Introduction
There were >3,000,000 cases of thyroid cancer
worldwide in 2015 and 31,900 deaths occurred as a result in the
same year (1). Thyroid cancer most
commonly occurs between the ages of 35 and 65 years, with women
affected more frequently (2).
Papillary thyroid cancer is the most common type of thyroid cancer,
accounting for 75–85% of all thyroid cancer cases (3). Its incidence has tripled between 1975
and 2012, with the incidence in 2012 being 14.9 per 100,000
individuals (4). Papillary thyroid
cancer is the most common thyroid cancer in children, as well as in
those who had previously received radiation therapy to the head and
neck (5). It has also been observed
in patients with a family history of syndromes, including multiple
endocrine neoplasia type 2 and familial adenomatous polyposis
(6).
Kruppel-like factor 4 (KLF4) has been implicated in
a number of different types of cancer, such as gastric cancer
(7), lung cancer (8) and colorectal cancer (9). MicroRNA (miR)-7 has been previously
demonstrated to suppress the metastasis of breast cancer stem-like
cells into the brain via KLF4 modulation (10). Epigenetic inactivation of KLF4 has
been reported to be associated with the progression and early
recurrence of urothelial cancer (11). The KLF4/Musashi 2 signaling pathway
is also known to regulate the growth and metastasis of pancreatic
cancer (12). Additionally, KLF4 was
revealed to suppress the proliferation of estrogen-dependent breast
cancer by inhibiting the transcriptional activity of estrogen
receptor-α (13). miR-25 has been
reported to enhance non-small cell lung cancer cell migration and
invasion by inhibiting KLF4 via the extracellular signal-regulated
kinase (ERK) signaling pathway (14). The progestin-induced suppression of
miR-29 has been demonstrated to promote the dedifferentiation of
breast cancer cells via KLF4 (15).
However, the role of KLF4 in papillary thyroid cancer remains
elusive.
E-cadherin is an adhesion molecule that suppresses
the invasion of various tumor cells (16). N-cadherin is also a transmembrane
protein that functions to mediate cell-cell adhesion (17). The co-expression of E-cadherin and
Vimentin is associated with the invasion and metastasis of breast
cancer (18). Additionally, matrix
metalloproteinase (MMP) 2 was revealed to be suppressed by
regulatory T cells and was determined to be involved the regulation
of urinary bladder cancer cell invasion (19). The gene expression of MMP9 is
regulated by epigenetic modifications in breast cancer (20). Collagen was also revealed to be
elevated in the serum of patients with non-small cell lung cancer
(21). However, how KLF4 affects the
expression of E-cadherin, N-cadherin, Vimentin, MMP2, MMP9 and
collagen in papillary thyroid cancer remains unclear.
Therefore, the present study aimed to investigate
the role of KLF4 in papillary thyroid cancer and to determine
potential underlying molecular mechanisms.
Materials and methods
Reagents and patients
Primers and probes, TRIzol® reagent,
SuperScript III Reverse Transcriptase, SYBR qPCR mix kit was
purchased from Invitrogen; Thermo Fisher Scientific, Inc. DMEM was
purchased from Gibco (Thermo Fisher Scientific, Inc.).
Thyroid cancer tissues together with the adjacent
non-tumor tissue were surgically removed from 8 patients who were
admitted to Baoshan District Integrated Traditional Chinese and
Western Medicine Hospital (Shanghai, China) between December 2016
and November 2017. The distance between adjacent non-tumor tissue
and the boundary of the cancer tissue was ~1 cm. All patients were
diagnosed with papillary thyroid cancer and their
clinicopathological characteristics were recorded (Table I) (22,23).
After the study was explained, all patients provided written
informed consent. The present study was approved by the
Institutional Research Board of Baoshan District Integrated
Traditional Chinese and Western Medicine Hospital (Shanghai,
China).
 | Table I.Clinicopathological features of
patients in the present study. |
Table I.
Clinicopathological features of
patients in the present study.
| Patient no. | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 |
|---|
| Age (years) | 66 | 50 | 57 | 35 | 55 | 48 | 48 | 52 |
| Sex | F | F | F | M | M | F | F | F |
| TNM | T1aN0M0 | T1aN1M0 | T1N1M0 | T1aN1M0 | T1aN1M0 | T1aN0M0 | T1bN0M0 | T1bN0M0 |
| Tumor stage | I | III | III | I | III | I | I | I |
Immunohistochemistry (IHC)
Tumor tissues and adjacent non-tumor tissues were
embedded in paraffin and cut into slides (5 µm thick). Slides were
fixed in 4% paraformaldehyde for 10 min at room temperature,
blocked with 5% bovine serum albumin (Bio-west, Inc.) at room
temperature for 30 min and incubated with primary antibodies
against KLF4 (1:500; cat. no. Bs-1064R; Bioss Antibodies, Inc.) at
4°C overnight. Following overnight incubation, slides were washed
with PBS and incubated in the dark with FITC-conjugated goat
anti-rabbit secondary antibodies (1:1,000; cat. no. ab6717; Abcam)
at room temperature for 1 h. Slides were subsequently washed with
PBS (3 times; each, 30 sec). Slides were prepared with mounting
media and Antifades (Invitrogen; Thermo Fisher Scientific, Inc.)
and observed using a fluorescence microscope (magnification, ×100).
The mean intensity, calculated by multiplying the area (size) and
average density of fluorescence, was evaluated using Image-Pro Plus
software (version 7; Media Cybernetics, Inc.).
Western blot analysis
The protein expression of KLF4 in tumor tissues and
adjacent non-cancerous tissues was detected via western blot
analysis. In addition, the protein expression of N-cadherin, MMP2,
MMP9 and collagen in KTC1 cells were detected via western blotting.
Tissues were digested and lysed in lysis buffer (cat. no. P0013;
Beyotime Institute of Biotechnology) at 4°C with inhibitors of
phosphatase and protease (cat. no. P1045; Beyotime Institute of
Biotechnology). The lysis mixture was centrifuged at 4°C for 10 min
at 10,000 × g and the supernatant containing cellular proteins was
utilized in the following experiments. Protein concentration was
determined using a BCA kit. Proteins were separated via SDS-PAGE
(10% gel; 40 µg loaded per lane; 120 V). Separated proteins were
then transferred to PVDF membranes (100 V for 120 min; Beyotime
Institute of Biotechnology), which were subsequently blocked with
5% non-fat milk at room temperature for 1 h. Membranes were then
incubated with the following primary antibodies obtained from Abcam
at 4°C overnight: Anti-KLF4, anti-N-cadherin (cat. no. ab18203),
anti-MMP2 (cat. no. ab97779), anti-MMP9 (cat. no. ab228402),
anti-collagen (cat. no. ab138492), anti-GAPDH (cat. no. ab181602)
and anti-β-actin (cat. no. ab8227; all 1:1,000). Membranes were
washed with Tris-buffered saline containing Tween 20 and incubated
with horseradish peroxidase-conjugated goat anti-rabbit secondary
antibodies (cat. no. ab6721; 1:2,000; Abcam) at room temperature
for 1 h. Membranes were incubated in enhanced chemiluminescence
solution (Beyotime Institute of Biotechnology). Images were
captured on film (Beyotime Institute of Biotechnology) in a dark
room. Experiments were repeated three times. The western blot
images were quantified in greyscale using ImageJ software (version
1.5.2; National Institutes of Health).
Construction of recombinant plasmids
and lentiviral packaging
cDNA sequence of KLF4 (NM_004235) was synthesized
and subcloned into lentivirus vector pL6.3-CMV-GFPa1-IRES-MCS
(Novobio) for lentivirus production. The KLF4 recombinant
lentivirus vector, pL6.3-CMV-GFPa1-IRES-KLF4, was confirmed by
Sanger sequencing (24).
Packaging mix (9 µg; Novobio) and KLF4 recombinant
lentiviral plasmids (3 µg) were added into Opti-Minimum Essential
Medium (Opti-MEM; Thermo Fisher Scientific Inc.) and mixed.
