Introduction
Thyroid cancer (TC) commonly presents in women of
all ages and is associated with a high morbidity, worldwide
(1,2). Despite the high survival rates of
affected patients, an increasing incidence combined with a high
rate of recurrence and a young age at diagnosis indicates that
additional research on TC is required (2,3).
Follicular thyroid carcinoma (FTC) accounts for almost 15% of all
cases of TC (4).
Long non-coding RNAs (lncRNAs) are a class of
non-protein-coding transcripts that are >200 nucleotides in
length (5). Increasing evidence
indicates that lncRNAs are crucial factors in tissue physiology and
the pathological processes of various diseases, including cancer
(6,7). A large number of dysregulated lncRNAs
have been identified in cancer, suggesting their oncogenic or
tumor-suppressive properties (8,9). lncRNAs
have been reported to serve an important role in the oncogenesis
and progression of TC (10,11). For example, the lncRNA PTC
susceptibility candidate 3, which is specifically expressed in
thyroid tissues and downregulated in PTC tissues, has been reported
to function as a tumor suppressor through the S100 calcium-binding
protein A4 pathway (12,13). In addition, the expression of
BRAF-activated lncRNA (BANCR) was found to be higher in PTC tissues
compared with that in matched normal tissues, and BANCR has been
reported to contribute to the genesis of PTC through regulating
cell proliferation and the cell cycle (14,15).
Recent studies have indicated that expression of
lncRNA H19 is upregulated in TC tissues and TC cell lines (4,16).
Furthermore, an increased expression of lncRNA H19 was reported to
be associated with poor prognosis of patients with TC (4). However, the underlying molecular
mechanisms remain to be fully elucidated. It has been reported that
the overexpression of H19 in TC cell lines, including TPC-1 and NIM
lines, promoted cell proliferation, migration and invasion, whereas
the knockdown of H19 had the opposite effect and enhanced cell
apoptosis (16). The exact molecular
mechanisms remain to be investigated and confirmed.
In the present study, the expression of H19 in TC
tissues was analyzed. Furthermore, the FTC-133 human TC cell line
was used to assess the role of H19 in TC in vitro and to
investigate the possible mechanisms.
Materials and methods
Tissue samples
The present study included 30 patients who underwent
thyroidectomy for TC between May 2016 and June 2017 at the Fourth
Hospital of Hebei Medical University (Shijiazhuang, China). The
characteristics of the patients are listed in Table I. Samples of cancer tissues and
adjacent normal tissues were collected, immediately frozen in
liquid nitrogen and stored at −80°C until use. The study protocol
was approved by the Ethics Committee of the Fourth Hospital of
Hebei Medical University and all patients provided written informed
consent prior to participation in the study.
 | Table I.Expression of H19 in tissues from
patients with thyroid cancer. |
Table I.
Expression of H19 in tissues from
patients with thyroid cancer.
| Factor | Cases | H19a | P-value |
|---|
| Gender |
|
| 0.623 |
| Male | 14 | 6.645±0.210 |
|
|
Female | 16 | 6.783±0.182 |
|
| Age (years) |
|
| 0.578 |
|
<60 | 17 | 6.661±0.239 |
|
|
≥60 | 13 | 6.848±0.215 |
|
| Histological
grade |
|
| 0.041 |
|
Well-intermediately
differentiated | 9 | 6.106±0.240 |
|
| Poorly
differentiated | 21 | 6.918±0.225 |
|
Cell culture
The human FTC-133 and TPC-1 TC cell lines were
purchased from the Shanghai Institute of Cell Biology of the
Chinese Academy of Sciences (Shanghai, China) and cultured in
RPMI-1640 medium (Invitrogen; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA) supplemented with 10% fetal bovine serum (Thermo
Fisher Scientific, Inc.), 100 IU/ml penicillin and 100 mg/ml
streptomycin (Invitrogen; Thermo Fisher Scientific, Inc.), and
incubated at 37°C in a humidified atmosphere containing 5%
CO2. The cells were passaged every other day and cells
in the logarithmic growth phase were used for the subsequent
experiments.
Cell transfection
For the knockdown of lncRNA H19, two complementary
oligonucleotides of small interfering (si)RNA, H19-siRNA1 and
H19-siRNA2, and non-targeting control siRNA (NC) were designed. The
sequences were chemically synthesized by Shanghai GeneChem Co.,
Ltd. (Shanghai, China). Using Lipofectamine 2000™ reagent
(Invitrogen; Thermo Fisher Scientific, Inc.) following the
manufacturer's protocol, the TC cells were transiently transfected
with H19 siRNA (siRNA-1 or siRNA-2) or NC control. The mRNA levels
were detected at 48 h post-transfection using a
microspectrophotometer at a wavelength of 260 nm. The most
efficient siRNA was selected for the subsequent experiments.
Cell viability assay
The cells were seeded into a 96-well culture plate
at a density of 5×104 cells/well. The cell viability was assessed
using an MTT assay (Bio-Rad Laboratories, Inc., Hercules, CA, USA).
Following the manufacturer's protocols, 10 µl of MTT reagent (5
mg/ml) was added to each well, followed by incubation for 4 h at
37°C. The supernatants were aspirated and the formazan crystals
were dissolved in dimethylsulfoxide. The optical density of each
well was measured using an ELISA reader (Bio-Rad Laboratories,
Inc.) at a wavelength of 490 nm at 12, 24 or 48 h.
RNA isolation and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
analysis
Total RNA from the thyroid tissue and cells was
extracted using TRIzol reagent (Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturer's protocol. Total
RNA was reverse-transcribed into complementary DNA with a
PrimeScript™ RT reagent kit (Takara Biotechnology Co., Ltd.,
Dalian, China) and qPCR was performed using a SYBR®
Premix Ex Taq™ kit (Takara Biotechnology Co., Ltd.) on an ABI 7300
Real-Time PCR system (Applied Biosystems; Thermo Fisher Scientific,
Inc.) according to the manufacturers' protocols. The reactions were
under the following conditions: 95°C for 7 min; 40 cycles of 95°C
for 15 sec, 60°C for 30 sec, and extension at 72°C for 30 sec. The
relative expression of mRNA in each sample was normalized to GAPDH
and calculated using the 2−ΔΔCq method (17). The primer sequences are listed in
Table II.
| Table II.Oligonucleotide primers used for
quantitative polymerase chain reaction analysis. |
Table II.
