Introduction
Colorectal cancer is one of the most common
malignant tumours worldwide, and has the third highest incidence
rates among all malignancies (1–3). Despite
significant advances in surgical resection combined with
radiochemotherapy, the prognosis of patients with advanced stage
colorectal cancer remains poor with a five-year survival rate of
<20% (4,5). This is thought to be mainly due to the
rapid growth and widespread metastasis of the cancer. The molecular
mechanisms underlying colorectal cancer growth and metastasis is
currently unclear. Understanding the molecular mechanisms may
facilitate the identification of novel and effective therapeutic
targets (6).
Long non-coding RNAs (lncRNAs) are a class of RNAs
(>200 nucleotides in length) with no protein-coding ability,
that regulate gene expression through lncRNA-mRNA interactions,
lncRNA-microRNA (miR) interactions and lncRNA-protein interactions
(7–9). Recently, an increasing number of
lncRNAs have been reported to serve key roles in various biological
processes, including cell proliferation, colony formation, cell
cycle progression, cell migration, cell invasion and
epithelial-mesenchymal transition (EMT) (10–12). In
addition, deregulation of various lncRNAs has been observed in
several human malignancies (including colorectal cancer) and
lncRNAs have been demonstrated to serve either oncogenic or tumour
suppressive roles (10–13). For instance, expression of the
lncRNA, HOX transcript antisense RNA, is significantly upregulated
in colorectal cancer and regulates the progression and
chemoresistance of colorectal cancer by modulating miR-203a-3p
expression and the activity of the Wnt/β-catenin signalling pathway
(5).
Cervical carcinoma expressed PCNA regulatory lncRNA
(CCEPR) is an lncRNA localized to the 10q21.1 chromosomal region.
CCEPR expression was initially identified to be significantly
upregulated in cervical cancer and is associated with poor patient
prognosis (14). Yang et al
(15) reported that CCEPR promotes
cervical cancer cell proliferation via the upregulation of
proliferating cell nuclear antigen (PCNA) expression. In addition,
CCEPR exerts oncogenic roles in gastric cancer, lung cancer,
bladder cancer and liver cancer (16–19).
Recently, Gaballah et al (20) identified that CCEPR expression is
significantly upregulated in colorectal cancer, and its expression
is positively correlated with the expression of phosphorylated
(p)-ERK1/2, cyclooxygenase (COX)-2, cyclin D1 and PCNA (20). This suggests that CCEPR may be
involved in colorectal cancer progression by modulating the
ERK/COX-2 signalling pathway and cell proliferation activity.
However, the detailed role of CCEPR during colorectal cancer
progression remains currently unclear.
Matrix metalloproteinase (MMP)-2 and MMP-9 are
members of the MMP gene family. They function as zinc-dependent
enzymes that cleave extracellular matrix components (21). It has been demonstrated that MMP-2
and MMP-9 serve crucial roles in tumour cell migration and invasion
(21). EMT is characterized by the
transition of cells from an epithelial-like phenotype to a
mesenchymal phenotype, which causes tumour cells to acquire
invasive and migratory capacities (22–24). In
addition, MMP-2 and MMP-9 have been implicated in the process of
EMT (25). However, whether CCEPR
affects the expression of MMP-2 and MMP-9 and the process of EMT in
colorectal cancer has not yet been explored.
The aims of the present study were to evaluate the
clinical significance of CCEPR expression in colorectal cancer, and
to investigate the function of CCEPR in regulating the malignant
phenotypes of colorectal cancer cells in vitro.
Materials and methods
Clinical tissue samples
The present study was approved by the Ethics
Committee of the Hunan Provincial People's Hospital (Changsha,
China). A total of 58 colorectal cancer tissues and paired adjacent
normal tissues were collected from 58 patients with primary
colorectal cancer admitted to Hunan Provincial People's Hospital
between April 2012 and May 2013. These patients included 38 male
and 20 female, from 33 years old to 75 years old. All patients
provided written informed consent. The inclusion criteria were that
all patients only had primary colorectal cancers. The exclusion
criteria were colorectal cancer patients who had received
chemotherapy or radiotherapy prior to undergoing surgery. The
collected tissues were frozen using liquid nitrogen shortly after
surgical resection, and stored at −80°C until experimentation. The
total duration of follow-up was 5 years.
Cell culture and transfection
Human colorectal cancer cell lines, HT29, Caco2,
SW480 and LS174T, and the normal human intestinal epithelial cell
line, HIEC, were purchased from the American Type Culture
Collection. Cells were cultured in DMEM (Thermo Fisher Scientific,
Inc.) containing 10% FBS (Thermo Fisher Scientific, Inc.) and
maintained in a humidified atmosphere at 37°C and 5%
CO2. The HT29 and Caco-2 cell lines were established
from primary adenocarcinomas of the colon. The SW480 and LS174T
cell lines were established from a Duke's type B adenocarcinoma of
the colon. HT29 and SW480 cells (5×106 cells per well)
were seeded in six-well plates and transfected with 100 nM CCEPR
small interfering (si) RNA (siCCEPR; Shanghai GenePharma Co., Ltd.)
or 100 nM scrambled negative control (NC) siRNA (siNC; Shanghai
GenePharma Co., Ltd.) using Lipofectamine 2000 (Thermo Fisher
Scientific, Inc.) in accordance with the manufacturer's
instructions. The target sequence of the CCEPR siRNA was
5′-CGAGGGCGAGCATGTTTGTTGTTTA-3′ (15). The NC siRNA sequences were
5′-TCAAGUCCACGACGACTTTG-3′. At 48-h post-transfection, the
subsequent experiments were conducted.
