Introduction
Osteosarcoma is the most common type of primary
malignant tumor in the bone tissue (1). Whilst most younger patients (10–30
years old) suffer from primary conventional osteosarcoma, elderly
patients (>65 years) are more likely have secondary osteosarcoma
(2). Osteosarcoma mainly originates
from the long bones, but other bones may also be affected by the
disease (3). It is estimated that
between 0.8 and 11 individuals per 100,000 are affected by this
disease (4), with 1,000 new cases of
osteosarcoma reported in the United States every year (2). Despite the low incidence rate,
osteosarcoma is considered to be an important cause of
cancer-related mortality (3).
Patients with osteosarcoma are usually treated with aggressive
adjuvant chemotherapy and surgical resection (3,4). With
the development of novel treatment strategies, treatment outcomes
of osteosarcoma have improved significantly during the past decades
(1). However, the prognosis for
osteosarcoma patients remains fairly poor, particularly for
patients with metastatic or recurrent osteosarcoma, for whom the
5-year survival rate is <25% (3).
The development of osteosarcoma is a complex process
that involves a number of internal and external factors. High
levels of genomic instability are common in patients with
osteosarcoma, and activation of oncogenes and mutations of tumor
suppressor genes have been found to be at least partially
responsible for the occurrence of this disease (1). The role of noncoding RNAs has been
extensively studied in a variety of pathophysiological processes
(5). Long non-coding RNA (lncRNA) is
a group of noncoding RNAs that are >200 nucleotides in length
(6), and significantly longer than
miRNA, siRNA and other short noncoding RNAs. One such lncRNA,
taurine-upregulated gene 1 (TUG1), has been demonstrated to serve
different functions in different types of malignancies (7,8). In
glioma, TUG1 acts as a tumor suppressor gene by promoting cancer
cell apoptosis (7), whereas it has
been reported to be upregulated in esophageal squamous cell
carcinoma, where it promotes cancer cell proliferation and
migration (8). In addition, TUG1 has
been reported to be upregulated in osteosarcoma, with this
upregulation being closely correlated with poor prognosis of
osteosarcoma patients (9). However,
the functionality of this remains unclear.
In the present study, the expression of TUG1 in the
tumor tissues and adjacent healthy tissues of osteosarcoma
patients, and in plasma samples from osteosarcoma patients and
healthy controls was analyzed. The diagnostic and prognostic values
of TUG1 for osteosarcoma were analyzed. In addition, the effects of
TUG1 overexpression and silencing on osteosarcoma cell viability,
migration and invasion were tested. The effects of TUG1
overexpression on proteins associated with the
epithelial-mesenchymal transition (EMT) pathway were also
explored.
Materials and methods
Patients
A total of 40 patients with osteosarcoma (22 males
and 18 females; age range, 11–78 years; average age, 41±11.1 years)
were selected in the 2nd Affiliated Hospital of Zhejiang University
School of Medicine (Zhejiang, China) from January 2009 to January
2011. Inclusion criteria: i) All patients were diagnosed by
pathological and imaging examination; ii) newly diagnosed cases.
Exclusion criteria: i) Recurrent patients; ii) patients who have
been treated; iii) patients with multiple newly diagnosed clinical
disorders. Distant metastasis was detected in 26 patients. All
patients received surgical resections, with tumor tissues and
adjacent healthy tissues within the region 0.5 cm from the tumor
boundary being collected during surgery. Within the same timeframe,
40 healthy individuals (20 males and 20 females; age range, 16–72
years; average age, 40±9.8 years) were selected as controls. Blood
(5 ml) was extracted from each patient and control before therapies
under fasting conditions. Blood was centrifuged in EDTA tubes at
room temperature for 10 min at 1,200 × g to obtain plasma. There
were no significant differences in age, sex and other basic
information between the two groups. This study was approved by the
ethics committee of The 2nd Affiliated Hospital of Zhejiang
University School of Medicine, and all patients provided signed
informed consent. Follow-up was performed for 60 months to monitor
the survival of the patients.
Cell lines and cell culture
Human normal bone cell line hFOB and osteosarcoma
cell lines MG-63 and U2OS were purchased from American Type Culture
Collection (ATCC). According to the supplier's protocol, hFOB cells
were cultivated in a mixture of 45% Dulbecco's modified Eagle's
medium, 45% Ham's F12 Medium and 10% fetal bovine serum (all
Sigma-Aldrich; Merck KGaA), U2OS cells were cultured using
ATCC-formulated McCoy's 5A Medium (cat no. 30-2007; ATCC)
containing 10% fetal bovine serum, whilst MG-63 cells were cultured
with Eagle's Minimum Essential Medium (cat no. 30-2003; ATCC)
containing 10% heat-inactivated fetal bovine serum (Sigma-Aldrich;
Merck KGaA) maintained in a humidified atmosphere at 37°C under 5%
CO2. Cells were harvested upon reaching the logarithmic
growth phase for subsequent experiments.
Cell transfection
Silencer™ Select Negative Control No. 1 siRNA
(5′-UUCGAGAGAUGCACGGAAAU-3′; cat. no. 4390843) and TUG1 siRNA
(5′-GGGAUAUAGCCAGAGAACAAUUCU-3′; cat. no. 1299001; both Thermo
Fisher Scientific, Inc.) were used to establish TUG1-silenced cell
lines. A TUG1 overexpression vector was established using
pIRSE2-EGFP vector backbone and empty pIRSE2-EGFP vector backbone
vectors were also used (Clontech Laboratories, Inc.). The vector
construction service was provided by Sangon Biotech Co., Ltd. Cells
(106 in 10 ml media; a different medium for different
cell lines) were cultured overnight to reach 80–90% confluence, and
transfection (40 nM siRNA or 10 nM vector) was performed using
Lipofectamine® 2000 reagent according to the
manufacturer's protocol (Invitrogen; Thermo Fisher Scientific,
Inc.). Following experiments were performed using cells harvested
at 24 h post-transfection.