Lipofectamine™ 2000 (36 µl; Thermo Fisher Scientific Inc.) was
mixed with Opti-MEM (1.5 ml) and incubated at room temperature for
5 min. The plasmid solution and diluted Lipofectamine 2000 were
then mixed and incubated at room temperature for 5 min. The mixture
was added into a culture dish with 293 T cells (Novobio), and cells
were cultured for 48 h. Cell supernatants were then collected,
centrifuged at 1,500 × g for 10 min at room temperature and
filtered. The lentivirus solution was then condensed via
centrifugation at 50,000 × g for 2 h at 4°C and re-suspended in
DMEM. KLF4 recombinant lentivirus was derived.
The human papillary thyroid carcinoma cell line KTC1
(3×105 cells/well in six-well plates) was transfected
with the pL6.3-CMV-GFPa1-IRES-KLF4 and pL6.3-CMV-GFPa1-IRES-MCS
(control). The transduction MOI was 30. Quantitative PCR (qPCR) was
utilized to detect the efficiency of KLF4 overexpression after 48
h. There was no transfection in blank group.
Reverse transcription-quantitative PCR
(RT-qPCR)
The expression of E-cadherin, N-cadherin, Vimentin,
MMP2, MMP9 and collagen in KTC1 cells was detected via qPCR. Total
RNA was extracted using TRIzol® reagent, according to
the manufacturer's protocol. A universal cDNA synthesis kit
(Invitrogen Thermo Fisher Scientific, Inc.) was utilized for
reverse transcription at 42°C for 1 h. Each reaction contained 0.5
µl random hexamers primers (dN6; 0.2 µg/µl; Novobio) and 1 µl
SuperScript III reverse transcriptase (200 U/µl). The specific
primers used are listed in Table
II. PCR was performed using a SYBR qPCR mix kit (Invitrogen;
Thermo Fisher Scientific, Inc.). The PCR conditions were as
follows: Pre-denaturation at 95°C for 2 min; 40 cycles of
denaturation at 95°C for 10 sec annealing at 60°C for 30 sec and
polymerization at 70°C for 45 sec. qPCR was performed using a CFX96
Touch™ Real-Time PCR Detection system (Bio-Rad Laboratories, Inc.).
Gene expression was determined and normalized to β-actin. The
primer used for rat β-actin was as follows: Forward,
5′-AGGGAAATCGTGCGTGAC-3′ and reverse, 5′-CGCTCATTGCCGATAGTG-3′. The
2−∆∆Cq method was utilized to measure PCR results
(25).
| Table II.Primers for quantitative PCR. |
Table II.
Primers for quantitative PCR.
| Primer | Sequences (5′ to
3′) |
|---|
| KLF4-F |
TTCCCATCTCAAGGCACACC |
| KLF4-R |
CATGTGTAAGGCGAGGTGGT |
| MMP2-F |
GATACCCCTTTGACGGTAAGGA |
| MMP2-R |
CCTTCTCCCAAGGTCCATAGC |
| MMP9-F |
GTACTCGACCTGTACCAGCG |
| MMP9-R |
TTCAGGGCGAGGACCATAGA |
| Collagen I-F |
AGTGGTTTGGATGGTGCCAA |
| Collagen I-R |
GCACCATCATTTCCACGAGC |
| Vimentin
(VIM)-F |
TGGACCAGCTAACCAACGAC |
| Vimentin
(VIM)-R |
GCCAGAGACGCATTGTCAAC |
| E-cadherin
(CDH1)-F |
TCATGAGTGTCCCCCGGTAT |
| E-cadherin
(CDH1)-R |
TCTTGAAGCGATTGCCCCAT |
| N-cadherin
(CDH2)-F |
TGACAATGACCCCACAGCTC |
| N-cadherin
(CDH2)-R |
GTCCTGCTCACCACCACTAC |
Cell viability assay
The viability of KTC1 cells was measured using a
cell counting kit-8 (CCK-8; Dojindo Molecular Technologies, Inc.)
cell viability assay at 24, 48 and 72 h after KTC1 cells were
transfected with the aforementioned viruses. CCK-8 solution was
added to each well and incubated at 37°C for 4 h. The absorbance
was subsequently measured using a microplate reader at 490 nm.
Relative tumor cell viability rate was calculated by dividing the
reading of each group at 24, 48 and 72 h by the baseline reading at
0 h. Experiments were repeated three times.
Transwell invasion assay
The membrane of the upper compartment was coated
with Matrigel (1 g/l; 50 µl), which was allowed to solidify via
incubation at 37°C for 1 h. KTC1 cell suspension (1×104
cells/ml, 200 µl) in 2% DMEM was added to the upper compartment of
each Transwell insert, while 800 µl DMEM with 10% FBS (HyClone; GE
Healthcare Life Sciences) was added to the lower compartment. Cells
were incubated at 37°C for 24 h. Subsequently, 4% paraformaldehyde
was utilized to fix cells on the microporous membrane at room
temperature for 30 min. Cells on the lower side of the membrane
were stained with 1% crystal violet at room temperature for 10 min
and washed with PBS twice. Cells were then observed under Olympus
IX50 fluorescent microscope (magnification, ×400; Olympus
Corporation) and the number of cells that had transgressed through
the membrane was counted. Relative tumor cell invasion was
calculated by dividing the average number of cells that invaded
through the membrane in the experimental groups by that in the
blank group. Experiments were repeated three times.
Scratch migration assay
A confluent monolayer of KTC1 cells was used in the
scratch migration assay. A marker pen was used to draw a straight
line at the back of plate. Pippet tips were utilized to draw
scratch lines vertical to the straight line on the second day. PBS
was used to wash cells three times and DMEM without serum was
added. Images were taken under Olympus IX50 (magnification, ×400)
and cultured in an incubator with 5% CO2 at 37°C. Images
were then taken at 24 h and migration distances were calculated
under the same field using Image-Pro Plus software. Migration
distance at 24 h relative to 0 h was recorded for both negative
control and overexpression groups. Then relative migration
distances were normalized to the negative control group.
Statistical analysis
Experiments were repeated three times. Statistical
data was analyzed using GraphPad Prism software (version 5.0;
GraphPad Software Inc.). The results are presented as the mean ±
standard deviation. Differences among more than three groups were
compared by a one-way analysis of variance followed by the
Bonferroni post-hoc test. Differences between two groups were
compared using Student's unpaired t-test. P<0.05 was considered
to indicate a statistically significant difference.
