Introduction
Sepsis is broadly defined as a dysregulated immune
response to a pathogenic insult which damages host tissues, and
represents a highly complex and often fatal syndrome which varies
widely in its clinical presentation. Currently, sepsis treatment is
primarily limited to supportive medications and broad spectrum
antibiotics, however currently no clinically validated approach
exists to treat the underlying immune dysregulation in sepsis. The
majority of immune-modulating therapeutics that show efficacy in
preclinical sepsis models do not exhibit similar clinical benefits
(1) which is often attributed to the
pervasive use of inappropriate animal models. The use of relevant
animal models is imperative to gain insights into sepsis
pathophysiology, to improve treatment paradigms and to investigate
long-term effects in survivors. Small animal models favor quick,
cost effective and easily executed studies but larger animal models
are often more appropriate to measure multiple clinical parameters
within more relevant physiological contexts to humans.
The availability of genetically defined rodent
strains has enabled in-depth evaluation of immunological parameters
that are not achievable in large animal species (2). Despite controversy surrounding the
applicability of murine studies to human diseases (3), recent evidence suggests that mouse
models parallel many aspects of human inflammatory and
immunological diseases (4).
Nevertheless, large animal models including pigs, sheep and dogs
are more likely to recapitulate pathophysiologic changes seen in
humans (5). The reclassification of
sepsis according to Sepsis-3 has ignited a renewed focus on organ
damage rather than systemic inflammatory criteria as the most
useful diagnostic and prognostic measures for sepsis (6). Consequently, preclinical approaches are
adopting a similar shift away from immunological endpoints towards
more integrated measures (7,8). Ovine models are particularly
advantageous for investigation of physiological outcomes due to the
hemodynamic similarity between sheep and humans (5), the availability of sheep, and the
ability to repurpose clinical instrumentation. Sheep are comparable
to humans for most common coagulation tests (9), and exhibit similar metabolic changes in
response to sepsis (10).
Instrumented septic ovine models have been used to study acute lung
injury (11), fluid resuscitation
(12) and renal inflammation
(13). Recently, an ovine septic
shock incorporating intensive care supports (OSSICS) model which
recapitulates multiple pathophysiological hallmarks of human septic
shock (14) has been used to
investigate renal pathology (15)
and thyroid hormone therapy (16) in
sepsis. However, the utility of this model for studying the
physiologic effects of immunomodulatory therapies has not been
established.
High mobility group box protein 1 (HMGB1) is a
potential therapeutic target in sepsis with a wide therapeutic
window (17). Normally located
within the nucleus, HMGB1 is released during sepsis and promotes
inflammation as a damage associated molecular pattern (DAMP).
Preclinical modelling has shown that methods used to neutralize or
inhibit HMGB1 activity reduce mortality (18).
Previously, our laboratory has demonstrated that
ovine anti-HMGB1 polyclonal antibodies (pAbs) can reduce mortality
in murine endotoxemia (19) and
polymicrobial sepsis (20) in a
similar manner to other studies (21). While the effects of anti-HMGB1 pAb
therapy on the immune phenotype of sepsis may be assessed in the
murine model, little can be ascertained about the impact of such
therapy on sepsis pathophysiology. Therefore, we aimed to determine
the suitability of the OSSICS model for the evaluation of
anti-HMGB1 pAb using clinically-relevant parameters.
Materials and methods
Ovine polyclonal anti-HMGB1
production
Ovine anti-HMGB1 pAbs were generated via
immunization of sheep with recombinant HMGB1 in Freund's adjuvant
and purified from anti-HMGB1 serum via Protein G chromatography as
previously described (19).
Neutralization capacity of ovine anti-HMGB1 pAbs were evaluated as
previously described (19). Purified
pAbs were sterile filtered and adjusted to 5 mg/ml in PBS for
administration.
HMGB1 and anti-HMGB1 quantitation by
ELISA
Sheep serum samples were analyzed by a human HMGB-1
ELISA kit (Cloud-Clone Corp; standard curve range 4,000-63 pg/ml)
according to the manufacturer's instructions. Anti-HMGB1 was
quantified in sheep serum using a custom ELISA method. Briefly,
ELISA plates were coated with recombinant HMGB1 antigen (5 µg/ml)
produced as previously described (19) and blocked with BSA. Sheep serum
samples were diluted 1:2 and bound ovine antibodies were detected
with anti-ovine IgG-HRP (Donkey anti-sheep IgG A3415; Sigma
Aldrich) and OPD substrate (SigmaFAST; Sigma Aldrich). Readings
were compared to a standard curve prepared from an aliquot of
anti-HMGB1 pAb preparation.
Sheep preparation and monitoring
All sheep studies were approved by at The South
Australian Health and Medical Research Institute (Adelaide,
Australia) Animal Ethics Committee and conducted following
institutional and national ethical guidelines according to the
‘Australian code for the care and use of animals for scientific
purposes (2013)’. Border Leicester × Merino ewes (2.4–4 years old)
were group housed at The South Australian Health and Medical
Research Institute and given free access to food and water. A model
of OSSICS was performed on six sheep previously published protocols
(15,16). Briefly, anesthesia was induced in
overnight fasted randomly selected sheep by intravenous (IV)
injection of Thiopentone (15 mg/kg) before intubation and
ventilation with isoflurane (2%) in oxygen the morning following
fasting (8–10 am). Catheters were placed under surgical conditions:
an arterial catheter (Angiocath 1.7×133 mm; Becton Dickinson) was
placed into the right carotid artery, and a quad-lumen central
venous catheter (8.5 Frx20 cm; Arrow International Inc.) and a
large-bore single lumen catheter (8.5 Frx10 cm; Arrow International
Inc.) were both inserted into the right jugular vein. The
large-bore single lumen catheter was used for the placement of a
pulmonary artery catheter (Swan Ganz CCOmbo 7.5 Frx110 cm; Edwards
Lifesciences) performed under radiology imaging.
A tracheostomy tube was inserted into the proximal
trachea (Portex, ID 9.0, 24 cm; Smith's Medical) and a rumen tube
(16 gauge, Bard Medical) was inserted via the nose. An indwelling
urethral catheter (14Fr, Covington; Bard Medical) was placed into
the bladder and attached to a burette for measurement. Sheep were
moved onto a surgical table, lying prone with head and neck
supported in an upright position to prevent edemous fluid
accumulation in the lower limbs, and mechanically ventilated
(Puritan Bennett 7200). Central venous and arterial catheters were
attached to a pressure monitor (Intellivue MP 50; Phillips) and a
pulmonary artery catheter to cardiac output monitor (Vigilance
Monitor, Edwards Lifesciences). Cardiac output was continuously
monitored and indexed to sheep body surface area according to the
calculation (weight (kg) 0.67×0.0842).
All sheep were monitored by a dedicated intensive
care research nurse throughout the study (Table I), and mean arterial pressure (MAP)
was maintained according to the protocol described in Table II. Previous studies have
demonstrated that anti-HMGB1 pAbs are efficacious in reducing
mortality in murine endotoxemia within 24 h (19), thus a short time frame was adopted in
the current study to allow comparison to previous OSSICS cohorts.
Sheep were euthanized under anesthesia at 26 h post-E. coli
administration by IV injection of pentobarbitone (6.5 g). Death was
apparent by unrecordable arterial blood pressure, and confirmed by
a trained intensive care nurse. All laboratory and biochemical
measurements were conducted by technicians blinded to the treatment
group identity.
 | Table I.Sheep monitoring and maintenance. |
Table I.