Reverse transcription-quantitative PCR
(RT-qPCR)
TRIzol reagent (Invitrogen; Thermo Fisher
Scientific, Inc.) was used to extract total RNA from tissues,
plasma samples and cell lines according to the manufacturer's
protocol. RNA quality was assessed using the NanoDrop™ 2000
Spectrophotometer (Thermo Fisher Scientific, Inc.) to determine the
absorbance values at 260 nm (A260) and A280. Only RNA samples with
A260/A280 ratios of between 1.8 and 2.0 were used for RT to
synthesize cDNA. RT was carried out using RevertAid RT Reverse
Transcription kit (Thermo Fisher Scientific, Inc.) with the
following conditions: 55°C for 30 min and 80°C for 10 min. qPCR
systems were prepared using SYBR® Green Real-Time PCR
Master mix according to the manufacturer's protocol (Thermo Fisher
Scientific, Inc.). The following primers were used for the qPCR
step: TUG1 forward, 5′-TTGTCACGTCCACCGGACCTG-3′ and reverse,
5′-CACAAATTCCCATCATTCCC-3′; runt-related transcription factor 2
(RUNX2) forward, 5′-CCTGAACTCTGCACCAAGTC-3′ and reverse,
5′-GAGGTGGCAGTGTCATCATC-3′; and β-actin forward,
5′-GACCTCTATGCCAACACAGT-3′ and reverse, 5′-AGTACTTGCGCTCAGGAGGA-3′.
CFX96 Touch™ Real-Time PCR Detection System (Bio-Rad Laboratories,
Inc.) was used to perform qPCR. The thermocycling conditions for
qPCR were: Initial denaturation at 95°C for 30 sec, followed by 40
cycles of 95°C for 12 sec and 60°C for 30 sec. Data were processed
using the 2−ΔΔCq method (10). Relative expression levels of TUG1
were normalized to those of the endogenous control β-actin.
Cell viability assay
Cells were seeded into 96-well plates at
6×103 cells/well. After incubation for 3–5 h to allow
cell adhesion, 100 µl aforementioned media was added. A total of 10
µl Cell Counting Kit-8 (CCK-8; Sigma-Aldrich; Merck KGaA) reagent
was added after 24, 48, 72 and 96 h of incubation. Following
incubation for a further 3 h, optical density values at 450 nm were
measured for each well using a microplate reader.
Cell migration and invasion assay
Transwell® cell migration assay (BD
Biosciences) was performed. Briefly, the upper chamber was seeded
with 3×104 cells diluted in serum-free aforementioned
media, while RPMI-1640 medium (Thermo Fisher Scientific, Inc.)
containing 20% fetal calf serum (Sigma-Aldrich; Merck KGaA) was
used to fill the lower chamber. The membranes of the Transwell
chambers were collected after 24 h incubation, membranes were fixed
in 70% ethanol for 1 min at room temperature, followed by staining
with 0.5% crystal violet (Sigma-Aldrich; Merck KGaA) for 20 min at
room temperature. Stained cells were counted under a light
microscope. Five visual fields were selected to count cells. In
cell invasion assays, the upper chamber was first pre-coated with
200 mg/nl Matrigel® (cat. no. 356234; EMD Millipore;
Merck KGaA) for 12 h at 37°C, and all other procedures were
identical to those of the migration assays. Cell migration and
invasion rates were normalized to cell viability rates at 24 h
using the CCK-8 assay.
Western blot analysis
Total protein extraction was performed using RIPA
solution (Thermo Fisher Scientific, Inc.), and protein
concentration was assessed using a bicinchoninic acid assay. A
total of 30 µg protein from each sample was separated by 10%
SDS-PAGE and then transferred to PVDF membranes. Blocking was
performed by incubation with 5% skimmed milk in PBS at room
temperature for 2 h. Following washing, membranes were incubated
with rabbit polyclonal anti-RUNX2 (1:1,000; cat. no. ab23981) or
rabbit polyclonal anti-GAPDH (1:1,000; cat. no. ab9485) primary
antibodies (all Abcam) overnight at 4°C. Membranes were washed and
incubated with goat anti-rabbit IgG secondary antibody conjugated
to horseradish peroxidase (1:1,000; cat. no. MBS435036;
MyBioSource, Inc.) at room temperature for 3 h. Following another
round of washing, ECL (Sigma-Aldrich; Merck KGaA) was added to
visualize the protein bands. Signals were then scanned using the
MYECL™ Imager (Thermo Fisher Scientific, Inc.). Relative expression
levels of each protein were normalized to those of the endogenous
control β-actin using Image J v1.48 software (National Institutes
of Health).
Statistical analysis
SPSS 19.0 software (IBM Corp.) was used for all
statistical analyses. Normally distributed data are presented as
the mean ± standard deviation. Comparisons between two groups were
performed using unpaired t-test, comparisons between tumor and
healthy tissues were performed using paired test, and comparisons
among multiple groups were performed using one-way ANOVA followed
by Tukey's test. Patients with osteosarcoma were divided into two
groups according to the median expression level of TUG1 mRNA in
plasma (3.98). Survival curves were plotted using the Kaplan-Meier
method and compared using the log-rank test. Non-normally
distributed data were analyzed using the Mann-Whitney U test. The
associations between clinicopathological features and TUG1
expression levels in plasma were analyzed using Chi-square test.
