Introduction
Multiple sclerosis (MS) is a disease of the central
nervous system (CNS) characterized by multifocal demyelination
along with astrocyte gliosis. These processes affect several
structures in the CNS, resulting in severe neurological deficits
(1). It has been reported that
43–70% of patients with MS experience cognitive deficits (2). Cognitive impairment in patients with MS
is variable depending on the disease stage and type of clinical
course (3). MS-associated cognitive
impairment is has a negative effect on the quality of life of
patients (4), and makes them less
likely to be employed or actively engaged in daily activities
(5). Although the causes of
cognitive impairment in patients MS are currently unclear,
depression is believed to be a contributing factor (6–8).
Depression often burdens people with MS-related cognitive
dysfunction (8), and lifetime
prevalence of major depression in MS patients is as high as 50%
(9). Amelioration of depression has
been suggested as a mean to improve cognitive functioning in MS.
Factors that can contribute to the reduction of depression include
social, physical and cognitive stimulation. The effect of these
factors is mimicked in laboratory animals housed in an enhanced
environment (EE), which has been shown to improve brain development
and memory in both normal and pathological conditions (10,11).
The expression of brain-derived neurotrophic factor
and tyrosine kinase receptor B, key neuronal differentiation and
survival factors, is increased with exposure to EE (12). A host of studies have strengthened
the notion that EE is beneficial to a variety of neurodegenerative
diseases by protecting against cognitive impairment and improving
cognitive performance in Alzheimer's and Huntington's mouse models,
respectively (13). Upon studying
the effect of EE on experimental autoimmune encephalomyelitis in
rodents, a model of MS, results favored functional recovery
(14). However, there is
insufficient data to draw a conclusion regarding the effect of EE
on MS mouse models.
The cuprizone-induced demyelination model has
attracted increasing interest during the last decade as a method of
inducing MS (15). Indeed,
cuprizone-induced demyelination in distinct brain regions,
including the corpus callosum, is the most frequently investigated
white matter tract in animal models (15,16). The
addition of 0.2% cuprizone (bis-cyclohexanone oxaldihydrazone), a
copper chelating agent, to the diet of male C57BL/6 strain mice
induces spatially and temporally well-defined histopathological
alterations in the CNS (17).
Furthermore, cuprizone-fed animals in the cuprizone model
experience oligodendrocyte death and steady demyelination (18). This demyelination effect was evident
5–6 weeks after cuprizone induction (18,19),
whereas remyelination occurred two weeks after the discontinuation
of cuprizone. In addition, cuprizone-induced toxicity has been
widely used to identify treatments for demyelinating diseases
(15,20–22).
The present study investigated the impact of induced
depression on a cuprizone mouse model of demyelination and the
effectiveness of EE as a method of intervention.
Materials and methods
Animals
Male C57BL/6 strain mice were bred in the animal lab
of Arabian Gulf University. All animal protocols have been approved
by the Ethical Committee for Animal Experiments of Arabian Gulf
University.
Housing conditions
Three-week-old mice were housed in either standard
housing or EE conditions for a period of 9 weeks. Standard cages
(33×15×13 cm), containing 2–3 mice per cage, were cleaned twice a
week. All mice were maintained on a 12-h light/dark cycle and fed
ad libitum. All tests were performed during the dark cycle.
Experimental design
Experimental group
Mice were subdivided into five groups, based on the
intervention received. A total of 46 mice, dived into 5 groups,
were tested: i) Cuprizone mouse model with no intervention (Cup-O;
n=9); ii) cuprizone mouse model undergoing induced depression
(Cup-Dep; n=9); iii) cuprizone mouse model housed in an EE (Cup-EE;
n=10); iv) cuprizone mouse model housed in an EE and undergoing
induced depression (Cup-ED; n=9), and v) a control group, (n=9),
which was neither injected with cuprizone nor subjected to
depression induction.
Cuprizone model of demyelination
Demyelination was induced by continuously feeding
six-week-old C57BL/6 male mice a 0.2% cuprizone-enriched diet
purchased from Specialty Feeds (Western Australia, Australia).
After a six-week period of cuprizone diet, all groups were tested.
As the drug is temperature sensitive, the food containing the
cuprizone was kept at 4°C and was replenished every other day. The
presence of demyelination in the hippocampus of cuprizone-treated
mice was evaluated using Luxol Fast Blue (LFB) staining of
formalin-fixed sections as previously described (23,24).
EE
Three-week-old mice were housed in an EE, in groups
of 5–7 per cage, achieving social stimulation. The EE consisted of
two-story cages (40×26×38 cm), colorful walls, and equipped with a
climbing slide and an exercise wheel. The EE method in this study
design was based on a previous report by our group (25).