Oligonucleotide primers used for
quantitative polymerase chain reaction analysis.
| Name | Primer
sequence |
|---|
| H19 | Forward
5′-ACTCAGGAATCGGCTCTGGAA-3′ |
|
| Reverse
5′-CTGCTGTTCCGATGGTGTCTT-3′ |
| U6 | Forward
5′-CTCGCTTCGGCAGCACA-3′ |
|
| Reverse
5′-AACGCTTCACGAATTTGCGT-3′ |
| Bax | Forward
5′-CACCAGCTCTGAACAGATCATGA-3′ |
|
| Reverse
5′-TCAGCCCATCTTCTTCCAGATGT-3′ |
| Bcl-2 | Forward
5′-CACCCCTGGCATCTTCTCCTT-3′ |
|
| Reverse
5′-AGCGTCTTCAGAGACAGCCAG-3′ |
| Caspase-3 | Forward
5′-CATGGAAGCGAATCAATGGACT-3′ |
|
| Reverse
5′-CTGTACCAGACCGAGATGTCA-3′ |
| GAPDH | Forward
5′-GGAGCGAGATCCCTCCAAAAT-3′ |
|
| Reverse
5′-GGCTGTTGTCATACTTCTCATGG-3′ |
Protein isolation and western blot
analysis
The proteins were extracted from cells using
ice-cold radioimmunoprecipitation assay lysis buffer (Beyotime
Institute of Biotechnology, Haimen, China). The concentration of
protein was determined using a BCA kit (Thermo Fisher Scientific,
Inc.). Equal quantities of protein (80 µg) were denatured in a
boiling water bath and then separated by 10% SDS-PAGE, followed by
transfer onto polyvinylidene difluoride membranes. Subsequently,
the membranes were blocked with 5% non-fat milk, and incubated with
primary antibodies specific for B-cell lymphoma 2 (Bcl-2; cat. no.
ab32124, 1:1,000, Abcam, Cambridge, MA, USA), Bcl-2-associated X
protein (Bax; cat. no. ab32503, 1:1,000, Abcam), caspase 3 (cat.
no. ab32351, 1:5,000, Abcam), phosphoinositide-3 kinase (p-PI3K;
cat. no. ab109006, 1:1,000, Abcam), AKT (cat. no. ab133458,
1:1,000, Abcam) or GAPDH (cat. no. ab181603, 1:10,000, Abcam) at
37°C for 1 h. Following washing with PBS five times, the membranes
were incubated with horseradish peroxidase-conjugated secondary
antibodies (cat. no. ab205718, 1:2,000, Abcam) at room temperature
for 60 min, and bands were visualized with a Novex® ECL
Chemiluminescent Substrate Reagent kit (Thermo Fisher Scientific,
Inc.). Images were captured using a ChemiDoc™
XRS+ imaging system (Bio-Rad Laboratories, Inc.). The
signal intensities were quantified using ImageJ software 1.46
(National Institutes of Health, Bethesda, MD, USA).
Apoptosis assay
The apoptotic cells were quantified using an Annexin
V-fluorescein isothiocyanate (FITC)/propidium iodide (PI) Apoptosis
Detection kit (BD Biosciences, Franklin Lakes, NJ, USA) following
the manufacturer's protocol. The cells were seeded in 6-well plates
and stained with Annexin V/FITC for 15 min at room temperature in
the dark, followed by the addition of PI for 5 min at room
temperature in the dark. Finally, the cells were subjected to flow
cytometric analysis (BD Biosciences) and analyzed using CellQuest
software 5.1 (BD Biosciences).
Statistical analysis
All data were analyzed using SPSS 20.0 software (IBM
Corp., Armonk, NY, USA). Values are expressed as the mean ±
standard deviation. Statistical significance was analyzed using
Student's t-test. The difference among multiple groups were
evaluated with one-way ANOVA followed by a Newman-Keuls post hoc
test. P<0.05 was considered to indicate a statistically
significant difference.
Results
Characteristics of patients
As shown in Table I,
the expression levels of H19 were significantly associated with
histological grade (P=0.041), whereas there was no significant
association between the expression of H19 and the gender (P=0.623)
or ages of the patients (0.578).
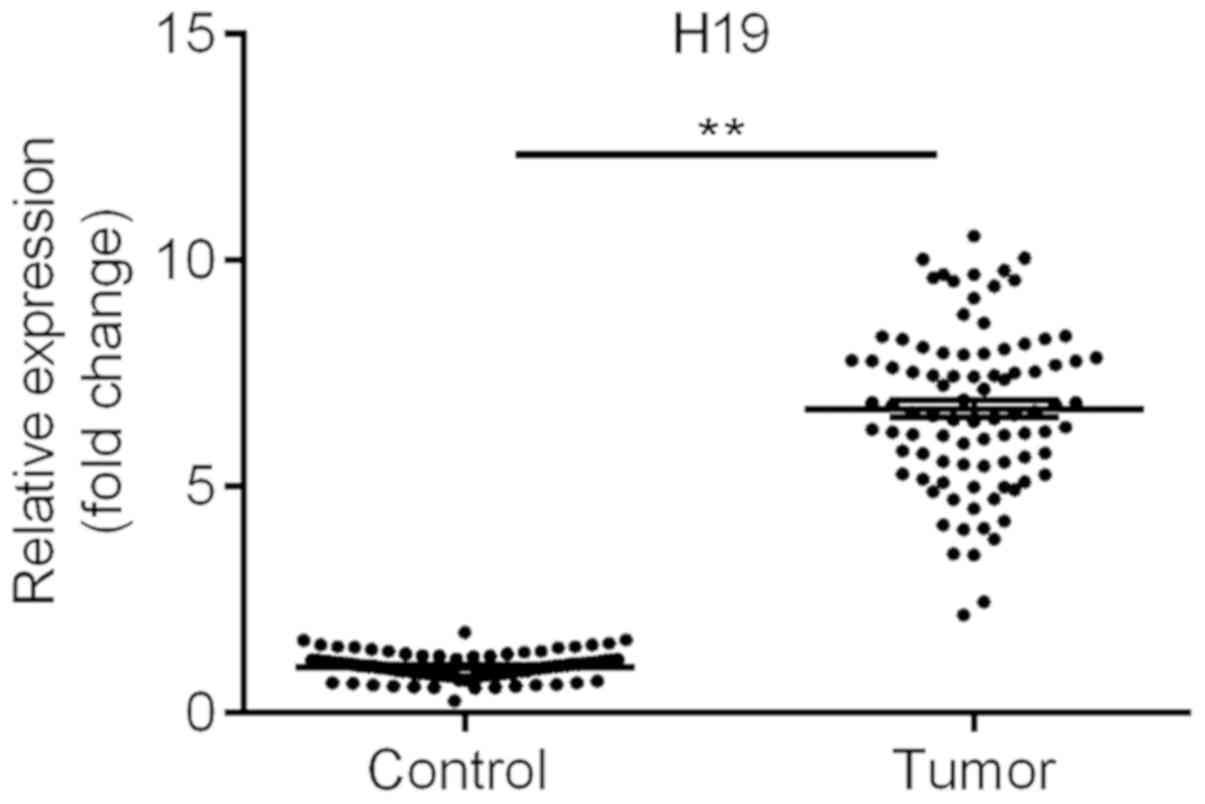
lncRNA H19 is upregulated in TC
tissues
RT-qPCR analysis was used to examine the expression
of H19. The results suggested that the levels of H19 in the TC
tissues were significantly upregulated, by over six-fold, compared
with those in the adjacent normal tissues (Fig. 1), which is consistent with the
results of recent studies (4,16). These
results suggested that H19 may be involved in the development of
TC.
