Introduction
Breast cancer is the second most common malignancy
worldwide accounting up to 29% of all new cancers in women and the
leading cause of cancer-related mortality in women. In the last
years, significant reduction in breast cancer mortality has been
achieved in the developed world through implementing breast cancer
screening methods, particularly mammography, and developing new
therapeutic strategies. However, there are still 522,000 annual
cases of death attributed to breast cancer (1).
Breast cancer is a complex and heterogeneous disease
encompassing five distinct molecular subtypes. These include
luminal A and B, human epidermal growth factor receptor 2 (HER2)
positive, basal-like (lacking hormone receptor and HER2 expression
and therefore also called ‘triple-negative’), and normal
breast-like subtypes (2). In the
last two decades, several molecular targeting therapies against
estrogen receptor (ER), HER2 and vascular endothelial growth factor
(VEGF) have been developed (3).
However, the absence of these receptors in some breast cancer
subtypes, inherent and acquired resistance to endocrine and/or
cytostatic treatments as well as the breast cancer intra-tumor
heterogeneity make it unlikely that targeting of a single pathway
will be suitable for developing efficient and long-term breast
cancer therapy (4).
RNA interference (RNAi) holds great promise for
improving cancer management through inhibiting the pathways that
promote or sustain the growth of cancer cells by degrading
corresponding mRNAs in a sequence-specific manner (5). For the application of RNAi in cancer
therapy, one major aim is to inhibit the expression of genes that
are crucial for cell viability, allowing specific elimination of
diseased cells (6) as well as to
overcome multidrug resistance of cancer cells against chemotherapy
and radiotherapy (7).
Due to the heterogeneity of breast cancer, we have
selected a panel of siRNAs that can inhibit four vital pathways in
cancer cells and enable the specific tailoring of therapies for
particular, patient-specific needs. The siRNAs described have the
ability to inhibit vital cell pathways by silencing the gene
expression of eukaryotic elongation factor 2 (EEF2), Polo-like
kinase 1 (PLK1), G-protein coupled receptor kinase 4 (GRK4), and
sphingosine kinase interacting protein (SKIP5).
The activity of EEF2 is tightly regulated by a
specific calmodulin-dependent protein kinase. ADP ribosylation of
EEF2 by diphtheria toxin or Pseudomonas A toxin was shown to
inhibit protein biosynthesis efficiently and thus induce apoptosis
(8). These toxins are effective in
various cell types and have been used for the development of
targeted anti-cancer agents, so-called immunotoxins (8).
Several studies have reported elevated PLK-1 level
in breast cancer cells (9), whereas
it is solely overexpressed in healthy proliferative tissues
(10). PLK1 is the
best-characterized member of the human PLK family, which plays a
pivotal role in mitotic entry, spindle assembly, and DNA damage
response (11).
GRKs regulate the activity of G-protein coupled
receptors (GPCRs) via phosphorylation of their intracellular
domains after release and activation of their associated G-proteins
(12). GRKs regulate cell signaling
by phosphorylating heptahelical receptors, thereby promoting GPCR
interaction with β-arrestins.
Sphingosine kinases (SKs) are conserved lipid
kinases catalysing the formation of Sphingosine-1-phosphate (S1P)
from the precursor sphingolipid, sphingosine. S1P is characterized
as a signalling molecule with dual function. On one hand, it binds
to five different S1P receptors that are coupled to a variety of
G-proteins allowing to regulate diverse biological functions, on
the other hand it appears to act as an intracellular second
messenger but potential binding partner are still unknown (13). Inhibiting these pathways will reduce
the cell viability and promote cell apoptosis.
Considering the high therapeutic potential of RNAi,
the inefficient systemic delivery of siRNAs sets a real challenge
in the development of RNAi-based cancer therapies in general
(14). Incorporating siRNAs into
different kind of nanocarriers has improved siRNA cell
internalization and facilitate siRNA accumulation in tumor tissues
passively due to the enhanced permeability and retention effect
(EPR) (15).
Using aptamers to deliver siRNA to specific target
cells is one of the promising approaches due to some advantages
compared to nanocarriers and other ligands. For example, aptamers
can be generated against any target including toxins and
non-immunogenic targets through an in vitro chemical
process. Furthermore, they are generally generated by chemical
synthesis which reduces the structural variation from batch to
batch and allows the chemical modification of aptamer molecules as
well as unlimited shelf life (16).
In the present study, we have used the RNA aptamer
A30 that binds to the extracellular domain of human epidermal
growth factor receptor-3 (HER3) to deliver the described siRNAs
into breast cancer cells. HER3 is a member of receptor tyrosine
kinases (RTKs) family. These receptors are frequently overexpressed
on the surface of several cancers including breast cancer cells,
and are involved in several cell growth and differentiation
pathways (17).
The RNA aptamer A30 binds to the extracellular
domain of HER3 and does not compete with heregulin (HRG) binding or
inhibit HRG-dependent HER2 phosphorylation (18). However, it does reduce cell
proliferation by inhibiting HRG signaling. We set out to enhance
the anti-cancer activity of the aptamer by using it to deliver
cytotoxic siRNAs specifically to HER3-expressing breast cancer
cells (19, 20).
We found that the combined siRNAs against EEF2,
PLK-1, GRK4 and SKIP5 with aptamer A30, were taken up specifically
by HER3-expressing breast cancer cells, induced target-specific
gene silencing and ultimately suppress cell proliferation. The
limited immunogenicity and minimal off-target effects achieved by
the specific targeting of tumor cells suggest that these
aptamer-siRNA chimeras could represent an option for cancer
therapy.
Materials and methods
siRNA preparation
Synthetic 21-nt RNAs were purchased from Qiagen GmbH
(Hilden, Germany) in their deprotected, desalted and annealed form.
The EEF2-specific siRNA sequences were: sequence siEEF2-1,
5′-AGGCCUAUCUGCCCGUCAAdTdT (sense) and 5′-UUGACGGGCAGAUAGGCCUdTdG
(antisense); sequence siEEF2-2, 5′GCGCCAUCAUGGACAAGAAUUdTdT (sense)
and 5′UUCUUGUCCAUGAUGGCGCGGdGdG (antisense). The PLK1, GRK-4 and
SKIP5-specific siRNA sequences were provided by the Department of
Molecular Biology, Max Planck Institute for Infection Biology
(Berlin, Germany). siPLK1-1, 5′-CCAUAUGAAUUGUACAGAAdTdT (sense 1)
and 5′-UUCUGUACAAUUCAUAUGGdTdG (antisense 1); siPLK1-2,
5′-GGAUCAAGAAGAAUGAAUAdTdT (sense 2) and 5′-UAUUCAUUCUUCUUGAUCCdGdG
(antisense 2). siGRK-4-1, 5′-GGAUGUUACUCACCAAGAAdTdT (sense) and
5′-UUCUUGGUGAGUAACAUCCdTdG (antisense); siGRK4-2,
5′GGGUGUUUCAAAGACAUCAdTdT (sense) and 5′UGAUGUCUUUGAAACACCCdGdG
(antisense). The siSKIP5 sequences, 5′-CGUCUGGCUGCUGAUGGAAdTdT
(sense) and 5′-UUCCAUCAGCAGCCAGACGdTdT (antisense). Non-silencing
controls were also purchased from Qiagen GmbH, with the following
sequences: 5′-UUCUCCGAACGUGUCACGUdTdT (sense) and
5′-ACGUGACACGUUCGGAGAAdTdT (antisense). The siTOX siRNA (GE
Healthcare Dharmacon, Inc., Lafayette, CO, USA) was used as a
toxicity control.
Cell lines
The human HER3+ mammary adenocarcinoma cell line
MCF-7 (ATCC HTB-22) (21–23) was cultured in GIBCO™ RPMI 1640 medium
(Invitrogen; Thermo Fisher Scientific, Inc., Waltham, MA, USA)
supplemented with 10% heat-inactivated fetal bovine serum (FBS;
Invitrogen; Thermo Fisher Scientific, Inc.), 100 µg/ml penicillin
and 100 µg/ml streptomycin (Invitrogen; Thermo Fisher Scientific,
Inc.). HER3– mammary carcinoma cell line MDA-MB-231 (HTB-26)
(21–23) was maintained in DMEM tissue culture
medium (Invitrogen; Thermo Fisher Scientific, Inc.) supplemented
with 10% FBS, 100 U/ml penicillin and 100 µg/ml streptomycin. The
cells were maintained at 37°C in a humidified 5% CO2
atmosphere.
Transfections
All transfections were carried out using RNAifect
(Qiagen GmbH) following the manufacturer's recommendations.
Briefly, RNAifect (1.5 µl) was mixed with 50 µl RPMI medium
containing the appropriate concentration of siRNA and incubated for
10 min at room temperature, then added dropwise in 96-well plates
(Greiner Bio-One GmbH, Frickenhausen, Germany). Approximately
1–3×104 cells/well were seeded on top of the siRNA drop and grown
overnight (final volume 100 µl). The cells were harvested after 12,
24 or 48 h incubation for gene silencing analysis. Cell viability
was assayed after 24 or 48 h.