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was extracted from the tissue samples or
cells using TRIzol reagent (Thermo Fisher Scientific, Inc.) in
accordance with the manufacturer's instructions. Subsequently, 1 µg
total RNA was used to perform RT-qPCR using a SuperScript III
Platinum SYBR Green One-Step RT-qPCR kit (Thermo Fisher Scientific,
Inc.) and the ABI 7500 real-time PCR system (Thermo Fisher
Scientific, Inc.) in accordance with the manufacturer's
instructions. The thermocycling conditions for reverse
transcription were as follows: 16°C for 30 min, 42°C for 30 min and
85°C for 5 min. The thermocycling conditions for PCR were as
follows: 95°C for 1 min, followed by 40 cycles of 95°C for 15 sec
and 60°C for 30 sec. Relative mRNA expression levels were
calculated using the 2−ΔΔCq method (26) normalized to the internal reference
gene GAPDH. The primer sequences were as follows: CCEPR forward,
5′-AAGGTCCCAGGATACTCGC-3′, and reverse, 5′-GTGTCGTGGACTGGCAAAAT-3′;
GAPDH forward, 5′-ACAACTTTGGTATCGTGGAAGG-3′, and reverse,
5′-GCCATCACGCCACAGTTTC-3′. This assay was repeated 3 times.
Cell proliferation assay
Cell proliferation was examined using a Cell
Counting kit-8 (CCK-8) assay (Thermo Fisher Scientific, Inc.).
Briefly, HT29 and SW480 cells (10,000 cells/well) were seeded in
96-well plates and incubated at 37°C for 0, 24, 48 or 72 h. At the
indicated time points, 10 µl CCK-8 reagent (Thermo Fisher
Scientific, Inc.) was added to each well. The cells were
subsequently incubated at 37°C for 2 h. The optical density at 450
nm was determined using a microplate reader (Bio-Rad Laboratories,
Inc.).
Colony formation assay
Transfected HT29 and SW480 cells were seeded into
six-well plates at a density of 200 cells/well. Following culture
at 37°C for 14 days, the cells were washed with PBS (Thermo Fisher
Scientific, Inc.) twice before they were stained with 0.5% crystal
violet (Thermo Fisher Scientific, Inc.) at room temperature for 10
min. The number of colonies (containing >50 cells) were then
counted under a light microscope (magnification, ×200).
Cell cycle analysis
Flow cytometry was used for cell cycle analysis.
Briefly, transfected HT29 and SW480 cells were first fixed in 75%
ethanol at 4°C overnight, and washed with PBS three times. The
cells were then permeabilized using eBioscience™ Permeabilization
Buffer (Thermo Fisher Scientific, Inc.) at 37°C for 30 min then
stained with 500 µl propidium iodide solution (Thermo Fisher
Scientific, Inc.) at 4°C for 30 min. The cell cycle distribution
was determined using a FACScan flow cytometer (Becton, Dickinson
and Company) and BD Accuri C6 system (32-bit) software version 1.0
(BD Biosciences).
Cell migration assay
A wound healing assay was performed to examine cell
migration. Briefly, the transfected HT29 and SW480 cells were
seeded into 12-well plates and incubated at 37°C without serum to
~90% confluence. Wounds were generated using a 100-µl pipette tip
and the cells were then incubated at 37°C for 24 h. The wounds were
photographed at 0 and 24-h using an inverted microscope
(magnification, ×200; Olympus Corporation) and measured using
ImageJ software version 1.8 (National Institutes of Health).
Cell invasion assay
Cell invasion was examined using a 24-well Transwell
chamber (8 mm pore size; Corning, Inc.) pre-coated with Matrigel
for 1-h at room temperature (EMD Millipore; Merck KGaA).
Transfected HT29 and SW480 cells (50,000 cells/well) were seeded in
300 µl serum-free DMEM to the upper chamber. A total of 500 µl DMEM
containing 20% FBS was added into the bottom chamber. Following
incubation at 37°C for 24 h, cells on the insert were carefully
removed with a cotton-tipped swab. The cells that had invaded
through the membrane were stained with 0.5% crystal violet at room
temperature for 10 min and photographed under an inverted
microscope (magnification, ×200).
Western blot analysis
Transfected HT29 and SW480 cells were lysed using
radioimmunoprecipitation buffer (Beyotime Institute of
Biotechnology) according to the manufacturer's protocols. A
bicinchoninic acid protein assay kit (Pierce; Thermo Fisher
Scientific, Inc.) was then used to determine the protein
concentrations. Protein samples (60 µg/lane) were separated by 12%
SDS-PAGE before they were transferred to polyvinylidene difluoride
membranes (Beyotime Institute of Biotechnology). The membranes were
blocked with 5% non-fat dry milk overnight at 4°C. After washing
with PBS at room temperature three times, the membranes were
incubated with rabbit anti-human E-cadherin (dilution, 1:200; cat.
no. ab40772; Abcam), rabbit anti-human N-cadherin (dilution, 1:500;
cat. no. ab18203; Abcam), rabbit anti-human vimentin (dilution,
1:200, cat. no. ab92547; Abcam), rabbit anti-human MMP-2 (dilution,
1:500, cat. no. ab181286; Abcam), rabbit anti-human MMP-9
(dilution, 1:200, cat. no. ab137867; Abcam), or rabbit anti-human
GAPDH (dilution 1:200, cat. no. ab181602; Abcam) primary antibodies
at room temperature for 4 h. After washing with PBS at room
temperature three times, the membranes were then incubated with a
horseradish peroxidase-conjugated goat anti-rabbit secondary
antibody (dilution, 1:5,000, cat. no. ab6721; Abcam) at room
temperature for 30 min. Chemiluminescence was examined using
SuperSignal™ West Femto Maximum Sensitivity Substrate (Thermo
Fisher Scientific, Inc.) according to the manufacturer's protocol.
ImageJ software (v1.46; National Institutes of Health) was used for
densitometry analysis with GAPDH as the loading control.
Statistical analysis
Data are presented as the mean ± standard deviation
and were analysed using GraphPad Prism 3.0 software (GraphPad
Software, Inc.). Differences between 2 groups were analysed using
paired or unpaired Student's t-tests. Differences between multiple
groups were analysed by one-way analysis of variance followed by a
post hoc Tukey's test. The association between CCEPR expression and
clinicopathological characteristics of colorectal cancer patients
was analysed using chi-square tests. Survival analysis was
performed using Kaplan-Meier survival curves and log-rank tests.
P<0.05 was considered to indicate a statistically significant
difference.