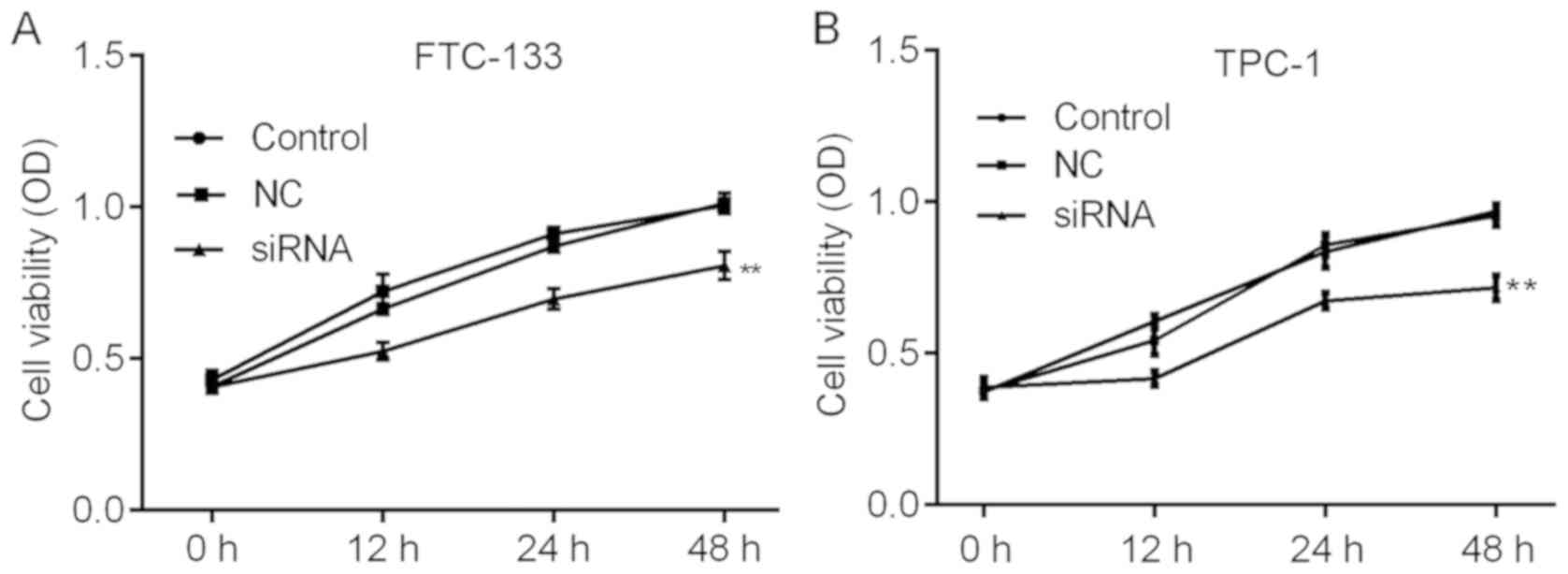
H19 knockdown inhibits TC cell
viability and induces apoptosis in vitro
The roles of H19 in TC were further investigated by
assessing the cell viability and apoptosis of TC cells subjected to
H19 knockdown. The knockdown efficiencies are shown in Fig. 2A and B. H19 siRNA-2 was selected for
subsequent experiments due to its relatively higher efficiency. As
indicated in Fig. 3A and B, H19
knockdown inhibited cell viability in a time-dependent manner, with
a significant reduction at 48 h post-transfection. To further
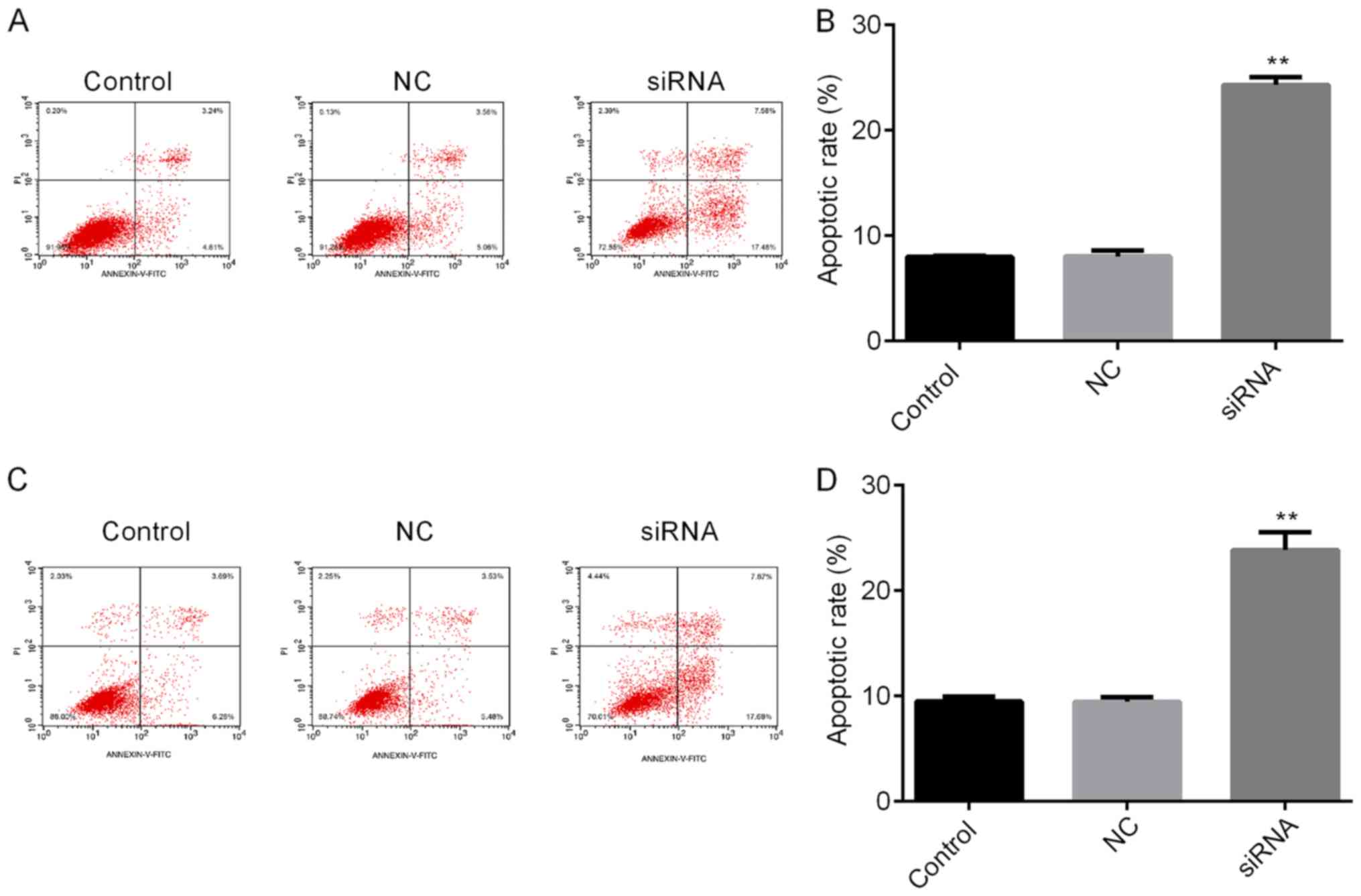
investigate the mechanisms by which H19 inhibits TC cell viability,
the apoptotic rates of TC cells with H19 knockdown were assessed by
flow cytometry. The results indicated that H19 silencing
significantly enhanced the apoptotic rate (Fig. 4A-D). Subsequently, whether H19