Receiver operating characteristic (ROC) curve analysis was
performed to analyze the diagnostic value of TUG1 expression for
osteosarcoma. P<0.05 was considered to indicate a statistically
significant difference.
Results
Relative expression levels of lncRNA
TUG1 in osteosarcoma and adjacent healthy tissues
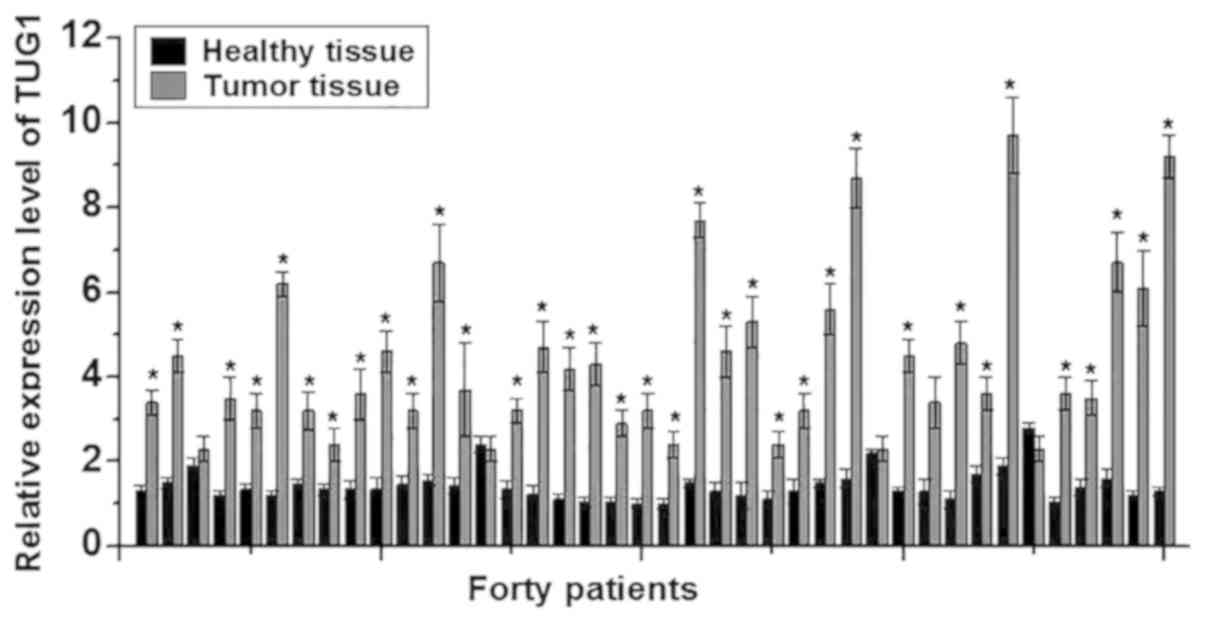
In this study, RT-qPCR was performed to detect the
expression of TUG1 lncRNA in the cancer tissues and adjacent
healthy tissues of 40 patients with osteosarcoma. The expression
levels of TUG1 were significantly higher in cancer tissues compared
with adjacent healthy tissues in 37/40 patients (P<0.05;
Fig. 1), suggesting that lncRNA TUG1
is likely to be involved in the development of osteosarcoma.
Relative expression levels of lncRNA
TUG1 in the plasma of osteosarcoma patients and healthy
controls
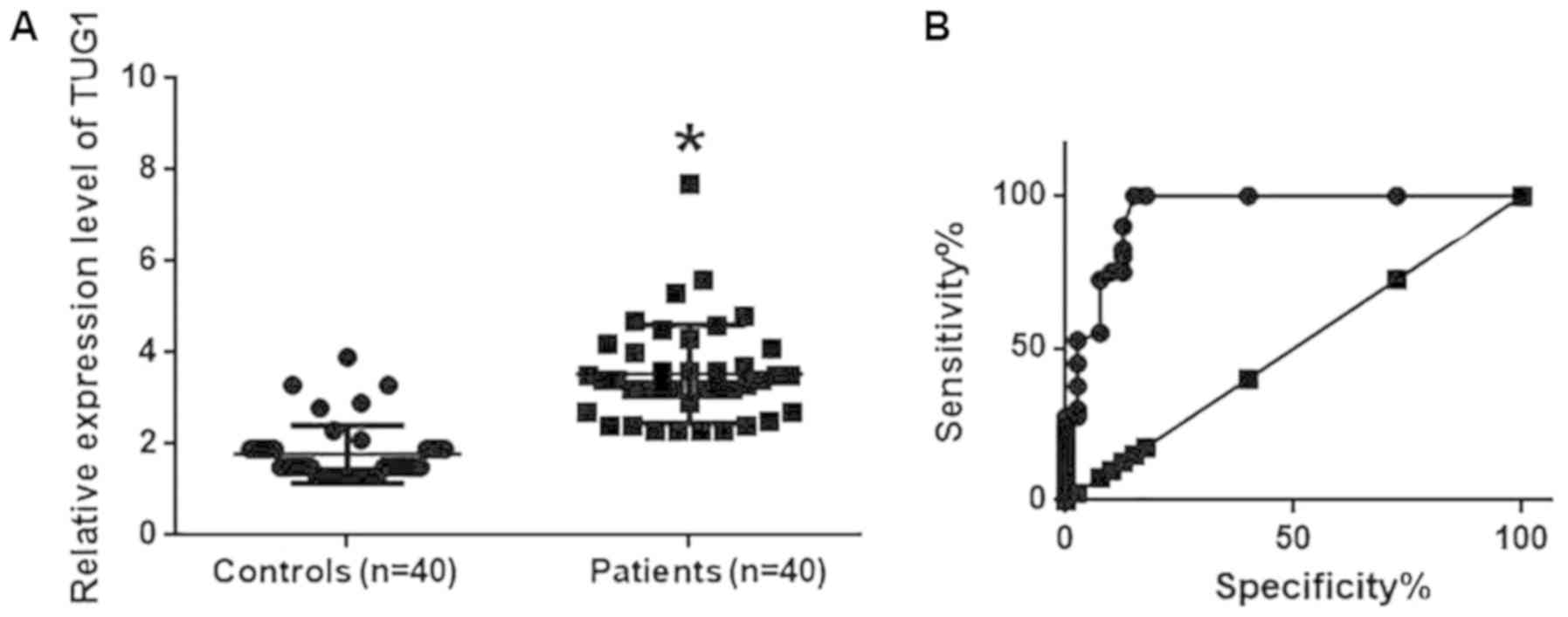
Expression levels of TUG1 in the plasma samples of
40 osteosarcoma patients and 40 healthy controls were also detected
using RT-qPCR. TUG1 levels in the plasma of osteosarcoma patients
were significantly higher compared with those in healthy controls
(P<0.05; Fig. 2A). ROC curve
analysis was also performed to analyze the diagnostic value of TUG1
expression for osteosarcoma. The area under the curve was
calculated to be 0.9447 with a 95% confidence interval of
0.8943–0.9960 (P<0.0001; Fig.
