia, CA, USA) following the manufacturer's protocol and eluted
in 100 µl. The percentage of agarose gel used in semi-quantitative
PCR was 2% and samples were visualized using ethidium bromide. The
primers of PDK1 were as follows: Forward,
5′CTCTACCTCCACCATGCCAAGT3′ and reverse,
5′GCTGCGCTGATAGACATCCA3′.
The protocol was as follows: For primary
amplification (15 cycles of 10 sec at 95°C; 20 sec at 60°C and 20
sec at 72°C), 5 µl of genomic DNA representing each test-sample or
5 µl of water (negative control) were dispensed into 0.2 ml PCR
strips and placed into a 24-well thermocycling block within the
Gene-Plex robotic platform (Applied Biosystems™; Thermo Fisher
Scientific, Inc.). Following the dispensing of each sample and the
initiation of the assay, the following set-up process and analysis
were executed by the program Assay Setup (cat. no. 9001550; Qiagen,
Inc.), with the secondary amplification in PCR and the melting
curve analysis being semi-automated.
Hemodynamic measurements
After 21 days following PAH induction the mice were
anesthetized. Right ventricular (RV) pressures were measured using
a high-fidelity pressure sensor catheter inserted directly into the
right ventricle. Pressure waveforms were recorded for 2 min for
each mouse using the PowerLab Chart 5 version 5.3 data acquisition
system and analysis software. RV systolic pressures (RVSPs) were
calculated by averaging ≥20 cardiac cycles for each mouse. The RV
weight-to left ventricular (LV) plus septum (S) weight ratio
(RV/LV+S) was calculated as the right ventricle hypertrophy index
(RVHI).
Histology
The histology of PAH mice was detected using
hematoxylin and eosin (H&E) staining as previously described
(16,17). Briefly, the pulmonary artery tissues
were perfused, inflated, separated and fixed with 4% formaldehyde
at room temperature for 24 h, embedded in paraffin, hydrated and
sectioned (thickness, 5 µm) using a Leica RM2255 rotary paraffin
section machine (Leica Microsystems GmbH, Wetzlar, Germany) at room
temperature with a humidity of 70%. Tissue sections were then dried
at 60°C, deparaffinizated by xylene and rehydrated by gradient of
ethanol (100, 95, 85 and 70%) at room temperature for 5 min.
Hematoxylin (Sigma-Aldrich; Merck KGaA) staining (100 µl) was
performed for 5 min at room temperature after the section was
rinsed using distilled water. The section was then rinsed with PBS.
Eosin (Merck KGaA) staining was performed for 3–5 min at room
temperature and then sections were rinsed using distilled water.
Gradient alcohol (70, 85, 95 and 100%) was adopted for dehydration
for 5 min at each gradient. H&E-staining was quantitatively
evaluated by a point-counting technique in 10 randomly selected
microscopic fields, at a final magnification of ×200 under a
100-point grid by light microscopy. Blind analysis was performed on
all sections by the same observer.
Western blot analysis
Pulmonary tissue was homogenized, lysed in a
radioimmunoprecipitation buffer (Amyjet Scientific, Wuhan, China)
and centrifuged to obtain the supernatant. Proteins were separated
using 10% SDS-PAGE. Total protein (20 µg/lane) was then separated
on 10% SDS polyacrylamide gel and blotted onto HyBond N membranes
(EMD Millipore, Billerica, MA, USA). Blocking was then performed
with 5% skimmed milk solution in TBS with 0.1% Tween-20 for 2 h at
room temperature. The membranes were incubated with primary
antibodies at 4°C overnight against PDK1, phosphorylated
(p)-AktT308, Akt, ribosomal protein S6 kinase (p70S6K;
cat. no. ab8811; 1:1,000), p-p70S6K, (cat. no. ab8892; 1:1,000),
proline-rich Akt1 substrate 1 (PRAS40; cat. no. ab3323; 1:1,000),
p-PRAS40 (cat. no. ab5505; 1:1,000), p-S6KT229 (cat. no. ab1223;
1:1,000), p-S6T40/244 (cat. no. ab4435; 1:1,000) and S6 (cat. no.
ab2334; 1:1,000) (all from Abcam, Cambridge, UK). All antibodies
were purchased from Cell Signaling Technologies Inc. (Beverly, MA,
USA) β-actin (cat. no. ab8227; 1:1,000; Abcam) was used as the
internal reference protein. Membranes were then incubated with
horseradish peroxidase-conjugated goat anti-mouse immunoglobulin G
secondary antibodies (cat. no. ab6789; 1:5,000; Abcam) for 2 h at
room temperature. The antigen-antibody complexes were visualized
using an enhanced chemiluminescence detection plus kit (cat. no.