regulates the expression of the apoptosis-associated molecules Bax,
Bcl-2 and caspase 3 was assessed (18,19). As
presented in Figs. 5A-E and 6A-E, significant increases in Bax and
caspase 3, and repression of Bcl-2 were observed at the mRNA and
protein expression levels in TC cells following H19 silencing.
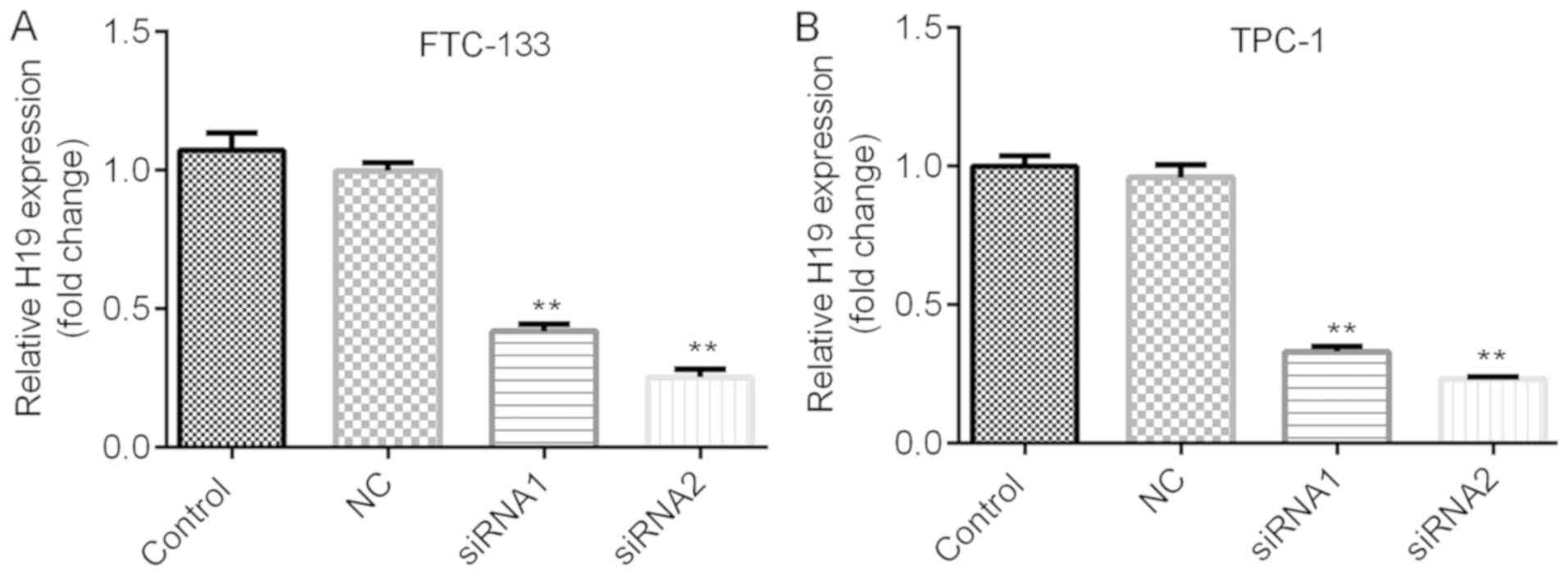 | Figure 2.Expression of H19 in thyroid cancer
cells. Reverse transcription-quantitative polymerase chain reaction
analysis was performed to examine the expression of H19. (A)
FTC-133 cells were transfected with NC siRNA, H19 siRNA1 or H19
siRNA2. Compared with that in the NC group, the expression of H19
was significantly decreased. (B) TPC-1 cells were transfected with
NC siRNA, H19 siRNA1 or H19 siRNA2. Compared with that in the NC
group, the expression of H19 was significantly decreased. Of note,
H19 siRNA2 was more potent at suppressing the expression of H19.