Sheep monitoring and maintenance.
| Parameter | Monitoring | Maintenance |
|---|
| Sedation | Depth of sedation
continuously monitored; medication dose recorded hourly. | IV infusion of
midazolam (0.1–0.5 mg/kg/h) and ketamine (1–5 mg/kg/h). |
| Ventilation | Respiratory rate,
tidal volume, peak and plateau pressures and O2 and
CO2 saturation recorded hourly. Tracheal suctioning was
performed hourly or as required. | Synchronised
intermittent mandatory ventilation with 10 ml/kg tidal volume and
positive end expiratory pressure of 5 cm H2O.
Respiration adjusted to maintain end tidal CO2 of 30
mmHg. Fraction of inspired oxygen was adjusted to maintain pulse
O2 saturation >95%. |
| Hemodynamics | HR, CVP and
pulmonary arterial pressure were recorded hourly. | Parenteral fluid
was administered at 3 ml/kg/h via peristaltic pump. |
|
| MAP and CO recorded
every 10 min. | CVP was maintained
≥5 mmHg via fluid bolus administration (250 ml saline). |
|
|
| MAP was maintained
≥75 mm/Hg (Table II). |
| Temperature | Recorded every
hour. | Temperature was
maintained <42°C with the application of cool damp drapes to the
back |
| Blood analysis | – | Samples taken every
2 h for cytokine and lactate analysis, every 4 h for blood gas
analysis and every 6 h for pathology analysis including
coagulation, hematological and biochemical tests. |
| Fluid balance | Fluids administered
and urine output recorded hourly. | Total urine
collected at 0, 12 and 26 h was subjected to protein and creatinine
analysis. |
| Table II.Ovine model hemodynamic management
protocol. |
Table II.
Ovine model hemodynamic management
protocol.
| Blood pressure
parameters | Action |
|---|
| MAP <75 mmHg and
CVP <5 mmHg | 250 ml saline
bolus |
| 75 mmHg and CVP ≥5
mmHg | NorA infusion as
below; adjust every 2 min |
| MAP <55
mmHg | ↑ 10 µg/min
NorA |
| MAP 55–59 mmHg | ↑ 5 µg/min
NorA |
| MAP 60–64 mmHg | ↑ 3 µg/min
NorA |
| MAP 65–69 mmHg | ↑ 2 µg/min
NorA |
| MAP 70–74 mmHg | ↑ 1 µg/min
NorA |
| MAP 75–80 mmHg | No change to NorA
dose |
| MAP 81–85 mmHg | ↓ 1 µg/min
NorA |
| MAP 86–90 mmHg | ↓ 2 µg/min
NorA |
| MAP 91–95 mmHg | ↓ 3 µg/min
NorA |
| MAP 96–00 mmHg | ↓ 5 µg/min
NorA |
| MAP >100
mmHg | ↓ 10 µg/min
NorA |
Preparation and administration of E.
coli and anti-HMGB1 polyclonal antibodies
E. coli (Serotype O6, Biotype 1; ATCC 25922)
inoculum was prepared from colonies (8–10) grown
overnight on fresh Columbia horse blood agar (bioMérieux Australia)
emulsified in 100 ml sterile saline to an optical density of 0.25
at 550 nm (empirically determined to yield 1.5×108
CFU/ml). The number of CFUs/ml of each inoculum was determined by
serial dilution and plating onto Columbia Horse Blood agar.
The inoculum (1 ml/kg) was administered to prepared,
anaesthetized sheep as an IV infusion in the over one hour to
achieve 1–1.5×108 CFU/kg based on previous studies
(15,16). Anti-HMGB1 antibodies (10 mg/kg) were
administered via IV infusion over one hour commencing two hours
after the induction of sepsis. This dose rate was based on previous
murine studies and is in molar excess (180-fold) of detectable
plasma HMGB1. Placebo-treated sheep were administered IV saline in
place of the antibody infusion. Bacterial doses and treatments
administered to the six experimental sheep are summarized in
Table III.
| Table III.Characteristics of sheep and survival
outcomes within the study as compared to archive data. |
Table III.
Characteristics of sheep and survival
outcomes within the study as compared to archive data.
| Sheep | Weight, kg | E. coli
dose, CFU/kg | Treatment | Survival time,
h | Cumulative NorA
administered, µg/kg | NorA dose rate mean
AUC, µg/kg |
|---|
| 1 | 68 |
1.08×108 | Placebo (normal
saline) | 26 | 26.6 | 13.3a |
| 2 | 74 |
0.94×108 | Placebo (normal
saline) | 26 | 0 |
|
| 3 | 69 |
1.47×108 | Placebo (normal
saline) | 26 | 718.5 | 715 |
| 4 | 58 |
1.62×108 | 10 mg/kg anti-HMGB1
pAbs | 26 | 556.2 | 794b |
| 5 | 71 |
1.50×108 | 10 mg/kg anti-HMGB1
pAbs | 12 | 234.4 |
|
| 6 | 76 |
1.58×108 | 10 mg/kg anti-HMGB1
pAbs | 25 | 869.4 |
|
| Archive data (n=8;
mean ± SD) | 63±8 |
1.08±0.5×108 | Placebo (normal
saline) | 25±2.1 | 330±403 | 340 |
Statistical analysis
Analysis was performed using GraphPad Prism V3.0
software. Measurements at individual time points were compared
using student's two-tailed t-tests and time course analysis was
performed by two-way analysis of variance.