PE0010; EMD Millipore) according to the manufacturer's
protocol.
RT-qPCR
Total RNA was extracted from mice lung vessels using
TRIzol® (Invitrogen; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA) and first-strand cDNA was synthesized at 37°C for
15 min (cat. no. 00345; Qiagen, Inc.). The mRNA expression of PDK1
was determined using a SYBR Premix Ex Taq™ kit (Takara Bio, Inc.,
Otsu, Japan) on an ABI 7500 fast real time PCR platform (Applied
Biosystems; Thermo Fisher Scientific, Inc.). The following
thermocycling conditions were used: 95°C for 3 min, 40 cycles of
95°C for 30 sec, 60°C for 30 sec and 72°C 10 sec, followed by 65°C
for 3 min. The relative expression level was calculated using the
2−ΔΔCq method (18). All
reactions were run in triplicate. β-actin was used as the internal
reference gene for mRNA detection. The forward and reverse primers
were as following: PDK1, forward, CTGTGATACGGATCAGAAACCG and
reverse, TCCACCAAACAATAAAGAGTGCT; β-actin forward,
GGTGTGGCATCAGGATTCAAG and reverse, TTTCATACCGATTGCTGTTGGA.
Statistical analysis
Statistical analyses were performed using SPSS 17.0
software (SPSS, Inc., Chicago, IL, USA). All data are expressed as
the mean + standard deviation from three independent experiments
with each measured in triplicate. Differences between two groups
and among three groups were assessed by Student's t-test and
one-way analysis of variance, respectively. A post-hoc test
(Tukey's test) was performed following ANOVA. P<0.05 was
considered to indicate a statistically significant difference.
Results
PDK1 is upregulated in lung tissue in
a mouse model of PAH
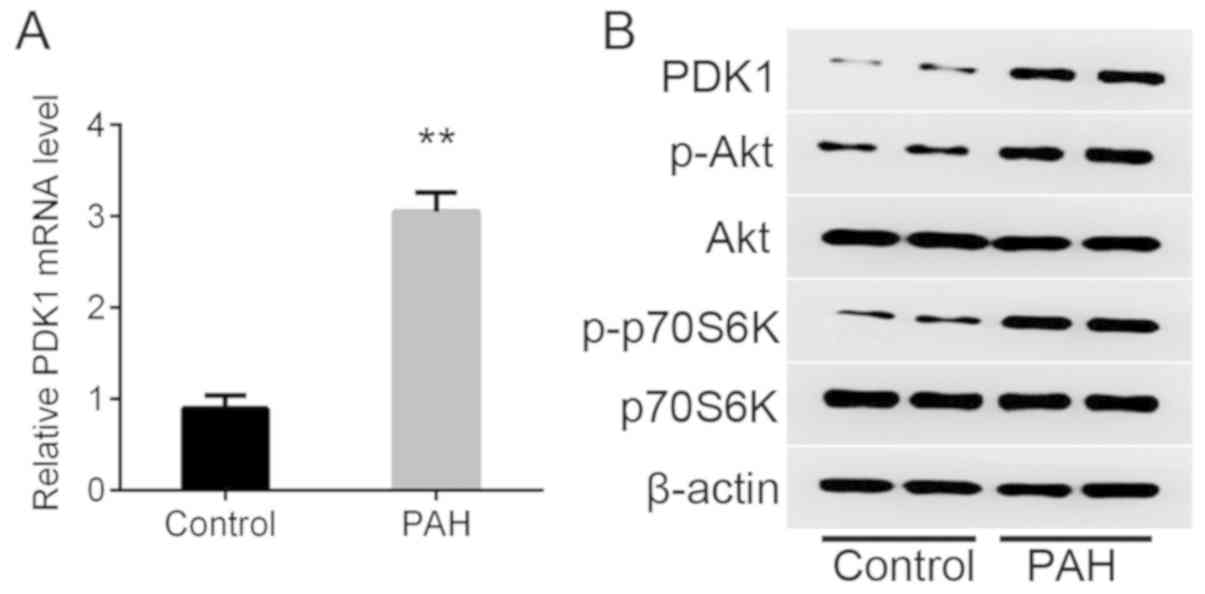
PDK1 mRNA was significantly upregulated in a mouse
model of hypoxia-induced PAH compared with the control as detected
by RT-qPCR (Fig. 1A). The protein
expression of PDK1 was increased in the PAH group compared with the
control as determined by western blot analysis (Fig. 1B). It was also observed that p-Akt
and p-p70S6K were increased markedly in the PAH group compared with
the control, which indicated that the PDK1/Akt/p70S6K signaling
pathway was activated.