Results
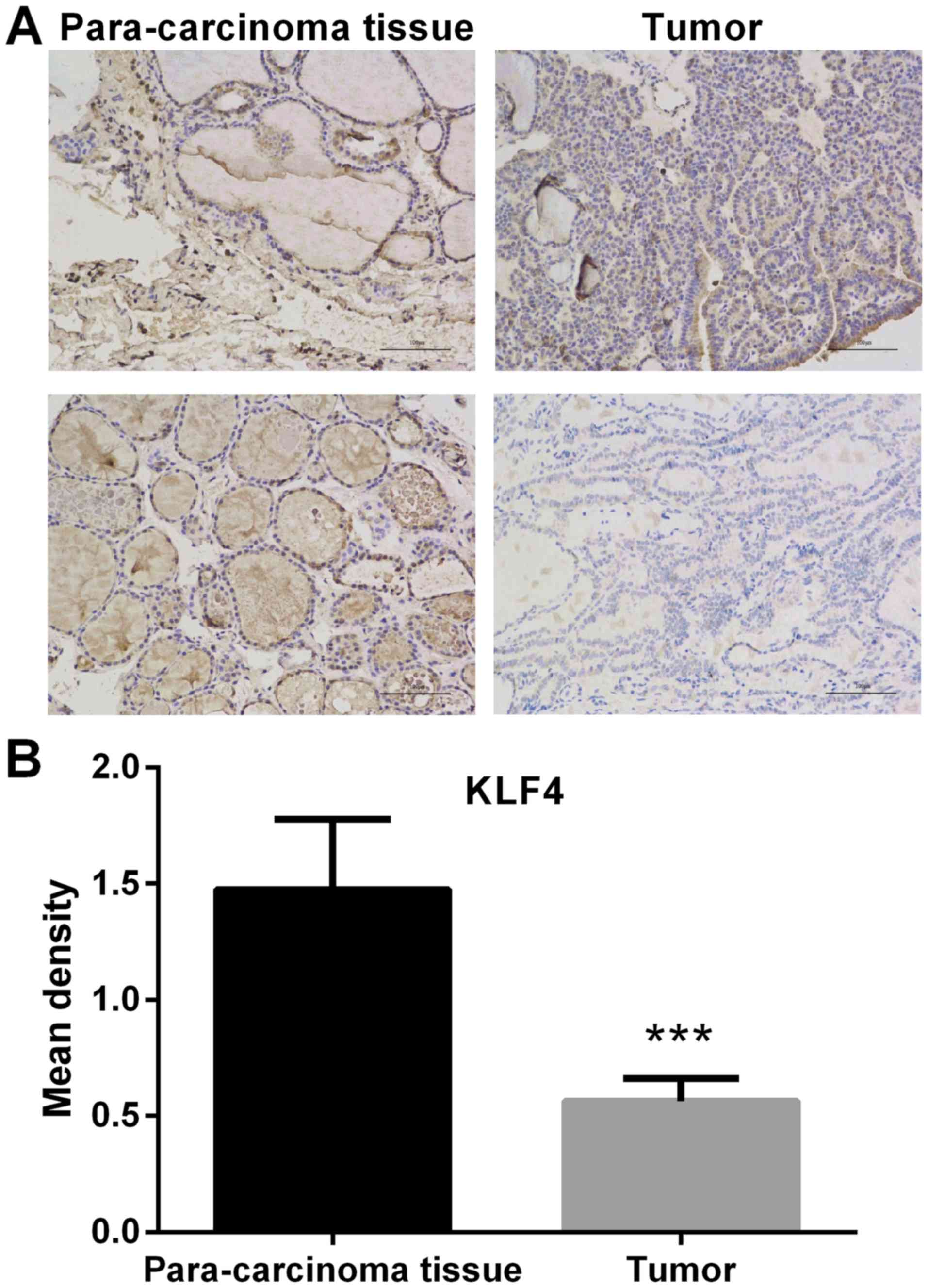
Expression of KLF4 is significantly
lower in thyroid tumor tissue
The expression of KLF4 in thyroid tumor tissue and
paracarcinoma tissue was detected by IHC. In the microscopic image
of KLF4 staining (Fig. 1), positive
cell nucleus was stained by diaminobenzidine (DAB) and appeared as
brown. Negative cell nucleus was stained by hematoxylin and
appeared as blue. Compared with adjacent non-cancerous tissue, the
protein expression of KLF4 was significantly lower in thyroid tumor
tissue (P<0.001; Fig. 1). In
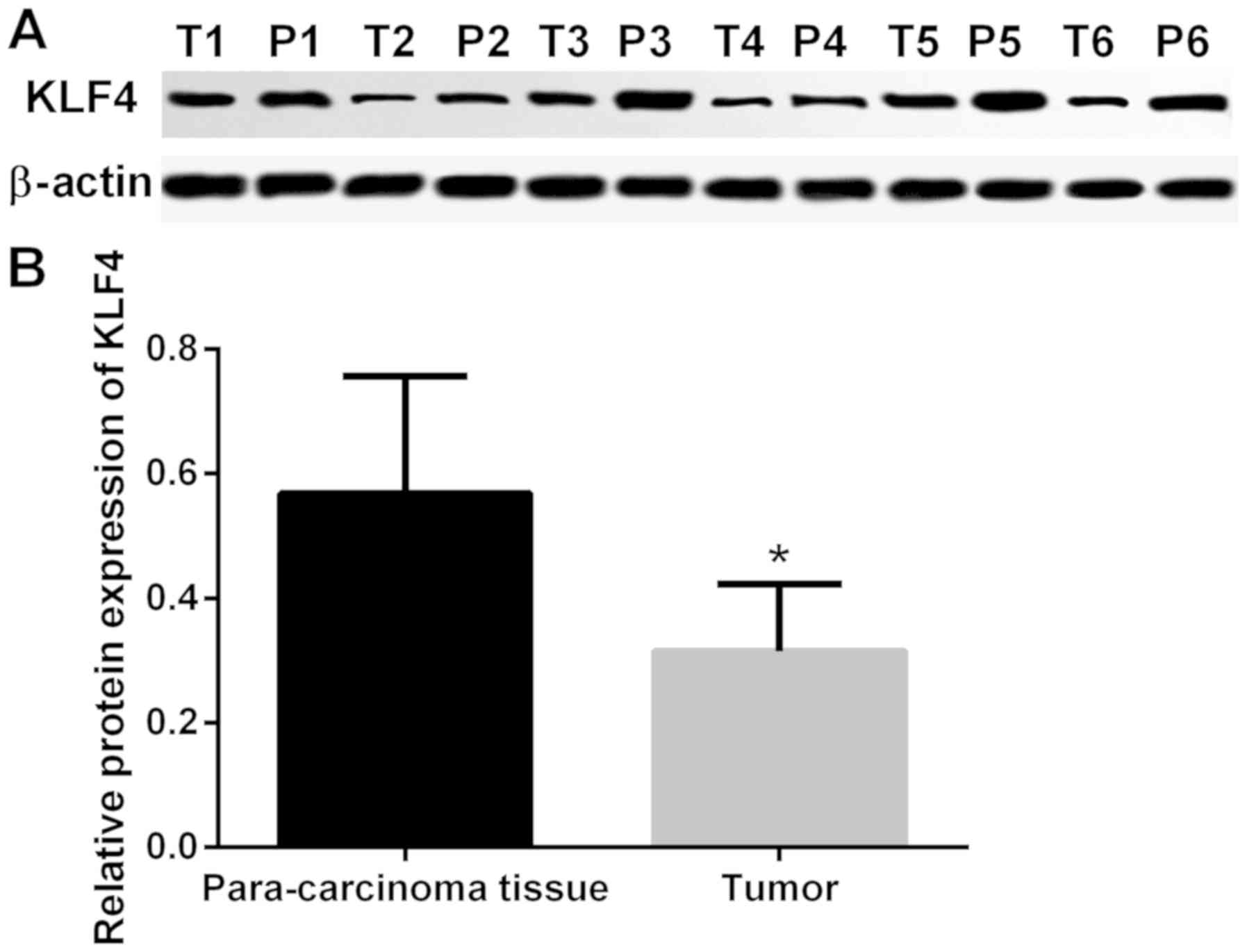
addition, the protein expression of KLF4 in tumor and adjacent
non-cancerous tissue was detected via western blot analysis. The
protein expression of KLF4 was markedly lower in thyroid tumor
tissue when compared with adjacent non-cancerous tissues
(P<0.05; Fig. 2).