Results
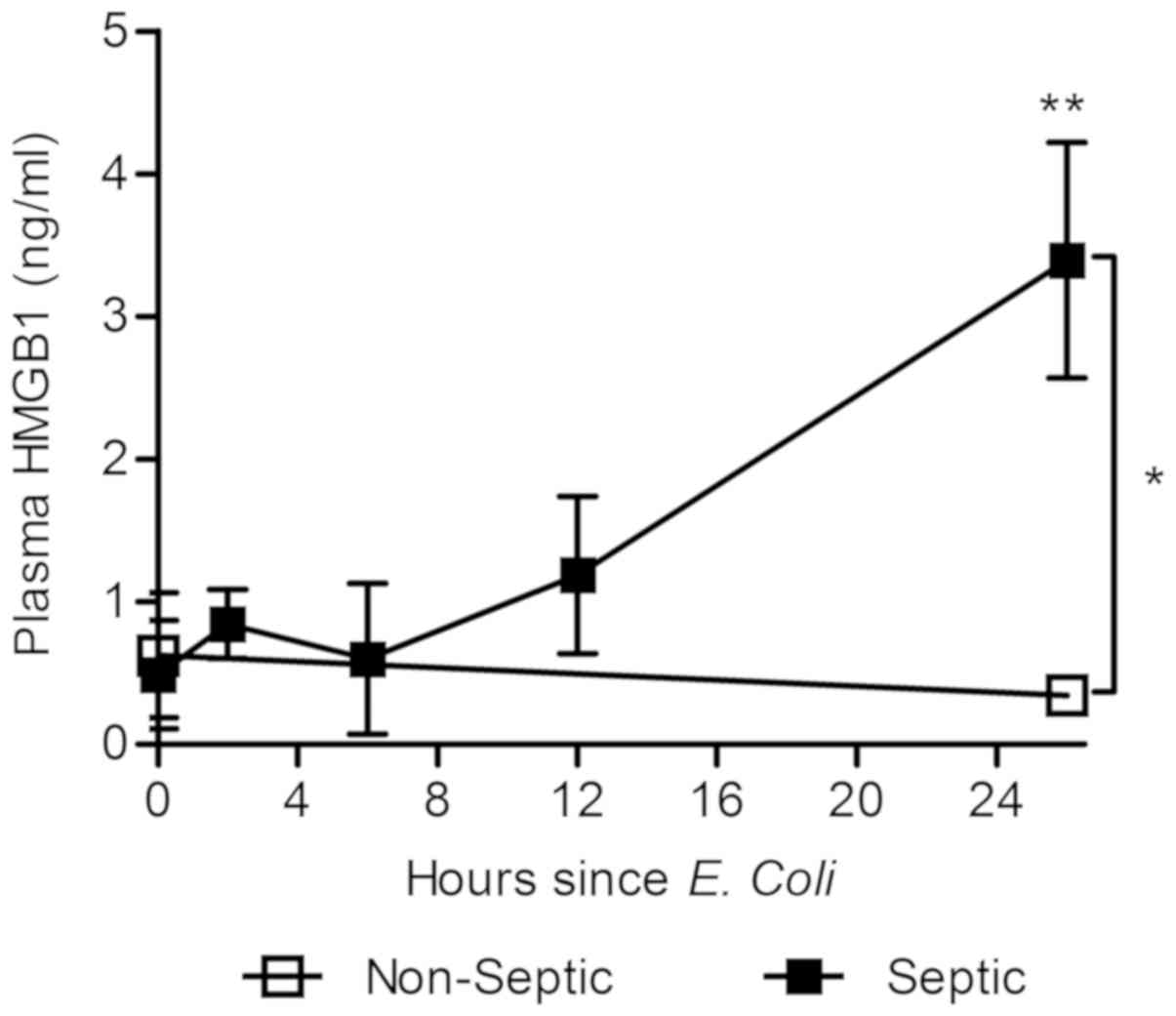
Ovine septic shock induces elevation
of plasma HMGB1
First we determined whether HMGB1 release kinetics
were similar between sheep and humans. Banked plasma samples from
previous OSSICS subjects (see references 15 and 16) and analyzed
for HMGB1 content by commercial ELISA. Elevated HMGB1 was
detectable (Fig. 1) from 12 h post
E. coli infusion (1.08±0.05×108 CFU/kg) while no
HMGB1 elevation was observed in non-septic animals. Significantly,
plasma HMGB1 levels at 26 h were three-fold higher in septic sheep
than non-septic sheep, or those measured at the onset of sepsis
(Fig. 1). Septic sheep exhibited
HMGB1 levels between 2–6 ng/ml over the course of sepsis. These
results support the use of the ovine septic shock model for the
study of anti-HMGB1 therapy in sepsis.
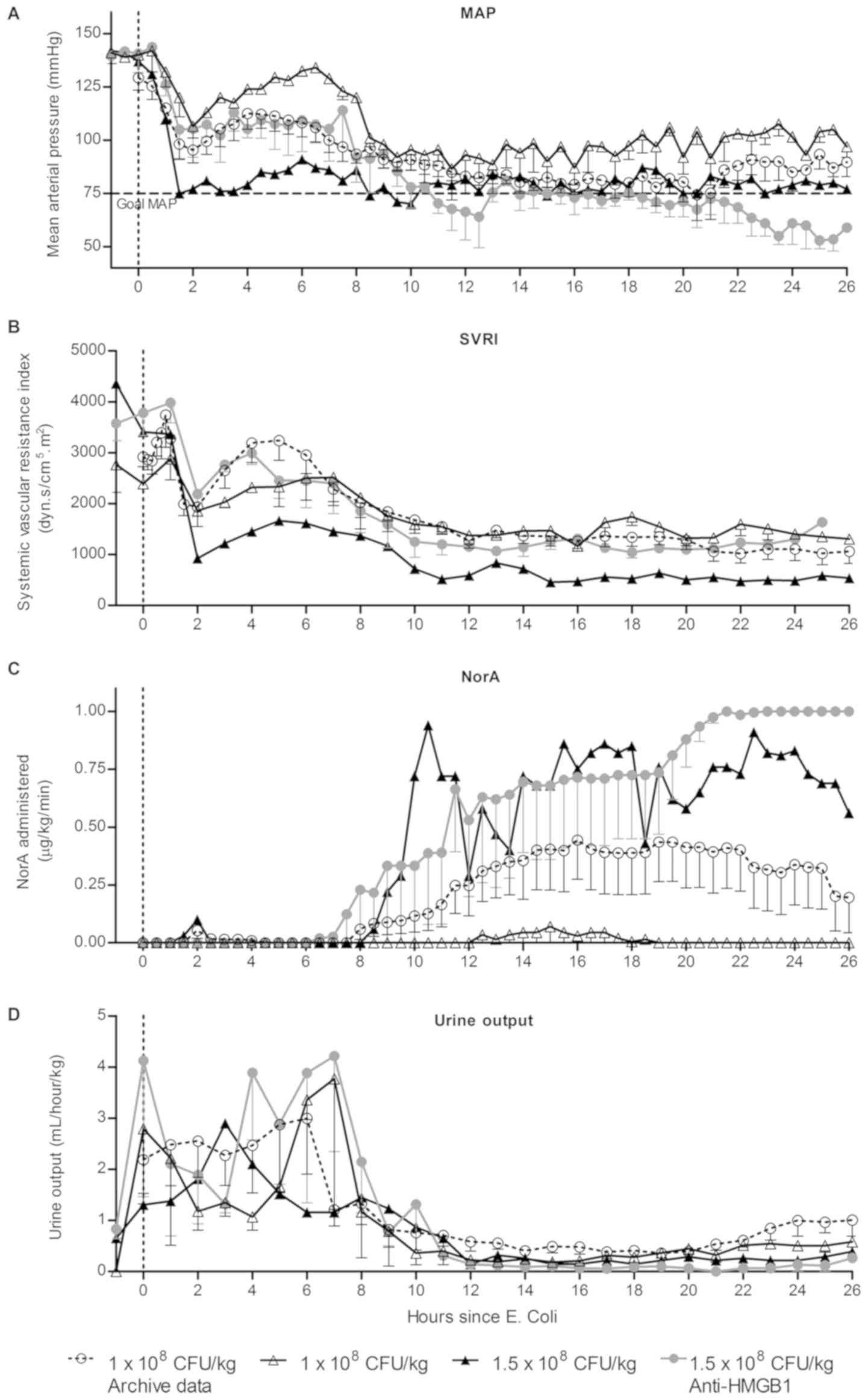
Anti-HMGB1 therapy did not reduce the
development of shock or noradrenaline requirement in the OSSICS
model
The requirement for vasopressor therapy in the form
of noradrenaline (NorA) to maintain adequate blood pressure has
previously been used as the primary outcome measure in the OSSICS
model. Within this study, shock was generated in six sheep and
compared to data from previous studies utilising the same protocol.
Characteristics of study sheep and outcomes are depicted in
Table III. Initially, two sheep
(#1 and #2) were administered the same E. coli dose as used
in previous studies (1×108 CFU/kg) (15,16),
however the sheep required little NorA and did not progress to
prolonged septic shock. To replicate a degree of sickness severity
observed in previous studies, an increased E. coli dose
(1.5×108 CFU/kg) was employed for the remaining sheep
within the study (#3–6). Three sheep were administered anti-HMGB1
pAbs (10 mg/kg; sheep 4–6) two hours after the induction of sepsis.