Gene silencing assay
Gene silencing was assessed by quantitative RT-PCR
(qRT-PCR), with a Lightcycler Faststart DNA Master SYBR Green I
(Roche Applied Science, Penzberg, Germany) and a Roche Lightcycler
1.0 system. The reaction volume was 20 µl. The primers for human
GAPDH were: GAPDH forward, 5′-CTCACTGGCATGGCCTTCCGTG-3′; GAPDH
reverse, 5′-GTACTCCAGGTGGTGGGACAACG-3′. The primers for EEF2 were:
EEF2 forward, 5′-ATGGTGAACTTCACGGTAGAC-3′; EEF2 reverse,
5′-GACTTGATGGTGATGCAACGGACTTGATGGTGATGCAACG-3′. Primers for GRK-4,
SKIP-5 and PLK-1 were the same as shown above. Briefly, 1–3×104
MCF-7 and MDA-MB-231 cells were seeded in 96-well plates (Greiner
Bio-One GmbH) and incubated overnight at 37°C. Cells were
transfected the following day with 200 nM non-silencing siRNA or
varying amounts of siRNA (15–150 nM for siRNA sequences 1 and 2)
using RNAifect Reagent (Qiagen GmbH) following the manufacturer's
recommendations. Cells were lysed after 12 h. Total RNA was
extracted using the RNeasy kit (Qiagen GmbH) and treated with DNase
I (Qiagen GmbH). RNA was eluted with 30 µl of RNase-free water.
Equal amounts of RNA were used for cDNA synthesis using random
hexamer primers and Superscript III RNA polymerase (Invitrogen;
Thermo Fisher Scientific, Inc.) following the manufacturer's
recommendations. Appropriate amounts of cDNA (2 µl) were used for
qRT-PCR. The relative amount of target gene mRNA was normalized to
GAPDH mRNA by the ΔΔCq method (24). The specificity of amplified PCR
products was verified through melting curve analysis.
Cell proliferation and viability
assay
Cell proliferation and viability was assessed using
the CellTiter 96® AQueous One solution cell
proliferation assay (MTS; Promega GmbH, Mannheim, Germany). This
assay contains a tetrazolium compound (MTS) and an electron
coupling reagent, phenazine ethosulfate (PES). MTS can be
bioreduced by cells into formazan, which is soluble in cell culture
medium. The quantity of the formazan product as measured
spectrophotometrically is directly proportional to the number of
viable cells. The assay measures dehydrogenase enzyme activity
found in metabolically active cells. MCF-7 or MDA-MB-231 cells were
seeded in 96-well plates at a density of 1–3×104 cells per well in
100 µl RPMI 1640 medium, and incubated overnight at 37°C. The cells
were washed 2 times with PBS then different concentration (10, 25,
50,75, 100, 250, 500, 1000 nM) of siRNAs were mixed with 1.5 µl
RNAifect (Qiagen GmbH) and added at a final volume of 100 µl per
well. After 48 h, the Aqueous One Solution containing MTS/PES was
added and the optical density was measured at 490 nm using an
ELx808 microplate reader (BioTek Instruments GmbH, Bad
Friedrichshall, Germany) after 6 h. This experiment was performed
in triplicate wells. The absorbance measured from control siRNA
(siNON) treatment was normalized to 100% viability.
Predicting RNA secondary
structure
The RNA structure program MFOLD (http://unafold.rna.albany.edu/?q=mfold/download-mfold)
was used to predict the secondary structures of aptamer A30 and
aptamer-siRNA transcripts. The negative control A30-siGFP was
designed accordingly. The most stable structures with the lowest
free energies for each construct were compared.
In vitro RNA transcription
The template DNAs for the aptamer-siRNA transcripts
were synthesized by assembly-PCR using six overlapping primer
sequences (Table I) as described by
Rydzanicz et al (25). The
aptamer (A30) and the aptamer-siRNA transcripts (Table II) were synthesized by in
vitro transcription from a dsDNA template containing a T7 RNA
polymerase promoter. Transcriptions were performed in a 100-µl
reaction volume using T7 RNA transcription kit (Agilent
Technologies GmbH, Waldbronn, Germany). Reactions were carried out
overnight at 37°C and then treated with DNaseI for 20 min at 37°C.
The RNA products were purified by denaturing (7 M urea) gel
electrophoresis on an 8% polyacrylamide gel. RNA bands were
excised, and RNA was eluted in 0.3 M sodium acetate for 1 h at 60°C
and recovered by ethanol precipitation.
 | Table I.DNA sequences of synthetic
oligonucleotides for RNA transcripts. |
Table I.
DNA sequences of synthetic
oligonucleotides for RNA transcripts.
| Primer name | Sequence
(5′-3′) |
|---|
| A30 |
|
| 5′
A30 | TAA TAC GAC TCA CTA
TAG GGA ATT CCG CGT GTG CC |
| 3′
A30 | GAG GAT CCC GAA CGG
ACC GCC |
|
A30-1 | GGG AAT TCC GCG TGT
GCC AGC GAA AGT TGC GTA TGG GTC ACA |
|
A30-2 | ACG GAC CGC CCA GAT
GAC ATG TGC CTG CGA TGT GAC CCA TAC GCA ACT T |
|
A30-3 | CAT CTG GGC GGT CCG
TTC GGG ATC CTC |
| A30-siEEF2 |
|
| 5′
A30-EEF2 | 5′ A30 |
|
3′A30-EEF2 | CCG CGC CAT CAT GGA
CAA GAA GAA GC |
|
A30-EEF2-1 | GGG AAT TCC GCG TGT
GCC AGC GAA AGT TGC GTA TGG GTC ACA |
|
A30-EEF2-2 | ACG GAC CGC CCA GAT
GAC ATG TGC CTG CGA TGT GAC CCA TAC GCA ACT T |
|
A30-EEF2-3 | CAT CTG GGC GGT CCG
TTC GGG ATC CTC GAA GCT AGC GCC ATC ATG GAC A |
|
A30-EEF2-4 | ATG GAC AAG AAG AAG
CTT CAA TTC TTG TCC ATG ATG GCG CTA G |
| A30-siGRK4 |
|
| 5′
A30-GRK4-1 | 5′ A30 |
| 3′
A30-GRK4-1 | CAG GAT GTT ACT CAC
CAA GAA GAA GC |
|
A30-GRK4-1-1 | A30-EEF2-1 |
|
A30-GRK4-1-2 | A30-EEF2-2 |
|
A30-GRK4-1-3 | CAT CTG GGC GGT CCG
TTC GGG ATC CTC GAA GCT AGG ATG TTA CTC ACC A |
|
A30-GRK4-1-4 | CTC ACC AAG AAG AAG
CTT CAA TTC TTG GTG AGT AAC ATC CTA G |
| 5′
A30-GRK4-2 | 5′ A30 |
| 3′
A30-GRK4-2 | CCG GGT GTT TCA AAG
ACA TCA GAA GC |
|
A30-GRK4-2-1 | A30-EEF2-1 |
|
A30-GRK4-2-2 | A30-EEF2-2 |
|
A30-GRK4-2-3 | CAT CTG GGC GGT CCG
TCC GGG ATC CTC GAA GCT AGG GTG TTT CAA AGA C |
|
A30-GRK4-2-4 | CAA AGA CAT CAG AGG
CTT CAA TTC TTG GTG AGT ACC ATC CAT G |
| A30-siPLK1 |
|
| 5′
A30-PLK1-1 | 5′ A30 |
| 3′
A30-PLK1-1 | CAC CAT ATG AAT TGT
ACA GAA GAA GC |
|
A30-PLK1-1-1 | A30-EEF2-1 |
|
A30-PLK1-1-2 | A30-EEF2-2 |
|
A30-PLK1-1-3 | CAT CTG GGC GGT CCG
TTC GGG ATC CTC GAA GCT ACC ATA TGA ATT GTA C |
|
A30-PLK1-1-4 | ATT GTA CAG AAG AAG
CTT CAA TTC TGT ACA ATT CAT ATG GT G |
| 5′
A30-PLK1-2 | 5′ A30 |
| 3′
A30-PLK1-2 | CCG GAT CAA GAA GAA
TGA ATA GAA GC |
|
A30-PLK1-2-1 | A30-EEF2-1 |
|
A30-PLK1-2-2 | A30-EEF2-2 |
|
A30-PLK1-2-3 | CAT CTG GGC GGT CCG
TTC GGG ATC CTC GAA GCT AGG ATC AAG AAG AAT G |
|
A30-PLK1-2-4 | AAG AAT GAA TAG AAG
CTT CAA TAT TCA TTC TTC TTG ATC CTA G |
| A30-siSKIP5 |
|
| 5′
A30-SKIP5 | 5′ A30 |
| 3′
A30-SKIP5 | AAC GTC TGG CTG CTG
ATG GAA GAA GC |
|
A30-SKIP5-1 | A30-EEF2-1 |
|
A30-SKIP5-2 | A30-EEF2-2 |
|
A30-SKIP5-3 | CAT CTG GGC GGT CCG
TTC GGG ATC CTC GAA GCT ACG TCT GGC TGC TGA T |
| A30-GFP |
|
| 5’
A30-GFP | 5’ A30 |
| 3’
A30-GFP | CGG CAA GCT GAC CCT
GAA GTT CCA AGC |
|
A30-GFP-1 | A30-EEF2-1 |
|
A30-GFP-2 | A30-EEF2-2 |
|
A30-GFP-3 | CAT CTG GGC GGT CCG
TTC GGG ATC CTC GGA AGC TTG CAA GCT GAC CCT G |
|
A30-GFP-4 | CTG AAG TTC CAA GCT
TCA TGA ACT TCA GGG TCA GCT TGC AAG C |
| Table II.Sequences of RNA constructs. |
Table II.