**P<0.01. Groups: NC, cells transfected with NC siRNA; siRNA1,
cells transfected with H19 siRNA1; siRNA2, cells transfected with
H19 siRNA2. NC, negative control; siRNA, small silencing RNA. |
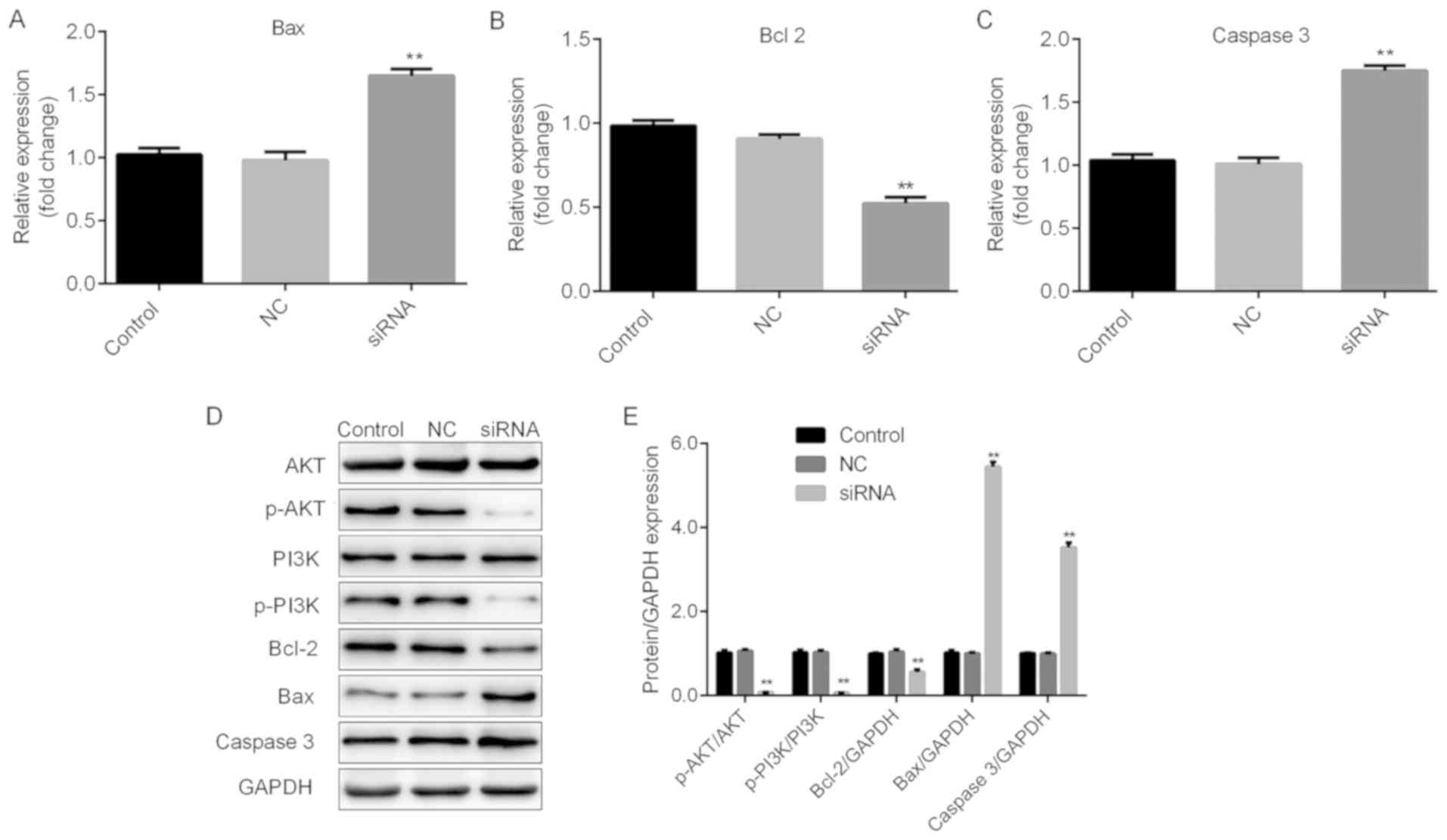 | Figure 5.H19 may function through the Bax,
Bcl2, caspase 3 and the PI3K/AKT pathway in FTC-133 cells. Thyroid
cancer cells were transfected with H19 siRNA, NC or control,
respectively. Reverse transcription-quantitative polymerase chain
reaction analysis of (A) Bax, (B) Bcl-2 and (C) caspase 3
expression was performed. Results are expressed relative to the
value of the NC group (set at 1 as the reference). (D)
Representative western blot images for determining the expression
of Bax, Bcl-2 and caspase 3, and the levels of p-PI3K and p-AKT
protein. GAPDH was used as the loading control. (E) Quantitative
analysis of relative changes in the proteins indicated above.