2B), suggesting that TUG1 expression can be used to diagnose
osteosarcoma effectively.
Factors affecting TUG1 expression and
prognostic value of TUG1 expression for osteosarcoma
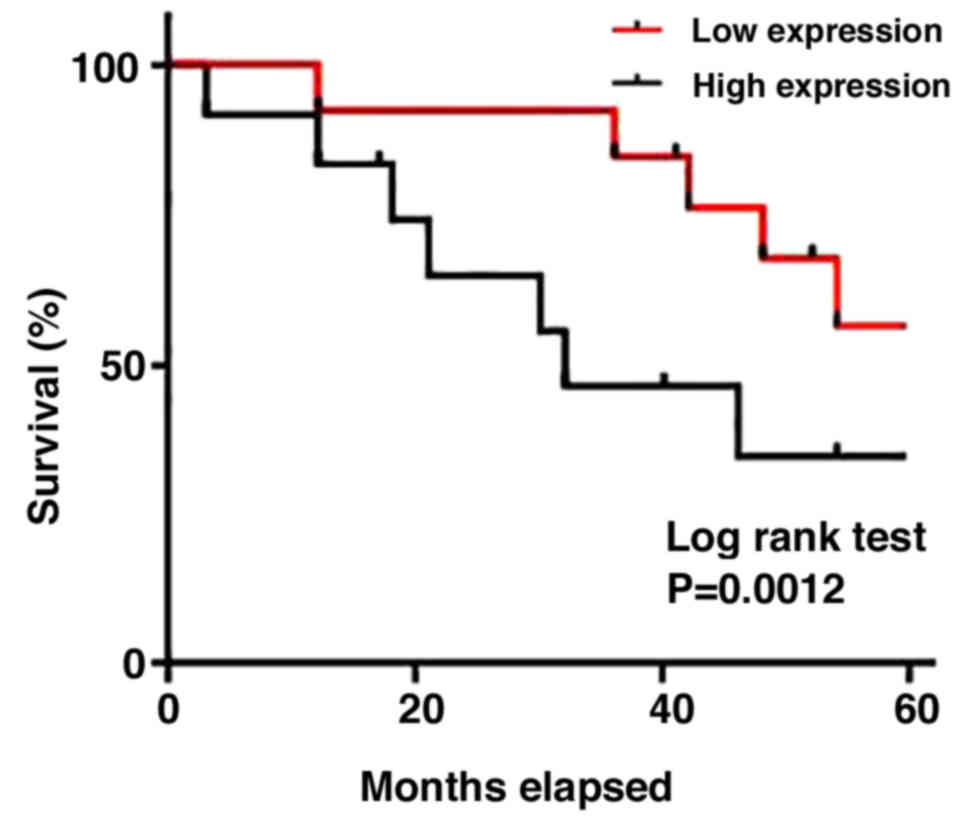
Patients with osteosarcoma were divided into two
groups according to the median expression levels of TUG1 in plasma.
LncRNA expression has been found to be dependent on lifestyle
factors, including smoking and drinking (11). In the present study, the expression
level of TUG1 was not associated with sex, age, or history of
drinking or smoking, but was associated significantly with tumor
metastasis (P<0.001; Table I),
suggesting that TUG1 is a valid diagnostic marker for osteosarcoma.
The Kaplan-Meier method was next applied to produce a survival
curve. Overall patient survival in the group with high TUG1
expression was found to be significantly lower compared with that
in patients with low TUG1 expression (P<0.005; Fig. 3). These results suggest that TUG1 can
serve as a prognostic marker for osteosarcoma.
 | Table I.Association between
clinicopathological features and TUG1 expression levels in
plasma. |
Table I.