Results
Increased CCEPR expression is
associated with colorectal cancer progression
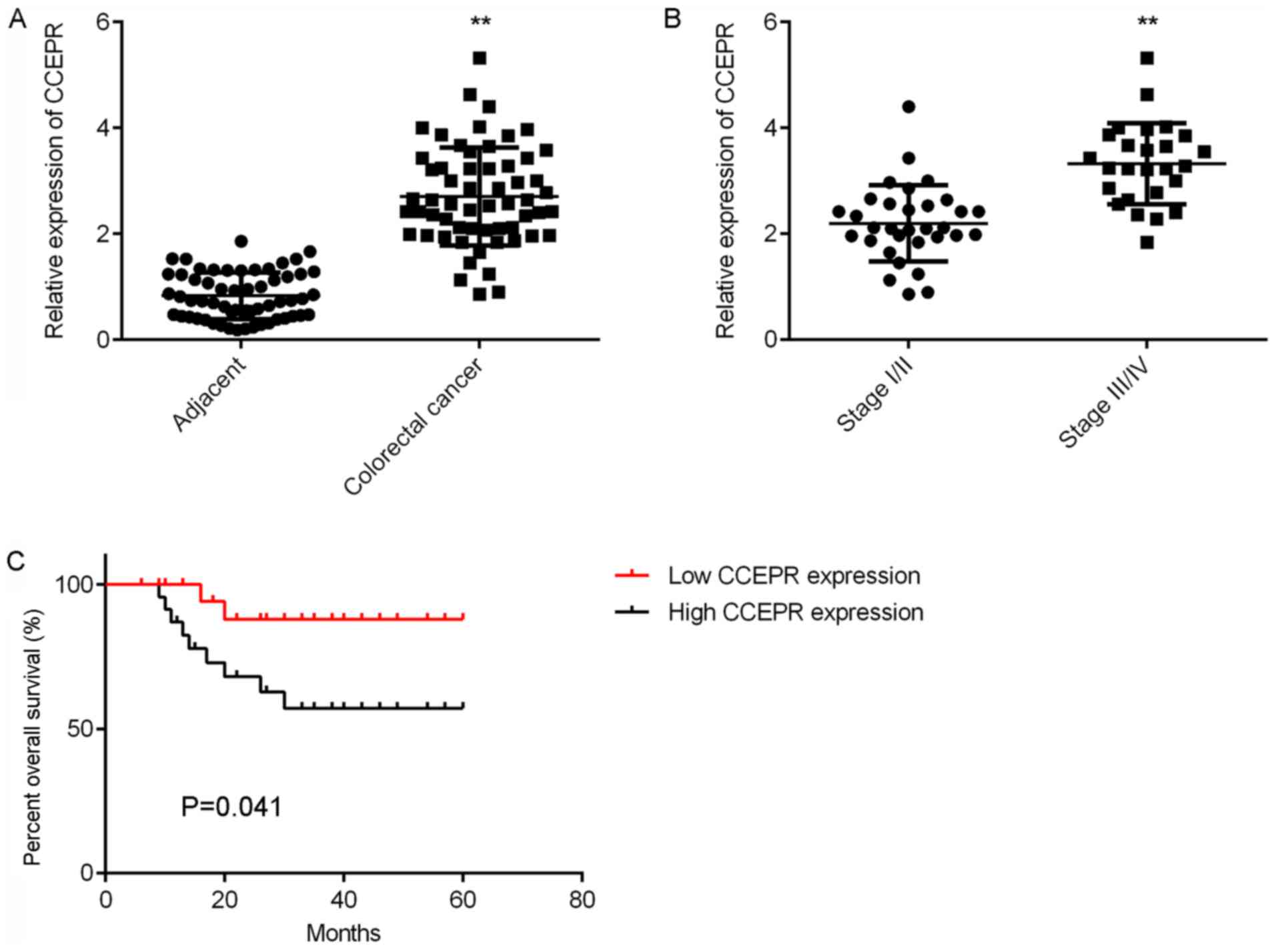
In the present study, the expression of CCEPR in 58
primary colorectal cancer tissues and adjacent normal tissues was
first examined. RT-qPCR results indicated that CCEPR expression was
significantly upregulated in colorectal cancer tissues compared
with adjacent normal tissues (Fig.
1A). In addition, CCEPR expression levels were significantly
higher in advanced colorectal cancer tissues (III/IV) than in
early-stage colorectal cancer tissues (I/II; Fig. 1B). The association between CCEPR
expression and the clinicopathological characteristics of
colorectal cancer patients was then determined. Using the median
expression value of CCEPR as a cut-off value (2.71), patients were
divided into low and high CCEPR expression groups. Statistical
analysis of the results demonstrated that high CCEPR expression was
significantly associated with poor differentiation, positive lymph
node metastasis, distant metastasis and advanced clinical stage
(Table I). This suggests that
upregulation of CCEPR expression may serve a key role during
colorectal cancer progression. Of particular note, patients with
high CCEPR expression levels exhibited shorter survival rates than
patients with low CCEPR expression levels over 5 years (Fig. 1C), suggesting that CCEPR expression
may be a promising predictive marker for colorectal cancer
prognosis.
 | Table I.Association between CCEPR expression
and clinicopathological characteristics of patients with colorectal
cancer. |
Table I.
Association between CCEPR expression
and clinicopathological characteristics of patients with colorectal
cancer.