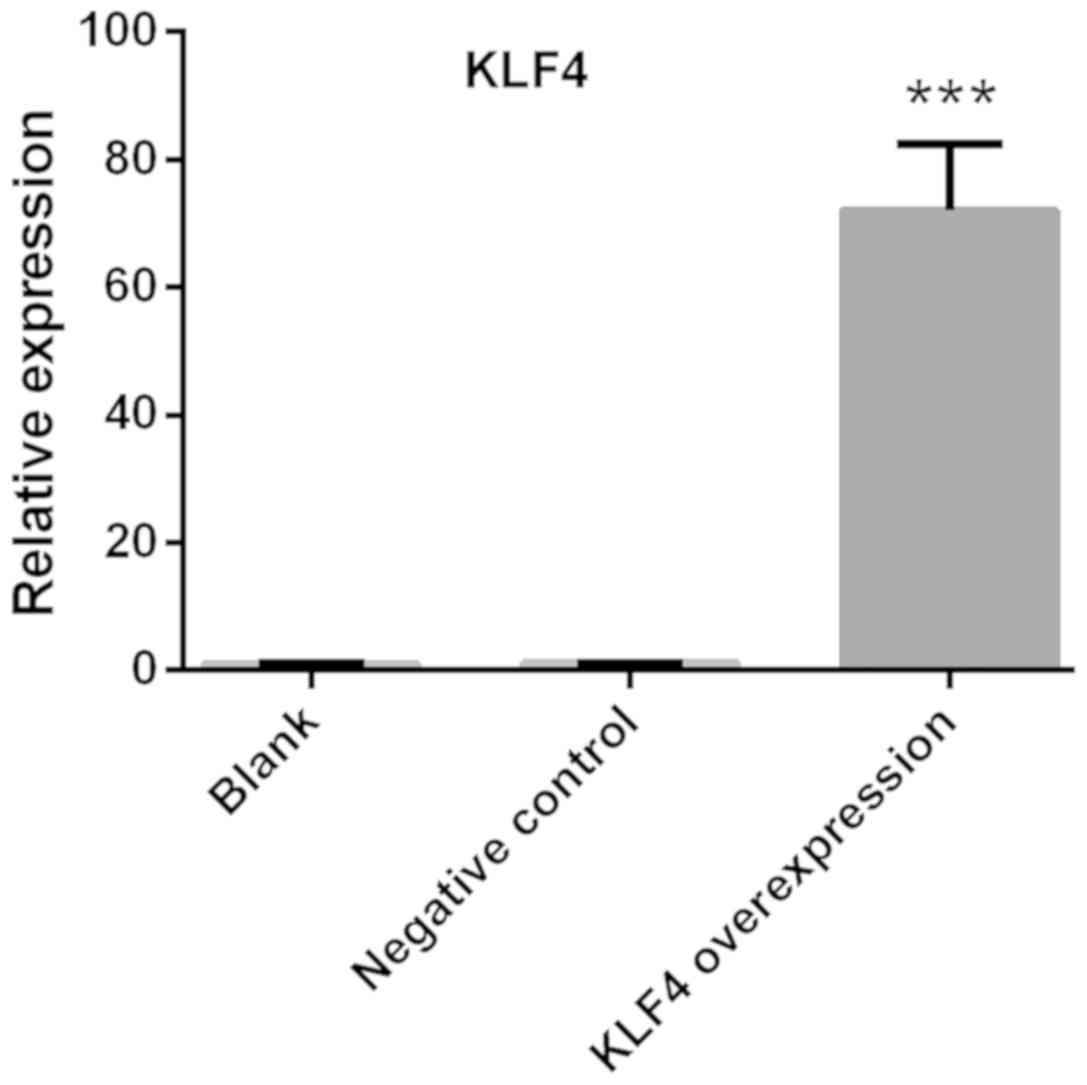
Confirmation of KLF4
overexpression
KTC1 cells were transfected with viruses carrying
KLF4 overexpression vectors. The relative expression of KLF4 was
detected via qPCR. The expression of KLF4 in KTC1 cells was
significantly increased compared with the blank or negative control
groups (P<0.001; Fig. 3).
Expression of N-cadherin, MMP2, MMP9
and collagen is significantly decreased in the KLF4 overexpression
group
The mRNA levels of E-cadherin, N-cadherin, Vimentin,
MMP2, MMP9 and collagen in KTC1 cells were detected via qPCR. The
protein expression of N-cadherin, MMP2, MMP9 and collagen in KTC1
cells were confirmed by western blotting. Among all the genes
screened, the mRNA expression of N-cadherin, MMP2, MMP9 and
collagen were significantly decreased in the KLF4 overexpression
group when compared with the blank or negative control groups
(P<0.05 for N-cadherin and MMP2; P<0.01 for MMP9; P<0.001
for collagen; Fig. 4). No
significant differences were observed in the levels of E-cadherin
and Vimentin (Fig. 4). In addition,
the protein expression of N-cadherin, MMP2, MMP9 and collagen were
significantly decreased in the KLF4 overexpression group when
compared with blank or negative control group (P<0.01 for
N-cadherin; P<0.001 for MMP-2, MMP-9 and collagen; Fig. 5).
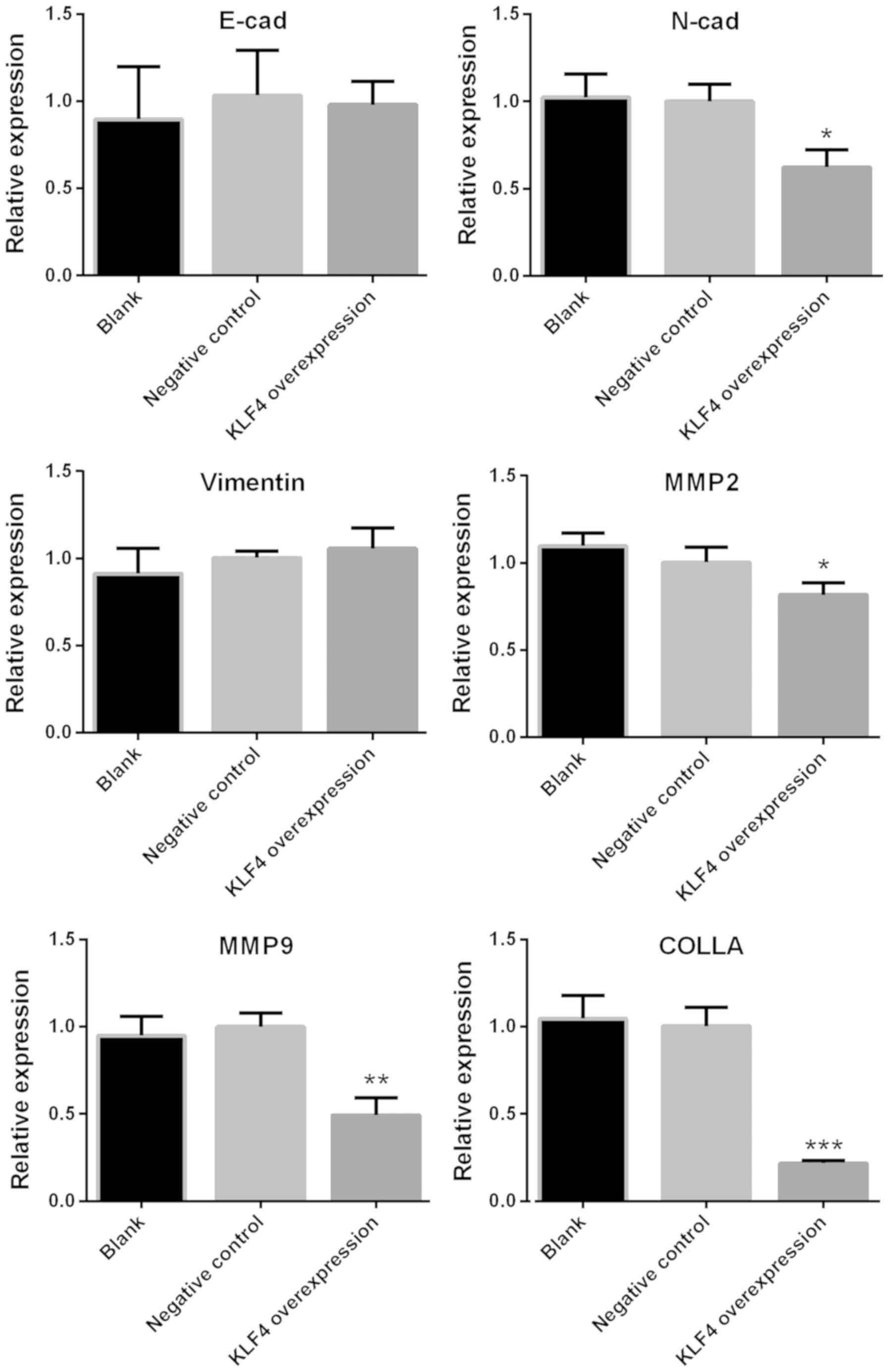 | Figure 4.mRNA expression of N-cadherin, MMP2,
MMP9 and collagen was significantly decreased in the KLF4
overexpression group. mRNA expression of E-cadherin, N-cadherin,
Vimentin, MMP2, MMP9 and collagen in KTC1 cells were detected via
quantitative PCR. Among all genes screened, the expression levels
of N-cadherin, MMP2, MMP9 and collagen were significantly decreased
in the KLF4 overexpression group, as compared with the blank or
negative control groups (mean ± standard deviation; n=3 per group).