Association between
clinicopathological features and TUG1 expression levels in
plasma.
|
|
| TUG1 expression |
|
|---|
|
|
|
|
|
|---|
| Clinicopathological
features | N | High | Low | P-value |
|---|
| Sex |
|
|
| 0.53 |
| Male | 22 | 12 | 10 |
|
|
Female | 18 | 8 | 10 |
|
| Age (years) |
|
|
| 0.11 |
|
>40 | 21 | 13 | 8 |
|
| ≤40 | 19 | 7 | 12 |
|
| Drinking |
|
|
| 0.17 |
| Yes | 28 | 16 | 12 |
|
| No | 12 | 4 | 8 |
|
| Smoking |
|
|
| 1.00 |
| Yes | 24 | 12 | 12 |
|
| No | 16 | 8 | 8 |
|
| Metastasis |
|
|
| <0.001 |
| Yes | 26 | 19 | 7 |
|
| No | 14 | 1 | 13 |
|
Effects of TUG1 knockdown by siRNA and
vector-induced overexpression on osteosarcoma cell viability
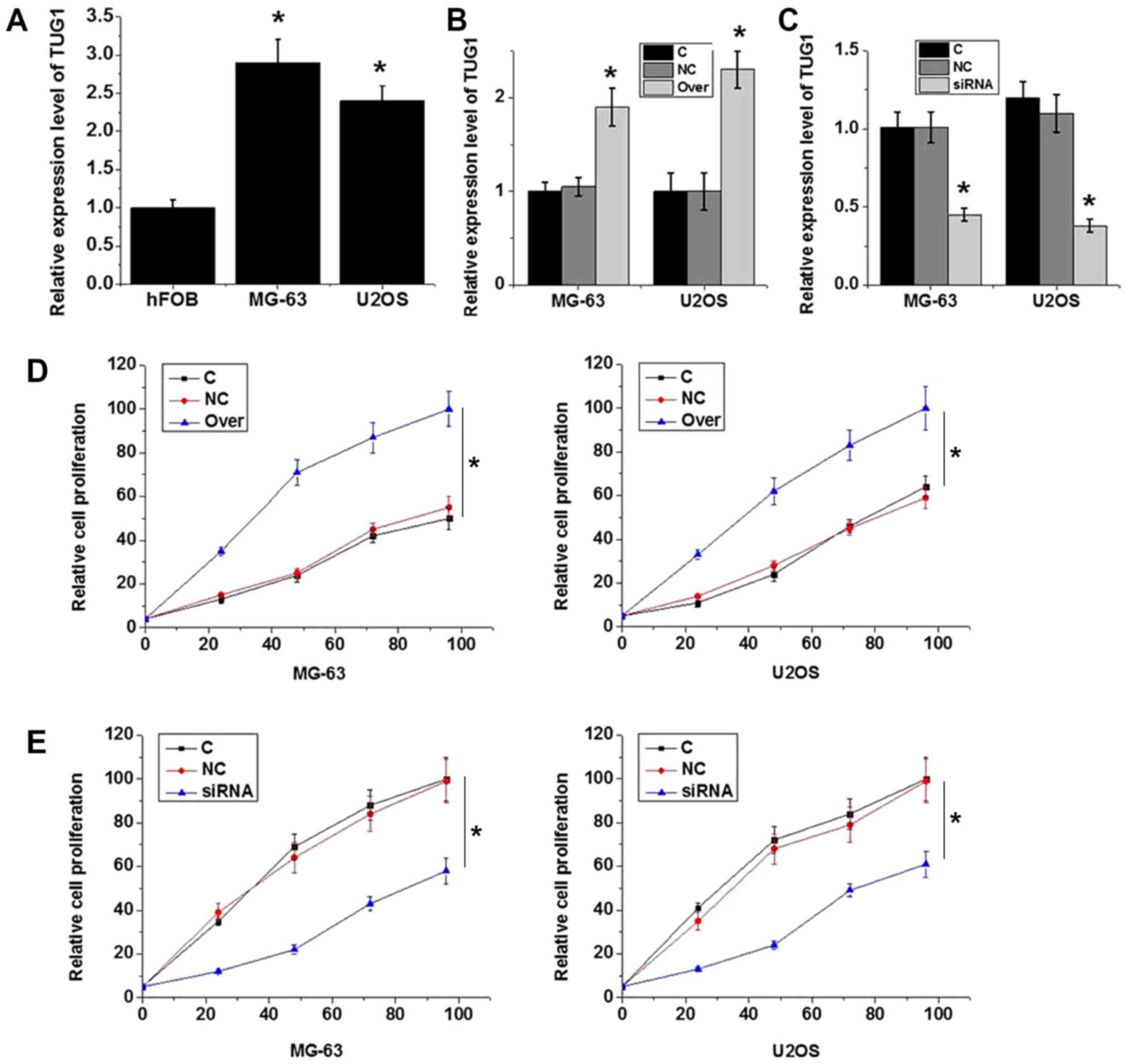
The expression levels of TUG1 were found to be
significantly lower in the hFOB human normal bone cell line
compared with those in the MG-63 and U2OS osteosarcoma cell lines
(P<0.05; Fig. 4A). According to
RT-qPCR results, transfection with plasmid expressing TUG1
significantly increased TUG1 expression (P<0.05; Fig. 4B), whereas transfection with TUG1
siRNA significantly reduced TUG1 expression in MG-63 and U2OS cells
(P<0.05; Fig. 4C), demonstrating
that the transfection was efficient. TUG1 overexpression
significantly increased MG-63 and U2OS cell viability (P<0.05;
Fig. 4D), while TUG1 knockdown
significantly reduced cell viability (P<0.05; Fig. 4E). These data suggest that the
expression level of TUG1 is positively associated with viability in
osteosarcoma cells.