Generation of PDK1f/+
genotype mice
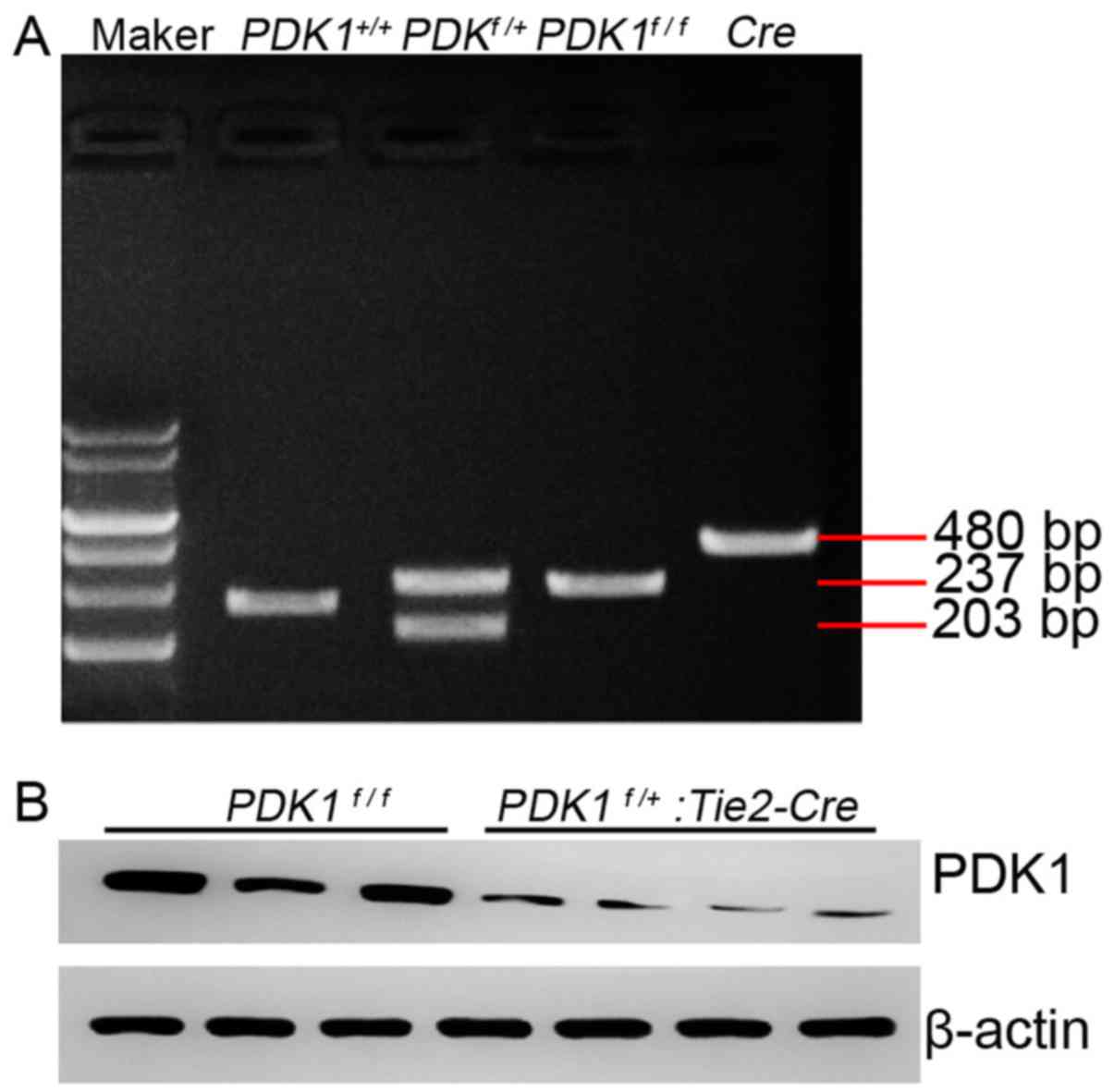
PDK1flox/flox male mice and
Tie2-Cre female mice were hybridized and the tails were
obtained 7 days after birth. sqPCR was performed and the embryos
with different PDK1 genotypes were revealed (Fig. 2A). Mice with the
PDK1f/+ genotype had clear bi-stripe features
(Fig. 2A). The protein expression of
PDK1 in PDK1 knockout mice was notably reduced compared with the
wild type mice (Fig. 2B), which
indicated that the PDK1f/+: Tie2-Cre mice were
successfully established.