*P<0.05, **P<0.01 and ***P<0.001 compared with the control
or negative control group. MMP, matrix metalloproteinase; KLF4,
Kruppel-like factor 4; COLLA, collagen; cad, cadherin. |
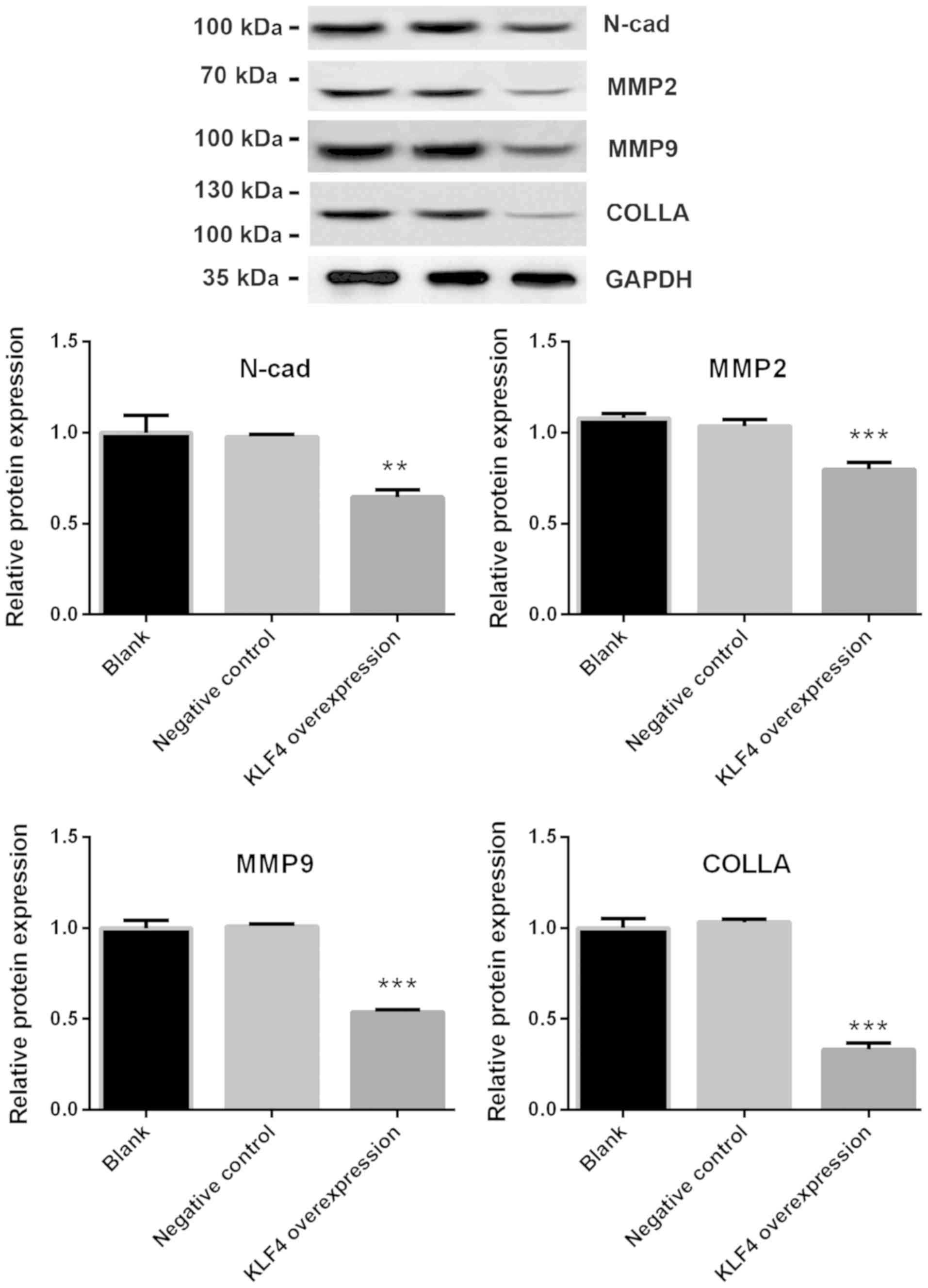 | Figure 5.Protein expression of N-cadherin,
MMP2, MMP9 and collagen was significantly decreased in the KLF4
overexpression group. Protein expression of E-cadherin, N-cadherin,
Vimentin, MMP2, MMP9 and collagen in KTC1 cells were detected via
western blotting. The protein expression of N-cadherin, MMP2, MMP9
and collagen was significantly decreased in the KLF4 overexpression
group compared with the blank or negative control groups (mean ±
standard deviation; n=3 per group). **P<0.01 and ***P<0.001
vs. the control or negative control groups. MMP, matrix
metalloproteinase; KLF4, Kruppel-like factor 4; COLLA, collagen;
N-cad, N-cadherin. |
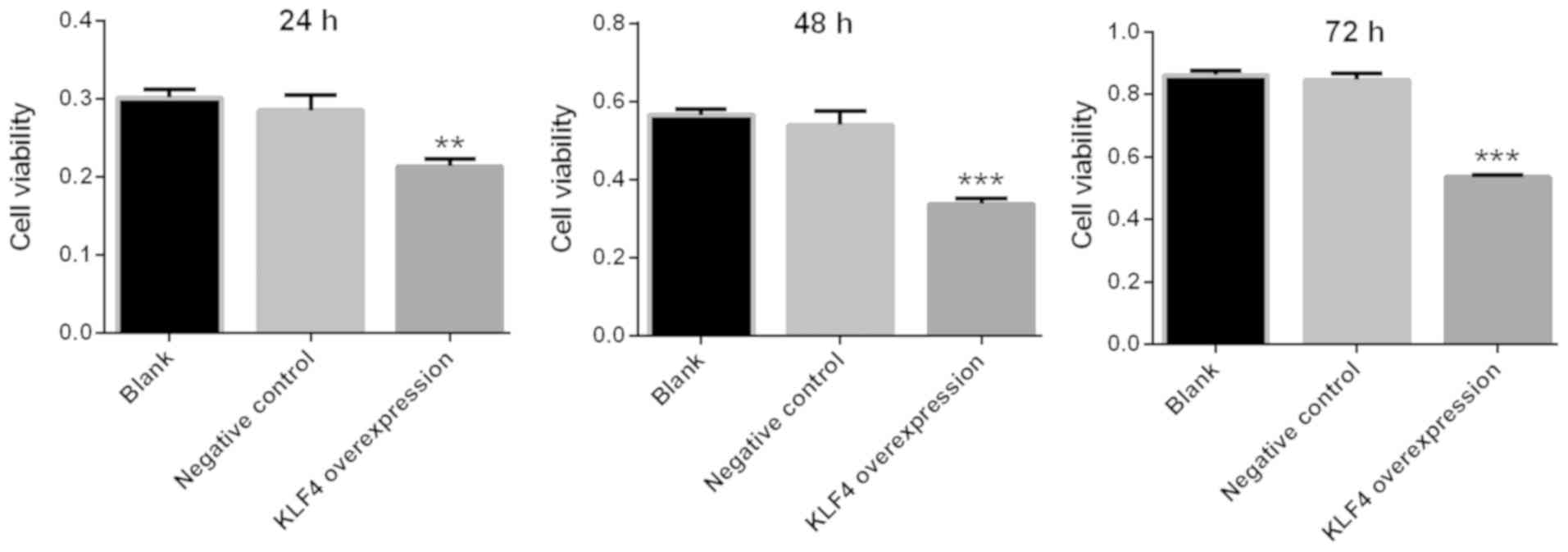
Viability of papillary thyroid cancer
cells is markedly decreased in the KLF4 overexpression group
The viability of KTC1 cells was detected via a CCK-8
assay at 24, 48 and 72 h. The viability of KTC1 cells was markedly
decreased in the KLF4 overexpression group at 24, 48 and 72 h when
compared with the blank or negative control group (P<0.01 for 24
h; P<0.001 for 48 and 72 h; Fig.
6).