Induced depression
There has not been enough evidence to prove the
development of depression (major behavioral defects) in the
cuprizone mouse model; therefore, a repeated open-space forced swim
test was used to induce chronic depression-like state in mice to
mimic the comorbid depression seen in MS patients. The forced swim
test is the most commonly used test for the study of
depressive-like behavior in rodents (26). It is also considered as a preclinical
test to the screening of antidepressant activity in rats and mice
(27–29). Mice were made to swim in lukewarm
water (32±2°C) in an inescapable plastic container (43×24×23 cm)
for 15 min daily for 4 consecutive days. The container was filled
to a depth of 13 cm water; mice were not able to touch the bottom
with their feet or tails. This procedure resembles an inescapable
stressor producing an alternation in neural activity and brain cell
proliferation that is characteristic of depression. The protocol
was initiated 6 days prior to the testing period, as the effect of
depression lasts for 3 weeks (30).
Tests
All tests were conducted, scored and analyzed
blindly. Morris water maze (MWM) and open field test (OFT) trials
were videotaped. Each group was scored by all experimenters to
avoid bias.
MWM
Spatial learning and memory were tested over a
period of 6 days by the MWM (31).
The main component of the MWM set up is a circular pool (140×60 cm,
diameter × height) filled to a depth of 30 cm water (28±1°C). A
submerged platform (transparent, round and 8 cm in diameter) was
placed in a fixed location, 1 cm below water level. Various visible
cues were added on the pool's internal walls aiding spatial
discrimination. Each mouse was placed in the pool facing one of 5
starting positions (north, south, east, west and southeast), and
allowed to swim until it located and climbed onto the submerged
platform. Any mouse that failed to locate the platform within 120
sec was placed on the platform by the experimenter. All mice were
allowed to stay on the platform for 30 sec. The mouse was tracked
via a video computer system (Noldus Information Technology,
Wageningen, Netherlands). Latency and distance travelled to reach
the platform, in addition to swimming velocity, were analyzed using
the ANY-maze software on 5 consecutive days. Probe trials were
conducted on day 6, where the platform was removed and time spent
in the target quadrant was measured.
Rotarod performance test
Rotarod performance test is used to assess motor
function, coordination and balance and consists of a rotating drum
with a grooved surface for gripping (32). Mice underwent three, 2-min