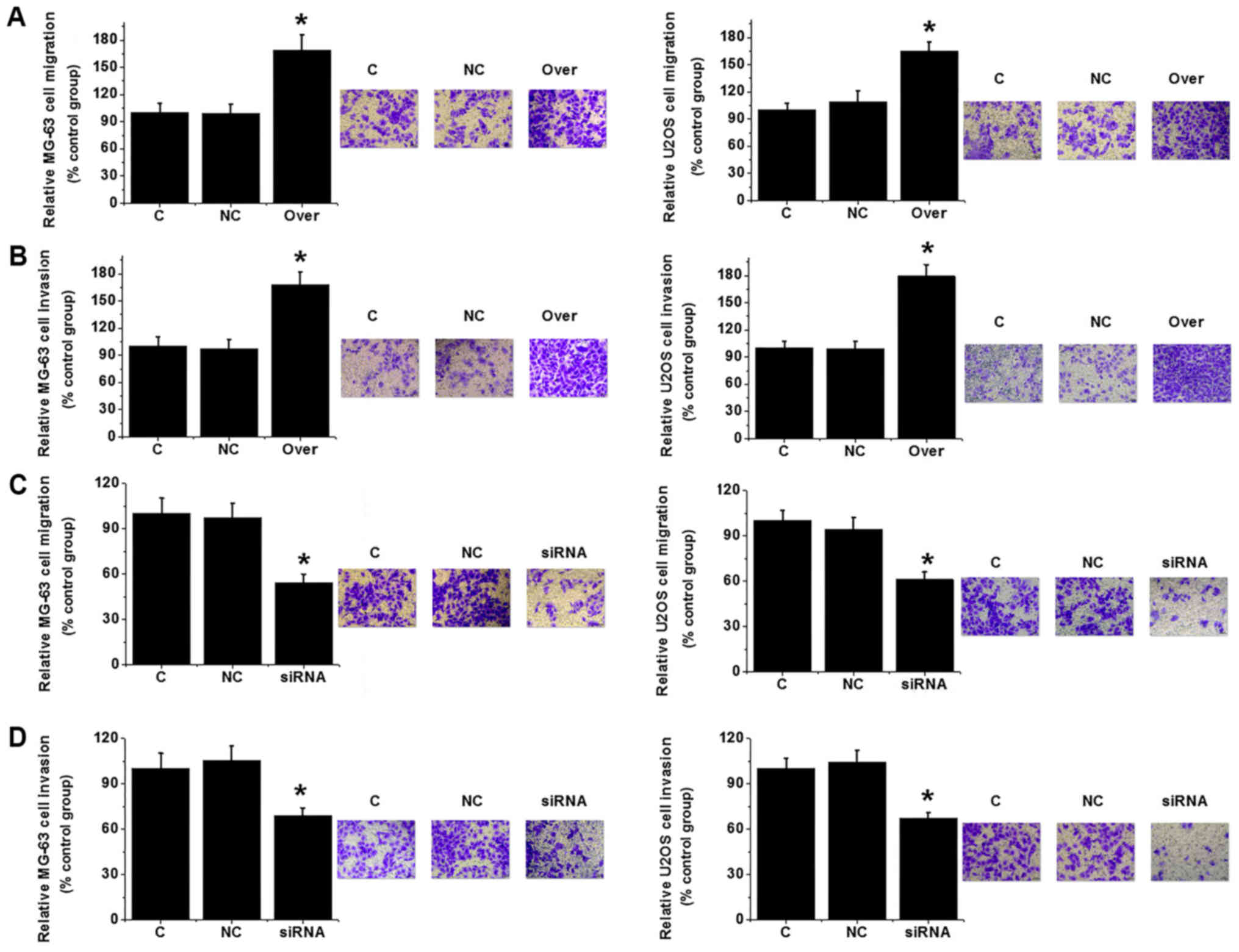
Effects of TUG1 silencing and
overexpression on osteosarcoma cell migration and invasion
Transwell migration and invasion assays were
performed to investigate the effects of changes in TUG1 expression
on cell migration and invasion. TUG1 overexpression significantly
increased MG-63 and U2OS cell migration and invasion (P<0.05;
Fig. 5A and B). By contrast, TUG1
knockdown significantly reduced the migratory and invasive
capabilities of the same cell lines (P<0.05; Fig. 5C and D). Those observations suggest
that expression level of TUG1 is positively associated with the
migration and invasion abilities of osteosarcoma cells.
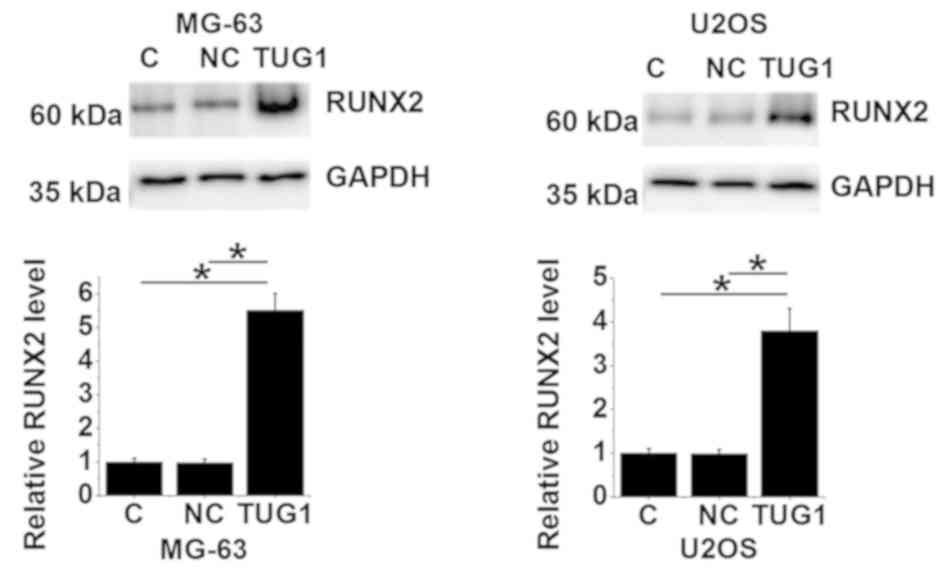
TUG1 overexpression upregulates RUNX2
expression in osteosarcoma cells
The RUNX2 protein expression levels of MG-63 and
U2OS cells overexpressing TUG1 were evaluated using western blot
analysis. Compared with un-transfected control and negative control
cells transfected with empty vectors, lncRNA TUG1 overexpression
resulted in significantly upregulated RUNX2 expression in both cell
lines (P<0.05; Fig. 6).