Two early deaths occurred within this group, as reflected by the
recorded survival time. Hemodynamic parameters were evaluated in
sheep for 1 h before the induction of sepsis and for 26 h
thereafter (Fig. 2). Placebo-treated
sheep from previous studies (n=8; hereafter referred to as archive
data) displayed a rapid decrease in MAP and SVRI within 2 h of
bacterial infusion before increasing to a stable level within 4 h
of infusion (Fig. 2A and B). Between
6–8 h after infusion, MAP and SVRI gradually declined until a MAP
<75 mmHg was achieved and NorA therapy was initiated (Fig. 2C). Two sheep within the current study
were administered 1×108 CFU/kg E. coli and these
sheep displayed a markedly reduced need for NorA compared to
archive data. One sheep with an increased bacterial dose
(1.5×108 CFU/kg) exhibited consistently low MAP and SVRI
and had high NorA requirements. Infusion of anti-HMGB1 pAbs two
hours post-challenge in three sheep that received the increased
bacterial dose of 1.5×108 CFU/kg did not appear to
impact on NorA requirements compared to the equivalently-dosed
control sheep. Urine output (Fig.
2D) in the OSSICS model appeared consistent with archive data,
where measured urine output was between 1–4 ml/hour/kg and
subsequently fell to less than 1 ml/h/kg by 8 h post-challenge.
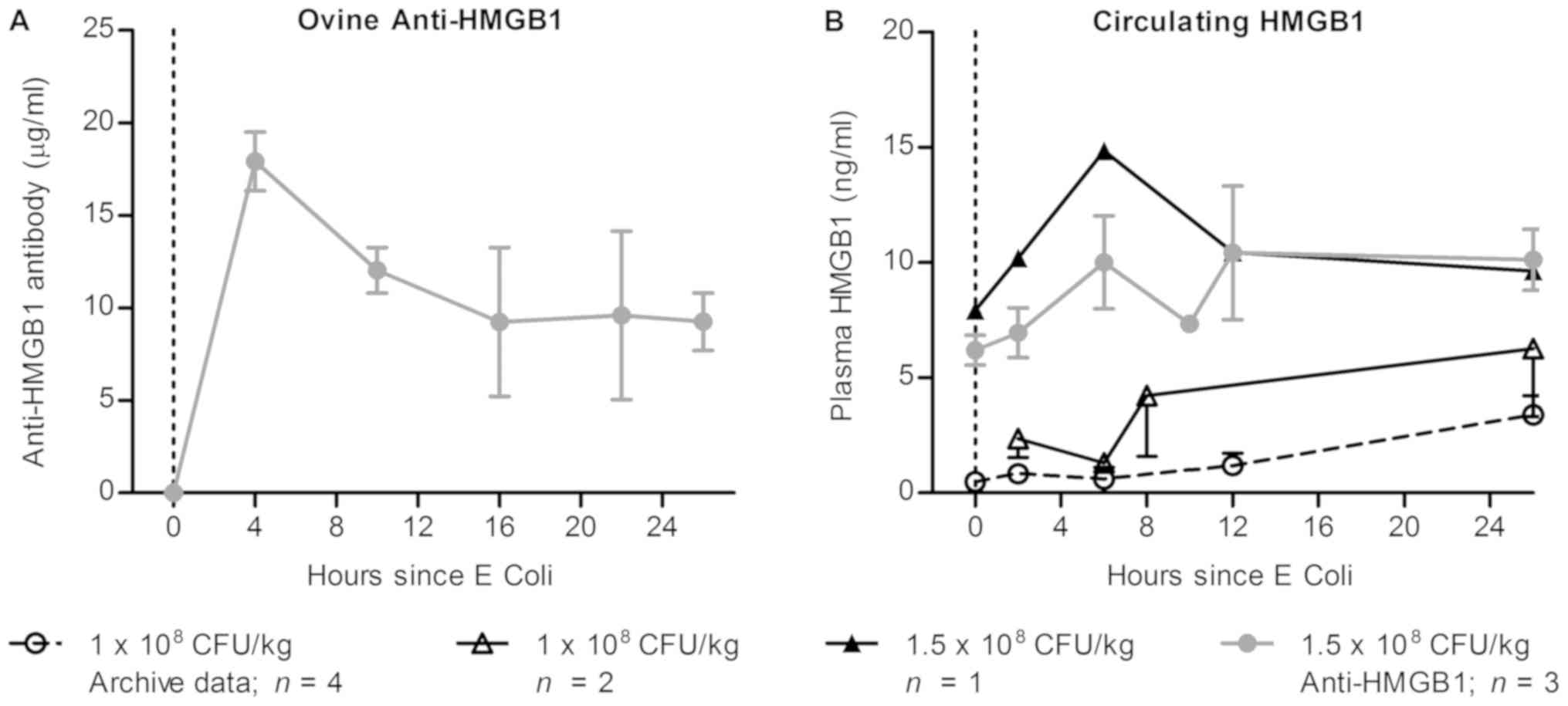
Administration of anti-HMGB1 pAb did
not alter detectable HMGB1 levels in septic sheep plasma
In this preclinical model, a dose rate of 10 mg/kg
anti-HMGB1 pAb was selected based on previous murine studies and on
concentrations of HMGB1 detected within the circulation of septic
sheep. In order to establish whether sufficient anti-HMGB1 pAb was
administered in this study to observe effects, kinetic serum
samples from the three Ab dosed sheep were analyzed by ELISA to
detect anti-HMGB1 ovine IgG (Fig.
3A) and circulating HMGB1 (Fig.
3B). Anti-HMGB1 pAbs were detectable two hours following Ab
infusion at 18 µg/ml (18 mg/l), and decreased thereafter. As one
sheep died 11 h following E. coli infusion, measurements at
later time points are the mean values of two remaining sheep. These
sheep exhibited vastly different kinetics of pAb clearance, as the
detected concentration decreased rapidly in one sheep and remained
constant in another, which may be due to metabolic variability in
the sheep following sepsis induction.
Sepsis-associated alterations in
hematological, coagulative and metabolic parameters were not
improved by anti-HMGB1 treatment
In the current study, blood and plasma samples drawn
from septic sheep were subjected to standard pathology testing and
compared to archive values from placebo-treated septic sheep. Ovine
septic shock has previously been associated with a progressive
decline in packed cell volume (PCV). In the current study,
pre-operative samples were drawn which showed increased PCV and
hemoglobin (data not shown) compared to pre-septic baseline
samples. Measured PCV sharply increased in all groups 2 h post
challenge and declined similarly for all groups for the duration of
the study (Fig. 4Aa). Conversely to
PCV, measured WBC counts increased from pre-operative to baseline
time points (Fig. 4Ab), potentially
due to inflammation caused by surgical catheter placement or
isoflurane exposure. WBC counts decreased sharply by 80% 2 h post
challenge before gradually increasing over time. Further analysis
showed that this decrease was due to marked depletion of
neutrophils, which decreased by 98% from baseline values (Fig. 4Ac), whereas lymphocytes (Fig. 4Ad) and other granulocytes (data not
shown) did not change significantly.