**P<0.01. Groups: NC, cells transfected with NC siRNA; siRNA,
cells transfected with H19 siRNA. NC, negative control; siRNA,
small silencing RNA; p-PI3K, phosphorylated phosphoinositide-3
kinase; Bax, Bcl-2-associated X protein; Bcl-2, B-cell lymphoma
2. |
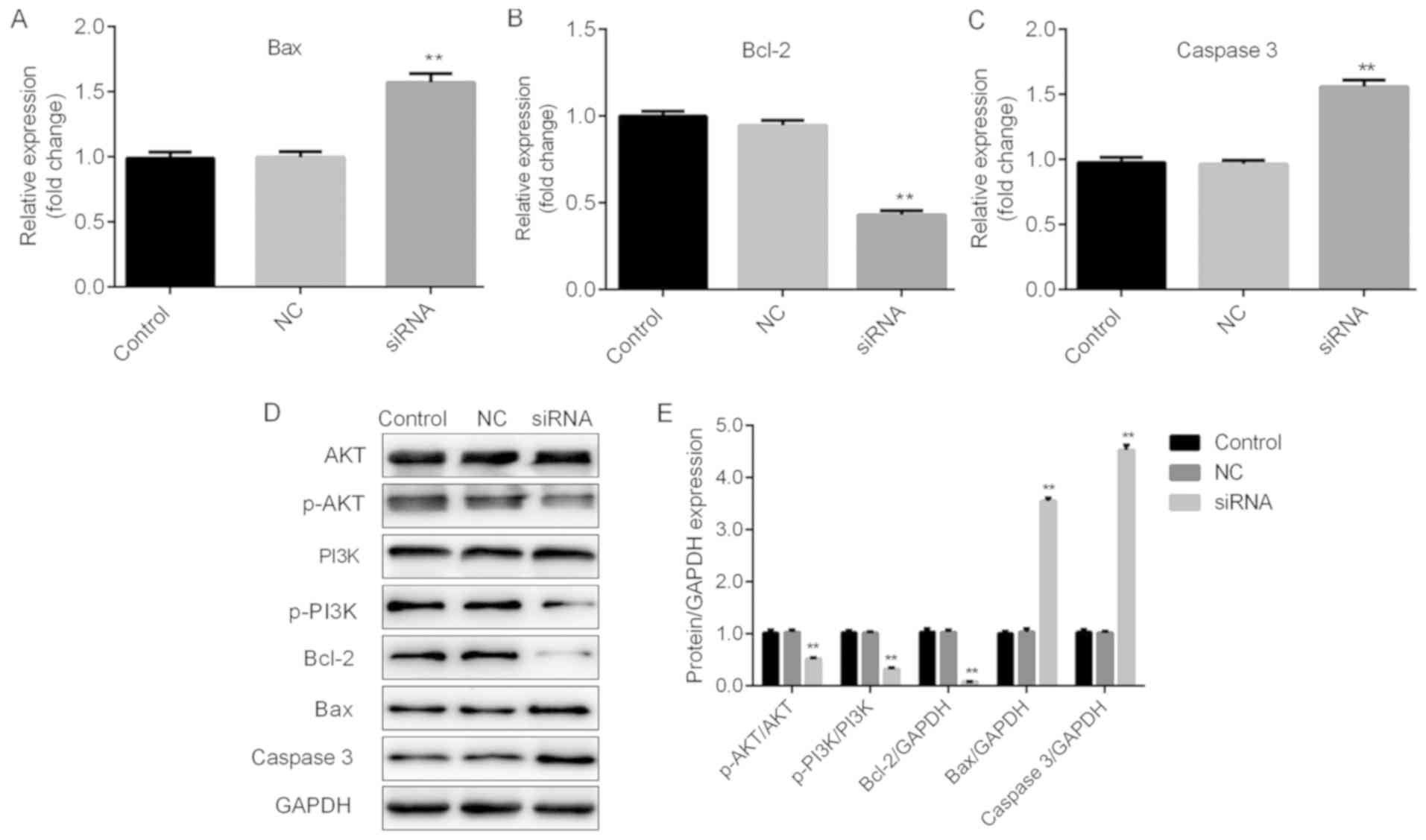 | Figure 6.H19 may function through Bax, Bcl2,
caspase 3 and the PI3K/AKT pathway in TPC-1 cells. Thyroid cancer
cells were transfected with H19 siRNA, NC or control, respectively.
Reverse transcription-quantitative polymerase chain reaction
analysis of (A) Bax, (B) Bcl-2 and (C) caspase 3 expression was
performed. Results are expressed relative to the value of the NC
group (set at 1 as the reference). (D) Representative western blot
images for determining the expression of Bax, Bcl-2 and caspase 3,
and the levels of p-PI3K and p-AKT protein. GAPDH was used as the
loading control. (E) Quantitative analysis of relative changes in
the proteins indicated above. **P<0.01. Groups: NC, cells
transfected with NC siRNA; siRNA, cells transfected with H19 siRNA.
NC, negative control; siRNA, small silencing RNA; p-PI3K,
phosphorylated phosphoinositide-3 kinase; Bax, Bcl-2-associated X
protein; Bcl-2, B-cell lymphoma 2. |
H19 may exert its functions in TC via
the PI3K/AKT pathway
The PI3K/AKT signaling pathway has been reported to
be associated with cell viability and apoptosis (20,21).
Therefore, whether PI3K/AKT signaling is involved in H19-mediated
cellular responses was examined. As presented in Figs. 5D and E and 6D and E, H19 silencing induced a
significant repression of p-PI3K and p-AKT, indicating that H19
exerts its cellular functions at least in partly through the
PI3K/AKT signaling pathway.
Discussion
Increasing efforts have contributed to elucidating
the molecular and cellular mechanisms underlying the progression of
cancer, among which lncRNAs have attracted increasing attention
(22). However, effective therapy
for cancer remains limited (23),
therefore, novel molecular signatures or potential molecular
targets are urgently required. lncRNA H19 has been reported to have
an oncogenic role in various types of cancer, including gastric
cancer and cholangiocarcinoma (24,25).
However, current knowledge of H19 in TC is limited. In the present
study, the expression of H19 was increased in TC tissues,
indicating that H19 may be an oncogene in TC, which was consistent
with previous studies (24,25). The main types of TC are papillary
thyroid cancer (PTC, ~85%) and follicular thyroid cancer (FTC,
~10%) (26). FTC is more aggressive
and harder to diagnose than PTC and the 10-year survival rate is
lower (27). Although the potential
role of H19 has been reported in TC (28), the underlying mechanisms have
remained unclear.