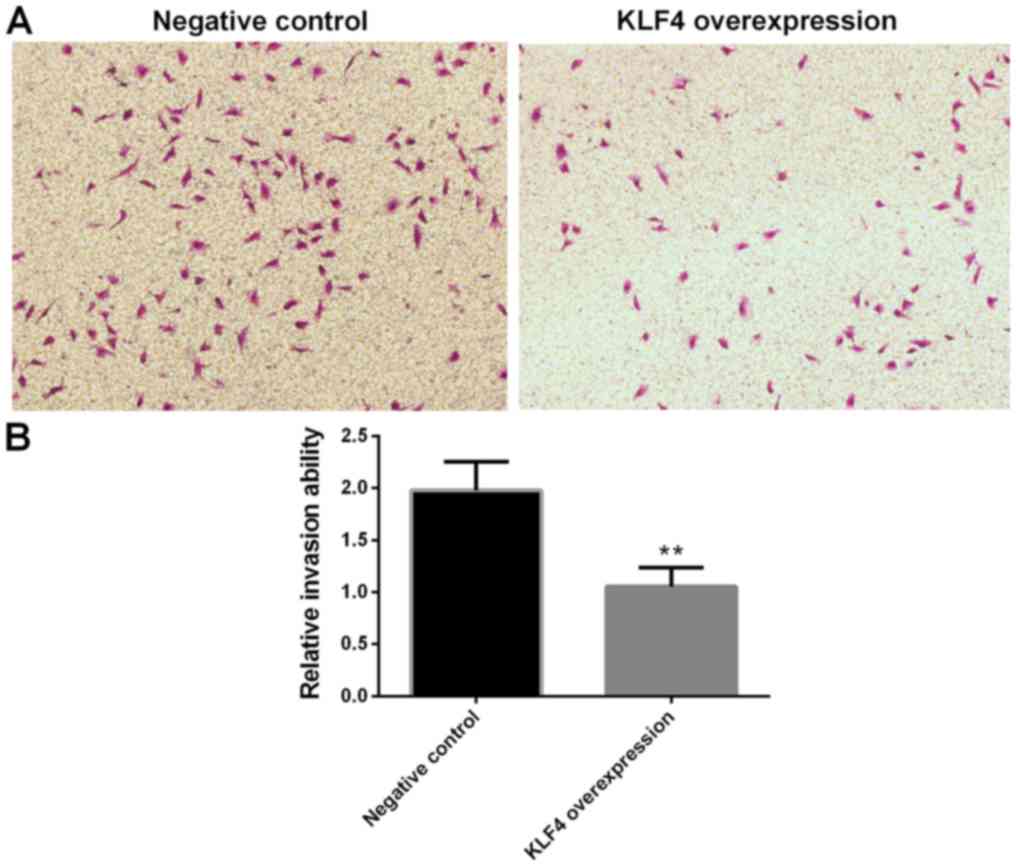
Invasion of papillary thyroid cancer
cells in the KLF4 overexpression group is significantly
decreased
Cell invasion was investigated using a transwell
invasion assay. Compared with the negative control group, the
invasion of KTC1 cells in the KLF4 overexpression group were
significantly decreased (P<0.01; Fig.
7).
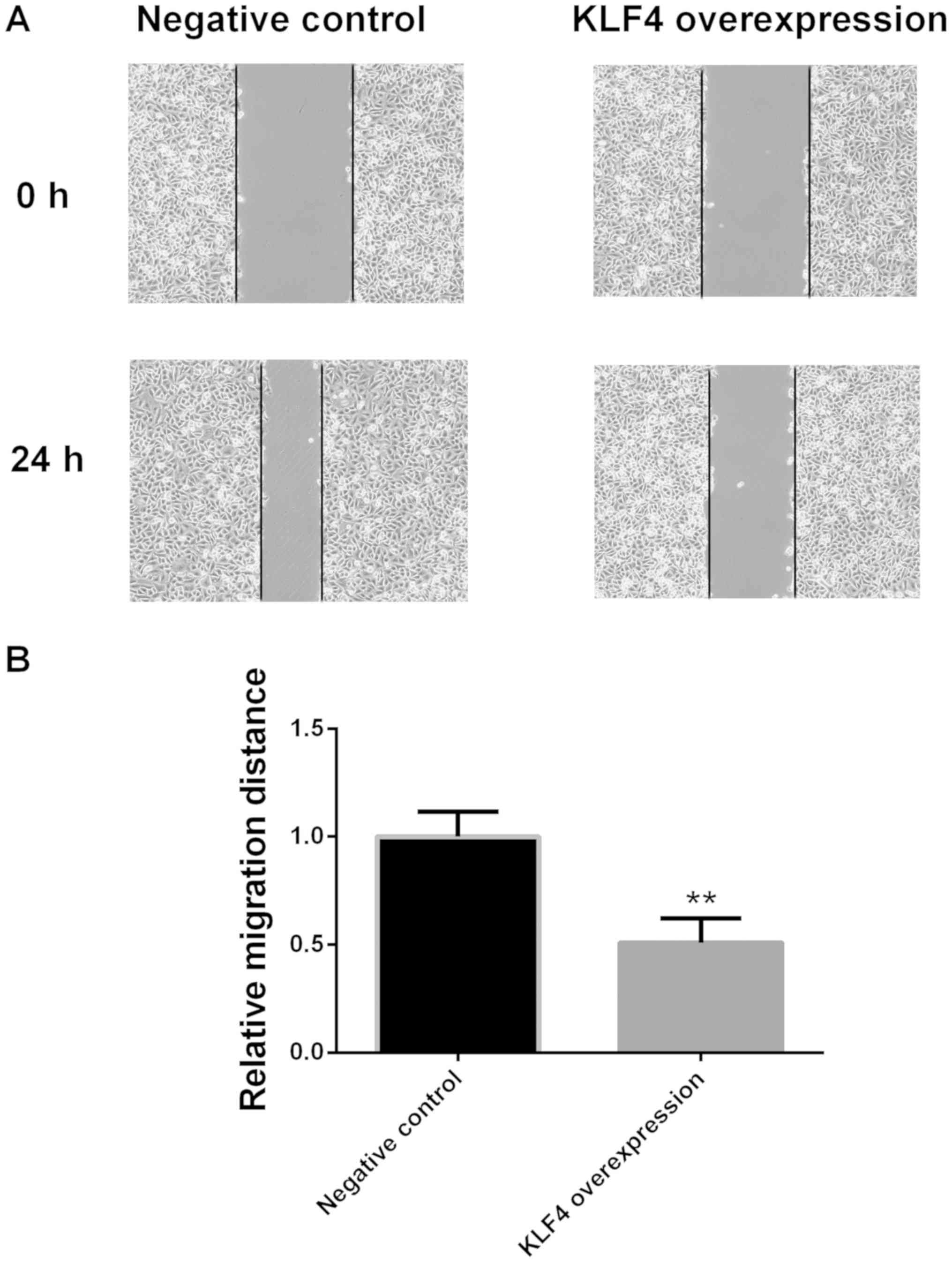
Migration of papillary thyroid cancer
cells in the KLF4 overexpression group is significantly decreased
at 24 h
Cell migration ability was examined via a scratch
migration assay at 0 and 24 h. Compared with the negative control
group, the migration of KTC1 cells in the KLF4 overexpression group
was significantly decreased at 24 h (P<0.01; Fig. 8).
Discussion
The present study demonstrated that the expression
of KLF4 was significantly lower in thyroid tumor tissue compared
with adjacent non-cancerous tissue. The viability, invasion and
migration of cells, and the expression of N-cadherin, MMP2, MMP9
and collagen in papillary thyroid cancer cells were markedly
decreased following KLF4 overexpression.
KLF4 has been reported to inhibit the proliferation
of colorectal cancer cells via NMYC downstream-regulated gene 2
(26). Furthermore, KLF4 suppressed
estrogen-dependent breast cancer growth by inhibiting the
transcriptional activity of the estrogen receptor (27). KLF4 has also been demonstrated to
inhibit the invasion of lung cancer cells by suppressing secreted
protein acidic and cysteine rich gene expression (27). F-box protein-32 suppressed the
tumorigenesis of breast cancer by targeting KLF4 for proteasomal
degradation (28). In addition, the
long non-coding RNA small nucleolar RNA host gene 5/miR-32 axis was
revealed to regulate gastric cancer cell proliferation and
migration by targeting KLF4 (29).
KLF4 and KLF5 regulated the proliferation, apoptosis and invasion
of esophageal cancer cells (30).