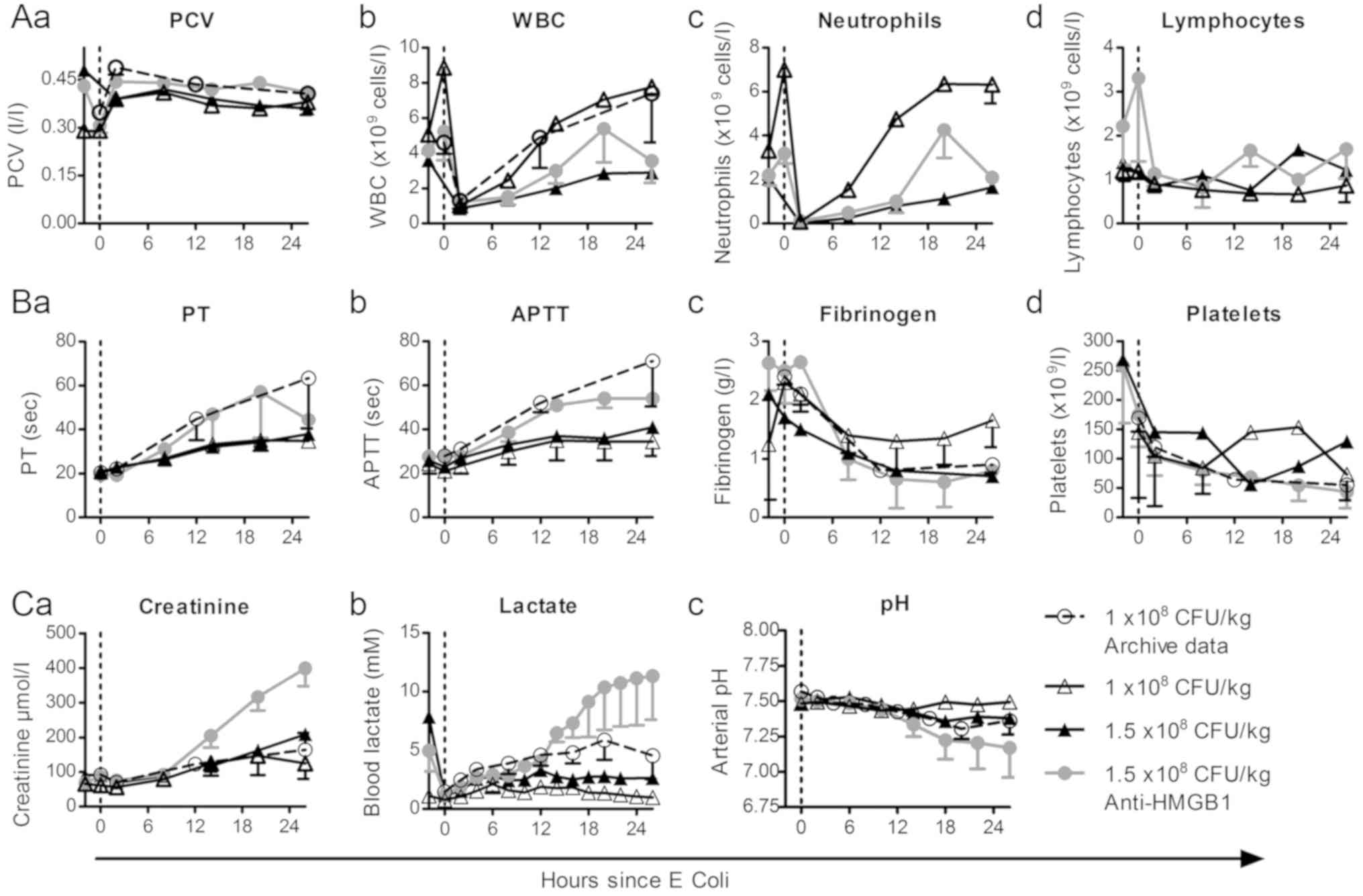 | Figure 4.Measured hematological, coagulative
and metabolic parameters in experimental sheep. Sheep were
administered 1.5×108 CFU/ml E. coli (IV) to
induce sepsis and either treated with anti-HMGB1 2 h post challenge
(n=3) or placebo (saline) treated (n=1). A total of two sheep were
administered a lower E. coli dose (1×108 CFU/ml).
Hematological parameters were measured from whole blood samples
collected every 6 h, with (Aa) PCV, (Ab) WBC, (Ac) neutrophil count
and (Ad) lymphocyte count are presented. Coagulation tests were
performed on plasma samples collected every 6 h, with (Ba) PT, (Bb)
APTT, (Bc) fibrinogen level and (Bd) platelet count from whole
blood represented. Metabolic parameters were assessed via (Ca)
creatinine measurement from six-hourly plasma samples with (Cb)
lactate measurement from two-hourly snap-frozen plasma samples and
(Cc) pH assessment via bedside blood gas analysis (Cc).
Experimental groups were compared to archive data (n=8). Data are
presented as the mean-standard error of the mean of treatment
groups. PCV, packed cell volume; WBC, white blood count; PT,
prothrombin time; APTT, activated partial thromboplastic time;
HMGB1, high mobility group box 1. |
Sheep that received the lower E. coli dose
(1×108 CFU/kg) exhibited mild coagulopathy, with only a
modest increase in PT (Fig. 4Ba) and
APTT (Fig. 4Bb) and mild fibrinogen
depletion (Fig. 4Bc) from baseline
values compared to archive data. These sheep also exhibited reduced
platelet depletion (Fig. 4Bd),
although this was not consistent throughout the septic period.
Sheep treated with anti-HMGB1 pAbs exhibited more profound clotting
dysfunction than a placebo-treated sheep given the same E.
coli dose, however these did not appear altered from archive
data and low subject numbers preclude statistical comparison. As
anti-HMGB1 pAb treated sheep exhibited the highest mortality of the
groups, these results support the notion of coagulation function
tests as a marker of disease severity in ovine septic shock,
however further investigation is required to determine if
anti-HMGB1 therapy may influence clotting dysfunction.
Creatinine, lactate and pH were monitored throughout
the experimental period as a measure of metabolic status. Plasma
creatinine (Fig. 4Ca) increased in
all groups from 2 h post challenge, however a marked increase was
observed in sheep treated with anti-HMGB1 pAbs. Similarly, the
anti-HMGB1 pAb group exhibited the most elevated lactate (Fig. 4Cb) measurements and the highest
degree of acidosis (Fig. 4Cc). The
group that received a low E. coli dose exhibited mild
increases in creatinine, and no increase in plasma lactate.
Correspondingly, the development of acidosis was largely absent in
this group, as pH only decreased 0.06 units below baseline. These
results support the notion that metabolic parameters are useful
clinical tests in ovine septic shock that reflect disease
severity.