Discussion
Osteosarcoma is a rare but devastating disease.
Although genetic factors, including tumor protein P53 and
retinoblastoma tumor suppressor gene, have been demonstrated to be
involved in the development of osteosarcoma, the pathogenesis of
this disease remains poorly characterized (12,13).
LncRNA is a group of functional RNAs that do not encode proteins.
Despite the lack of protein-coding ability, lncRNAs have been shown
serve a role in almost every aspect of critical biological and
pathological processes (14).
Previous studies have found that the development of osteosarcoma is
closely associated with the abnormal expression of a number of
lncRNAs (9,15,16). In
particular, high levels of lncRNA TUG1 have been shown to be
associated with poor prognosis in patients with osteosarcoma
(9). In osteosarcoma cell lines,
lncRNA TUG1 expression is upregulated compared with that in normal
osteoblastic cells, and the overexpression of lncRNA TUG1 promotes
osteosarcoma cell proliferation (15). By contrast, the downregulation of
lncRNA TUG1 has been shown to reduce osteosarcoma cell
proliferation and increase cancer cell apoptosis (16). In addition, an association of the
upregulation of lncRNA TUG1 with the poor survival of osteosarcoma
patients has been reported (9).
Consistent with these findings from previous studies, the present
study revealed that the expression levels of TUG1 were
significantly higher in osteosarcoma tissues compared with adjacent
healthy tissues. TUG1 levels were also demonstrated to be higher in
the plasma samples of osteosarcoma patients compared with those
from healthy controls. Taken together, this suggests that TUG1 may
serve a role as an oncogene in osteosarcoma.
The pathogenesis of certain human malignancies is
frequently accompanied by changes in biological molecules such as
lncRNA in the plasma (17). Notably,
plasma lncRNA has been widely applied in cancer diagnosis; a recent
study reported that the plasma levels of lncRNA SOX2-OT were
significantly increased in patients with osteosarcoma compared with
healthy controls, which indicated poor prognosis (18). Although lncRNA TUG1 has been reported
to be involved in the development of osteosarcoma, its diagnostic
and prognostic potential for osteosarcoma remain unreported. In the
present study, ROC curve analysis showed that increased levels of
TUG1 expression could be used effectively to predict osteosarcoma.
In addition, survival times of patients with higher levels of TUG1
expression were significantly shorter compared with those of
patients with lower expression levels of TUG1, further supporting
the significant diagnostic and prognostic values of TUG1 expression
for osteosarcoma. It is well accepted that the expression of some
lncRNAs can be influenced by lifestyle factors, including drinking
and smoking (11). In the present
study, the levels of TUG1 expression did not associate
significantly with age, sex, or history of drinking or smoking
among patients with osteosarcoma. Instead, TUG1 expression levels
were found to be associated significantly with tumor metastasis.
These data suggest serum TUG1 expression to be a promising
biomarker for osteosarcoma.
TUG1 may participate in the development of a number
of human malignancies by regulating cancer cell proliferation,
migration and invasion (9,15,16).
Zhang et al (19) found that
the downregulation of TUG1 significantly inhibited proliferation,
migration and invasion but promoted apoptosis in renal cell
carcinoma cells, suggesting that TUG1 is a promising target for
this disease. In addition, downregulation of TUG1 was also found to
inhibit proliferation and promote cell apoptosis in osteosarcoma
cells (20). In the present study,
TUG1 overexpression significantly increased osteosarcoma cell
viability, migration and invasion, respectively, while
siRNA-mediated TUG1 knockdown and significantly suppressed them.
EMT is one of the key steps for cancer cell migration and invasion
(21). RUNX2 contributes to the
growth and metastasis of osteosarcoma (22). In the present study, expression
levels of RUNX2 mRNA positively correlated with those of lncRNA
TUG1 in tumor tissues but not in adjacent healthy tissues. LncRNA
TUG1 overexpression led to significantly upregulated RUNX2 in
osteosarcoma cell lines. These findings suggest that TUG1 can
upregulate RUNX2 to promote osteosarcoma cell migration and
invasion.
Due to the limited number of young participants, the
average age of the research subjects in the present study was
41±11.1 years, which does not reflect the general demographics of
this disease and, therefore, serves as a limitation. In future
studies analysis including samples from younger patients is
necessary to confirm the conclusions. In addition, this present
study lacks in vivo experimental data. Experiments using
animal models should also be included in any future studies.