Sequences of RNA constructs.
| Aptamer | Sequence
(5′-3′) |
|---|
| A30 |
GGGAAUUCCGCGUGUGCCAGCGAAAGUUGCGUAUGGGUCACAUCGCA
GGCACAUGUCAUCUGGGCGGUCCGUUCGGGAUCCUC |
| A30-siEEF2 |
GGGAAUUCCGCGUGUGCCAGCGAAAGUUGCGUAUGGGUCACAUCGCA
GGCACAUGUCAUCUGGGCGGUCCGUUCGGGAUCCUCGAAGCUAGCGC
CAUCAUGGACAAGAAUUGAAGCUUCUUCUUGUCCAUGAUGGCGCGG |
| A30-siPLK1-1 |
GGGAAUUCCGCGUGUGCCAGCGAAAGUUGCGUAUGGGUCACAUCGCA
GGCACAUGUCAUCUGGGCGGUCCGUUCGGGAUCCUCGAAGCUAGGGU
GUUUCAAAGACAUCAUUGAAGCUUCUGAUGUCUUUGAAACACCCGG |
| A30-siPLK1-2 |
GGGAAUUCCGCGUGUGCCAGCGAAAGUUGCGUAUGGGUCACAUCGCA
GGCACAUGUCAUCUGGGCGGUCCGUUCGGGAUCCUCGAAGCUAGGAU
CAAGAAGAAUGAAUAUUGAAGCUUCUAUUCAUUCUUCUUGAUCCGG |
| A30-siSKIP5 |
GGGAAUUCCGCGUGUGCCAGCGAAAGUUGCGUAUGGGUCACAUCGCA
GGCACAUGUCAUCUGGGCGGUCCGUUCGGGAUCCUCGAAGCUACGUC
UGGCUGCUGAUGGAAUUGAAGCUUCUUCCAUCAGCAGCCAGACGUU |
| A30-siGRK4-1 |
GGGAAUUCCGCGUGUGCCAGCGAAAGUUGCGUAUGGGUCACAUCGCA
GGCACAUGUCAUCUGGGCGGUCCGUUCGGGAUCCUCGAAGCUAGGAU
GUUACUCACCAAGAAUUGAAGCUUCUUCUUGGUGAGUAACAUCCUG |
| A30-siGRK4-2 |
GGGAAUUCCGCGUGUGCCAGCGAAAGUUGCGUAUGGGUCACAUCGCA
GGCACAUGUCAUCUGGGCGGUCCGUUCGGGAUCCUCGAAGCUAGGGU
GUUUCAAAGACAUCAUUGAAGCUUCUGAUGUCUUUGAAACACCCGG |
Fluorescence labeling of RNA
In order to fluorescently labeling the 5′-ends of
generated RNAs, 25 mM guanosine-5′-O-monophosphothioate (GMPS) was
added to the transcription reaction. After DNase I digestion the
thiol group of GMPS was reduced using 100 mM DTT and incubated for
1 h. Then, 1.5 nmol of RNA transcript was incubated with 0.5 mg/ml
5′-iodoacetamidofluoresceine (IAF), 10 mM EDTA, 1 M urea, 100 mM
Tris-HCl (pH 7.4) and 10% v/v dimethylformamide s(DMF). The
reaction was incubated at 4°C in the dark overnight. Labeled
transcripts were purified by ethanol precipitation, washed several
times with 70% ethanol and then resuspended in RNase free
water.
Cell surface binding studies of
aptamer-siRNA transcripts
Specific binding of the aptamer transcripts to HER3+
cells was determined by flow cytometry. Briefly, IAF-labeled RNA
transcripts were first heated to 80°C for 3 min and then incubated
at 37°C for 10 min in binding buffer (20 mM HEPES, 150 mM NaCl, 1
mM MgCl2, 1 mM CaCl2, pH 7.4). HER3+ MCF-7 cells and HER3–
MDA-MB-231 cells were washed twice in ice-cold PBS and resuspended
in binding buffer, then 2–5×105 cells were incubated at 4°C for 1 h
with the IAF-labeled aptamer A30 at a concentration of 100 nM or
aptamer-siRNA transcripts at a concentration of 10 and 100 nM,
respectively. After an additional washing step, FACS profiles were
acquired based on the analysis of at least 10,000 events in a
FACSCalibur (Becton Dickinson, Heidelberg, Germany). Data analysis
was performed with CellQuest Software (Becton Dickenson). HER3–
control cell lines were treated as described above using RNA
transcript concentrations of 100 nM.
Internalization assay
HER3+ MCF-7 and MDA-MB-231 control cells (2×105)
were washed with 500 µl HEPES binding buffer and incubated with 100
nM FITC-labeled aptamer-siRNA in binding buffer supplemented with
16 mM glucose. The ‘non-internalization’ controls for each
transcript were kept on ice. All other samples were incubated in
the dark at 37°C for 30–120 min. Thereafter, cells were cooled in
ice for 5 min. In order to remove aptamer-siRNA transcripts bound
to HER3 on cell surface, 0.5 mg/ml proteinase K was added to the
cells in HEPES binding buffer. For the ‘100% internalization’ value
of each transcript, the proteinase K treatment was omitted so that
the maximum number of internalized transcripts reflected the amount
of bound fluorescence. After 15 min, cells were collected, washed
twice, resuspended in 400 µl binding buffer and analyzed by flow
cytometry. For each transcript, data were normalized to the
untreated samples (100% internalization) and the no internalization
controls (0% internalization).
Aptamer-siRNA transcripts-mediated
cytotoxicity
Cell proliferation and viability assay was performed
using MTS assay as described before. Briefly, Approximately, 2×104
MCF-7 or MDA-MB-231 cells were seeded in 96-well plates, and
allowed to attach overnight at 37°C. The cells were washed 2 times
with PBS then the cells were treated with increased concentrations
(10, 25, 50, 75, 100, 250, 500, 1000 nM) of A30-siRNAs. After 48 h,
the Aqueous One Solution containing MTS/PES was added and the
optical density was measured at 490 nm using an ELx808 microplate
reader after 6 h. The absorbance measured from control siRNA
(siNON) treatment was normalized to 100% viability.
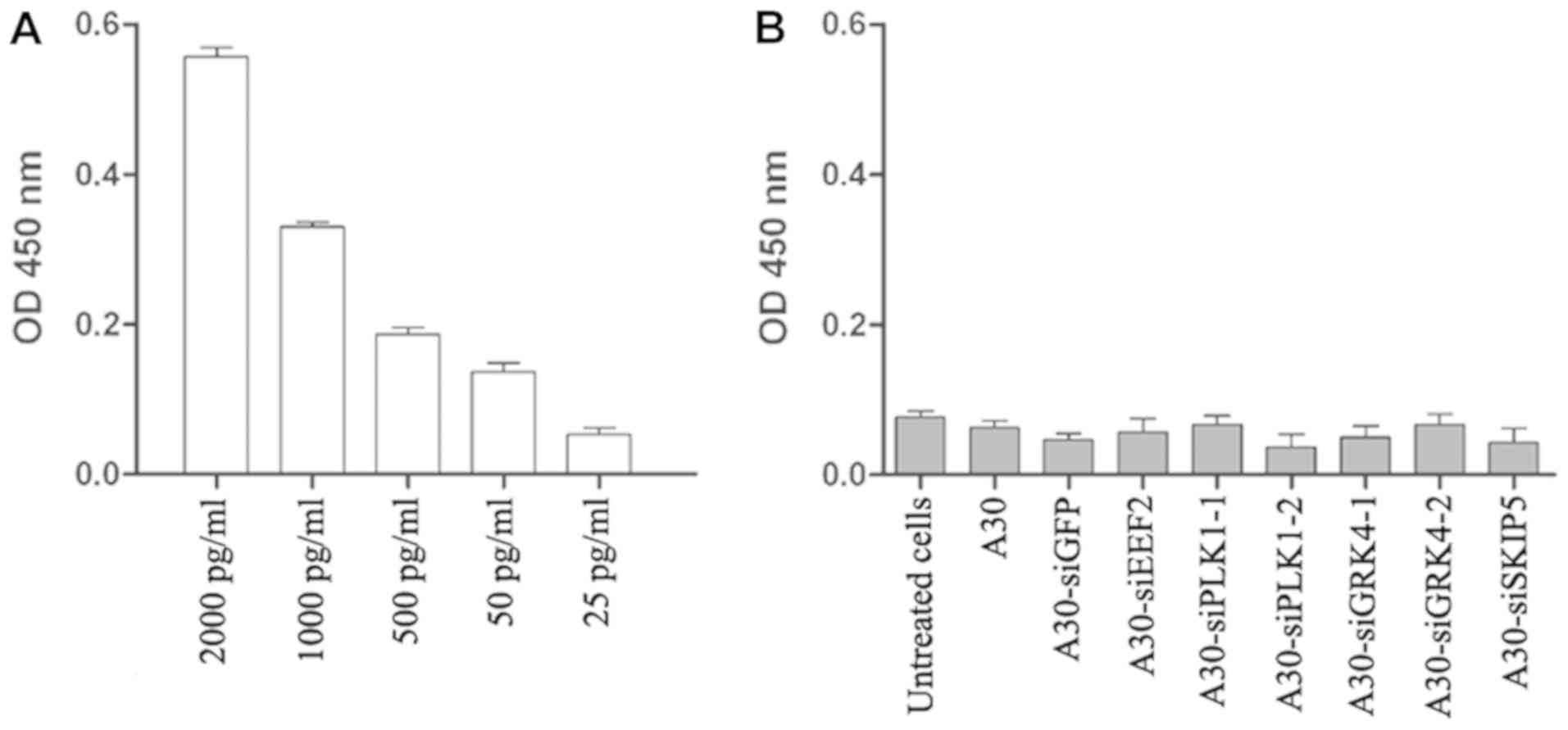
Interferon assay
In order to analyze the aptamer-siRNA
immunogenicity, the secretion of interferon β into the supernatant
of MCF-7 cells was examined. This was done after treating the cells
with the aptamer-siRNA chimera using the human interferon β ELISA
kit (PBL Biomedical Laboratories, New Jersey, United States).