pre-testing trials at a speed of 4 m/min. Each mouse had three
trials, where one constant speed (4.5 m/min) was employed. Latency
to fall was recorded.
OFT
The exploration of a novel environment and general
locomotor activity were evaluated by the OFT. The open field is a
square arena (44×44 cm) surrounded by 32 cm high walls. The floor
of the arena was divided into 16 even squares. The test apparatus
was placed in a dark room and the central four squares were
illuminated. Each mouse was tested for three, 10-min trials.
Following each trial, the arena was cleaned with 70% ethanol.
Distance travelled and time spent in the central zone were
recorded.
Statistical analysis
All data were presented as the mean ± standard error
of mean. T-test was used to establish the validity of the results
when comparing groups for the rotarod performance test. In tests
with multiple trials, like the MWM and OFT, a two-way repeated
Analysis of Variance (ANOVA) with Tukey's post hoc test, was used
to calculate the P-value. Statistical analysis was performed using
Microsoft Excel (version 2007). P<0.05 was considered to
indicate a statistically significant difference.
Results
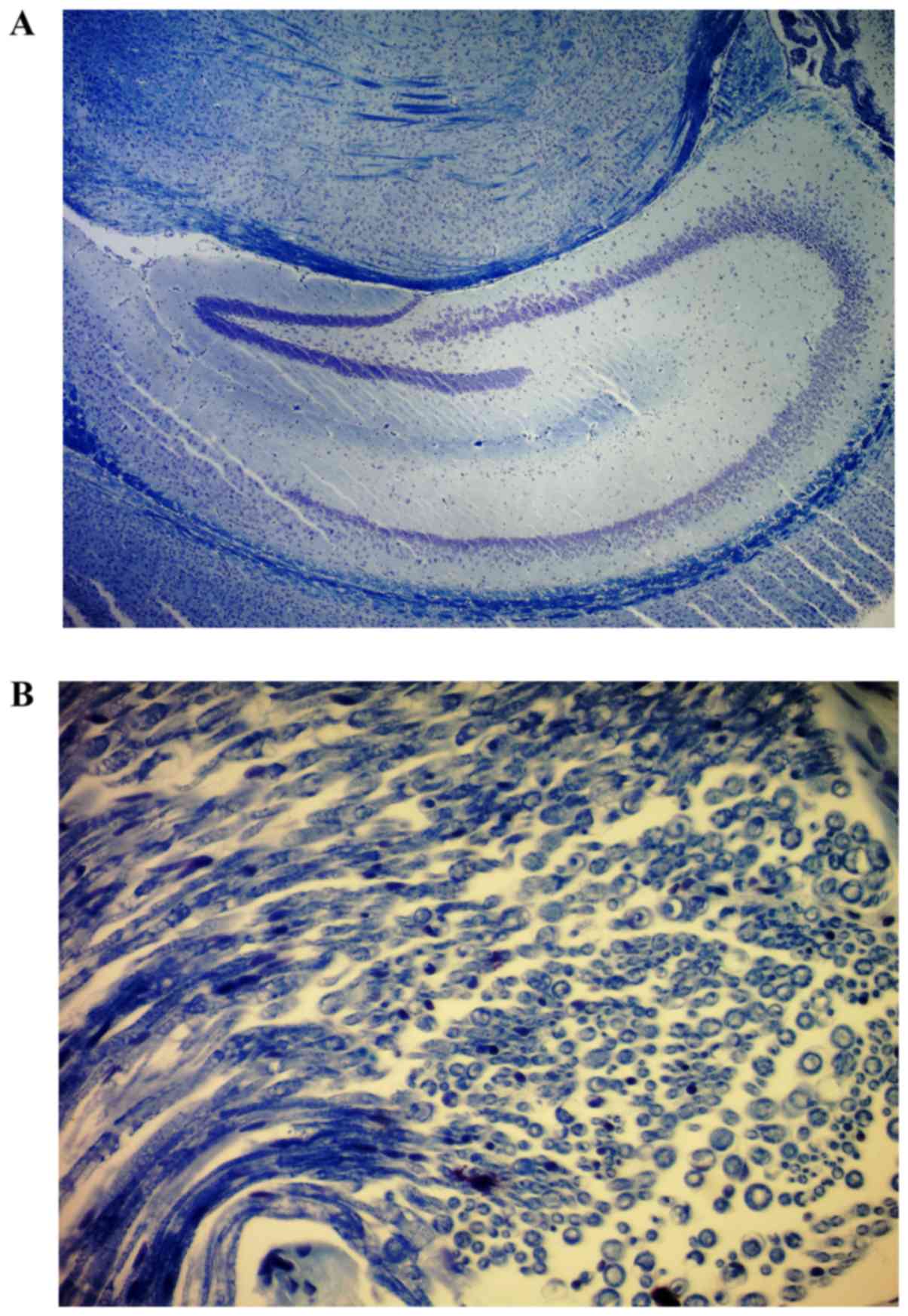
Cuprizone-induced demyelination
Demyelination was induced by continuously feeding
six-week-old C57BL/6 male mice a 0.2% cuprizone-enriched diet. The
presence of demyelination was observed in the hippocampus of
treated mice based on LFB staining (Fig.
1A and B).
Body weight
The weight of the animals was measured in the
beginning of the experiment and 1 week later, to determine the
cuprizone effect. The results of the effect of cuprizone
administration on the body weight of animals are displayed in
Table I. Cuprizone-fed animals
exhibited a significant reduction in body weight, when comparing
their initial body weight (Cup, 22.2±0.32; Cup-Dep, 22.1±0.2;
Cup-EE, 22±0.24; Cup-ED, 22.3±0.17) to the weight following a week
of cuprizone treatment (Cup 20.9±0.26, ANOVA, F=11.320 and
P=0.0039; Cup-Dep 21.1±0.2, ANOVA, F=12.462 and P=0.0023; Cup-EE
21.1±0.31, ANOVA, F=5.224 and P=0.0362; Cup-ED 21.3±0.17, ANOVA,
F=18 and P=0.00062). The control animals exhibited a significant
increase in weight (from 22.3±0.23 vs. 23.03±0.21; ANOVA, F=5.8255
and P=0.02815).
 | Table I.The effect of cuprizone
administration on the body weight of animals. |
Table I.
The effect of cuprizone
administration on the body weight of animals.
| Group | Weight (gm) at the
start of experiment | Weight (gm) after
one week of cuprizone | P-value |
|---|
| Control (no
cuprizone injection) | 22.3±0.23 | 23.03±0.21 | >0.05 |
| Cup | 22.2±0.32 | 20.9±0.26 | 0.0039 |
| Cup-Dep | 22.1±0.2 | 21.1±0.2 | 0.0027 |