| Variables | Cases (n=58) | Low CCEPR levels
(n=33) | High CCEPR levels
(n=25) | P-value |
|---|
| Age (years) |
|
|
| 0.384 |
| ≤55 | 31 | 16 | 15 |
|
|
>55 | 27 | 17 | 10 |
|
| Sex |
|
|
| 0.832 |
| Male | 38 | 22 | 16 |
|
|
Female | 20 | 11 | 9 |
|
| Differentiation |
|
|
| 0.032a |
| Well and
moderately | 41 | 27 | 14 |
|
|
Poor | 17 | 6 | 11 |
|
| Node
metastasis |
|
|
| 0.043a |
|
Present | 26 | 11 | 15 |
|
|
Absent | 32 | 22 | 10 |
|
| Distant
metastasis |
|
|
| 0.003a |
|
Present | 9 | 1 | 8 |
|
|
Absent | 49 | 32 | 17 |
|
| Clinical stage |
|
|
| 0.043a |
|
I–II | 32 | 22 | 10 |
|
|
III–IV | 26 | 11 | 15 |
|
Knockdown of CCEPR inhibits the growth
of colorectal cancer cells
In vitro experiments were performed to
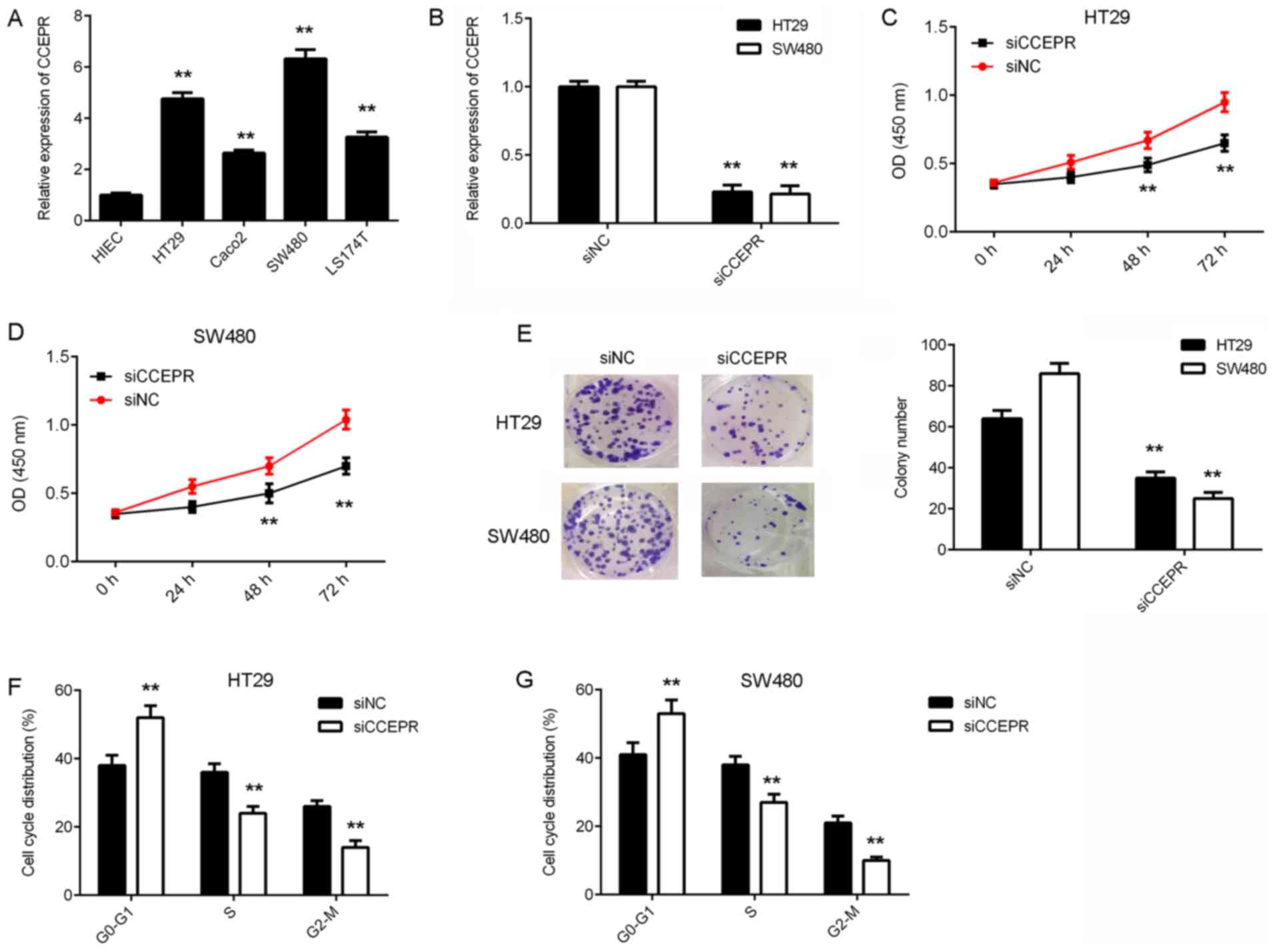
investigate the function of CCEPR in colorectal cancer further. The
expression levels of CCEPR were examined in several human
colorectal cancer cell lines including HT29, Caco-2, SW480 and
LS174T, as well as in the normal human intestinal epithelial cell
line, HIEC. RT-qPCR results demonstrated that CCEPR expression was
significantly increased in colorectal cancer cell lines compared
with HIEC cells (Fig. 2A). HT29 and
SW480 cells exhibited the highest CCEPR expression levels and were
therefore selected for subsequent experiments. As CCEPR expression
was observed to be significantly upregulated in colorectal cancer
samples, CCEPR siRNA was transfected into HT29 and SW480 cells to
reduce its expression levels. Following transfection, CCEPR levels
were significantly decreased in the siCCEPR group compared with the
siNC group (Fig. 2B). A CCK-8 assay
was then performed to assess cell proliferation. The proliferation
of the HT29 and SW480 cells in the siCCEPR group was significantly
suppressed compared with cells in the siNC group (Fig. 2C and D). Thus, CCEPR may serve an
oncogenic role in colorectal cancer growth. A colony formation
assay was then performed to examine the effects of CCEPR
downregulation on the colony formation capacity of colorectal
cancer cells. The results indicated that the colony formation
capacity of cells in the siCCEPR group was significantly inhibited
when compared with cells in the siNC group (Fig. 2E). To further confirm these findings,
flow cytometry was utilised to examine the effects of CCEPR
downregulation on cell cycle progression in colorectal cancer
cells. The results indicated that silencing CCEPR in HT29 and SW480
cells led to significant cell cycle arrest in the G1 stage
(Fig. 2F and G). Therefore, these
results demonstrated that knockdown of CCEPR inhibited the growth
of colorectal cancer cells.