In conclusion, the expression levels of TUG1 were
found to be significantly higher in osteosarcoma tissues compared
with adjacent healthy tissues, and to be significantly higher in
the plasma samples of osteosarcoma patients compared with healthy
controls. This suggests that TUG1 expression has significant
diagnostic and prognostic value for osteosarcoma. Mechanistically,
TUG1 likely promotes osteosarcoma cell proliferation, migration and
invasion by upregulating RUNX2.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
KS designed experiments. KS and YL performed
experiments and analyzed data. KS drafted the paper and both
authors approved the paper.
Ethics approval and consent to
participate
This study was approved by the ethics committee of
The 2nd Affiliated Hospital of Zhejiang University School of
Medicine, and all patients provided signed informed consent.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Durfee RA, Mohammed M and Luu HH: Review
of osteosarcoma and current management. Rheumatol Ther. 3:221–243.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lindsey BA, Markel JE and Kleinerman ES:
Osteosarcoma overview. Rheumatol Ther. 4:25–43. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Lin YH, Jewell BE, Gingold J, Lu L, Zhao
R, Wang LL and Lee DF: Osteosarcoma: Molecular pathogenesis and
iPSC modeling. Trends Mol Med. 23:737–755. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Isakoff MS, Bielack SS, Meltzer P and
Gorlick R: Osteosarcoma: Current treatment and a collaborative
pathway to success. J Clin Oncol. 33:3029–3035. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Esteller M: Non-coding RNAs in human
disease. Nat Rev Genet. 12:861–874. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Perkel JM: Visiting ‘noncodarnia’.
Biotechniques. 54:301, 303–304. 2013. View Article : Google Scholar
|
|
7
|
Li J, Zhang M, An G and Ma Q: LncRNA TUG1
acts as a tumor suppressor in human glioma by promoting cell
apoptosis. Exp Biol Med (Maywood). 241:644–649. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Xu Y, Wang J, Qiu M and Xu L, Li M, Jiang
F, Yin R and Xu L: Upregulation of the long noncoding RNA TUG1
promotes proliferation and migration of esophageal squamous cell
carcinoma. Tumor Biol. 36:1643–1651. 2015. View Article : Google Scholar
|
|
9
|
Ma B, Li M, Zhang L, Huang M, Lei JB, Fu
GH, Liu CX, Lai QW, Chen QQ and Wang YL: Upregulation of long
non-coding RNA TUG1 correlates with poor prognosis and disease
status in osteosarcoma. Tumor Biol. 37:4445–4455. 2016. View Article : Google Scholar
|
|
10
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Thai P, Statt S, Chen CH, Liang E,
Campbell C and Wu R: Characterization of a novel long noncoding
RNA, SCAL1, induced by cigarette smoke and elevated in lung cancer
cell lines. Am J Respir Cell Mol Biol. 49:204–211. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wang L, Zhao Z, Feng W, Ye Z, Dai W, Zhang
C, Peng J and Wu K: Long non-coding RNA TUG1 promotes colorectal
cancer metastasis via EMT pathway. Oncotarget. 7:51713–51719.
2016.PubMed/NCBI
|
|
13
|
Kansara M and Thomas DM: Molecular
pathogenesis of osteosarcoma. DNA Cell Biol. 26:1–18. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Chen X and Yan GY: Novel human
lncRNA-disease association inference based on lncRNA expression
profiles. Bioinformatics. 29:2617–2624. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yun-Bo F, Xiao-Po L, Xiao-Li L, Guo-Long
C, Pei Z and Fa-Ming T: LncRNA TUG1 is upregulated and promotes
cell proliferation in osteosarcoma. Open Med (Wars). 11:163–167.
2016.PubMed/NCBI
|
|
16
|
Zhang Q, Geng PL, Yin P, Wang XL, Jia JP
and Yao J: Down-regulation of long non-coding RNA TUG1 inhibits
osteosarcoma cell proliferation and promotes apoptosis. Asian Pac J
Cancer Prev. 14:2311–2315. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Hori SS, Lutz AM, Paulmurugan R and
Gambhir SS: Correlation of plasma biomarker levels with early-stage
tumor viability in an orthotopic ovarian cancer mouse model. Cancer
Res. 74 (19 Suppl):Abstract nr 873. 2014.PubMed/NCBI
|
|
18
|
Wang Z, Tan M, Chen G, Li Z and Lu X:
LncRNA SOX2-OT is a novel prognostic biomarker for osteosarcoma
patients and regulates osteosarcoma cells proliferation and
motility through modulating SOX2. IUBMB Life. 69:867–876. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhang M, Lu W, Huang Y, Shi J, Wu X, Zhang
X, Jiang R, Cai Z and Wu S: Downregulation of the long noncoding
RNA TUG1 inhibits the proliferation, migration, invasion and
promotes apoptosis of renal cell carcinoma. J Mol Histol.
47:421–428. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhang Q, Geng PL, Yin P, Wang XL, Jia JP
and Yao J: Down-regulation of long non-coding RNA TUG1 inhibits
osteosarcoma cell proliferation and promotes apoptosis. Asian Pac J
Cancer Prev. 14:2311–2315. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Rokavec M, Öner MG, Li H, Jackstadt R,
Jiang L, Lodygin D, Kaller M, Horst D, Ziegler PK, Schwitalla S, et
al: IL-6R/STAT3/miR-34a feedback loop promotes EMT-mediated
colorectal cancer invasion and metastasis. J Clin Invest.
124:1853–1867. 2014. View
Article : Google Scholar : PubMed/NCBI
|
|
22
|
Martin JW, Zielenska M, Stein GS, van
Wijnen AJ and Squire JA: The role of RUNX2 in osteosarcoma
oncogenesis. Sarcoma. 2011:2827452011. View Article : Google Scholar : PubMed/NCBI
|