PDK1f/+ genotype reduces
hypoxia-induced PAH in mice
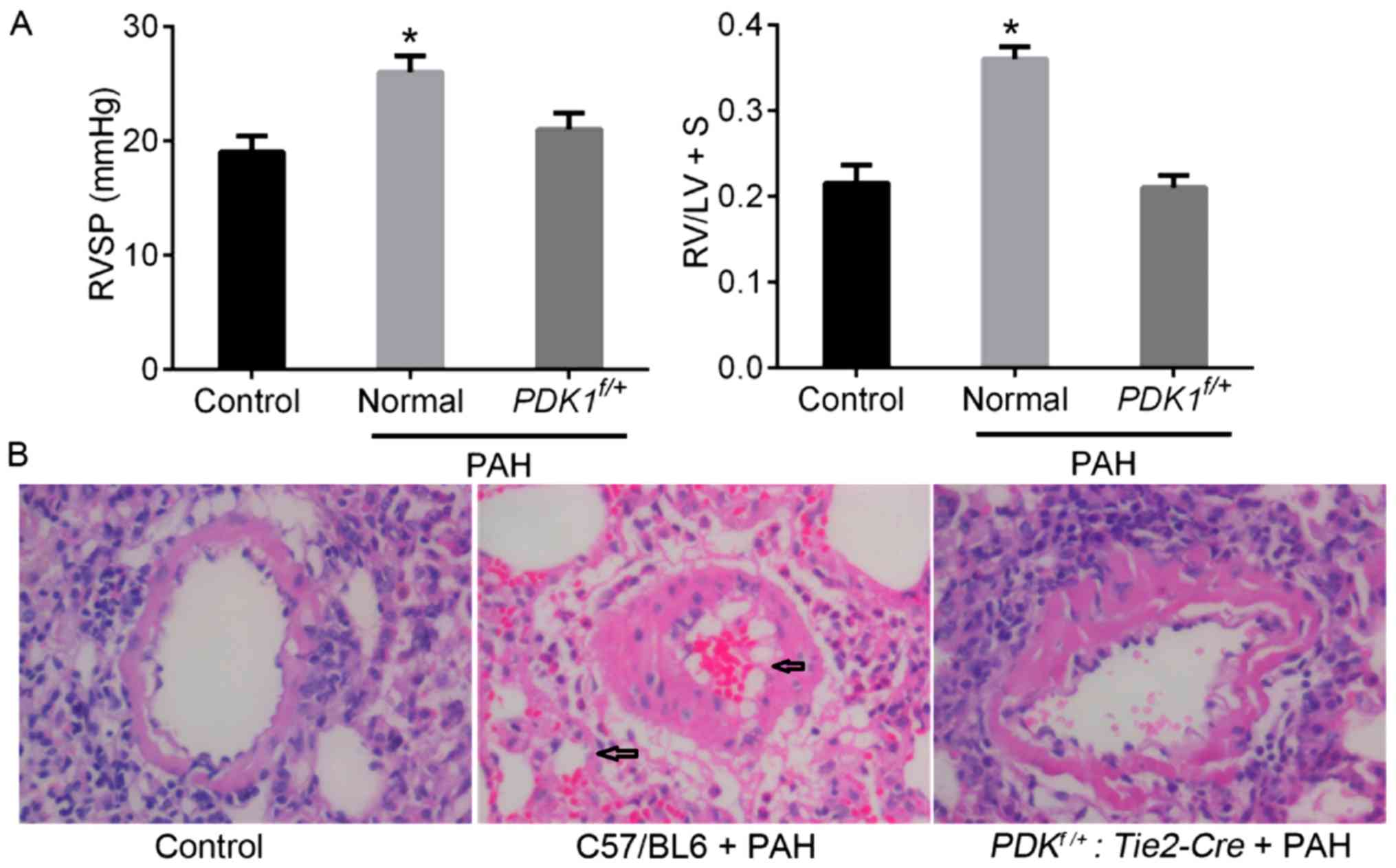
After 21 days following PAH induction RVSP and RVHI
(RV/LV+S) were significantly increased in the experimental group
compared with the control (Fig. 3A).
Hypoxia induced a notable increase in the thickness of the lung
vessels and the proportion of muscularized pulmonary vessels (as
indicated by black arrows in Fig.