| Cup-EE | 22±0.24 | 21.1±0.31 | 0.0362 |
| Cup-ED | 22.3±0.17 | 21.3±0.17 | 0.0006 |
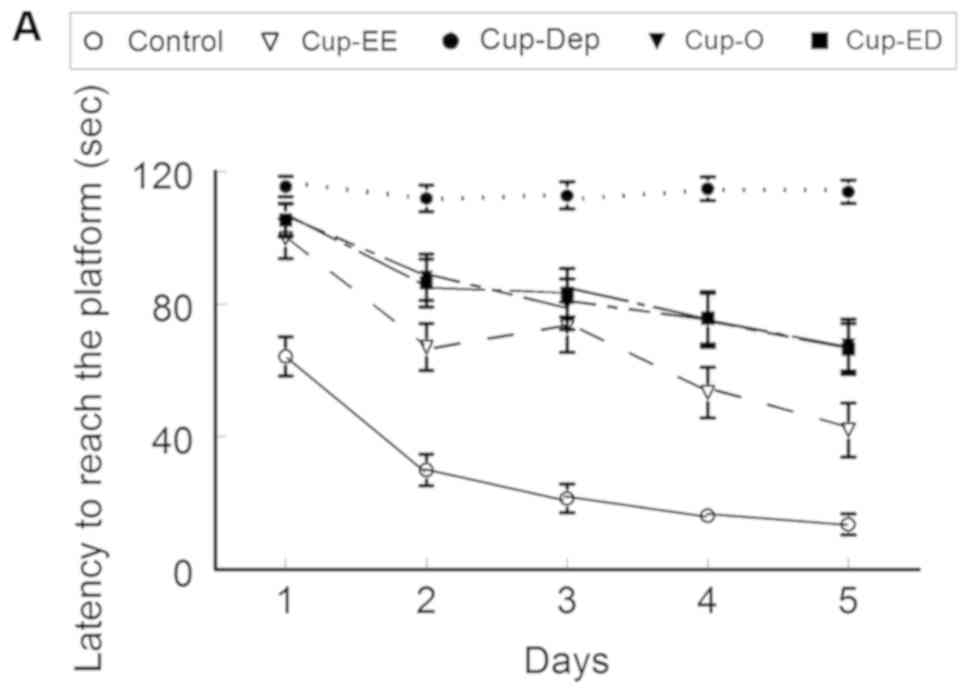
MWM
Latency and distance travelled to
reach the platform
Upon measuring the latency to reach the platform on
day 5, significant results were obtained. All cuprizone groups had
worse results than the control group. Cup-EE did significantly
better than Cup-O (42.0±8.2 vs. 67.7±7.8 sec, respectively; ANOVA,
F=43.788 and P<0.0001). Cup-Dep did significantly worse than
Cup-O (113.9±3.5 vs. 67.7±7.8 sec; ANOVA, F=29.517 and P<0.001)
and Cup-ED (66.5±7 sec; ANOVA, F=31.916 and P<0.0001), with the
latter two groups having comparable results (ANOVA, F=0.0121 and
P=0.9128; Fig. 2A). Similar results
were obtained with regards to distance travelled to reach the
platform on day 5 (Fig. 2B).
Swimming velocity
All cuprizone groups exhibited a slower swimming
velocity than the control group on days 1–4; which was also
reflected on the latency results. The swimming velocity of
cuprizone groups varied during the testing period with no
identifiable pattern (Fig. 2C).
Similar swimming velocities indicate that the results were not
influenced by differences in muscle power among the mice of the
cuprizone groups. No significant differences in swimming velocity
were observed between the groups (P>0.05).
Percentage of time spent in the target
quadrant
Apart from the control group, which spent 42.9±3% of
the time in the target quadrant, none of the other groups exhibited
a learning behavior, since they spent <25% of the time in the
target quadrant. However, a variation was noted among cuprizone
groups. Cup-EE (19.2±2.3%) had significantly better results than
Cup-Dep (11.1±2.5%; ANOVA, F=5.8964 and P=0.01721) and Cup-O
(12.3±3.1; ANOVA, F=5.1110 and P=0.026), whereas no significant
difference was observed between Cup-O and Cup-ED (13.3±2.9; ANOVA,
F=0.0596, and P=0.8076; Fig.
2D).
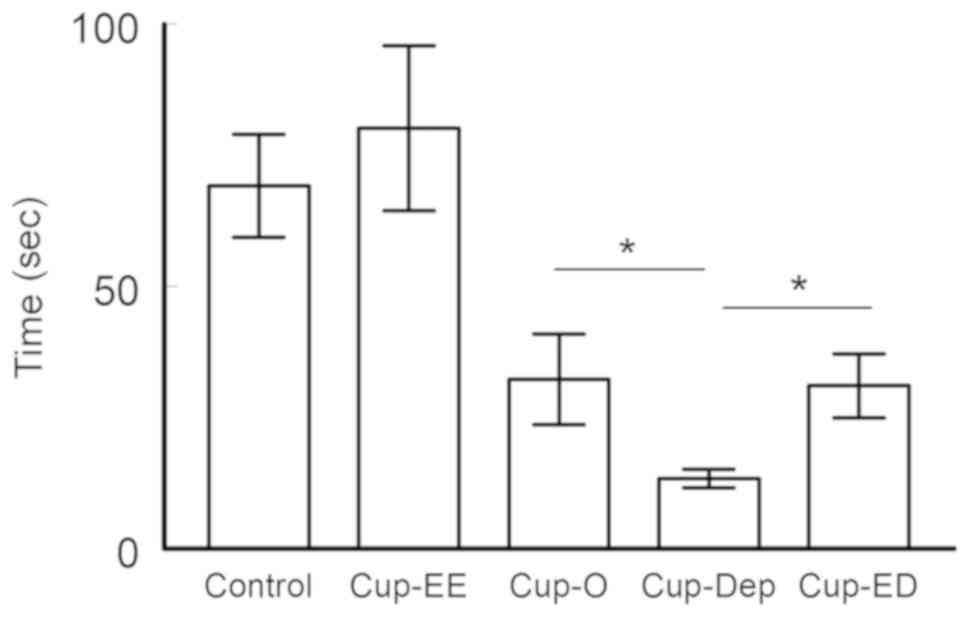
Rotarod performance test
Latency to fall
At the speed of 4.5 m/min, no significant difference
was observed between the control (70.1±10.6 sec) and Cup-EE
(80.1±15.7 sec; ANOVA, F=0.2799 and P=0.599). A variation was noted
among cuprizone groups; Cup-EE mice spent significantly more time
on the rotating rod than Cup-O mice (80.1±15.7 vs. 32.2±8.7,
respectively; ANOVA, F=7.1026 and P=0.0102). The results of the
Cup-Dep group were significantly worse than those of Cup-O
(13.3±1.8 vs. 32.2±8.7; ANOVA, F=4.594 and P=0.0368), indicating
declined motor functioning. No significant difference was observed
between the Cup-ED and Cup-O groups (31.0±6.1 vs. 32.2±8.7; ANOVA,
F=0.0133 and P=0.909; Fig. 3).