Previous studies have demonstrated that H19
suppresses the migration, invasion and cell viability of TC cells
(29,30). In the present study, it was found
that downregulated H19 inhibited the cell viability and promoted
apoptosis of FTC-133 and TPC-1 TC cells, which further verified H19
as an oncogene in TC. Liu et al reported that H19 regulated
the proliferation, migration and invasion of TC cells via
regulating the expression of miR-17-5p and YES1 (16). In addition, the overexpression of H19
has been shown to predict poor prognosis in patients with TC via
regulating let-7 (31). The present
study investigated whether H19 can regulate the viability and
apoptosis of TC cells via other pathways.
The PI3K/AKT signaling pathway has an important role
in tumorigenesis (32).
Additionally, H19 has been shown to exert an oncogenic function by
regulating the PI3K/AKT pathway in various types of cancer
(33,34). Bcl-2 and Bax are crucial in tumor
cell apoptosis (35). Feng et
al demonstrated that caspase 3 was associated the apoptosis of
TC cells (36). In the present
study, RT-qPCR and western blot analyses were performed to examine
the mRNA and protein levels of Bcl-2/Bax, p-PI3K/AKT and caspase 3.
The knockdown of H19 significantly suppressed Bcl-2, p-PI3K and
p-AKT, and upregulated Bax and caspase 3, indicating that the
suppression of H19 may have inhibited cell viability and promoted
the apoptosis of TC cells via regulating Bcl-2/Bax, p-PI3K/AKT and
caspase 3.
However, the present study had limitations. First,
the study was limited to by number of the patients enrolled.
Second, all the experiments performed in the study was in
vitro, thus in vivo experiments are required in further
investigations.
In conclusion, H19 was upregulated in TC tissues.
The effects of decreased H19 were broadly similar in suppressing
the proliferation and promoting the apoptosis of the FTC-133 FTC
cell line and TPC-1 PTC cell line, suggesting that H19 may be
applicable as a target across different types of TC.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
XL prepared the manuscript and performed experiments
with the assistance of QL. XJ collected the data and provided
interpretation with the assistance of HG. YL conceived and designed
the current study.
Ethics approval and consent to
participate
The study protocol was approved by the Ethics
Committee of the Fourth Hospital of Hebei Medical University and
all patients provided written informed consent prior to
participation in the study.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Klubo-Gwiezdzinska J, Yang L, Merkel R,
Patel D, Nilubol N, Merino MJ, Skarulis M, Sadowski SM and Kebebew
E: Results of screening in familial non-medullary thyroid cancer.
Thyroid. 27:1017–1024. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Aschebrook-Kilfoy B, James B, Nagar S,
Kaplan S, Seng V, Ahsan H, Angelos P, Kaplan EL, Guerrero MA, Kuo
JH, et al: Risk factors for decreased quality of life in thyroid
cancer survivors: Initial findings from the North American thyroid
cancer survivorship study. Thyroid. 25:1313–1321. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Grogan RH, Kaplan SP, Cao H, Weiss RE,
Degroot LJ, Simon CA, Embia OM, Angelos P, Kaplan EL and Schechter
RB: A study of recurrence and death from papillary thyroid cancer
with 27 years of median follow-up. Surgery. 154:1436–1447. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Gillanders SL and O'Neill JP: Prognostic
markers in well differentiated papillary and follicular thyroid
cancer (WDTC). Eur J Surg Oncol. 44:286–296. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Jeong S, Lee J, Kim D, Seol MY, Lee WK,
Jeong JJ, Nam KH, Jung SG, Shin DY, Lee EJ, et al: Relationship of
focally amplified long noncoding on chromosome 1 (FAL1) lncRNA with
E2F transcription factors in thyroid cancer. Medicine (Baltimore).
95:e25922016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Mercer TR, Dinger ME and Mattick JS: Long
non-coding RNAs: Insights into functions. Nat Rev Genet.
10:155–159. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
7
|
Hao S, Yao L, Huang J, He H, Yang F, Di Y,
Jin C and Fu D: Genome-wide analysis identified a number of
dysregulated long noncoding RNA (lncRNA) in human pancreatic ductal
adenocarcinoma. Technol Cancer Res Treat. 17:15330346177484292018.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ponzio G, Rezzonico R, Bourget I, Allan R,
Nottet N, Popa A, Magnone V, Rios G, Mari B and Barbry P: A new
long noncoding RNA (lncRNA) is induced in cutaneous squamous cell
carcinoma and down-regulates several anticancer and cell
differentiation genes in mouse. J Biol Chem. 292:12483–12495. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhao JH, Sun JX, Song YX, Chen XW, Yang
YC, Ma B, Wang J, Gao P and Wang ZN: A novel long noncoding
RNA-LOWEG is low expressed in gastric cancer and acts as a tumor
suppressor by inhibiting cell invasion. J Cancer Res Clin Oncol.
142:601–609. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Chen C, Zhou L, Wang H, Chen J, Li W, Liu
W, Shen M, Liu H and Fu X: Long noncoding RNA CNALPTC1 promotes
cell proliferation and migration of papillary thyroid cancer via
sponging miR-30 family. Am J Cancer Res. 8:192–206. 2018.PubMed/NCBI
|
|
11
|
Li T, Yang XD, Ye CX, Shen ZL, Yang Y,
Wang B, Guo P, Gao ZD, Ye YJ, Jiang KW and Wang S: Long noncoding
RNA HIT000218960 promotes papillary thyroid cancer oncogenesis and
tumor progression by upregulating the expression of high mobility
group AT-hook 2 (HMGA2) gene. Cell Cycle. 16:224–231. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Jendrzejewski J, He H, Radomska HS, Li W,
Tomsic J, Liyanarachchi S, Davuluri RV, Nagy R and de la Chapelle
A: The polymorphism rs944289 predisposes to papillary thyroid
carcinoma through a large intergenic noncoding RNA gene of tumor
suppressor type. Proc Natl Acad Sci USA. 109:8646–8651. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Jendrzejewski J, Thomas A, Liyanarachchi
S, Eiterman A, Tomsic J, He H, Radomska HS, Li W, Nagy R, Sworczak
K and de la Chapelle A: PTCSC3 is involved in papillary thyroid
carcinoma development by modulating S100A4 gene expression. J Clin
Endocrinol Metab. 100:E1370–E1377. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zheng H, Wang M, Jiang L, Chu H, Hu J,
Ning J, Li B, Wang D and Xu J: BRAF-activated long noncoding RNA
modulates papillary thyroid carcinoma cell proliferation through
regulating thyroid stimulating hormone receptor. Cancer Res Treat.