3B). However, the partial deletion of PDK1 (PDK1f/+:
Tie2-Cre mice) reduced hypoxia-induced damage to the pulmonary
vessels, resulting in notably lower RVSP and RVHI values compared
with the control mice. The histology of the PDK1f/+ mice
was similar to that of the control group. These results
demonstrated that the PDK1f/+ genotype may limit
the procession of hypoxia-induced PAH in mice, potentially by
reducing pulmonary vascular remodeling.
PDK1f/+ genotype reduces
hypoxia-induced activation of the PDK1/Akt/p70S6K signaling
pathway
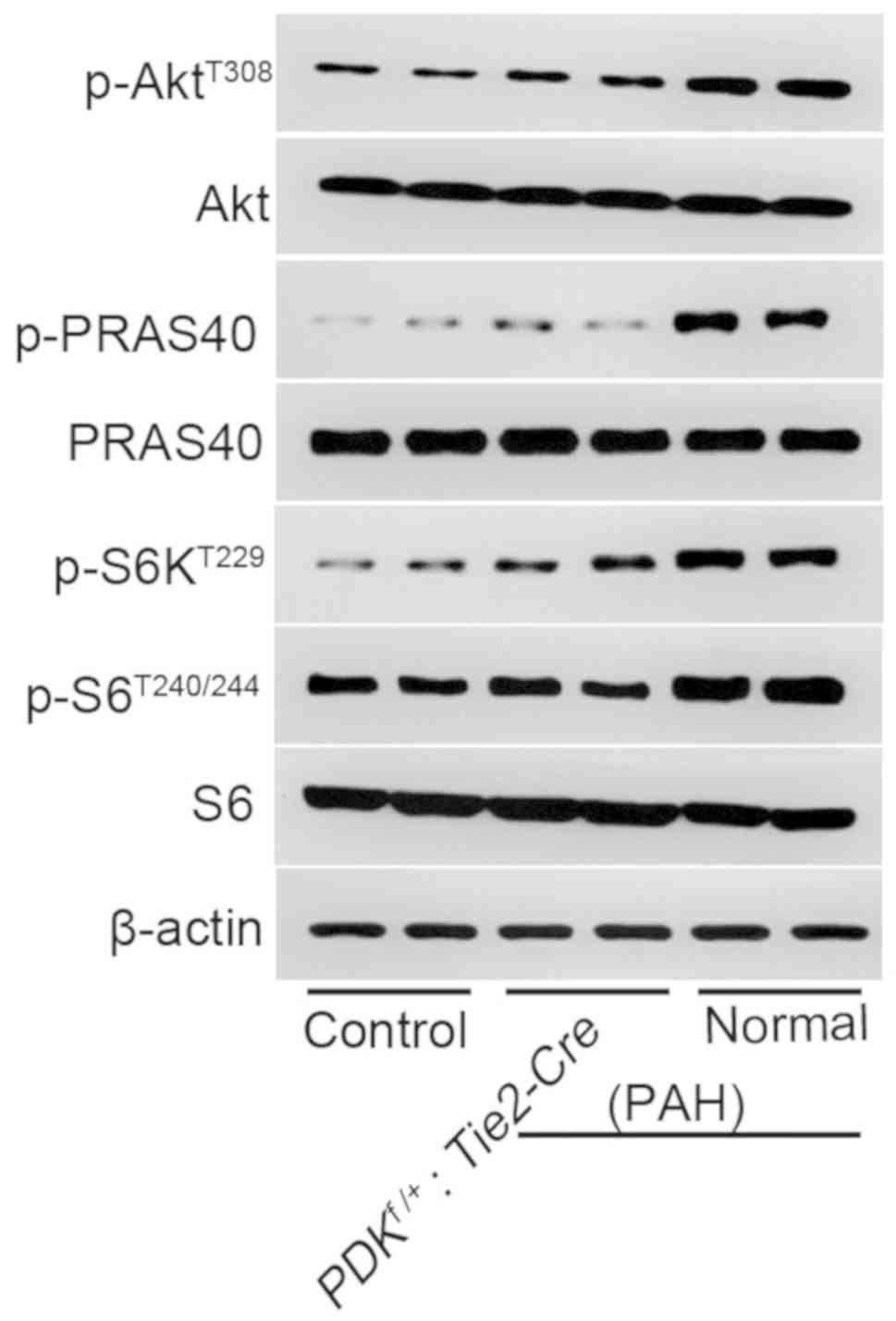
The phosphorylation levels of AktT308,
PRAS40, S6KT229 and S6T240/244 were
upregulated in the pulmonary vessels of the PDK1f/+ PAH
group (Fig. 4). This suggested that
the PDK1/Akt/p70S6K signaling pathway was activated. By contrast,
the phosphorylation of these proteins in the
PDK1f/+ group was like the control group and
notably reduced compared with the normal PAH group. This indicated
that the partial knockout of PDK1 protected against the
hypoxia-induced activation of the PDK1/Akt/p70S6K signaling pathway
in mice.