Knockdown of CCEPR suppresses the
migration, invasion and EMT of colorectal cancer cells
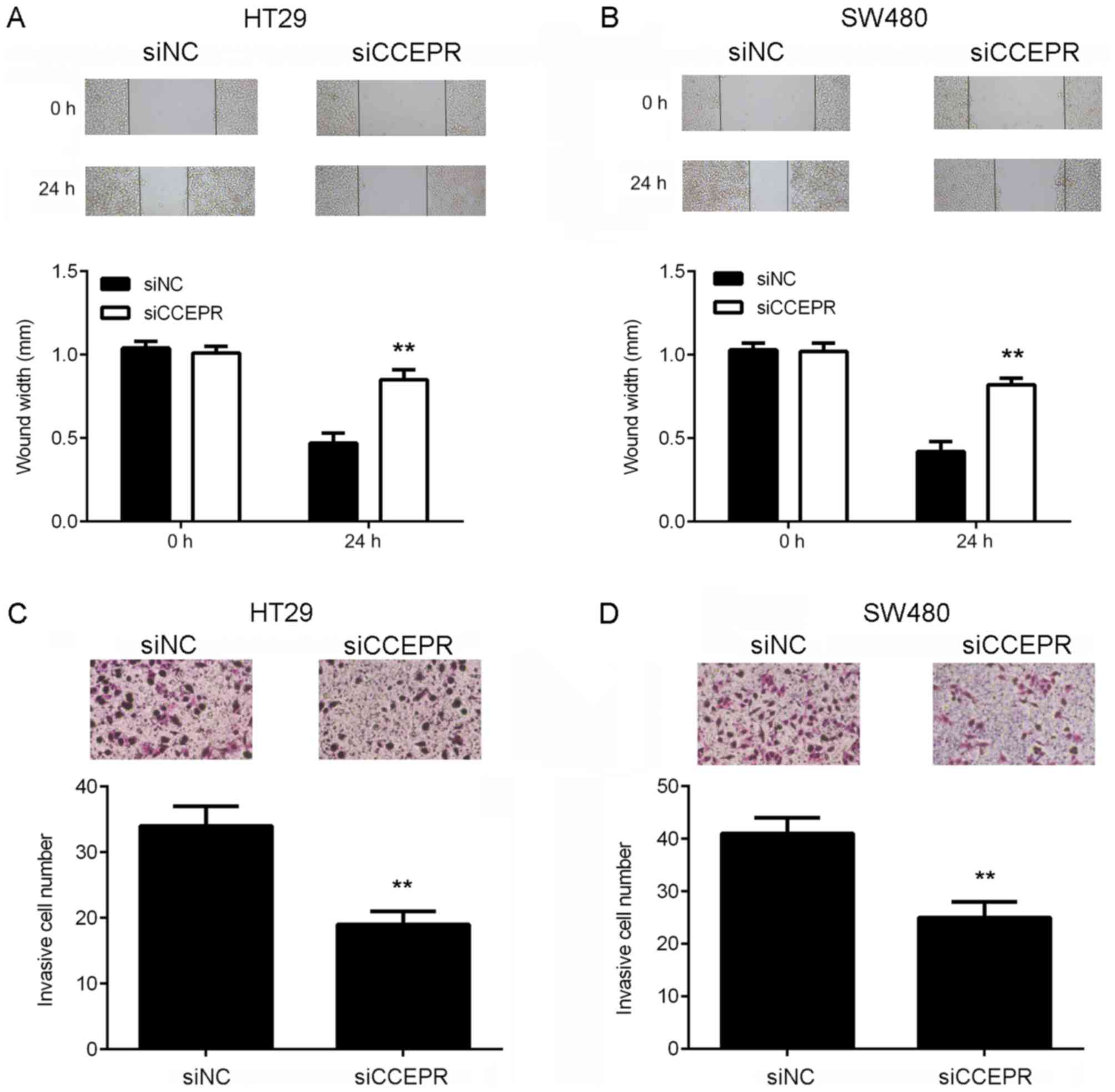
Following the cell growth experiments, the function
of CCEPR in colorectal cancer metastasis in vitro was
investigated. Wound healing and Transwell assays were performed to
examine the effects of CCEPR downregulation on colorectal cancer
cell migration and invasion, respectively. The results of the wound
healing assay demonstrated that the migratory capacity of the HT29
and SW480 cells in the siCCEPR group was significantly attenuated
when compared with the cells in the siNC group (Fig. 3A and B). In addition, the Transwell
assay revealed that the number of invasive cells in the siCCEPR
group was significantly decreased when compared with the siNC group
(Fig. 3C and D), indicating that
knockdown of CCEPR suppressed colorectal cancer cell invasion.