Briefly, 1×104 MCF-7 cells were seeded in 96-well plates and
incubated overnight at 37°C. Thereafter, 1 µM of A30 or
aptamer-siRNA was added to the cells and incubated for a further 48
h. Then 100 µl of the supernatant was transferred to a pre-coated
ELISA plate and as a positive control different concentration (25,
50, 500, 1000 and 2000 pg/ml) of human IFNβ have been used and
incubated for further 24 h at room temperature. Interferon β was
detected using an antibody specific to human Interferon β following
the manufacturer's recommendations.
Data analysis
GraphPad Prism software v5.00 for Windows (GraphPad
Software, San Diego, California, USA) was used for statistical
analyses and curve fitting. Data represent the average of
triplicates ± SEM. The Student's t-test and and two-way
repeated-measure ANOVA followed by Sidak's multiple comparisons
test were used to assess the significance of independent
experiments. P<0.05 was considered to indicate a statistically
significant difference.
Results
Silencing potency of selected
siRNAs
RNAi-based genome-wide screens have provided a
powerful tool for the identification of cytotoxic siRNAs. We have
recognized siRNAs that induced the knock-down of PLK1-1 (83.6 ±
8.6%), PLK1-2 (91.5 ± 1.3%), and SNW1-5 (86.9 ± 1.2%) in HeLa cells
(data not shown). A further target, GRK4, was discovered by
phenotypic analysis of different cell lines (data not shown). Short
interfering RNA sequences targeting EEF2, PLK1, GRK4 and SNW1 mRNA
were designed by Qiagen GmbH, using their HP OnGuard siRNA Design
algorithm. Their silencing activity in MCF-7 and MDA-MB-231 human
mammary adenocarcinoma cells was evaluated at two concentrations
(15 and 150 nM). The corresponding mRNA levels were monitored by
quantitative real time RT-PCR (qPCR) 12 h post-transfection, to
ensure that any knock-down effects were siRNA-specific and not
artifacts caused by interferon-induced apoptosis. At 150 nM, all
the tested siRNAs significantly reduced the expression of their
target mRNAs with the exception of the non-silencing control
(siNON). Among them, the siPLK1-1 and siSNW1-5 constructs were the
most potent, resulting in up to 80% knock-down of target mRNA
levels. Transfection of siEEF2 and siGRK4-1 reduced the target mRNA
levels up to 75% (Fig. 1A).
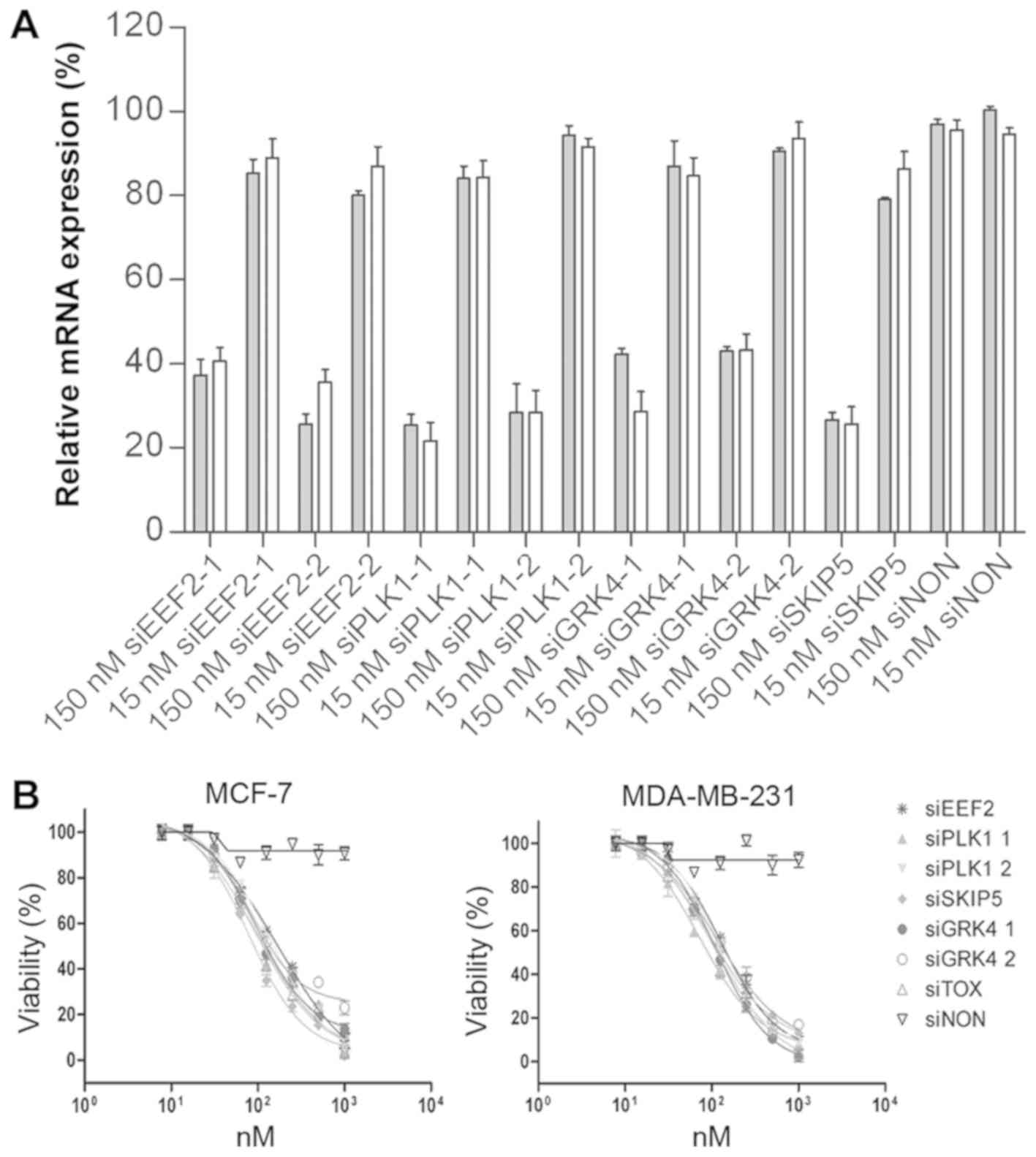 | Figure 1.Gene silencing activity of cell-death
promoting siRNAs. (A) RT-qPCR analysis of MCF-7 and MDA-MB-231cells
transfected with siRNA. Both cell lines were transfected with 15
and 150 nM siRNA against different target genes (EEF2, PLK1, GRK4
and SNW1 mRNA). A siNON was used as the negative control and
reference for normalisation. After 12 h, total RNA was isolated and
cDNA synthesized using random hexamer primers (Invitrogen; Thermo
Fisher Scientific, Inc.). The mRNA levels were normalized to GAPDH
mRNA. Error bars represent SEM (n=3). (B) Toxic activity of
cell-death promoting siRNAs. The cytotoxicity of siEEF2, siPLK1-1,
siPLK1-2, siSKIP5, siGRK4-1 and siGRK4-2, was evaluated against
MCF-7 and MDA-MB-231 cells using different concentrations (10, 25,
50,75, 100, 250, 500, 1000) nM of each siRNA. A siNON was used as
the negative control and the siTOX as the positive control at the
same concentrations. EEF2, eukaryotic elongation factor 2; PLK1,
polo-like kinase 1; GRK4, G protein-coupled receptor kinase 4;
siNON, non-silencing siRNA; siTOX, toxic siRNA |
siRNA toxic activity
The potent ability of six selected siRNAs to
eliminate breast cancer cells was determined by analyzing the cell
proliferation and viability of treated cells. To determine the
cytotoxicity of selected siRNAs to MCF-7 and MDA-MB-231 cells, cell
proliferation and viability assay was performed 48 h after
transfection. The viability of treated cells was reduced
significantly, in a concentration-dependent manner. The IC50 values
ranged from 79 nM for SKIP-5 to 158 nM for siEEF2. Exposing MCF-7
and MDA-MB-231 cells to negative control siRNA had no effect on
cell viability, while universally cytotoxic siTOX siRNA (Dharmacon,
Chicago, USA) showed comparable toxic effect on treated cells
(Table III and Fig. 1B).
| Table III.IC50 values of cytotoxic
siRNAs. |
Table III.