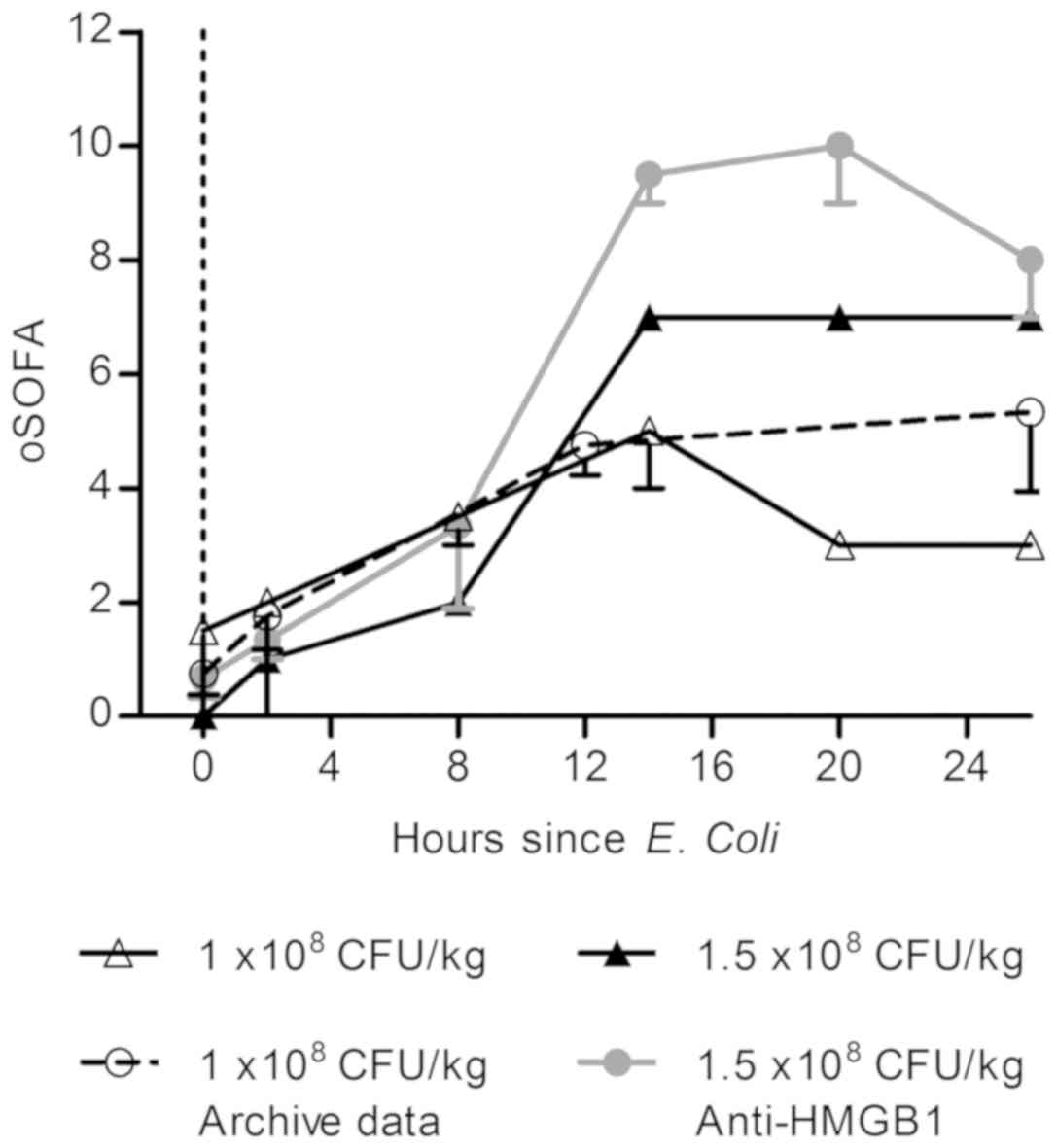
Modified SOFA scores may reflect
severity of illness in the OSSICS model
Clinically, the sequential organ failure assessment
(SOFA) score is used to assess the morbidity associated with sepsis
and incorporates measures of respiratory, hemostatic, liver,
cardiovascular, neurological and renal function. Within this study,
data collected from experimental sheep over time were collated into
a modified SOFA score representing cardiovascular, hemostatic and
renal function using human clinical reference ranges in the absence
of validated ovine ranges (Table
IV). All sheep exhibited an increase in modified SOFA score
over time (Fig. 5), however
increases were most apparent in sheep administered the highest
bacterial dose. These results demonstrate that combined morbidity
scores may have value as endpoints for preclinical sepsis
studies.
| Table IV.Parameters used to calculate the
modified sequential organ failure assessment for experimental
sheep. |
Table IV.
Parameters used to calculate the
modified sequential organ failure assessment for experimental
sheep.
|
| Score assigned |
|---|
|
|
|
|---|
| Parameter | 0 | 1 | 2 | 3 | 4 |
|---|
| Respiratory
PaO2/FiO2, mmHg | >400 | ≤400 | ≤300 | ≤200 | ≤100 |
| Coagulation
platelets, ×103/µl | >150 | ≤150 | ≤100 | ≤50 | ≤20 |
| Cardiovascular MAP,
mmHg or NorA, µg/kg/min | None | MAP <70 |
| NorA ≤0.1 | NorA >0.1 |
| Renal Creat, µmol/l
or urine output, ml/day | Creat
<106.1 | Creat ≥106.1 | Creat ≥176.85 | Creat ≥309.5 or UO
<500 | Creat ≥442.1 or UO
<200 |
Discussion
Sepsis still causes significant mortality in
developed nations and remains the primary cause of death from
infection (22). The
reclassification of sepsis definitions (Sepsis-3) have highlighted
the importance of organ dysfunction for diagnosis6.
Consequently, there is an increased need for animal models than can
recapitulate septic organ damage including circulatory, metabolic,
renal and coagulative dysfunction and efforts to assess
therapeutics should investigate multiple organ systems (8).
Within the present study, the feasibility of using
the OSSICS model for evaluating the impact of anti-HMGB1 pAb
therapy on physiological manifestations of septic shock was
assessed in six sheep. Analysis of banked plasma samples collected
from septic and non-septic OSSICS sheep revealed similar septic
HMGB1 release profiles to that found in septic humans. Ovine HMGB1
has been demonstrated to have inflammatory effects and release
kinetics that mirror those found in humans in studies of
chorioamnionitis (23), cell
migration (24) and fetal brain
injury (25). Consequently, sheep
may respond to HMGB1 blockade in the context of sepsis in a similar
manner to humans.
Initially, trials aimed to reproduce a sickness
severity achieved in previous studies (see references 15 and 16),
however administration of the same E. coli dose
(1×108 CFU/kg) resulted in decreased morbidity with
little requirement for NorA. It was subsequently hypothesized that
a higher E. coli dose (1.5×108 CFU/kg) may
increase the sickness severity to a level observed in previous
studies, thus the remainder of trials were performed with this
higher E. coli dose. An increased bacterial dose increased
requirement for NorA to maintain blood pressure, considered the
primary study endpoint, however these sheep exhibited a very rapid
disease course which culminated in two early deaths. Three sheep
were treated with anti-HMGB1 antibodies via IV injection at a rate
of 10 mg/kg based on previous murine studies. Anti-HMGB1 therapy
was associated with no improvement to MAP, NorA requirement or
vascular resistance in anti-HMGB1 pAb treated sheep compared to
controls, however a high degree of variation within this group, a
low number of subjects and an early death complicated data
interpretation. Additional studies incorporating non-specific pAb
control groups would provide information as to whether the
increased hypotension and morbidity in anti-HMGB1 treated sheep was
specific to anti-HMGB1 or a consequence of IV pAb administration in
this model.
Kinetic analysis of administered anti-HMGB1 pAb in
treated sheep revealed that circulating pAb concentration decreased
over the study period. Similar fluctuations in mAb concentration in
the context of tumor therapy have been attributed to several
mechanisms including Ab deposition, FcR binding or consumption
within immune complexes (26). All
sheep exhibited vasodilation and developed a positive fluid balance
as fluids were administered while urinary production slowed, thus
loss of administered pAb into edematous tissues and peritoneal
fluid accumulations may have occurred. Homology of HMGB1 between
humans and sheep is predicted to be over 98% (27) thus a commercial ELISA manufactured
for measurement of human HMGB1 was able to measure plasma HMGB1 in
septic sheep. The administration of anti-HMGB1 pAb did not reduce
the detectable plasma HMGB1 in the timeframe however it is not
known if detected HMGB1 was bioactive or functionally neutralized
as the antibodies used in the ELISA method may bind different
epitopes to therapeutic pAbs.