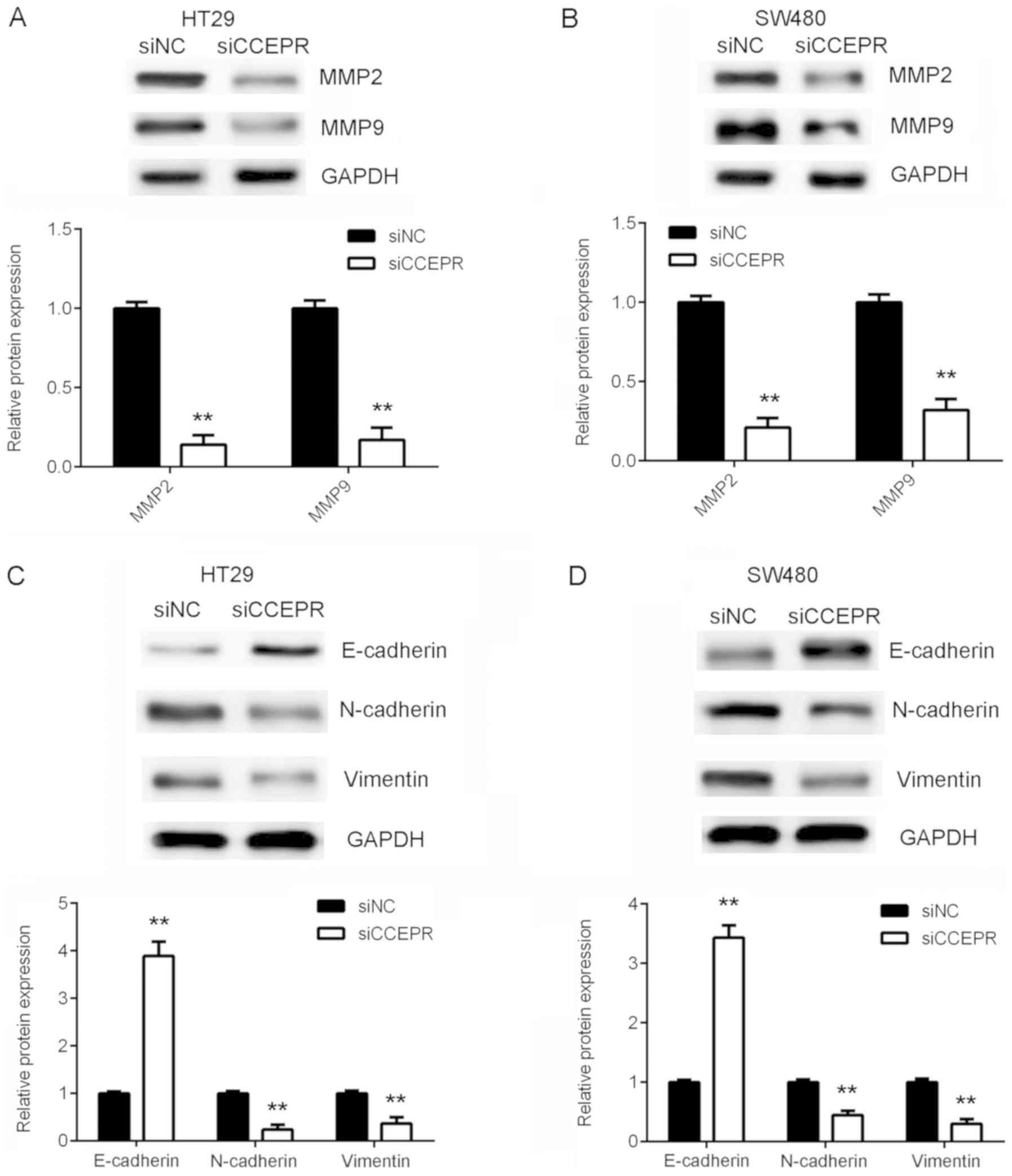
Taking the results presented thus far into consideration, the
authors of the present study hypothesized that CCEPR may serve an
oncogenic role in colorectal cancer metastasis. To further confirm
this hypothesis, the protein expression levels of MMP-2 and MMP-9
were then analysed. The results demonstrated that knockdown of
CCEPR significantly downregulated MMP-2 and MMP-9 expression in
HT29 and SW480 cells (Fig. 4A and
B). EMT is known to serve an essential role in cancer cell
migration and invasion (10).
Therefore, the effects of CCEPR downregulation on EMT in colorectal
cancer cells was subsequently investigated. In HT29 and SW480 cells
from the siCCEPR group, the protein expression levels of E-cadherin
(an epithelial marker) were significantly increased, while the
protein expression levels of N-cadherin and vimentin (mesenchymal
markers) were reduced when compared with cells from the siNC group
(Fig. 4C and D). This indicated that
knockdown of CCEPR significantly inhibited EMT in colorectal cancer
cells. Therefore, it was hypothesized that silencing CCEPR may
inhibit colorectal cancer cell migration and invasion via
suppression of EMT.
Discussion
The underlying molecular mechanisms of CCEPR in
colorectal cancer progression is currently unknown. The present
study observed that the expression of CCEPR was significantly
higher in colorectal cancer tissues when compared with paired
adjacent normal tissues. In addition, CCEPR expression was
significantly higher in advanced colorectal cancer tissues (stage
III/IV) than early-stage colorectal cancer tissues (stage I/II).
High CCEPR expression was significantly associated with poor
differentiation, advanced clinical stage, positive lymph node
metastasis and distant metastasis. Of particular note, patients
with colorectal cancer exhibiting high CCEPR expression had shorter
survival rates when compared with patients exhibiting low CCEPR
expression. Subsequent in vitro experiments determined that
the expression of CCEPR was significantly higher in colorectal
cancer cell lines when compared with a normal colon cell line.
Knockdown of CCEPR significantly inhibited colorectal cancer cell
proliferation, colony formation, cell cycle progression, cell
migration and invasion. In addition, silencing of CCEPR also
downregulated MMP-2 and MMP-9 expression, and suppressed EMT in
colorectal cancer cells.
An increasing number of lncRNAs have been reported
to serve as important mediators in human cancers (27,28).
Specific lncRNAs are significantly deregulated in colorectal cancer
and have been demonstrated to promote tumour growth and metastasis.
For instance, the growth arrest-specific 5 lncRNA is a prognostic
marker in colorectal cancer that has been demonstrated to inhibit
tumour cell proliferation, induce G0/G1 arrest and apoptosis
(29). In addition, the small
nucleolar RNA host gene 12 lncRNA promotes the growth and inhibits
apoptosis of colorectal cancer cells (30). The present study observed that the
expression levels of CCEPR were significantly increased in
colorectal cancer tissues and cell lines when compared with
adjacent normal tissues and non-tumour HIEC cells, respectively.
Consistent with these observations, Gaballah et al (20) also reported that CCEPR expression was
upregulated in 60 colorectal cancer tissues than in adjacent normal
tissues. The current study also demonstrated that the expression
levels of CCEPR were higher in advanced colorectal cancer tissues
(stage III/IV) compared with early-stage colorectal cancer tissues
(stage I/II). Further investigation demonstrated that high CCEPR
expression was associated with poor differentiation, positive lymph
node metastasis, distant metastasis and advanced clinical stage in
colorectal cancer. Consistent with these observations, Gaballah
et al (20) determined that
increased CCEPR expression was associated with increased tumour
size, poor differentiation, advanced Dukes' stage, positive lymph
node involvement and vascular invasion. Taking the results of the
present and previous studies into consideration, the authors
hypothesize that increased CCEPR expression may therefore serve a
role in colorectal cancer progression. In addition, the present
study is the first to investigate whether CCEPR expression may
predict the prognosis of patients with colorectal cancer. The
results demonstrated that patients with colorectal cancer
exhibiting high CCEPR expression levels had shorter survival times
than patients with low CCEPR expression levels.