IC50 values of cytotoxic
siRNAs.
| siRNA |
IC50 |
|---|
| siSKIP-5 | 79
nM |
| siPLK1-1 | 129 nM |
| siGRK4-1 | 130 nM |
| siPLK1-2 | 131 nM |
| siGRK4-2 | 140 nM |
| siEEF2 | 158 nM |
| siTOX | 126 nM |
| siNON | −− |
Design of aptamer-siRNA
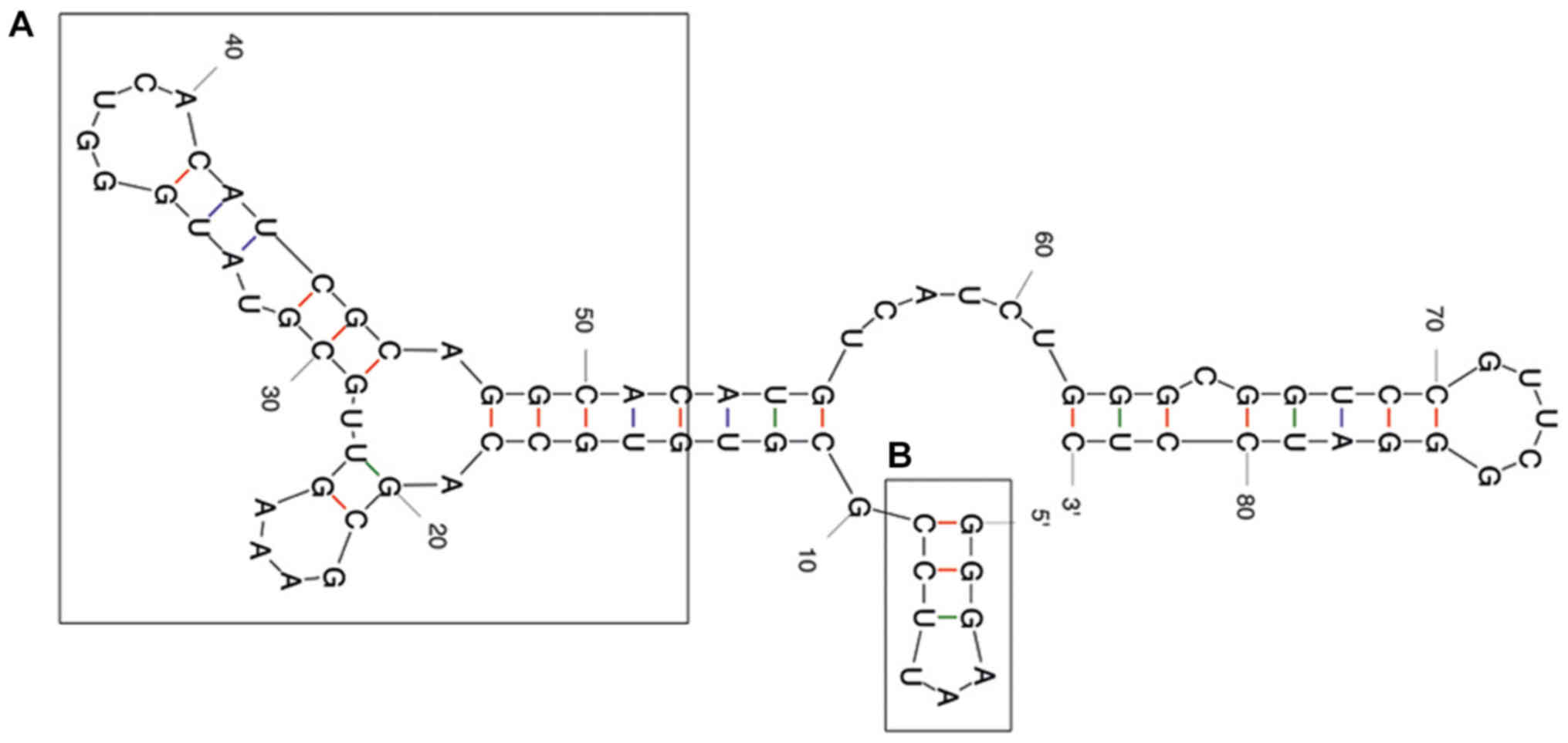
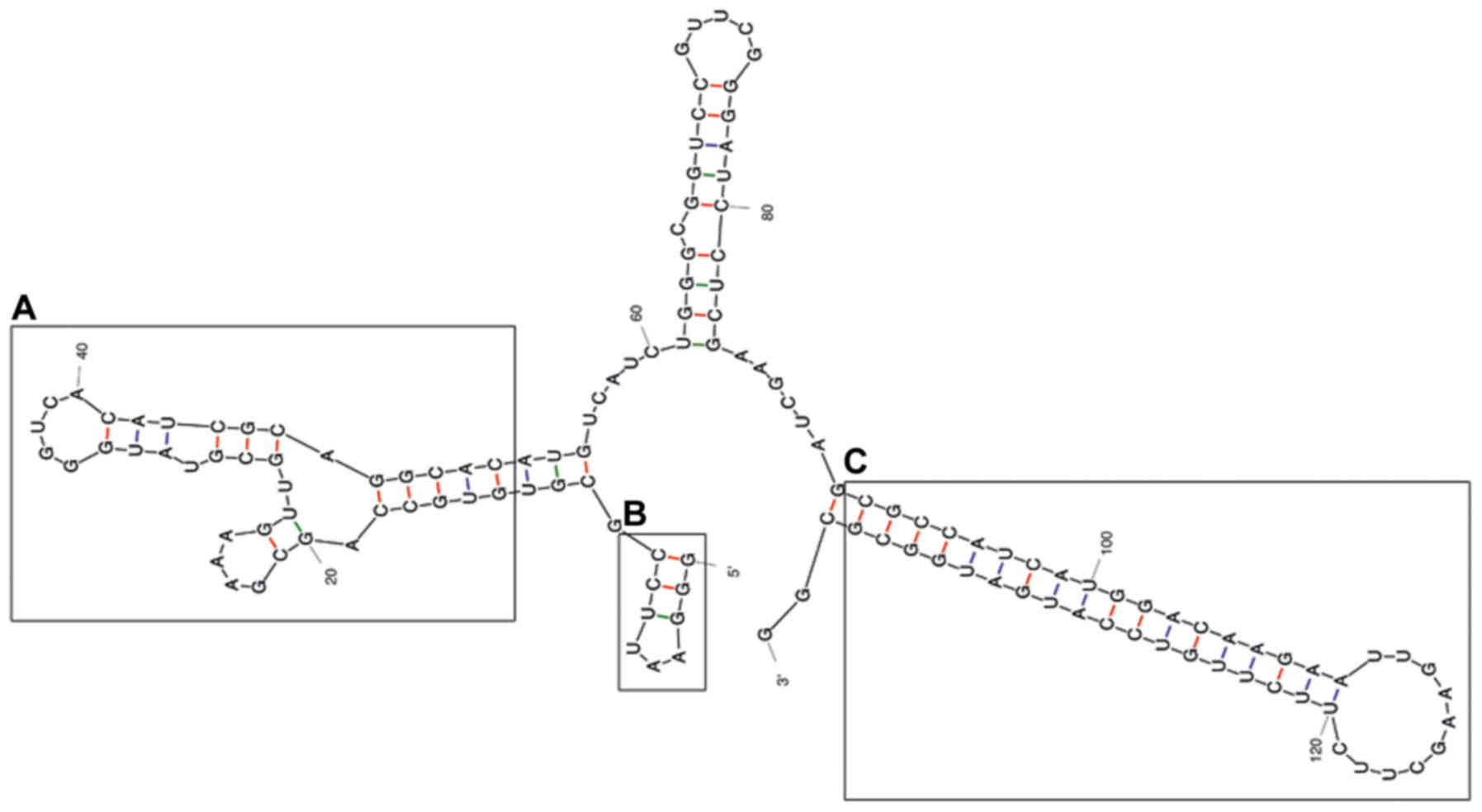
transcripts
The predicted secondary structure of the A30 aptamer
using the MFOLD web server showed that the full-length aptamer
folded into four main stem loop structures (Fig. 2). No significant alteration of
predicted secondary structure of the A30 aptamer has been observed
after adding siRNA sequence to the 3′ end of the aptamer as a short
hairpin RNA (Fig. 3). The putative
stabilization loop (Fig. 3B) has
been shown to have an important impact for functional folding of
both the (Fig. 3A) aptamer domain
and (Fig. 3C) shRNA stem loop. A
non-silencing control construct was generated with a siRNA sequence
against GFP (siGFP).