Discussion
PDK1 serves an important role in activating the AGC
subfamily of protein kinases, including Akt, which is essential to
PAH as well as pulmonary and liver fibrosis (18–20). The
results of the present study indicated that PDK1/Akt signaling
promoted PAH and the partial knockout of PDK1 reduced
hypoxia-induced Akt activation and PAH procession.
PAH is characterized by excessive proliferation of
pVSMC and pVEC, which results in pulmonary pressure (21). The activation of Akt signaling
promotes cell proliferation and survival, which accelerates disease
procession and the metastasis of various types of tumor (21–24).
Allard et al (24)
demonstrated that the activation of Akt promoted their survival of
VSMCs and inhibited the formation of intimal plaques in
atherosclerosis. Tang et al (21) recently suggested that the activation
of the Akt/mTOR signaling pathway and the knockout of Akt
significantly reduced hypoxia-induced PAH and vascular remodeling
in a mouse model. The present study demonstrated that hypoxia
significantly upregulated the expression of PDK1 and notably
upregulated the phosphorylation of Akt, whereas knockout of PDK1
notably reduced Akt phosphorylation and hypoxia-induced PAH. These
results indicated that PDK1 is essential for hypoxia-induced PAH,
which is associated with the activation of Akt signals.
PDK1 phosphorylates AktT308, which
subsequently activates Akt signaling pathways, including the Akt
survival pathway (25). PRAS40 (40
kDa) is a novel mTOR binding partner, which serves essential roles
in the transmission of Akt signaling by mediating Akt signals to
the mTORC1 kinase domain, thus inhibiting mTOR signalling (26). The Akt/mTOR signaling pathway serves
important roles in extracellular remodelling (19) as well as contributing to fibrosis in
non-alcoholic steatohepatitis (27)
and cancer cell growth and proliferation (28,29) by
crosstalk with signals, including p38/mitogen activated protein
kinase, 40S ribosomal protein S6 and extracellular signal-regulated
kinase (30–32). Activation of mTOR signaling may be
induced by hypoxia in vascular smooth muscle and endothelial cells.
In addition, silencing of PRAS40 inactivated Akt and uncouples the
Akt/mTOR signaling pathway. Humar et al revealed that the
activation of mTOR was essential for hypoxia-mediated VSMC
proliferation and angiogenesis. However, Tang et al
demonstrated that mTOR was not essential for Akt
activation-mediated regulation of hypoxia-induced PAH. These
contradicting results mean that any association between the
mTOR/p70S6K signaling pathway and PAH remains unclear. The present
study demonstrated that the phosphorylation of Akt, PRAS40 and
p70S6K was increased in hypoxia-induced PAH mice. However, the
partial knockout of PDK1 in PDK1f/+ mice reduced
the phosphorylation of these proteins and PAH procession, which
indicated that the PDK1/Akt/PRAS40/mTOR/p70S6K signaling pathway
contributed to hypoxia-induced PAH in a mouse model.
In conclusion, the present study determined that
PDK1 was essential for hypoxia-induced PAH, which was mediated by
the PDK1/Akt/PRAS40/mTOR/p70S6K signaling pathway. However, it
remains unclear whether the interaction or activation of the mTOR
mediated signaling pathway was necessary for transmitting
PDK1-mediated PAH in a mouse model. However, to some extent, this
study provides useful strategies in the foreseeable future and
sheds some lights in clinical diagnosis and treatment of PAH.
Acknowledgements
Not applicable.
Funding
The study was funded by the Shanghai Science and
Technology Commission (grant no. 14ZR1432800) and the Scientific
Research Project of Minhang District Health and Family Planning
Commission of Shanghai (grant no. 2012MW02).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
RD wrote the manuscript. ZY and YX for the
discussion of and PX for discussion and comments on an earlier
version of the manuscript. RD designed the experiments. ZY and YX
performed the experiments and acquired the data. PX analyzed the
data and prepared the manuscript. All authors read and approved the
final manuscript.
Ethics approval and consent to
participate
The present study was approved by the Research
Ethics Committee of Shanghai Fifth People's Hospital.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Sankhe S, Manousakidi S, Antigny F, Arthur
Ataam J, Bentebbal S, Ruchon Y, Lecerf F, Sabourin J, Price L,
Fadel E, et al: T-type Ca2+ channels elicit
pro-proliferative and anti-apoptotic responses through impaired
PP2A/Akt1 signaling in PASMCs from patients with pulmonary arterial
hypertension. Biochim Biophys Acta. 1864:1631–1641. 2017.