Based on the results observed in clinical samples,
the present study performed further in vitro experiments to
investigate the function of CCEPR in regulating the malignant
phenotypes of colorectal cancer cells. As CCEPR expression was
significantly upregulated in colorectal cancer samples, CCEPR siRNA
was used to transfect HT29 and SW480 cells to downregulate its
expression. The results of the in vitro experiments
demonstrated that silencing CCEPR significantly suppressed
colorectal cancer cell proliferation and colony formation capacity,
potentially via inducing cell cycle arrest at the G1 stage. These
results suggest that CCEPR serves an oncogenic role in colorectal
cancer growth. In addition, inhibition of CCEPR was associated with
a significant reduction in colorectal cancer cell migration and
invasion when compared with cells expressing normal endogenous
CCEPR levels. This suggests that CCEPR may promote colorectal
cancer metastasis, which is consistent with the clinical findings.
Similarly, Liao et al (17)
demonstrated that CCEPR promotes the proliferation, metastasis and
invasion of non-small cell lung cancer cells. Peng et al
(18) reported that silencing of
CCEPR significantly inhibits hepatocellular carcinoma cell growth
and induces cell apoptosis.
EMT is characterized by loss of the epithelial
phenotype and the acquisition of mesenchymal properties; a process
that is essential for cancer cells to escape their original sites
and gain invasion and migration capabilities (22,31). It
has been widely reported that inhibiting EMT effectively suppresses
cancer cell migration and invasion (22,31).
Recent studies have implicated several lncRNAs in the regulation of
EMT in colorectal cancer (32–34). For
instance, the X-inactive specific transcript lncRNA promotes
metastasis and modulates EMT in colorectal cancer (32). However, whether CCEPR regulates EMT
in colorectal cancer cells currently remains unclear. In the
present study, western blot analysis demonstrated that silencing of
CCEPR in HT29 and SW480 cells resulted in significant upregulation
of the epithelial marker, E-cadherin, but a significant
downregulation in the expression of two mesenchymal markers,
N-cadherin and vimentin, indicating that EMT was suppressed.
Therefore, the inhibitory effects of CCEPR downregulation on
colorectal cancer cell invasion and migration may be attributed to
the inhibition of EMT.
In conclusion, the results of the present study
demonstrated that CCEPR is upregulated in colorectal cancer and may
be associated with colorectal cancer progression, as well as poor
prognosis in patients. In addition, CCEPR may serve an oncogenic
role in regulating the malignant phenotypes of colorectal cancer
cells. Therefore, CCEPR may be a promising therapeutic target for
colorectal cancer treatment.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analysed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
ZF collected clinical tissues. LZ designed the study
and wrote the manuscript. YD and ZS performed experiments and the
statistical analyses. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of Hunan Provincial People's Hospital, Changsha, China.
All patients provided written informed consent.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Cusimano A, Balasus D, Azzolina A, Augello
G, Emma MR, Di Sano C, Gramignoli R, Strom SC, McCubrey JA,
Montalto G and Cervello M: Oleocanthal exerts antitumor effects on
human liver and colon cancer cells through ROS generation. Int J
Oncol. 51:533–544. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2017. CA Cancer J Clin. 67:7–30. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2015. CA Cancer J Clin. 65:5–29. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Munera JO, Sundaram N, Rankin SA, Hill D,
Watson C, Mahe M, Vallance JE, Shroyer NF, Sinagoga KL,
Zarzoso-Lacoste A, et al: Differentiation of human pluripotent stem
cells into colonic organoids via transient activation of BMP
signaling. Cell Stem Cell. 24:8292019. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Xiao Z, Qu Z, Chen Z, Fang Z, Zhou K,
Huang Z, Guo X and Zhang Y: LncRNA HOTAIR is a prognostic biomarker
for the proliferation and chemoresistance of colorectal Cancer via
MiR-203a-3p-Mediated Wnt/ss-catenin signaling pathway. Cell Physiol
Biochem. 46:1275–1285. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Xie B, Deng Z, Pan Y, Fu C, Fan S, Tao Y,
Zhou J and Xiao D: Post-transcriptional regulation DPC4 gene by
miR-190 in colorectal cancer cells. J Cancer Res Ther. 14:838–843.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Yu SY, Dong B, Tang L and Zhou SH: LncRNA
MALAT1 sponges miR-133 to promote NLRP3 inflammasome expression in
ischemia-reperfusion injured heart. Int J Cardiol. 254:502018.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Xu R, Zhu X, Chen F, Huang C, Ai K, Wu H,
Zhang L and Zhao X: LncRNA XIST/miR-200c regulates the stemness
properties and tumourigenicity of human bladder cancer stem
cell-like cells. Cancer Cell Int. 18:412018. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Xiong W, Huang C, Deng H, Jian C, Zen C,
Ye K, Zhong Z, Zhao X and Zhu L: Oncogenic non-coding RNA NEAT1
promotes the prostate cancer cell growth through the SRC3/IGF1R/AKT
pathway. Int J Biochem Cell Biol. 94:125–132. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Luo J, Chen J, Li H, Yang Y, Yun H, Yang S
and Mao X: LncRNA UCA1 promotes the invasion and EMT of bladder
cancer cells by regulating the miR-143/HMGB1 pathway. Oncol Lett.
14:5556–5562. 2017.PubMed/NCBI
|
|
11
|
Zhu H, Zheng T, Yu J, Zhou L and Wang L:
LncRNA XIST accelerates cervical cancer progression via
upregulating Fus through competitively binding with miR-200a.
Biomed Pharmacother. 105:789–797. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhang H and Lu W: LncRNA SNHG12 regulates
gastric cancer progression by acting as a molecular sponge of
miR320. Mol Med Rep. 17:2743–2749. 2018.PubMed/NCBI
|
|
13
|
Liu K, Yao H, Wen Y, Zhao H, Zhou N, Lei S
and Xiong L: Functional role of a long non-coding RNA
LIFR-AS1/miR-29a/TNFAIP3 axis in colorectal cancer resistance to
pohotodynamic therapy. Biochim Biophys Acta. 1864:2871–2880. 2018.