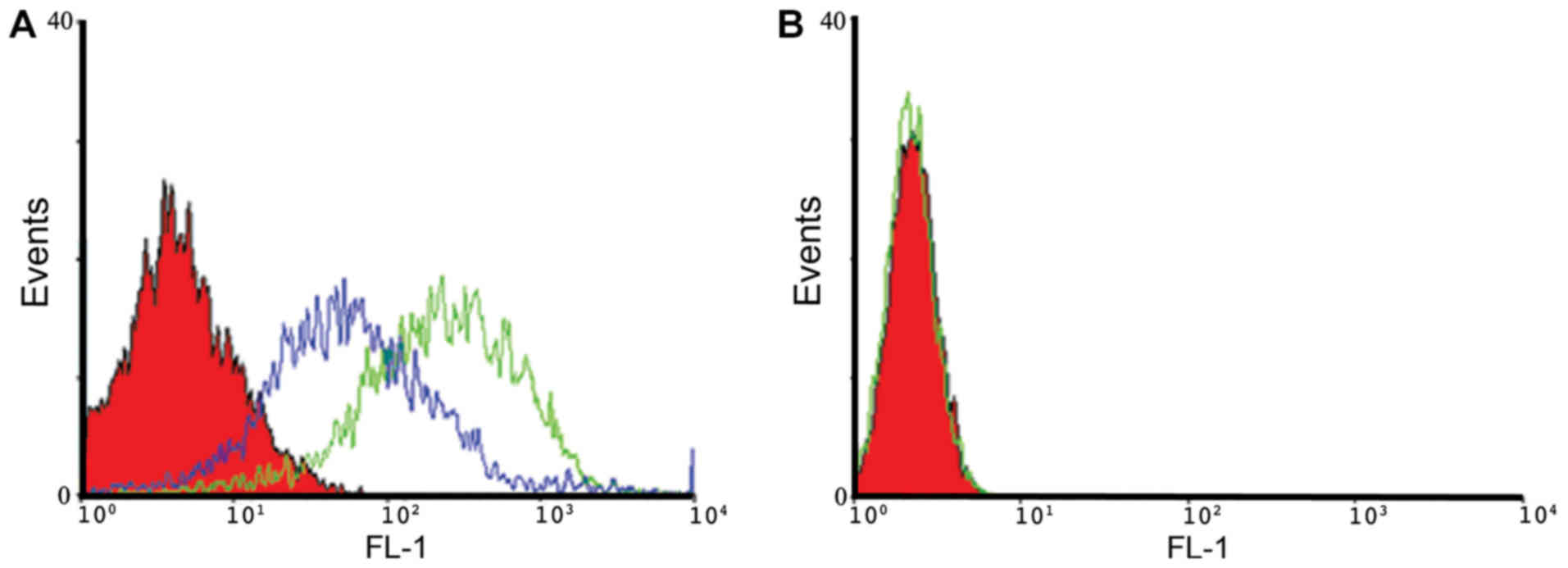
Binding and cellular uptake of
aptamer-siRNA transcripts
The binding properties of A30 aptamer to HER3
positive MCF-7 cells and MDA-MB-231 negative cells were analyzed
using flow cytometry. We observed specific binding of IAF-labeled
aptamer-siRNA transcripts to the HER3+ MCF-7 cells, as shown
exemplarily for A30-siEEF2 transcripts (Fig. 4). The binding activity of IAF-labeled
aptamer-siRNA transcripts was increased in a dose-dependent manner.
This was confirmed by using two different RNA concentrations of 10
and 100 nM. Furthermore, IAF-labeled RNA transcripts were solely
bound to HER3 receptor as no fluorescence signal has been detected
using the antigen-negative MDA-MB-231 cells.
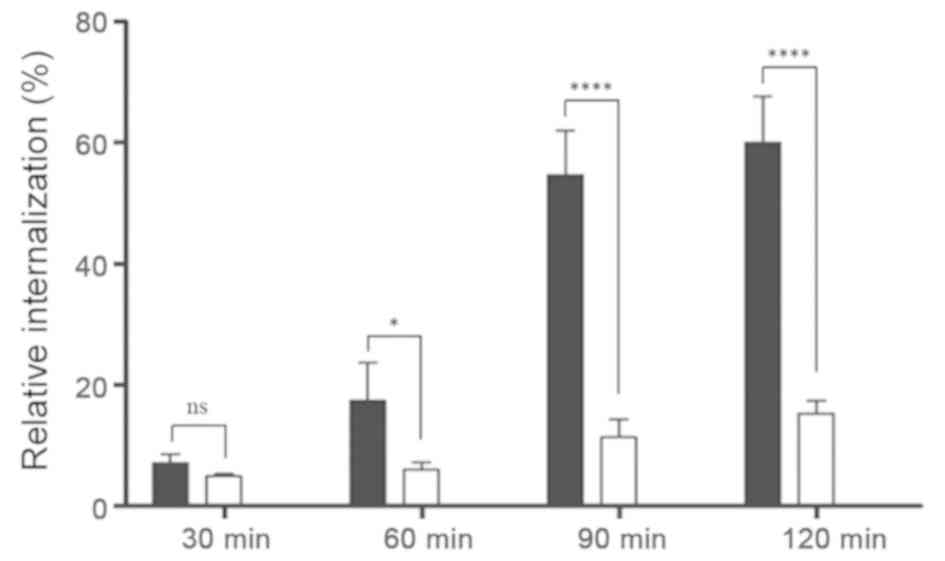
To determine whether aptamer-siRNA transcripts were
taken up by HER3+ MCF-7 cells, the rate of aptamer-siRNA
internalization was studied. Here, proteinase K was used to remove
the HER3 extracellular domain and attached transcripts, allowing to
detect only internalized IAF-labeled RNA transcripts. The results
of this experiment revealed that the amount of internal A30-siRNA
increased steadily until 60% was internalized (Fig. 5). This indicates that the
aptamer-siRNA transcripts were internalized via a receptor-mediated
endocytosis process in HER3+ cancer cells.
Aptamer-siRNA-mediated cellular
cytotoxicity
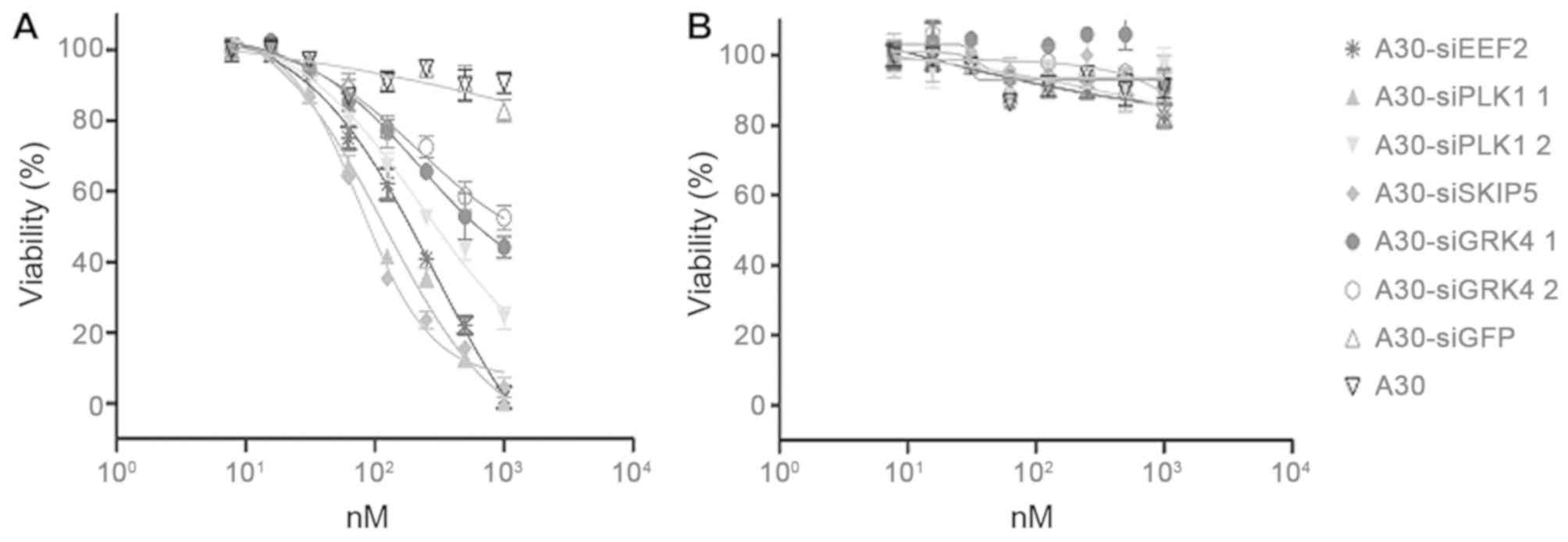
The ability of A30-siEEF2, A30-siPLK1, A30-siSKIP5
and A30-siGRK4 to inhibit HER3+ cell proliferation was analyzed
using a MTS-based colorimetric cell proliferation assay with the
HER3+ cell line MCF-7 and Her3- MDA-MB-231 cells as a negative
control. The cell viability of HER3+ MCF-7 cells was found to
decrease significantly in a concentration dependent manner after
incubation with the different A30-siRNA constructs. The IC50 values
were 89 nM for A30-siSKIP-5, 125 nM for A30-siPLK1-1, 199 nM for
A30-siEEF2 and 501 nM for A30-siGRK4-1, respectively (Fig. 6A and Table IV). MDA-MB-231 cells remained
unaffected even when treated with 1000 nM of the aptamer-siRNA
transcripts (Fig. 6B). Furthermore,
neither A30-siGFP nor A30 aptamer alone exerted any cytotoxicity to
both tumor cell lines.
| Table IV.IC50 values of
aptamer-siRNA transcripts. |
Table IV.
IC50 values of
aptamer-siRNA transcripts.
| siRNA |
IC50 |
|---|
| A30-siSKIP-5 | 89
nM |
| 30-siPLK1-1 | 125 nM |
| A30-siGRK4-1 | 501 nM |
| A30-siPLK1-2 | 316 nM |
| A30-siEEF2 | 199 nM |
| siGFP | −− |
Interferon assay
A major safety concern regarding the use of
siRNA-based therapy is the induction of nonspecific inflammatory
responses and subsequent cellular cytotoxicity. Therefore, we
determined the interferon β level after treating MCF-7 cells with
all aptamer-siRNA transcripts for 24 h. The interferon β levels in
treated cells were lower than the background levels observed in
untreated cells, indicating that under our experimental conditions
the aptamer-siRNA transcripts did not trigger a type I interferon
response (Fig. 7).