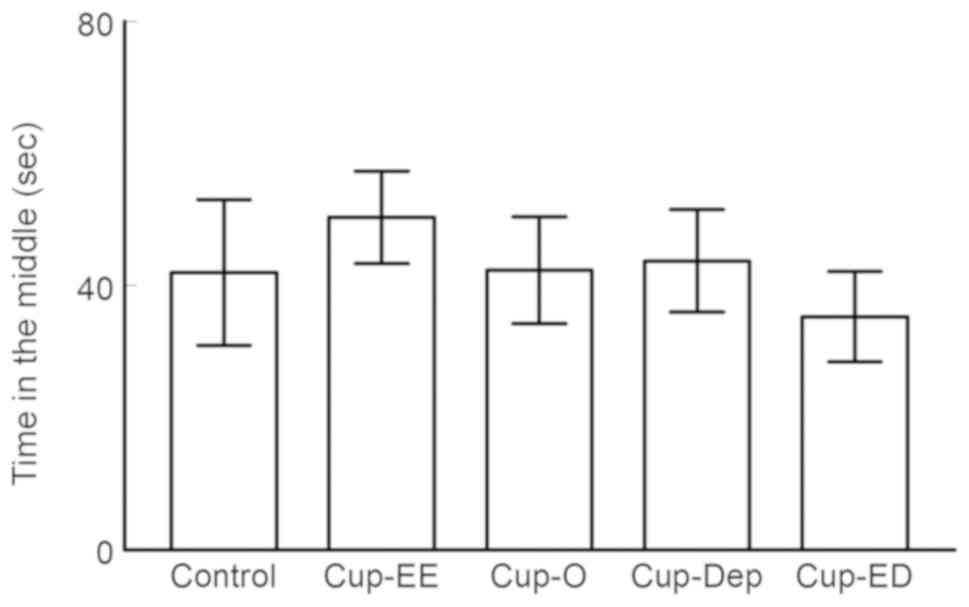
Open field
Time spent in the central zone. OFT results showed
no anxiety-like behavior in any of the groups, as they spent a
comparable amount of time in the central zone. No significant
difference was observed between the control and cuprizone groups
(P>0.05; Fig. 4).
Discussion
MS is a neurological chronic inflammatory disorder
of the CNS characterized by multifocal demyelination along with
astrocyte gliosis (1). Impaired
cognition and motor activity are commonly encountered in MS
patients (3) and depression is
believed to be a contributing factor (6–9).
Cuprizone-induced demyelination is considered a well-established
model of MS in animals (15,16), which leads to oligodendrocyte
apoptosis and steady demyelination and histopathological
alterations in the CNS (17,18). Cuprizone-induced toxicity has been
widely used to identify treatments for demyelinating diseases
(20–22).
The current study investigated the impact of induced
depression on a cuprizone mouse model of demyelination and the
effectiveness of EE as a method of intervention. C57BL/6 male mice
were divided into five groups: Cup-O, Cup-Dep, Cup-EE, Cup-ED and
the control group. In the present study, we first confirmed the
presence of demyelination in our cuprizone-treated mice using LFB
staining, a commonly used method to observe myelin under the light
to detect demyelination in the CNS (23,24).
Following a week of cuprizone treatment, our results showed a
significant reduction in the body weight of the animals, indicating
the effectiveness of cuprizone on these animals (33). The impact of induced depression was
studied in terms of cognitive and motor function. We tested the
spatial learning and memory in MWM as well as motor learning and
coordination by rotarod test. In the cuprizone model, induced
depression further worsened cognition, when compared to the
non-intervention group, as demonstrated by the results of the MWM.
Of note, further experiments can be conducted on transgenic and
knockout mice to ensure that abnormal responses, particularly in
learning and memory tasks, are indeed due to cognitive motor
impairment. Moreover, experiments using nutritional interventions
such as anti-oxidants-rich food in the diet of animals could be
conducted to slow the progression of cognitive decline.
Depression has been studied as an independent factor
influencing cognition and general outcome in MS patients. A study
conducted on MS patients with various degrees of depression
revealed greater neuropsychological dysfunction, as compared to
non-depressed MS patients (34).