Meanwhile, KLF4 was revealed to regulate adult lung
tumor-initiating cells and repress K-Ras-mediated lung cancer
(31). To the best of our knowledge,
the present study revealed for the first time that the expression
of KLF4 was significantly lower in thyroid tumor tissue when
compared with adjacent non-cancerous tissues, and the viability,
tumor invasion and migration of papillary thyroid cancer cells were
significantly decreased following the overexpression of KLF4. These
results may broaden the current understanding of the properties of
KLF4, and shed light on possible therapeutic treatment of papillary
thyroid cancer.
The present study revealed that the expression of
N-cadherin, MMP2, MMP9 and collagen in papillary thyroid cancer
cells were significantly decreased when KLF4 was overexpressed.
N-cadherin expression in breast cancer has been demonstrated to be
associated with an aggressive histological variant of invasive
micropapillary carcinoma (32).
N-cadherin, as a novel prognostic biomarker, was reported to drive
the malignant progression of colorectal cancer (33). N-cadherin expression was also
associated with enhanced invasion in erlotinib-resistant lung
cancer cell lines (34). In
addition, Trop2 has been indicated to enhance the invasion of
thyroid cancer by inducing MMP2 via ERK and Janus kinase pathways
(35). miR-29c suppressed the
adhesion of lung cancer cell to the extracellular matrix, as well
as metastasis, by targeting MMP2 (36). The association between the MMP2-1306
C/T polymorphism and prostate cancer susceptibility was revealed in
a meta-analysis based on 3,906 subjects (37). Meanwhile, the downregulation of
hepatoma-derived growth factor inhibited the migration and invasion
of prostate cancer cells by suppressing MMP2 and MMP9 (38). miR-133b was reported to inhibit the
cell growth, migration and invasion by targeting MMP9 in non-small
cell lung cancer (39). Furthermore,
the selective targeting of collagen IV in the cancer cell
microenvironment was revealed to decrease tumor burden (40). Losartan loaded liposomes improved the
antitumor efficacy of liposomal paclitaxel via the inhibition of
collagen in breast cancer (41). In
the present study, the significantly decreased expression of
N-cadherin, MMP2, MMP9 and collagen in papillary thyroid cancer
cells following KLF4 overexpression may impair the adhesion of
thyroid cancer cells to the extracellular matrix, thus disrupting
tumor invasion and migration.
Other factors may also account for the effects of
KLF4 overexpression on tumor invasion and migration. KLF4 inhibited
tumor growth and metastasis by targeting miR-31 in human
hepatocellular carcinoma (42).
Podocalyxin-like (PODXL) promoted the metastasis of gastric cancer,
whereas KLF4 downregulated PODXL and prevented metastasis (43). miR-543 was also revealed to promote
colorectal cancer proliferation and metastasis by targeting KLF4
(44). In addition, KLF4-mediated
suppression of CD44 signaling decreased the stemness and metastasis
of pancreatic cancer (45). Further
studies are required to elucidate whether the aforementioned
factors modulate the anti-metastasis effects of KLF4 in papillary
thyroid cancer.
There are certain limitations to the present study.
Normal thyroid cell lines were not used and there was no in
vivo study. However, the present study detected the expression
levels of KLF4 in human thyroid tumor tissue and adjacent normal
tissues via IHC and western blotting. The experimental results in
human tissues were consistent; therefore, similar experiments in
normal thyroid cell lines or animals were not performed, which may
or may not reflect the real situations in human.
In conclusion, the present study demonstrated that
the expression of KLF4 was significantly lower in thyroid tumor
tissue. The cell viability, tumor invasion and migration, and
expression levels of N-cadherin, MMP2, MMP9 and collagen in
papillary thyroid cancer cells were markedly decreased with the
overexpression of KLF4. Although further research is required to
elucidate the underlying molecular mechanisms, the present study
may provide the foundations for future therapeutic measures
targeting papillary thyroid cancer.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Baoshan
District Science and Technology Commission Fund (grant no.
16-E-10), and Baoshan District Integrated Traditional Chinese and
Western Medicine Hospital Fund (grant no. 201603).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
QW, LL and JX designed the experiments. QW and YC
carried out the experiments. YC performed the statistical analyses.
LL and JX gave advice on how to design and carry out experiments.
QW wrote the manuscript, which was revised by the other authors.
All authors read and approved the final manuscript.
Ethics approval and consent to
participate
The present study was approved by the Institutional
Research Board of Baoshan District Integrated Traditional Chinese
and Western Medicine Hospital. Each participant provided written
informed consent.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
GBD 2015 Disease and Injury Incidence and
Prevalence Collaborators: Global, regional, and national incidence,
prevalence, and years lived with disability for 310 diseases and
injuries, 1990–2015: A systematic analysis for the Global Burden of
Disease Study 2015. Lancet. 388:1545–1602. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chen J, Hou H, Chen H, Luo Y, He Y, Zhang
L, Zhang Y, Liu H, Zhang F, Liu Y, et al: Identification of
β-hydroxybutyrate as a potential biomarker for female papillary
thyroid cancer. Bioanalysis. 11:461–470. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
James BC, Timsina L, Graham R, Angelos P
and Haggstrom DA: Changes in total thyroidectomy versus thyroid
lobectomy for papillary thyroid cancer during the past 15 years.
Surgery. 166:41–47. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Galuppini F, Pennelli G and Rugge M: The
rising incidence of papillary thyroid cancer: More cancers or more
assessments? Indian J Cancer. 56:183–184. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kazaure HS, Roman SA and Sosa JA:
Aggressive variants of papillary thyroid cancer: Incidence,
characteristics and predictors of survival among 43,738 patients.
Ann Surg Oncol. 19:1874–1880. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Richards ML: Thyroid cancer genetics:
Multiple endocrine neoplasia type 2, non-medullary familial thyroid
cancer, and familial syndromes associated with thyroid cancer. Surg
Oncol Clin N Am. 1839–52. (viii)2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zhu M, Zhang N and He S: Transcription
factor KLF4 modulates microRNA-106a that targets Smad7 in gastric
cancer. Pathol Res Pract. 1524672019. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhuan B, Lu Y, Chen Q, Zhao X, Li P, Yuan
Q and Yang Z: Overexpression of the long noncoding RNA TRHDE-AS1
inhibits the progression of lung cancer via the miRNA-103/KLF4
axis. J Cell Biochem. May 22–2019.(Epub ahead of print). doi:
10.1002/jcb.29029. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yang VW, Liu Y, Kim J, Shroyer KR and
Bialkowska AB: Increased genetic instability and accelerated
progression of colitis-associated colorectal cancer through
intestinal epithelium-specific deletion of Klf4. Mol Cancer Res.
17:165–176. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Okuda H, Xing F, Pandey PR, Sharma S,
Watabe M, Pai SK, Mo YY, Iiizumi-Gairani M, Hirota S, Liu Y, et al:
miR-7 suppresses brain metastasis of breast cancer stem-like cells
by modulating KLF4. Cancer Res. 73:1434–1444. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Li H, Wang J, Xiao W, Xia D, Lang B, Wang
T, Guo X, Hu Z, Ye Z and Xu H: Epigenetic inactivation of KLF4 is
associated with urothelial cancer progression and early recurrence.
J Urol. 191:493–501. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Guo K, Cui J, Quan M, Xie D, Jia Z, Wei D,
Wang L, Gao Y, Ma Q and Xie K: The novel KLF4/MSI2 signaling
pathway regulates growth and metastasis of pancreatic cancer. Clin
Cancer Res. 23:687–696. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Akaogi K, Nakajima Y, Ito I, Kawasaki S,
Oie SH, Murayama A, Kimura K and Yanagisawa J: KLF4 suppresses
estrogen-dependent breast cancer growth by inhibiting the
transcriptional activity of ERalpha. Oncogene. 28:2894–2902. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ding X, Zhong T, Jiang L, Huang J, Xia Y
and Hu R: miR-25 enhances cell migration and invasion in
non-small-cell lung cancer cells via ERK signaling pathway by
inhibiting KLF4. Mol Med Rep. 17:7005–7016. 2018.PubMed/NCBI
|
|
15
|
Cittelly DM, Finlay-Schultz J, Howe EN,
Spoelstra NS, Axlund SD, Hendricks P, Jacobsen BM, Sartorius CA and
Richer JK: Progestin suppression of miR-29 potentiates
dedifferentiation of breast cancer cells via KLF4. Oncogene.