Discussion
Since the first description of the RNAi process by
Fire et al (26), significant
progress has been made in understanding the technical details and
in selecting siRNA against a wide range of target genes. The use of
siRNA has several therapeutic advantages compared to conventional
drugs. RNAi is an endogenous natural process generally used by
cells to selectively down-regulate gene expression or as defense
mechanism against viral infection (27,28). As
cancer diseases are associated with the up-regulation of specific
genes, their specific knock down in cancer cells with the help of
siRNA emerged as a highly promising therapeutic option.
Furthermore, siRNA-based therapies have the ability to overcome
multidrug resistance of cancer cells against chemotherapy,
photodynamic therapy and radiotherapy. Remarkable progress has been
made to select siRNA against target genes (27).
Due to breast cancer heterogeneity at the population
and single-cell level, its complexity and its metastasizing
potential (29), targeting different
pathways that could inhibit growth and invasion of cancer cells is
essential for developing effective therapies. Here we have tested
several siRNAs targeting not only one but a set of different
pathways to allow tailored development of siRNA-based therapeutic
agents to ideally hit the vast majority of breast cancer cells and
subtypes. As promising siRNAs candidates against four gene products
overexpressed and/or sustaining cancer cell growth have been
investigated. The corresponding toxic siRNAs are directed against
PLK-1 (30–32) and EEF-2 (33), as well as GRK4 and SKIP-5 (34). All tested siRNAs induced toxic effect
in breast cancer cells, however siSKIP-5 was two times more toxic
than other tested siRNAs. Recent reports indicated that sphingosine
kinase is overexpressed in breast cancer cells and serve as an
oncogene in tumorigenesis by enhancing tumor cell growth and
invasion and reducing cell apoptosis (35).
As described in previous studies, siSKIP-5 was able
to reduce the sphingosine kinase expression level and induce cell
death in HER3+ breast cancer cells (36). Although, this could reflect the high
toxic activity of siSKIP-5 comparing to other siRNAs, several
reasons could also play an essential role in knock down
experiments.
The inefficient systemic delivery of siRNAs, due to
their hydrophilicity, negative charge and sensitivity to nucleases,
poses challenges in the development of RNAi-based cancer therapies
in general (14). To solve the
aforementioned problems, siRNAs have been armed with several
classes of specific cell receptor ligands to direct them to the
target cells. These ligands include folic acid, aptamers,
monoclonal antibodies and their fragments (37–39).
Over the past years, several studies have reported
that arming of siRNAs with aptamers could significantly improve
therapeutic efficacy and pharmacokinetic properties of siRNA
(31).
In the present study, we chose the HER3-specific
aptamer A30 for the cell type-specific delivery of novel cytotoxic
siRNAs. HER3 is a member of the EGFR family, and is overexpressed
in diverse human cancers; it is associated with poor prognosis in
breast, lung and ovarian cancer (40,41).
Several publications emphasize the therapeutic potential of HER3
targeting, particularly for the treatment of drug-resistant tumors
(42). Here, several RNA transcripts
have been developed by fusing the HER3-specific aptamer A30 to
different cytotoxic siRNAs. The resulting transcripts maintained
high binding activity for cell-surface HER3. In comparison to
aptamer A30 alone, aptamer-siRNA conjugates showed an increased
binding to target cells. This improved binding activity might be
due to potential alterations in their RNA secondary structure that
increased duplex stability and nuclease resistance and thus
prevents degradation by RNases. However, the binding specificity of
A30 and A30-siRNA was confirmed using MDA-MB-231 cells which
express very low levels of HER3. The binding specificity need to be
further determined after knocking down HER3 gene in MCF-7
cells.
A30 aptamer-siRNA constructs were rapidly taken up
specifically into HER3+ MCF-7 cells even in the absence
of heregulin (HRG), which is normally required to trigger receptor
mediated endocytosis of HER3 (43).
Furthermore, the inhibition of HRG signaling in vivo by
aptamer A30 could have synergistic antitumor effects thus
increasing its therapeutic potential (18).
The A30-siRNAs specifically reduced the mRNA levels
of four different target genes to the same extent as the
transfected siRNAs indicating that the siRNAs also retained their
activities as part of a fusion transcript. In HER3+
MCF-7 cells, the knock-down of each target mRNA was sufficient to
reduce the cell viability whereas HER– MDA-MB-231 cells were not
affected. The IC50 values for the overall cytotoxicity
varied from 89 nM for SKIP-5 to 501 nM for GRK4-1, which was
comparable to previous reports with other cytotoxic siRNAs, and a
previously reported aptamer-siRNA targeting PSMA (31,33). In
the cell viability assay only MCF-7 cells were affected, and the
toxicity was dose-dependent. To confirm that these effects
reflected specific RNAi and not simply the binding of A30 to the
cell surface, we also analyzed A30 without a siRNA fusion and A30
fused to a non-silencing siRNA. Cytotoxic effects were observed
only in cells incubated with RNA transcripts that containing
cytotoxic siRNA.
Furthermore, simultaneous targeting of HER family
members might represent a promising therapeutic option to overcome
the heterogeneity of breast cancer. Nucleic acid aptamer-siRNA
constructs targeting HER2 and HER3 have recently demonstrated their
promising therapeutic properties in vivo against different
breast cancer cell lines. Therefore, combining siRNAs targeting
different gene transcripts with dual targeting aptamers could allow
the development of novel therapeutic options especially for breast
cancer patients (44).
Cell type-specific siRNA delivery is particularly
important for cytotoxic siRNAs because nonspecific uptake could
also kill healthy cells. HER3 is therefore a promising target for
cell-specific drug delivery because it is predominantly expressed
on breast, ovarian and lung cancer cells. Although more work is
clearly required to further confirm these results in vivo,
these initial experiments suggest that combining the highly
specific HER3 aptamer with cytotoxic siRNAs allows the development
of highly efficient agents for selective elimination of
HER3-expressing malignant cells.
Acknowledgements
The authors would like to thank Dr. Richard M.
Twyman (Twyman Research Management Ltd.) for critically reading the
manuscript.
Funding
The present study was partly supported by the
Florindon Foundation, the German Research foundation, DFG
(Graduiertenkolleg ‘Biointerface’ 1035), and by funding under the
Sixth Research Framework Programme of the European Union, Project
RIGHT (grant no. LSHB-CT-2004-005276).
Availability of data and materials
All data generated or analyzed during the present
study are included in this published article and its supplementary
information files.
Authors' contributions
SB, RF and MKT conceived the study. IN performed the
experiments with help from UW. NM and TFM identified the cytotoxic
siRNAs. SB, MKT, AFH, SG and IM discussed, analyzed and interpreted
the results. AFH and MKT wrote the manuscript with essential input
from all authors.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Conflicts of interest
The authors declare that they have no competing
interest.
Glossary
Abbreviations
Abbreviations:
|
DMF
|
dimethylformamide
|
|
EEF2
|
eukaryotic elongation factor 2
|
|
EGFR
|
epidermal growth factor receptor
|
|
EPR
|
enhanced permeability and retention
effect
|
|
ER
|
estrogen receptor
|
|
IAF
|
iodoacetamidofluoresceine
|
|
GPCRs
|
G-protein coupled receptors
|
|
GRK4
|
G protein-coupled receptor kinase
4
|
|
HER2
|
human epidermal growth factor
receptor-2
|
|
HER3
|
human epidermal growth factor
receptor-3
|
|
HRG
|
heregulin
|
|
PES
|
phenazine ethosulfate
|
|
PLK1
|
polo-like kinase 1
|
|
RNAi
|
RNA interference
|
|
RTKs
|
receptor tyrosine kinases
|
|
S1P
|
sphingosine-1-phosphate
|
|
siGFP
|
siRNA against GFP
|
|
siNON
|
control siRNA
|
|
siRNA
|
small interfering RNA
|
|
siTox
|
toxic siRNA
|
|
SKIP5
|
sphingosine kinase interacting
protein
|
|
SKs
|
sphingosine kinases
|
|
VEGF
|
vascular endothelial growth factor
|
References
|
1
|
Ferlay J, Soerjomataram I, Dikshit R, Eser
S, Mathers C, Rebelo M, Parkin DM, Forman D and Bray F: Cancer
incidence and mortality worldwide: Sources, methods and major
patterns in GLOBOCAN 2012. Int J Cancer. 136:E359–E386. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Subik K, Lee JF, Baxter L, Strzepek T,
Costello D, Crowley P, Xing L, Hung MC, Bonfiglio T, Hicks DG, et
al: The expression patterns of ER, PR, HER2, CK5/6, EGFR, Ki-67 and
AR by immunohistochemical analysis in breast cancer cell lines.
Breast Cancer (Auckl). 4:35–41. 2010.PubMed/NCBI
|
|
3
|
Toss A and Cristofanilli M: Molecular
characterization and targeted therapeutic approaches in breast
cancer. Breast Cancer Res. 17:602015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Schlotter CM, Vogt U, Allgayer H and
Brandt B: Molecular targeted therapies for breast cancer treatment.
Breast Cancer Res. 10:2112008. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Guo W, Chen W, Yu W, Huang W and Deng W:
Small interfering RNA-based molecular therapy of cancers. Chin J
Cancer. 32:488–493. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kim DH and Rossi JJ: Strategies for
silencing human disease using RNA interference. Nat Rev Genet.