View Article : Google Scholar
|
|
2
|
Humar R, Kiefer FN, Berns H, Resink TJ and
Battegay EJ: Hypoxia enhances vascular cell proliferation and
angiogenesis in vitro via rapamycin (mTOR)-dependent signaling.
FASEB J. 16:771–780. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Xia H, Diebold D, Nho R, Perlman D,
Kleidon J, Kahm J, Avdulov S, Peterson M, Nerva J, Bitterman P and
Henke C: Pathological integrin signaling enhances proliferation of
primary lung fibroblasts from patients with idiopathic pulmonary
fibrosis. J Exp Med. 205:1659–1672. 2015. View Article : Google Scholar
|
|
4
|
Lu Y, Azad N, Wang L, Iyer AK, Castranova
V, Jiang BH and Rojanasakul Y: Phosphatidylinositol-3-kinase/akt
regulates bleomycin-induced fibroblast proliferation and collagen
production. Am J Respir Cell Mol Biol. 42:432–441. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Sato S, Fujita N and Tsuruo T:
Interference with PDK1-Akt survival signaling pathway by UCN-01
(7-hydroxystaurosporine). Oncogene. 21:1727–1738. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Orlacchio A, Arciuch VA and Cristofano AD:
PDK1-dependent activation of AGC kinases is an absolute requirement
for AKT oncogenic activity. Cancer Res. 74 (Suppl 19):4776.
2014.
|
|
7
|
Dupuy F, Tabariès S, Andrzejewski S, Dong
Z, Blagih J, Annis MG, Omeroglu A, Gao D, Leung S, Amir E, et al:
PDK1-dependent metabolic reprogramming dictates metastatic
potential in breast cancer. Cell Metab. 22:577–589. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Li W, Qian L, Lin J, Huang G, Hao N, Wei
X, Wang W and Liang J: CD44 regulates prostate cancer
proliferation, invasion and migration via PDK1 and PFKFB4.
Oncotarget. 8:65143–65151. 2017.PubMed/NCBI
|
|
9
|
Fang B, Zhu J, Wang Y, Geng F and Li G:
MiR-454 inhibited cell proliferation of human glioblastoma cells by
suppressing PDK1 expression. Biomed Pharmacother. 75:148–152. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Liu Y, Yang K, Sun X, Fang P, Shi H, Xu J,
Xie M and Li M: MiR-138 suppresses airway smooth muscle cell
proliferation through the PI3K/AKT signaling pathway by targeting
PDK1. Exp Lung Res. 41:363–369. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yu ZL, Wang JN, Wu XH, Xie HJ, Han Y, Guan
YT, Qin Y and Jiang JM: Tanshinone IIA prevents rat basilar artery
smooth muscle cells proliferation by inactivation of PDK1 during
the development of hypertension. J Cardiovasc Pharmacol Ther.
20:563–571. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Mora A, Davies AM, Bertrand L, Sharif I,
Budas GR, Jovanović S, Mouton V, Kahn CR, Lucocq JM, Gray GA, et
al: Deficiency of PDK1 in cardiac muscle results in heart failure
and increased sensitivity to hypoxia. EMBO J. 22:4666–4676. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Belgardt BF, Husch A, Rother E, Ernst MB,
Wunderlich FT, Hampel B, Klöckener T, Alessi D, Kloppenburg P and
Brüning JC: PDK1 deficiency in POMC-expressing cells reveals
FOXO1-dependent and-independent pathways in control of energy
homeostasis and stress response. Cell Metab. 7:291–301. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Papandreou I, Cairns RA, Fontana L, Lim AL
and Denko NC: HIF-1 mediates adaptation to hypoxia by actively
downregulating mitochondrial oxygen consumption. Cell Metab.
3:187–197. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ciuclan L, Bonneau O, Hussey M, Duggan N,
Holmes AM, Good R, Stringer R, Jones P, Morrell NW, Jarai G, et al:
A novel murine model of severe pulmonary arterial hypertension. Am
J Respir Crit Care Med. 184:1171–1182. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Song Y, Coleman L, Shi J, Beppu H, Sato K,
Walsh K, Loscalzo J and Zhang YY: Inflammation, endothelial injury,
and persistent pulmonary hypertension in heterozygous BMPR2-mutant
mice. Am J Physiol Heart Circ Physiol. 295:H677–H690. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Song Y, Jones JE, Beppu H, Keaney JF Jr,
Loscalzo J and Zhang YY: Increased susceptibility to pulmonary
hypertension in heterozygous BMPR2-mutant mice. Circulation.