Analysis of hematological parameters within this
study supported previous evidence that progressive anemia develops
within the OSSICS model (15). Rapid
declines in WBC and neutrophil counts occurred upon challenge
(28), which failed to recover in
groups receiving a high E. coli dose. While the magnitude of
leukocytosis and neutrophilia have been correlated with the dose of
endotoxin in human challenge studies (29), leukopenia has been associated with
mortality in gram-negative sepsis in humans (30). Thus, in this model, delayed leukocyte
responses may indicate failure to control bacteremia resulting in a
more severe septic phenotype. Sheep from previous studies exhibited
coagulative deficiency and fibrinogen depletion, however similarly
dosed sheep in the current study did not exhibit coagulative
deficiency of this degree. This variation between experiments is
likely to represent alterations in sheep cohorts, as clotting
assays and challenge organism were consistent between the studies.
For future studies, additional controls or a rolling control group
may be employed to minimize the effect of variation between
cohorts. In the current study, anti-HMGB1 pAb treated sheep
exhibited the most profound clotting deficiencies, which may be a
result of marked fibrinogen depletion (apparent in anti-HMGB1 pAb
treated sheep 2 h following challenge) or the depletion of other
clotting factors. Antibody-induced type III hypersensitivity
reactions are associated with platelet activation and the formation
of clotting-factor depleting microthrombi (31), however intravenous immunoglobulin has
also been used to treat sepsis-associated coagulopathy (32). Anti-HMGB1 pAb treated sheep also
exhibited increased acidosis, with creatinine and lactate increased
compared to other groups. Comparison of the current study to OSSICS
sheep administered non-immune ovine pAbs may provide answers as to
the potential relationship between coagulopathy, acidosis and organ
dysfunction and anti-HMGB1 pAb therapy. The high degree of
variation in the OSSICS model compared to murine models does
however allow greater comparison to heterogeneous human sepsis, but
as with human studies large numbers must be included for reliable
conclusions to be drawn (33).
An ideal preclinical sepsis model would incorporate
relevant clinical endpoints to enable clinical translation.
Clinical endpoints for sepsis RCT's have been under scrutiny
(34), and the failure of multiple
sepsis-specific therapies to generate meaningful differences in
study endpoints highlights the need for the careful endpoint
selection. Measures of morbidity during ICU stay are frequently
employed as secondary outcomes in RCT's (35). In the context of animal models,
short-term morbidity endpoints also circumvent ethical and
practical issues associated with measuring long-term mortality
(2). Clinically, composite morbidity
scores such as the SOFA score are used to assess sepsis severity
and morbidity (6). Within this
study, human ranges for the SOFA score calculation were used to
develop a modified SOFA score incorporating measures of
cardiovascular, hemostatic and renal dysfunction (Table IV). Modified SOFA scores were
calculated for experimental sheep and archive data and septic sheep
exhibited increased scores over time, thus a modified SOFA score
may provide a useful assessment of sepsis severity. Practical
limitations prevent the use of the Glasgow coma scale in the OSSICS
model, however future studies could incorporate liver function
tests and respiratory measurements for the purpose of developing an
ovine SOFA. Inclusion of appropriate reference ranges for
physiological criteria and a validated ovine SOFA score equivalent
would increase the utility of the model (36). As the highest bacterial dose used in
this study caused early mortality in two sheep, a lower bacterial
dose or the administration of antibiotics may allow the analysis of
morbidity endpoints, an increasingly common outcome in clinical
trials (35). Short study periods
may only enable evaluation of agents that show marked efficacy in
disease, thus a longer septic period may allow the study of more
moderate differences between groups. Performing additional tests
could provide more insight into the course of OSSICS, such as liver
function tests for liver damage and D-dimer assessment for the
detection of DIC (37). These
foundation studies could help further develop a robust and
authentic large animal model for the assessment of
immunotherapeutics in sepsis.
The increased focus on organ damage in sepsis
monitoring highlights the need for measurement of multiple
parameters in preclinical studies8. While small animal
models are useful for the study of molecular processes and
screening potential therapeutics, large animal models remain an
important step for the study of physiological processes involving
multiple organ systems. While no singular model can possibly
recapitulate the complexities encountered in human sepsis, animal
models remain a crucial tool for preclinical evaluation of
therapeutics. With the refinements, the OSSICS model could become a
robust tool in assessing sepsis therapies via multiple organ
dysfunction measures and facilitate translation into the clinic by
using relevant endpoints. The intelligent and rational optimization
of such models may reduce the discrepancy seen in results between
preclinical and clinical studies and enable the development of
effective host-specific therapeutics targeting sepsis.
Acknowledgements
The authors would like to thank Dr Susan Porter
South (Australian Health and Medical Research Institute, Adelaide,
Australia) for her technical expertise and analysis of ovine
samples, and Mr. Jason Edwards (Royal Adelaide Hospital, Adelaide,
Australia) for his dedicated intensive care unit monitoring.
Funding
The present study was funded by an Australian
Research Council Linkage Project grant (grant no. LP120100606) to
JDH (University of South Australia), MJC (Royal Adelaide Hospital)
and TRK (South Australian Health and Medical Research
Institute).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
NES acquired and analyzed the data, and wrote the
main manuscript text. CHN contributed to the design of the study
and data acquisition. CKF contributed to the data analysis and data
interpretation, and edited the manuscript. TRK designed the
experiments and performed the sheep study. MJM designed the study,
interpreted the data and provided archive data. MJC contributed to
the design of the study and edited the manuscript. KRD contributed
to the design of the study interpreted data, and wrote and edited
the main manuscript. JDH contributed to the conception of the
study, interpreted the data and edited the manuscript.
Ethical approval and consent to
participate
All sheep studies were approved by The South
Australian Health and Medical Research Institute Animal Ethics
Committee (Adelaide, Australia) and conducted following
institutional and national ethical guidelines according to the
‘Australian code for the care and use of animals for scientific
purposes (2013)’.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
CO
|
cardiac output
|
|
CVP
|
central venous pressure
|
|
DAMP
|
damage-associated molecular
pattern
|
|
HMGB1
|
high mobility group box 1
|
|
HR
|
heart rate
|
|
MAP
|
mean arterial pressure
|
|
NorA
|
noradrenaline (Norepinephrine)
|
|
OSSICS
|
ovine septic shock incorporating
intensive care support
|
|
pAb(s)
|
polyclonal antibody(-ies)
|
|
PCV
|
packed cell volume
|
|
RCT
|
randomized controlled trial
|
|
SOFA
|
sequential organ failure
assessment
|
|
SVRI
|
systemic vascular resistance index
|
References
|
1
|
Fink MP: Animal models of sepsis.
Virulence. 5:143–153. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Nemzek JA, Hugunin KM and Opp MR: Modeling
sepsis in the laboratory: Merging sound science with animal
well-being. Comp Med. 58:120–128. 2008.PubMed/NCBI
|
|
3
|
Seok J, Warren HS, Cuenca AG, Mindrinos
MN, Baker HV, Xu W, Richards DR, McDonald-Smith GP, Gao H, Hennessy
L, et al: Genomic responses in mouse models poorly mimic human
inflammatory diseases. Proc Natl Acad Sci USA. 110:3507–3512. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Takao K and Miyakawa T: Genomic responses
in mouse models greatly mimic human inflammatory diseases. Proc
Natl Acad Sci USA. 112:1167–1172. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Di Giantomasso D, Morimatsu H, Bellomo R
and May CN: Effect of low-dose vasopressin infusion on vital organ
blood flow in the conscious normal and septic sheep. Anaesth
Intensive Care. 34:427–433. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Singer M, Deutschman CS, Seymour CW,
Shankar-Hari M, Annane D, Bauer M, Bellomo R, Bernard GR, Chiche
JD, Coopersmith CM, et al: The third international consensus
definitions for sepsis and septic shock (Sepsis-3). JAMA.