Moreover, psychotherapy has been suggested as a method of
decreasing cognitive impairments in MS patients (35).
As established by the results of the rotarod
performance test in the present study, induced-depression appeared
to cause a significant decline in the motor performance of animals,
when compared to those undergoing no intervention.
Animals exposed to an EE exhibited enhanced
stimulation in their sensory, motor and cognitive systems, when
compared to those under normal housing conditions (36). The components of EE have been proven
to enhance neurogenesis, which in turn reflects on motor function
and cognition (12).
While in the present study EE did not rescue the
cognitive deficits in the cuprizone mouse model, enriched mice
performed significantly better, as compared to other cuprizone
groups. Altogether, the results of the MWM in this study provided
evidence that EE improved spatial learning and memory in the MS
animal model. Similarly, EE significantly improved motor function
in the cuprizone mouse model, as evidenced by the ability of the
mice to spend more time on the rotating rod. Notably, the effect of
EE was more pronounced in motor activity, suggesting that spatial
learning and memory are more complex processes.
The present findings revealed a marked effect of EE
on mice in the cuprizone group undergoing induced depression. A
complete reversal of the effects of induced depression was noted in
terms of cognition and motor function. Mice in the cuprizone group
undergoing induced depression and exposed to EE performed similarly
to those from the cuprizone group with no intervention. The
aforementioned results suggested that, although EE exposure did not
regain control values in the cuprizone mouse model of
demyelination, it certainly reversed the effect of induced
depression. These findings indicated that the disease effects of MS
on its own could not be reversed by EE, but the additive effect of
induced depression could. Nevertheless, the underlying biological
mechanism of EE in the cuprizone-induced demyelination mouse model
needs to be further investigated. For example, genetic studies
including the expression of genes and receptors that influence
brain plasticity and function may be important to elucidate the
effect of changes in gene expression on EE in cuprizone-induced
demyelination in animal models.
In the present study, the performance of the Cup-Dep
group of animals in the MWM with a hidden platform was
significantly worse than that of all other groups. This deficit was
unlikely, due to a learned helplessness that may have been induced
by the forced swim test, since the Cup-ED group completely reversed
the effects of depression in the MWM. In addition, the performance
of the Cup-ED group in a maze with a visible platform (data not
shown) was indistinguishable from that of other groups. Although
all cuprizone groups in the present study exhibited no learning
behavior, induced depression further deteriorated performance,
which was ameliorated by EE.
Considerable evidence has suggested that MS patients
have higher rates of anxiety (37,38). In
the present study, the results of the OFT failed to show an
anxiety-like behavior in the cuprizone mouse model, since all
groups exhibited similar exploratory behavior. Similar results,
suggesting that anxiety-like behavioral changes are subtle or
absent in the animal model of MS, were reported by Rodrigues et
al (39).
EE and depression are known to induce molecular and
cellular changes in specific brain regions (40,41).
These changes included altered gene expression profiles and
enhanced neurogenesis and synaptic plasticity (40).
Long term potentiation (LTP) and long-term
depression (LTD), two opposing forms of long-term plasticity, are
believed to be the actual synaptic processes underlying learning
and memory (42). EE modulates
synaptic plasticity by enhancing LTP, thus improving memory
(25). Conversely, depression and
stressful conditions impair LTP and facilitate LTD (41).
These observations correlate possible synaptic
plasticity changes in the hippocampus with exposure to different
factors. The present study suggests synaptic plasticity involvement
in the cuprizone mouse model of demyelination, where memory was
modulated upon exposure to EE and depression.
Increasing evidence suggests that the decline in
daily activities and functions in MS patients is the result of
interplay, among several other factors including depression which,
however, is not solely caused by the disease. These findings
clearly demonstrated the deteriorating effects of depression on the
cuprizone mouse model, and the ameliorating effect of EE on the
decline in cognition and motor function. Data from the present
study are very encouraging in the context of using EE to improve
the quality of life of patients with MS.
A limitation of the present study was the lack of
morphological and biochemical markers to prove demyelination.
However, cuprizone-induced demyelination is a widely used model of
MS in animals (15,16).
In conclusion, the results of this study raise the
possibility of future applications of non-medical therapy for MS,
and encourage future researchers to explore the technological
advances of non-invasive manipulations of brain activity to
modulate the course of various neurological diseases.
Acknowledgements
The authors would like to thank the technical staff
of the animal house at the College of Medicine and Medical
Sciences, Arabian Gulf University.
Funding
The present study was supported by departmental fund
from the Department of Physiology, College of Medicine and Medical
Sciences, Arabian Gulf University (grant no. 20-16).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
AM, GA, AK developed the project and edited the
manuscript; AM, AAlma, ZA, HA, RA, FA, AAlmu, AAl-M, AK collected
the data; all authors performed data analysis, managed the data and
wrote the manuscript. The final version of the manuscript has been
read and approved by all authors.