8:173–184. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
7
|
Karagiannis TC and El-Osta A: RNA
interference and potential therapeutic applications of short
interfering RNAs. Cancer Gene Ther. 12:787–795. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hristodorov D, Mladenov R, von Felbert V,
Huhn M, Fischer R, Barth S and Thepen T: Targeting CD64 mediates
elimination of M1 but not M2 macrophages in vitro and in
cutaneous inflammation in mice and patient biopsies. MAbs.
7:853–862. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Maire V, Némati F, Richardson M,
Vincent-Salomon A, Tesson B, Rigaill G, Gravier E, Marty-Prouvost
B, De Koning L, Lang G, et al: Polo-like kinase 1: A potential
therapeutic option in combination with conventional chemotherapy
for the management of patients with triple-negative breast cancer.
Cancer Res. 73:813–823. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
King SI, Purdie CA, Bray SE, Quinlan PR,
Jordan LB, Thompson AM and Meek DW: Immunohistochemical detection
of Polo-like kinase-1 (PLK1) in primary breast cancer is associated
with TP53 mutation and poor clinical outcom. Breast Cancer Res.
14:R402012. View
Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lens SM, Voest EE and Medema RH: Shared
and separate functions of polo-like kinases and aurora kinases in
cancer. Nat Rev Cancer. 10:825–841. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Keever LB, Jones JE and Andresen BT: G
protein-coupled receptor kinase 4gamma interacts with inactive
Galpha(s) and Galpha13. Biochem Biophys Res Commun. 367:649–655.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Strub GM, Maceyka M, Hait NC, Milstien S
and Spiegel S: Extracellular and intracellular actions of
sphingosine-1-phosphate. Adv Exp Med Biol. 688:141–155. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Dominska M and Dykxhoorn DM: Breaking down
the barriers: siRNA delivery and endosome escape. J Cell Sci.
123:1183–1189. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Bartlett DW, Su H, Hildebrandt IJ, Weber
WA and Davis ME: Impact of tumor-specific targeting on the
biodistribution and efficacy of siRNA nanoparticles measured by
multimodality in vivo imaging. Proc Natl Acad Sci USA.
104:15549–15554. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lakhin AV, Tarantul VZ and Gening LV:
Aptamers: Problems, solutions and prospects. Acta Naturae. 5:34–43.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Krause DS and Van Etten RA: Tyrosine
kinases as targets for cancer therapy. N Engl J Med. 353:172–187.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Chen CH, Chernis GA, Hoang VQ and Landgraf
R: Inhibition of heregulin signaling by an aptamer that
preferentially binds to the oligomeric form of human epidermal
growth factor receptor-3. Proc Natl Acad Sci USA. 100:9226–9231.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Carraway KL III, Sliwkowski MX, Akita R,
Platko JV, Guy PM, Nuijens A, Diamonti AJ, Vandlen RL, Cantley LC
and Cerione RA: The erbB3 gene product is a receptor for heregulin.
J Biol Chem. 269:14303–14306. 1994.PubMed/NCBI
|
|
20
|
Sliwkowski MX, Schaefer G, Akita RW,
Lofgren JA, Fitzpatrick VD, Nuijens A, Fendly BM, Cerione RA,
Vandlen RL and Carraway KL III: Coexpression of erbB2 and erbB3
proteins reconstitutes a high affinity receptor for heregulin. J
Biol Chem. 269:14661–14665. 1994.PubMed/NCBI
|
|
21
|
Kim S, Han J, Shin I, Kil WH, Lee JE and
Nam SJ: A functional comparison between the HER2(high)/HER3 and the
HER2(low)/HER3 dimers on heregulin-β1-induced MMP-1 and MMP-9
expression in breast cancer cells. Exp Mol Med. 44:473–482. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Salameh A, Fan X, Choi BK, Zhang S, Zhang
N and An Z: HER3 and LINC00052 interplay promotes tumor growth in
breast cancer. Oncotarget. 8:6526–6539. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Suga J, Izumiyama K, Tanaka N and Saji S:
Estradiol promotes rapid degradation of HER3 in ER-positive breast
cancer cell line MCF-7. Biochem Biophys Rep. 16:103–109.
2018.PubMed/NCBI
|
|
24
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Rydzanicz R, Zhao XS and Johnson PE:
Assembly PCR oligo maker: A tool for designing
oligodeoxynucleotides for constructing long DNA molecules for RNA
production. Nucleic Acids Res. 33(web server issue): W521–525.
2005.https://doi.org/10.1093/nar/gki380 View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Fire A, Xu S, Montgomery MK, Kostas SA,
Driver SE and Mello CC: Potent and specific genetic interference by
double-stranded RNA in Caenorhabditis elegans. Nature. 391:806–811.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Whitehead KA, Langer R and Anderson DG:
Knocking down barriers: Advances in siRNA delivery. Nat Rev Drug
Discov. 8:129–138. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Hussain AF, Wullner U, Neef I, Tur MK and
Barth S.: Targeted delivery of short interfering RNAs - Strategies
for in vivo application. Topics in Anti-Cancer-Research.
Bentham Science Publishers. (Sharjah, U.A.E.). 228–253. 2013.
|
|
29
|
Martelotto LG, Ng CK, Piscuoglio S,
Weigelt B and Reis-Filho JS: Breast cancer intra-tumor
heterogeneity. Breast Cancer Res. 16:2102014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Bu Y, Yang Z, Li Q and Song F: Silencing
of polo-like kinase (Plk) 1 via siRNA causes inhibition of growth
and induction of apoptosis in human esophageal cancer cells.
Oncology. 74:198–206. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Dassie JP, Liu XY, Thomas GS, Whitaker RM,
Thiel KW, Stockdale KR, Meyerholz DK, McCaffrey AP, McNamara JO II
and Giangrande PH: Systemic administration of optimized
aptamer-siRNA chimeras promotes regression of PSMA-expressing
tumors. Nat Biotechnol. 27:839–849. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
McNamara JO II, Andrechek ER, Wang Y,
Viles KD, Rempel RE, Gilboa E, Sullenger BA and Giangrande PH: Cell
type-specific delivery of siRNAs with aptamer-siRNA chimeras. Nat
Biotechnol. 24:1005–1015. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wullner U, Neef I, Eller A, Kleines M, Tur
MK and Barth S: Cell-specific induction of apoptosis by rationally
designed bivalent aptamer-siRNA transcripts silencing eukaryotic
elongation factor 2. Curr Cancer Drug Targets. 8:554–565. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Machuy N, Thiede B, Rajalingam K, Dimmler
C, Thieck O, Meyer TF and Rudel T: A global approach combining
proteome analysis and phenotypic screening with RNA interference
yields novel apoptosis regulators. Mol Cell Proteomics. 4:44–55.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zhang Y, Wang Y, Wan Z, Liu S, Cao Y and
Zeng Z: Sphingosine kinase 1 and cancer: A systematic review and
meta-analysis. PLoS One. 9:e903622014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Xu Y, Dong B, Huang J, Kong W, Xue W, Zhu
Y, Zhang J and Huang Y: Sphingosine kinase 1 is overexpressed and
promotes adrenocortical carcinoma progression. Oncotarget.
7:3233–3244. 2016.PubMed/NCBI
|
|
37
|
Hussain AF, Tur MK and Barth S: An
aptamer-siRNA chimera silences the eukaryotic elongation factor 2
gene and induces apoptosis in cancers expressing αvβ3 integrin.
Nucleic Acid Ther. 23:203–212. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Li TS, Yawata T and Honke K: Efficient
siRNA delivery and tumor accumulation mediated by ionically
cross-linked folic acid-poly(ethylene glycol)-chitosan
oligosaccharide lactate nanoparticles: For the potential targeted
ovarian cancer gene therapy. Eur J Pharm Sci. 52:48–61. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Song E, Zhu P, Lee SK, Chowdhury D,
Kussman S, Dykxhoorn DM, Feng Y, Palliser D, Weiner DB, Shankar P,
et al: Antibody mediated in vivo delivery of small
interfering RNAs via cell-surface receptors. Nat Biotechnol.
23:709–717. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Reschke M, Mihic-Probst D, van der Horst
EH, Knyazev P, Wild PJ, Hutterer M, Meyer S, Dummer R, Moch H and
Ullrich A: HER3 is a determinant for poor prognosis in melanoma.
Clin Cancer Res. 14:5188–5197. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Servidei T, Riccardi A, Mozzetti S,
Ferlini C and Riccardi R: Chemoresistant tumor cell lines display
altered epidermal growth factor receptor and HER3 signaling and
enhanced sensitivity to gefitinib. Int J Cancer. 123:2939–2949.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Lee-Hoeflich ST, Crocker L, Yao E, Pham T,
Munroe X, Hoeflich KP, Sliwkowski MX and Stern HM: A central role
for HER3 in HER2-amplified breast cancer: Implications for targeted
therapy. Cancer Res. 68:5878–5887. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Jeschke M, Wels W, Dengler W, Imber R,
Stöcklin E and Groner B: Targeted inhibition of tumor-cell growth
by recombinant heregulin-toxin fusion proteins. Int J Cancer.
60:730–739. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Yu X, Ghamande S, Liu H, Xue L, Zhao S,
Tan W, Zhao L, Tang SC, Wu D, Korkaya H, et al: Targeting
EGFR/HER2/HER3 with a three-in-one aptamer-siRNA chimera confers
superior activity against HER2+ breast cancer. Mol Ther
Nucleic. 10:317–330. 2018. View Article : Google Scholar
|