315:801–810. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Pickkers P and Kox M: Towards precision
medicine for sepsis patients. Crit Care. 21:112017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Fujishima S: Organ dysfunction as a new
standard for defining sepsis. Inflamm Regen. 36:242016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Foley SR, Solano C, Simonova G, Spanevello
MM, Bird RJ, Semple JW, Jackson DE, Schibler A, Fraser JF and Fung
YL: A comprehensive study of ovine haemostasis to assess
suitability to model human coagulation. Thromb Res. 134:468–473.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Tapia P, Soto D, Bruhn A, Alegría L,
Jarufe N, Luengo C, Kattan E, Regueira T, Meissner A, Menchaca R,
et al: Impairment of exogenous lactate clearance in experimental
hyperdynamic septic shock is not related to total liver
hypoperfusion. Crit Care. 19:1882015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Murakami K, Bjertnaes LJ, Schmalstieg FC,
McGuire R, Cox RA, Hawkins HK, Herndon DN, Traber LD and Traber DL:
A novel animal model of sepsis after acute lung injury in sheep.
Crit Care Med. 30:2083–2090. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Su F, Wang Z, Cai Y, Rogiers P and Vincent
JL: Fluid resuscitation in severe sepsis and septic shock: Albumin,
hydroxyethyl starch, gelatin or ringer's lactate-does it really
make a difference? Shock. 27:520–526. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Fenhammar J, Rundgren M, Hultenby K,
Forestier J, Taavo M Kenne E, Weitzberg E, Eriksson S, Ozenci V,
Wernerson A and Frithiof R: Renal effects of treatment with a TLR4
inhibitor in conscious septic sheep. Crit Care. 18:4882014.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Chapman M, Maiden M, Fraser J, Nash C,
Crichton F, Sideris P and Kuchel T: An ovine intensive care model
of septic shock. Criti Care. 14 (Suppl 1):P42010. View Article : Google Scholar
|
|
15
|
Maiden MJ, Otto S, Brealey JK, Finnis ME,
Chapman MJ, Kuchel TR, Nash CH, Edwards J and Bellomo R: Structure
and function of the kidney in septic shock. A prospective
controlled experimental study. Am J Respir Crit Care Med.
194:692–700. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Maiden MJ, Chapman MJ, Torpy DJ, Kuchel
TR, Clarke IJ, Nash CH, Fraser JD and Ludbrook G: Triiodothyronine
administration in a model of septic shock: A randomized blinded
placebo-controlled trial. Crit Care Med. 44:1153–1160. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Gentile LF and Moldawer LL: HMGB1 as a
therapeutic target for sepsis: It's all in the timing! Expert Opin
Ther Targets. 18:243–245. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Andersson U and Tracey KJ: HMGB1 is a
therapeutic target for sterile inflammation and infection. Annu Rev
Immunol. 29:139–162. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lakhan N, Stevens NE, Diener KR and
Hayball JD: CoVaccine HT™ adjuvant is superior to Freund's
adjuvants in eliciting antibodies against the endogenous alarmin
HMGB1. J Immunol Methods. 439:37–43. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Stevens NE, Chapman MJ, Fraser CK, Kuchel
TR, Hayball JD and Diener KR: Therapeutic targeting of HMGB1 during
experimental sepsis modulates the inflammatory cytokine profile to
one associated with improved clinical outcomes. Sci Rep.
7:58502017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Suda K, Kitagawa Y, Ozawa S, Saikawa Y,
Ueda M, Ebina M, Yamada S, Hashimoto S, Fukata S, Abraham E, et al:
Anti-high-mobility group box chromosomal protein 1 antibodies
improve survival of rats with sepsis. World J Surg. 30:1755–1762.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Fleischmann C, Thomas-Rueddel DO, Hartmann
M, Hartog CS, Welte T, Heublein S, Dennler U and Reinhart K:
Hospital incidence and mortality rates of sepsis. Dtsch Arztebl
Int. 113:159–166. 2016.PubMed/NCBI
|
|
23
|
Regan JK, Kannan PS, Kemp MW, Kramer BW,
Newnham JP, Jobe AH and Kallapur SG: Damage-associated molecular
pattern and fetal membrane vascular injury and collagen
disorganization in lipopolysaccharide-induced intra-amniotic
inflammation in fetal sheep. Reprod Sci. 23:69–80. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Gabrielyan A, Knaak S, Gelinsky M, Arnhold
S and Rösen-Wolff A: Hypoxia-conditioned media allows
species-specific attraction of bone marrow stromal cells without
need for recombinant proteins. BMC Vet Res. 10:562013. View Article : Google Scholar
|
|
25
|
Frasch MG, Szynkaruk M, Prout AP, Nygard
K, Cao M, Veldhuizen R, Hammond R and Richardson BS: Decreased
neuroinflammation correlates to higher vagus nerve activity
fluctuations in near-term ovine fetuses: A case for the afferent
cholinergic anti-inflammatory pathway? J Neuroinflammation.
13:1032016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Tabrizi M, Bornstein GG and Suria H:
Biodistribution mechanisms of therapeutic monoclonal antibodies in
health and disease. AAPS J. 12:33–43. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ito I, Fukazawa J and Yoshida M:
Post-translational methylation of high mobility group box 1 (HMGB1)
causes its cytoplasmic localization in neutrophils. J Biol Chem.
282:16336–16344. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Jain S, Gautam V and Naseem S: Acute-phase
proteins: As diagnostic tool. J Pharm Bioallied Sci. 3:118–127.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Michel O, Nagy AM, Schroeven M, Duchateau
J, Nève J, Fondu P and Sergysels R: Dose-response relationship to
inhaled endotoxin in normal subjects. Am J Respir Crit Care Med.
156:1157–1164. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kreger BE, Craven DE and McCabe WR:
Gram-negative bacteremia. IV. Re-evaluation of clinical features
and treatment in 612 patients. Am J Med. 68:344–355. 1980.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Hara T, Shimizu K, Ogawa F, Yanaba K,
Iwata Y, Muroi E, Takenaka M, Komura K, Hasegawa M, Fujimoto M and
Sato S: Platelets control leukocyte recruitment in a murine model
of cutaneous arthus reaction. Am J Pathol. 176:259–269. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Ishikura H, Nakamura Y, Kawano Y, Tanaka
J, Mizunuma M, Ohta D, Nishida T and Murai A: Intravenous
immunoglobulin improves sepsis-induced coagulopathy: A
retrospective, single-center observational study. J Crit Care.
30:579–583. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Fink MP and Warren HS: Strategies to
improve drug development for sepsis. Nat Rev Drug Discov.
13:741–758. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Opal SM, Dellinger RP, Vincent JL, Masur H
and Angus DC: The next generation of sepsis clinical trial designs:
What is next after the demise of recombinant human activated
protein C?*. Crit Care Med. 42:1714–1721. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Mebazaa A, Laterre PF, Russell JA,
Bergmann A, Gattinoni L, Gayat E, Harhay MO, Hartmann O, Hein F,
Kjolbye AL, et al: Designing phase 3 sepsis trials: Application of
learned experiences from critical care trials in acute heart
failure. J Intensive Care. 4:242016. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Shrum B, Anantha RV, Xu SX, Donnelly M,
Haeryfar SM, McCormick JK and Mele T: A robust scoring system to
evaluate sepsis severity in an animal model. BMC Res Notes.
7:2332014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Sathe PM and Patwa UD: D Dimer in acute
care. Int J Crit Illn Inj Sci. 4:229–232. 2014. View Article : Google Scholar : PubMed/NCBI
|