View Article : Google Scholar
|
|
14
|
Chen Y, Wang CX, Sun XX, Wang C, Liu TF
and Wang DJ: Long non-coding RNA CCHE1 overexpression predicts a
poor prognosis for cervical cancer. Eur Rev Med Pharmacol Sci.
21:479–483. 2017.PubMed/NCBI
|
|
15
|
Yang M, Zhai X, Xia B, Wang Y and Lou G:
Long noncoding RNA CCHE1 promotes cervical cancer cell
proliferation via upregulating PCNA. Tumour Biol. 36:7615–7622.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Xu G, Zhang Y, Li N, Zhang JB and Xu R:
LncRNA CCHE1 in the proliferation and apoptosis of gastric cancer
cells. Eur Rev Med Pharmacol Sci. 22:2631–2637. 2018.PubMed/NCBI
|
|
17
|
Liao Y, Cheng S, Xiang J and Luo C: lncRNA
CCHE1 increased proliferation, metastasis and invasion of non-small
lung cancer cells and predicted poor survival in non-small lung
cancer patients. Eur Rev Med Pharmacol Sci. 22:1686–1692.
2018.PubMed/NCBI
|
|
18
|
Peng W and Fan H: Long noncoding RNA CCHE1
indicates a poor prognosis of hepatocellular carcinoma and promotes
carcinogenesis via activation of the ERK/MAPK pathway. Biomed
Pharmacother. 83:450–455. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhan Y, Li Y, Guan B, Chen X, Chen Z, He
A, He S, Gong Y, Peng D, Liu Y, et al: Increased expression of long
non-coding RNA CCEPR is associated with poor prognosis and promotes
tumorigenesis in urothelial bladder carcinoma. Oncotarget.
8:44326–44334. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Gaballah HH, Gaber RA, Elrashidy MA,
Elshahat DA, Hablus MA and Ebeid AM: Expression of long non-coding
RNA CCHE1 in colorectal carcinoma: Correlations with
clinicopathological features and ERK/COX-2 pathway. Mol Biol Rep.
46:657–667. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Pietruszewska W, Bojanowska-Pozniak K and
Kobos J: Matrix metalloproteinases MMP1, MMP2, MMP9 and their
tissue inhibitors TIMP1, TIMP2, TIMP3 in head and neck cancer: An
immunohistochemical study. Otolaryngol Pol. 70:32–43. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wu Y, Sarkissyan M and Vadgama JV:
Epithelial-mesenchymal transition and breast cancer. J Clin Med.
5(pii): E132016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Jiang Z, Song Q, Zeng R, Li J, Li J, Lin
X, Chen X, Zhang J and Zheng Y: MicroRNA-218 inhibits EMT,
migration and invasion by targeting SFMBT1 and DCUN1D1 in cervical
cancer. Oncotarget. 7:45622–45636. 2016.PubMed/NCBI
|
|
24
|
Barrette K, Van Kelst S, Wouters J,
Marasigan V, Fieuws S, Agostinis P, van den Oord J and Garmyn M:
Epithelial-mesenchymal transition during invasion of cutaneous
squamous cell carcinoma is paralleled by AKT activation. Br J
Dermatol. 171:1014–1021. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Yang F, Yu N, Wang H, Zhang C, Zhang Z, Li
Y, Li D, Yan L, Liu H and Xu Z: Downregulated expression of
hepatoma-derived growth factor inhibits migration and invasion of
prostate cancer cells by suppressing epithelial-mesenchymal
transition and MMP2, MMP9. PLoS One. 13:e01907252018. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhu L, Yang N, Du G, Li C, Liu G, Liu S,
Xu Y, Di Y, Pan W and Li X: LncRNA CRNDE promotes the
epithelial-mesenchymal transition of hepatocellular carcinoma cells
via enhancing the Wnt/β-catenin signaling pathway. J Cell Biochem.
2018.(Epub ahead of prin).
|
|
28
|
Zhou Y, Chen Y, Ding W, Hua Z, Wang L, Zhu
Y, Qian H and Dai T: LncRNA UCA1 impacts cell proliferation,
invasion, and migration of pancreatic cancer through regulating
miR-96/FOXO3. IUBMB Life. 70:276–290. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Yang Y, Shen Z, Yan Y, Wang B, Zhang J,
Shen C, Li T, Ye C, Gao Z, Peng G, et al: Long non-coding RNA GAS5
inhibits cell proliferation, induces G0/G1 arrest and apoptosis,
and functions as a prognostic marker in colorectal cancer. Oncol
Lett. 13:3151–3158. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wang JZ, Xu CL, Wu H and Shen SJ: LncRNA
SNHG12 promotes cell growth and inhibits cell apoptosis in
colorectal cancer cells. Braz J Med Biol Res. 50:e60792017.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Smith BN and Bhowmick NA: Role of EMT in
metastasis and therapy resistance. J Clin Med. 5(pii): E172016.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Chen DL, Chen LZ, Lu YX, Zhang DS, Zeng
ZL, Pan ZZ, Huang P, Wang FH, Li YH, Ju HQ and Xu RH: Long
noncoding RNA XIST expedites metastasis and modulates
epithelial-mesenchymal transition in colorectal cancer. Cell Death
Dis. 8:e30112017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Tao Y, Han T, Zhang T, Ma C and Sun C:
LncRNA CHRF-induced miR-489 loss promotes metastasis of colorectal
cancer via TWIST1/EMT signaling pathway. Oncotarget. 8:36410–36422.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Yang X, Liu W, Xu X, Zhu J, Wu Y, Zhao K,
He S, Li M, Wu Y, Zhang S, et al: Downregulation of long noncoding
RNA UCA1 enhances the radiosensitivity and inhibits migration via
suppression of epithelialmesenchymal transition in colorectal
cancer cells. Oncol Rep. 40:1554–1564. 2018.PubMed/NCBI
|