Ethical approval and consent to
participate
The study received ethical approval from the
Research and Ethics Committee of the College of Medicine and
Medical Sciences (Arabian Gulf University, Manama, Kingdom of
Bahrain).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Beer S, Khan F and Kesselring J:
Rehabilitation interventions in multiple sclerosis: An overview. J
Neurol. 259:1994–2008. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Langdon DW, Amato MP, Boringa J, Brochet
B, Foley F, Fredrikson S, Hämäläinen P, Hartung HP, Krupp L, Penner
IK, et al: Recommendations for a Brief International Cognitive
Assessment for Multiple Sclerosis (BICAMS). Mult Scler. 18:891–898.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Amato MP, Zipoli V and Portaccio E:
Multiple sclerosis-related cognitive changes: A review of
cross-sectional and longitudinal studies. J Neurol Sci. 245:41–46.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Cioncoloni D, Innocenti I, Bartalini S,
Santamecchi E, Rossi S, Rossi A and Ulivelli M: Individual factors
enhance poor health-related quality of life outcome in multiple
sclerosis patients. Significance of predictive determinants. J
Neurol Sci. 345:213–219. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Salter A, Thomas N, Tyry T, Cutter G and
Marrie RA: Employment and absenteeism in working-age persons with
multiple sclerosis. J Med Econ. 20:493–502. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Raimo S, Trojano L, Pappacena S, Alaia R,
Spitaleri D, Grossi D and Santangelo G: Neuropsychological
correlates of theory of mind deficits in patients with multiple
sclerosis. Neuropsychology. 31:811–821. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Gay MC, Bungener C, Thomas S, Vrignaud P,
Thomas PW, Baker R, Montel S, Heinzlef O, Papeix C, Assouad R and
Montreuil M: Anxiety, emotional processing and depression in people
with multiple sclerosis. BMC Neurol. 17:432017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Rahn K, Slusher B and Kaplin A: Cognitive
impairment in multiple sclerosis: A forgotten disability
remembered. Cerebrum. 2012:142012.PubMed/NCBI
|
|
9
|
Siegert RJ and Abernethy DA: Depression in
multiple sclerosis: A review. J Neurol Neurosurg Psychiatry.
76:469–475. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kazlauckas V, Pagnussat N, Mioranzza S,
Kalinine E, Nunes F, Pettenuzzo L, Souza DO, Portela LV,
Porciúncula LO and Lara DR: Enriched environment effects on
behavior, memory and BDNF in low and high exploratory mice. Physiol
Behav. 102:475–480. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhao YY, Shi XY, Qiu X, Lu W, Yang S, Li
C, Chen L, Zhang L, Cheng GH and Tang Y: Enriched environment
increases the myelinated nerve fibers of aged rat corpus callosum.
Anat Rec (Hoboken). 295:999–1005. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Cao W, Duan J, Wang X, Zhong X, Hu Z,
Huang F, Wang H, Zhang J, Li F, Zhang J, et al: Early enriched
environment induces an increased conversion of proBDNF to BDNF in
the adult rat's hippocampus. Behav Brain Res. 265:76–83. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wood NI, Carta V, Milde S, Skillings EA,
McAllister CJ, Ang YL, Duguid A, Wijesuriya N, Afzal SM, Fernandes
JX, et al: Responses to environmental enrichment differ with sex
and genotype in a transgenic mouse model of huntington's disease.
PLoS One. 5:e90772010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Magalon K, Cantarella C, Monti G, Cayre M
and Durbec P: Enriched environment promotes adult neural progenitor
cell mobilization in mouse demyelination models. Eur J Neurosci.
25:761–771. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kipp M, Clarner T, Dang J, Copray S and
Beyer C: The cuprizone animal model: New insights into an old
story. Acta Neuropathol. 118:723–736. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Torkildsen O, Brunborg LA, Myhr KM and Bø
L: The cuprizone model for demyelination. Acta Neurol Scand Suppl.
188:72–76. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kipp M, Nyamoya S, Hochstrasser T and Amor
S: Multiple sclerosis animal models: A clinical and
histopathological perspective. Brain Pathol. 27:123–137. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Komoly S: Experimental demyelination
caused by primary oligodendrocyte dystrophy. Regional distribution
of the lesions in the nervous system of mice [corrected]. Ideggyogy
Sz. 58:40–43. 2005.PubMed/NCBI
|
|
19
|
Stidworthy MF, Genoud S, Suter U, Mantei N
and Franklin RJ: Quantifying the early stages of remyelination
following cuprizone-induced demyelination. Brain Pathol.
13:329–339. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Matsushima GK and Morell P: The
neurotoxicant, cuprizone, as a model to study demyelination and
remyelination in the central nervous system. Brain Pathol.