48:698–707. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wang Y, Guo Q, Zhao Y, Chen J, Wang S, Hu
J and Sun Y: BRAF-activated long non-coding RNA contributes to cell
proliferation and activates autophagy in papillary thyroid
carcinoma. Oncol Lett. 8:1947–1952. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Liu L, Yang J, Zhu X, Li D, Lv Z and Zhang
X: Long noncoding RNA H19 competitively binds miR-17-5p to regulate
YES1 expression in thyroid cancer. FEBS J. 283:2326–2339. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Mei JM and Niu CS: Effects of CDNF on
6-OHDA-induced apoptosis in PC12 cells via modulation of Bcl-2/Bax
and caspase-3 activation. Neurol Sci. 35:1275–1280. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Pal MK, Jaiswar SP, Srivastav AK, Goyal S,
Dwivedi A, Verma A, Singh J, Pathak AK, Sankhwar PL and Ray RS:
Synergistic effect of piperine and paclitaxel on cell fate via
cyt-c, Bax/Bcl-2-caspase-3 pathway in ovarian adenocarcinomas
SKOV-3 cells. Eur J Pharmacol. 791:751–762. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Liao YX, Zhang ZP, Zhao J and Liu JP:
Effects of fibronectin 1 on cell proliferation, senescence and
apoptosis of human glioma cells through the PI3K/AKT signaling
pathway. Cell Physiol Biochem. 48:1382–1396. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhou Y, Li S, Li J, Wang D and Li Q:
Effect of microRNA-135a on cell proliferation, migration, invasion,
apoptosis and tumor angiogenesis through the IGF-1/PI3K/Akt
signaling pathway in non-small cell lung cancer. Cell Physiol
Biochem. 42:1431–1446. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
de Oliveira JC, Oliveira LC, Mathias C,
Pedroso GA, Lemos DS, Salviano-Silva A, Jucoski TS, Lobo-Alves SC,
Zambalde EP, Cipolla GA and Gradia DF: Long non-coding RNAs in
cancer: Another layer of complexity. J Gene Med.
21:e30652019.PubMed/NCBI
|
|
23
|
Tang Y, Soroush F, Tong Z, Kiani MF and
Wang B: Targeted multidrug delivery system to overcome
chemoresistance in breast cancer. Int J Nanomedicine. 12:671–681.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yan J, Zhang Y, She Q, Li X, Peng L, Wang
X, Liu S, Shen X, Zhang W, Dong Y, et al: Long noncoding RNA
H19/miR-675 axis promotes gastric cancer via FADD/Caspase 8/Caspase
3 signaling pathway. Cell Physiol Biochem. 42:2364–2376. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Xu Y, Wang Z, Jiang X and Cui Y:
Overexpression of long noncoding RNA H19 indicates a poor prognosis
for cholangiocarcinoma and promotes cell migration and invasion by
affecting epithelial-mesenchymal transition. Biomed Pharmacother.
92:17–23. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Rothhut B, Ghoneim C, Antonicelli F and
Soula-Rothhut M: Epidermal growth factor stimulates matrix
metalloproteinase-9 expression and invasion in human follicular
thyroid carcinoma cells through focal adhesion kinase. Biochimie.
89:613–624. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Dettmer M, Perren A, Moch H, Komminoth P,
Nikiforov YE and Nikiforova MN: Comprehensive MicroRNA expression
profiling identifies novel markers in follicular variant of
papillary thyroid carcinoma. Thyroidology. 23:1383–1389. 2013.
View Article : Google Scholar
|
|
28
|
Murugan AK, Munirajan AK and Alzahrani AS:
Long noncoding RNAs: Emerging players in thyroid cancer
pathogenesis. Endocr Relat Cancer. 25:R59–R82. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wang P, Liu G, Xu W, Liu H, Bu Q and Sun
D: Long noncoding RNA H19 inhibits cell viability, migration, and
invasion via downregulation of IRS-1 in thyroid cancer cells.
Technol Cancer Res Treat. 16:1102–1112. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Mahmoudian-Sani MR, Jalali A, Jamshidi M,
Moridi H, Alghasi A, Shojaeian A and Mobini GR: Long non-coding
RNAs in thyroid cancer: Implications for pathogenesis, diagnosis,
and therapy. Oncol Res Treat. 42:136–142. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Liu N, Zhou Q, Qi YH, Wang H, Yang L and
Fan QY: Effects of long non-coding RNA H19 and microRNA let7a
expression on thyroid cancer prognosis. Exp Mol Pathol. 103:71–77.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Lee KT, Gopalan V and Lam AK: Roles of
long-non-coding RNAs in cancer therapy through the PI3K/Akt
signalling pathway. Histol Histopathol. 34:593–609. 2019.PubMed/NCBI
|
|
33
|
Wang SH, Wu XC, Zhang MD, Weng MZ, Zhou D
and Quan ZW: Long noncoding RNA H19 contributes to gallbladder
cancer cell proliferation by modulated miR-194-5p targeting AKT2.
Tumour Biol. 37:9721–9730. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Liao Z, Zhao J and Yang Y: Downregulation
of lncRNA H19 inhibits the migration and invasion of melanoma cells
by inactivating the NF-κB and PI3K/Akt signaling pathways. Mol Med
Rep. 17:7313–7318. 2018.PubMed/NCBI
|
|
35
|
Renault TT, Dejean LM and Manon S: A
brewing understanding of the regulation of Bax function by Bcl-xL
and Bcl-2. Mech Ageing Dev. 161:201–210. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Feng K, Liu Y, Xu LJ, Zhao LF, Jia CW and
Xu MY: Long noncoding RNA PVT1 enhances the viability and invasion
of papillary thyroid carcinoma cells by functioning as ceRNA of
microRNA-30a through mediating expression of insulin like growth
factor 1 receptor. Biomed Pharmacother. 104:686–698. 2018.
View Article : Google Scholar : PubMed/NCBI
|