112:553–562. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wang J, Chu ES, Chen HY, Man K, Go MY,
Huang XR, Lan HY, Sung JJ and Yu J: microRNA-29b prevents liver
fibrosis by attenuating hepatic stellate cell activation and
inducing apoptosis through targeting PI3K/AKT pathway. Oncotarget.
6:7325–7338. 2015.PubMed/NCBI
|
|
19
|
Abdalla M, Sabbineni H, Prakash R, Ergul
A, Fagan SC and Somanath PR: The Akt inhibitor, triciribine,
ameliorates chronic hypoxia-induced vascular pruning and
TGFβ-induced pulmonary fibrosis. Br J Pharmacol. 172:4173–4188.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Larson-Casey JL, Murthy S, Ryan AJ and
Carter AB: Modulation of the mevalonate pathway by akt regulates
macrophage survival and development of pulmonary fibrosis. J Biol
Chem. 289:36204–36219. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Tang H, Chen J, Drennan AR, Fraidenburg
DR, Song S, Sysol JR, Smith KA, Machado RF, Makino A and Yuan JX:
Akt/mTOR signaling contributes to the development of pulmonary
arterial hypertension, in D27. Preclinical models of pulmonary
hypertension: Novel targets and pathways. Am Thoracic Soc.
189:A55592014.
|
|
22
|
Dai W, Xu X, Li S, Ma J, Shi Q, Guo S, Liu
L, Guo W, Xu P, He Y, et al: SOX4 promotes proliferative signals by
regulating glycolysis through AKT activation in melanoma cells. J
Invest Dermatol. 137:2407–2416. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Dong Y, Liang G, Yuan B, Yang C, Gao R and
Zhou X: MALAT1 promotes the proliferation and metastasis of
osteosarcoma cells by activating the PI3K/Akt pathway. Tumor Biol.
36:1477–1486. 2015. View Article : Google Scholar
|
|
24
|
Allard D, Figg N, Bennett MR and
Littlewood TD: Akt regulates the survival of vascular smooth muscle
cells via inhibition of FoxO3a and GSK3. J Biol Chem.
283:19739–19747. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Sato S, Fujita N and Tsuruo T:
Interference with PDK1-Akt survival signaling pathway by UCN-01
(7-hydroxystaurosporine). Oncogene. 21:1727–1738. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Vander Haar E, Lee SI, Bandhakavi S,
Griffin TJ and Kim DH: Insulin signalling to mTOR mediated by the
Akt/PKB substrate PRAS40. Nat Cell Biol. 9:316–323. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Cai CX, Buddha H, Castelino-Prabhu S,
Zhang Z, Britton RS, Bacon BR and Neuschwander-Tetri BA: Activation
of insulin-PI3K/Akt-p70S6K pathway in hepatic stellate cells
contributes to fibrosis in nonalcoholic steatohepatitis. Dig Dis
Sci. 62:968–978. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Seufferlein T and Rozengurt E: Rapamycin
inhibits constitutive p70s6k phosphorylation, cell proliferation,
and colony formation in small cell lung cancer cells. Cancer Res.
56:3895–3897. 2004.
|
|
29
|
Syed DN, Chamcheu JC, Khan MI, Sechi M,
Lall RK, Adhami VM and Mukhtar H: Fisetin inhibits human melanoma
cell growth through direct binding to p70S6K and mTOR: Findings
from 3-D melanoma skin equivalents and computational modeling.
Biochem Pharmacol. 89:349–360. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Chai X, Chu H, Yang X, Meng Y, Shi P and
Gou S: Metformin increases sensitivity of pancreatic cancer cells
to gemcitabine by reducing CD133+ cell populations and
suppressing ERK/P70S6K signaling. Scientific Reports. 5:144042015.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Li G, Shan C, Liu L, Zhou T, Zhou J, Hu X,
Chen Y, Cui H and Gao N: Tanshinone IIA inhibits HIF-1α and VEGF
expression in breast cancer cells via mTOR/p70S6K/RPS6/4E-BP1
signaling pathway. PLoS One. 10:e01174402015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Conejo R, de Alvaro C, Benito M, Cuadrado
A and Lorenzo M: Insulin restores differentiation of
Ras-transformed C2C12 myoblasts by inducing NF-kappaB through an
AKT/P70S6K/p38-MAPK pathway. Oncogene. 21:3739–3753. 2002.
View Article : Google Scholar : PubMed/NCBI
|