32:2555–2564. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wong AS and Gumbiner BM:
Adhesion-independent mechanism for suppression of tumor cell
invasion by E-cadherin. J Cell Biol. 161:1191–1203. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Makrigiannakis A, Coukos G, Blaschuk O and
Coutifaris C: Follicular atresia and luteolysis. Evidence of a role
for N-cadherin. Ann N Y Acad Sci. 900:46–55. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yamashita N, Tokunaga E, Iimori M, Inoue
Y, Tanaka K, Kitao H, Saeki H, Oki E and Maehara Y: Epithelial
paradox: Clinical significance of coexpression of E-cadherin and
vimentin with regard to invasion and metastasis of breast cancer.
Clin Breast Cancer. 18:e1003–e1009. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Winerdal ME, Krantz D, Hartana CA,
Zirakzadeh AA, Linton L, Bergman EA, Rosenblatt R, Vasko J,
Alamdari F, Hansson J, et al: Urinary bladder cancer Tregs suppress
MMP2 and potentially regulate invasiveness. Cancer Immunol Res.
6:528–538. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Klassen LMB, Chequin A, Manica GCM,
Biembengut IV, Toledo MB, Baura VA, de O Pedrosa F, Ramos EAS,
Costa FF, de Souza EM and Klassen G: MMP9 gene expression
regulation by intragenic epigenetic modifications in breast cancer.
Gene. 642:461–466. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Nielsen SH, Willumsen N, Brix S, Sun S,
Manon-Jensen T, Karsdal M and Genovese F: Tumstatin, a matrikine
derived from collagen type IVα3, is elevated in serum from patients
with non-small cell lung cancer. Transl Oncol. 11:528–534. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Fujimori M and Sakauchi G: TNM
classification-thyroid cancer. Gan No Rinsho. 13:320–322. 1967.(In
Japanese). PubMed/NCBI
|
|
23
|
Haugen BR, Alexander EK, Bible KC, Doherty
GM, Mandel SJ, Nikiforov YE, Pacini F, Randolph GW, Sawka AM,
Schlumberger M, et al: 2015 American thyroid association management
guidelines for adult patients with thyroid nodules and
differentiated thyroid cancer. Thyroid. 26:1–33. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Mancini P, Bonanno Ferraro G, Iaconelli M,
Suffredini E, Valdazo-González B, Della Libera S, Divizia M and La
Rosa G: Molecular characterization of human Sapovirus in untreated
sewage in Italy by amplicon-based Sanger and next-generation
sequencing. J Appl Microbiol. 126:324–331. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ma Y, Wu L, Liu X, Xu Y, Shi W, Liang Y,
Yao L, Zheng J and Zhang J: KLF4 inhibits colorectal cancer cell
proliferation dependent on NDRG2 signaling. Oncol Rep. 38:975–984.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhou Y, Hofstetter WL, He Y, Hu W, Pataer
A, Wang L, Wang J, Zhou Y, Yu L, Fang B and Swisher SG: KLF4
inhibition of lung cancer cell invasion by suppression of SPARC
expression. Cancer Biol Ther. 9:507–513. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhou H, Liu Y, Zhu R, Ding F, Wan Y, Li Y
and Liu Z: FBXO32 suppresses breast cancer tumorigenesis through
targeting KLF4 to proteasomal degradation. Oncogene. 36:3312–3321.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zhao L, Han T, Li Y, Sun J, Zhang S, Liu
Y, Shan B, Zheng D and Shi J: The lncRNA SNHG5/miR-32 axis
regulates gastric cancer cell proliferation and migration by
targeting KLF4. FASEB J. 31:893–903. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Yang Y, Goldstein BG, Chao HH and Katz JP:
KLF4 and KLF5 regulate proliferation, apoptosis and invasion in
esophageal cancer cells. Cancer Biol Ther. 4:1216–1221. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Yu T, Chen X, Zhang W, Liu J, Avdiushko R,
Napier DL, Liu AX, Neltner JM, Wang C, Cohen D and Liu C: KLF4
regulates adult lung tumor-initiating cells and represses
K-Ras-mediated lung cancer. Cell Death Differ. 23:207–215. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Nagi C, Guttman M, Jaffer S, Qiao R, Keren
R, Triana A, Li M, Godbold J, Bleiweiss IJ and Hazan RB: N-cadherin
expression in breast cancer: Correlation with an aggressive
histologic variant-invasive micropapillary carcinoma. Breast Cancer
Res Treat. 94:225–235. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Yan X, Yan L, Liu S, Shan Z, Tian Y and
Jin Z: N-cadherin, a novel prognostic biomarker, drives malignant
progression of colorectal cancer. Mol Med Rep. 12:2999–3006. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Zhang X, Liu G, Kang Y, Dong Z, Qian Q and
Ma X: N-cadherin expression is associated with acquisition of EMT
phenotype and with enhanced invasion in erlotinib-resistant lung
cancer cell lines. PLoS One. 8:e576922013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Guan H, Guo Z, Liang W, Li H, Wei G, Xu L,
Xiao H and Li Y: Trop2 enhances invasion of thyroid cancer by
inducing MMP2 through ERK and JNK pathways. BMC Cancer. 17:4862017.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Wang H, Zhu Y, Zhao M, Wu C, Zhang P, Tang
L, Zhang H, Chen X, Yang Y and Liu G: miRNA-29c suppresses lung
cancer cell adhesion to extracellular matrix and metastasis by
targeting integrin β1 and matrix metalloproteinase 2 (MMP2). PLoS
One. 8:e701922013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zhang K, Chen X, Zhou J, Yang C, Zhang M,
Chao M, Zhang L and Liang C: Association between MMP2-1306 C/T
polymorphism and prostate cancer susceptibility: A meta-analysis
based on 3906 subjects. Oncotarget. 8:45020–45029. 2017.PubMed/NCBI
|
|
38
|
Yang F, Yu N, Wang H, Zhang C, Zhang Z, Li
Y, Li D, Yan L, Liu H and Xu Z: Downregulated expression of
hepatoma-derived growth factor inhibits migration and invasion of
prostate cancer cells by suppressing epithelial-mesenchymal
transition and MMP2, MMP9. PLoS One. 13:e01907252018. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Zhen Y, Liu J, Huang Y, Wang Y, Li W and
Wu J: miR-133b inhibits cell growth, migration, and invasion by
targeting MMP9 in non-small cell lung cancer. Oncol Res.
25:1109–1116. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Revert F, Revert-Ros F, Blasco R, Artigot
A, López-Pascual E, Gozalbo-Rovira R, Ventura I,
Gutiérrez-Carbonell E, Roda N, Ruíz-Sanchis D, et al: Selective
targeting of collagen IV in the cancer cell microenvironment
reduces tumor burden. Oncotarget. 9:11020–11045. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Xia T, He Q, Shi K, Wang Y, Yu Q, Zhang L,
Zhang Q, Gao H, Ma L and Liu J: Losartan loaded liposomes improve
the antitumor efficacy of liposomal paclitaxel modified with pH
sensitive peptides by inhibition of collagen in breast cancer.
Pharm Dev Technol. 23:13–21. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Tian C, Yao S, Liu L, Ding Y, Ye Q, Dong
X, Gao Y, Yang N and Li Q: Klf4 inhibits tumor growth and
metastasis by targeting microRNA-31 in human hepatocellular
carcinoma. Int J Mol Med. 39:47–56. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Zhang J, Zhu Z, Wu H, Yu Z, Rong Z, Luo Z,
Xu Y, Huang K, Qiu Z and Huang C: PODXL, negatively regulated by
KLF4, promotes the EMT and metastasis and serves as a novel
prognostic indicator of gastric cancer. Gastric Cancer. 22:48–59.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Zhai F, Cao C, Zhang L and Zhang J:
miR-543 promotes colorectal cancer proliferation and metastasis by
targeting KLF4. Oncotarget. 8:59246–59256. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Yan Y, Li Z, Kong X, Jia Z, Zuo X, Gagea
M, Huang S, Wei D and Xie K: KLF4-mediated suppression of CD44
signaling negatively impacts pancreatic cancer stemness and
metastasis. Cancer Res. 76:2419–2431. 2016. View Article : Google Scholar : PubMed/NCBI
|