11:107–116. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Morell P, Barrett CV, Mason JL, Toews AD,
Hostettler JD, Knapp GW and Matsushima GK: Gene expression in brain
during cuprizone-induced demyelination and remyelination. Mol Cell
Neurosci. 12:220–227. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Skripuletz T, Gudi V, Hackstette D and
Stangel M: De-and remyelination in the CNS white and grey matter
induced by cuprizone: The old, the new, and the unexpected. Histol
Histopathol. 26:1585–1597. 2011.PubMed/NCBI
|
|
23
|
Dumitrascu OM, Mott KR and Ghiasi H: A
comparative study of experimental mouse models of central nervous
system demyelination. Gene Ther. 21:599–608. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ingram G, Loveless S, Howell OW, Hakobyan
S, Dancey B, Harris CL, Robertson NP, Neal JW and Morgan BP:
Complement activation in multiple sclerosis plaques: An
immunohistochemical analysis. Acta Neuropathol Commun. 2:532014.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Artola A, von Frijtag JC, Fermont PC,
Gispen WH, Schrama LH, Kamal A and Spruijt BM: Long-lasting
modulation of the induction of LTD and LTP in rat hippocampal CA1
by behavioural stress and environmental enrichment. Eur J Neurosci.
23:261–272. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yankelevitch-Yahav R, Franko M, Huly A and
Doron R: The forced swim test as a model of depressive-like
behavior. J Vis Exp. 2–Mar;2015.doi: 10.3791/52587. View Article : Google Scholar
|
|
27
|
Castagné V, Moser P and Porsolt RD:
Behavioral assessment of antidepressant activity in rodents.
Methods of behavior analysis in neuroscience. 2nd. Boca Raton (FL):
CRCPress/Taylor & Francis; 2009
|
|
28
|
Porsolt RD, Le Pichon M and Jalfre M:
Depression: A new animal model sensitive to antidepressant
treatments. Nature. 266:730–732. 1977. View
Article : Google Scholar : PubMed/NCBI
|
|
29
|
Porsolt RD, Bertin A and Jalfre M:
‘Behavioural despair’ in rats and mice: Strain differences and the
effects of imipramine. Eur J Pharmacol. 51:291–294. 1978.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Stone EA, Lin Y and Quartermain D:
Evaluation of the repeated open-space swim model of depression in
the mouse. Pharmacol Biochem Behav. 91:190–195. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
D'Hooge R and De Deyn PP: Applications of
the Morris water maze in the study of learning and memory. Brain
Res Brain Res Rev. 36:60–90. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Bohlen M, Cameron A, Metten P, Crabbe JC
and Wahlsten D: Calibration of rotational acceleration for the
rotarod test of rodent motor coordination. J Neurosci Methods.
178:10–14. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Sachs HH, Bercury KK, Popescu DC,
Narayanan SP and Macklin WB: A new model of cuprizone-mediated
demyelination/remyelination. ASN Neuro. 6(pii):
17590914145519552014.PubMed/NCBI
|
|
34
|
Boeschoten RE, Braamse AMJ, Beekman ATF,
Cuijpers P, van Oppen P, Dekker J and Uitdehaag BMJ: Prevalence of
depression and anxiety in Multiple Sclerosis: A systematic review
and meta-analysis. J Neurol Sci. 372:331–341. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Bilgi E, Özdemir HH, Bingol A and Bulut S:
Evaluation of the effects of group psychotherapy on cognitive
function in patients with multiple sclerosis with cognitive
dysfunction and depression. Arq Neuropsiquiatr. 73:90–95. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Alwis DS and Rajan R: Environmental
enrichment and the sensory brain: The role of enrichment in
remediating brain injury. Front Syst Neurosci. 8:1562014.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Serra-de-Oliveira N, Boilesen SN, Prado de
França Carvalho C, LsSueur-Maluf L, Zollner Rde L, Spadari RC,
Medalha CC and Monteiro de Castro G: Behavioural changes observed
in demyelination model shares similarities with white matter
abnormalities in humans. Behav Brain Res. 287:265–275. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Acharjee S, Nayani N, Tsutsui M, Hill MN,
Ousman SS and Pitman QJ: Altered cognitive-emotional behavior in
early experimental autoimmune encephalitis-Cytokine and hormonal
correlates. Brain Behav Immun. 33:164–172. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Rodrigues DH, Vilela Mde C,
Lacerda-Queiroz N, Miranda AS, Sousa LF, Reis HJ and Teixeira AL:
Behavioral investigation of mice with experimental autoimmune
encephalomyelitis. Arq Neuropsiquiatr. 69:938–942. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Spires TL and Hannan AJ: Nature, nurture
and neurology: Gene-environment interactions in neurodegenerative
disease. FEBS Anniversary Prize Lecture delivered on 27 June 2004
at the 29th FEBS Congress in Warsaw. FEBS J. 272:2347–2361. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Von Frijtag JC, Kamal A, Reijmers LG,
Schrama LH, van den Bos R and Spruijt BM: Chronic imipramine
treatment partially reverses the long-term changes of hippocampal
synaptic plasticity in socially stressed rats. Neurosci Lett.
309:153–156. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Bliss TV and Cooke SF: Long-term
potentiation and long-term depression: A clinical perspective.
Clinics (Sao Paulo). 66 (Suppl 1):S3–S17. 2011. View Article : Google Scholar
|