Introduction
Tongue squamous cell carcinoma (TSCC) is the most
common type of oral malignancy, accounting for 40–50% of all cases
(1). TSCC exhibits the
characteristics of high malignancy, rapid growth and marked levels
of infiltration; furthermore, the tongue has abundant lymphatic
vessels and blood circulation within its structure, which often
results in TSCC progression to lymph node metastasis at earlier
stages, and it seriously affects the voice, chewing and swallowing
ability of the patients (2–4). At present, treatment of TSCC primarily
relies on surgery, and treatment via radiotherapy and chemotherapy
is auxiliary. However, the 5-year survival rate of patients is only
32–54% following comprehensive treatment (5,6).
Therefore, identifying effective treatment methods is essential to
improve survival rates for these patients.
Current research regarding TSCC focuses on microRNAs
(miRNAs), and the function of miRNAs has preliminarily been
identified. miRNA are small non-coding RNA molecules measuring
18–23 nucleotides in length. Although miRNAs have a small molecular
weight, they serve a role in the transcriptional level of human
cells and regulates numerous important biological functions of both
plant and animal organisms, and in human tumors (7,8).
mi-21-5p is one of the most important members of the
miRNA family, and it is closely associated with the occurrence and
development of cancer. miRNA-21-5p, located on chromosome 17q23.1,
has been identified in numerous different types of tumors, which
indicates that it may exhibit enhanced expression levels that
correspond with disease development. Jiang et al (9) revealed that miRNA-21-5p was upregulated
in gastric cancer tissues and SGC-7901 cells, and that the
knockdown of miRNA-21-5p suppressed cell proliferation, migration
and invasion, and the inflammatory response. The identification of
miRNAs and their expression profiles among different diseases
indicates that miRNA-21 may serve as a potential biomarker
(9). However, the role of miR-21-5p
in TSCC and its associated underlying molecular mechanisms have not
yet been reported.
In the present study, the expression levels of
miR-21-5p in TSCC were investigated, and the apoptotic effect of
miRNA-21-5p on human TSCC Cal 27 and SCC9 cell lines was examined.
The results revealed that the PI3K/AKT signaling pathway serves a
role in the underlying molecular mechanism of the disease.
Materials and methods
Patients and tissue samples
In total, 40 tumor tissue samples were obtained from
patients with TSCC who had been admitted to the Department of Oral
and Maxillofacial Surgery, Second Affiliated Hospital of Jinzhou
Medical University (Jinzhou, China) between January 2017 and June
2018, including 24 males and 16 females, aged 38–76 years, with a
median age of 54 years. None of the patients received chemotherapy
or radiotherapy. In addition, 40 cases of normal tissues (adjacent
non-cancerous tissues) were obtained from the Second Affiliated
Hospital of Jinzhou Medical University. The inclusion criteria were
as follows: All patients were diagnosed with TSCC via pathology,
and no radiation therapy or chemotherapy was performed prior to
biopsy. The exclusion criteria were as follows: Patients with one
or more of the following conditions were excluded: i) Infectious
disease; ii) acute cardiovascular and cerebrovascular diseases;
iii) rheumatic disease; iv) diabetes; or v) other tumors.
The present study was approved by the Ethical
Committee of Jinzhou Medical University on October 26, 2016
(approval no. JZH2016052). Written informed consent was obtained
from all patients included in the present study.
Hematoxylin and eosin (H&E)
staining
TSCC tissues were fixed (>24 h at room
temperature) in 4% paraformaldehyde and embedded in paraffin.
Paraffin-embedded samples were then sliced into 4 µm sections and
resected specimens were dewaxed in xylene, washed in distilled
water and stained with hematoxylin and eosin at room temperature
for 5 min. Pathological alterations of myocardial tissue were
observed under a light microscope (magnification, ×200).
Cell culture
TSCC Cal 27 and SCC9 cell lines were purchased from
The Cell Bank of Type Culture Collection of Chinese Academy of
Sciences. The Cal 27 cell line was cultured with Dulbecco's
modified Eagle's medium (DMEM; Gibco; Thermo Fisher Scientific,
Inc.) containing 10% fetal bovine serum (FBS; Gibco; Thermo Fisher
Scientific, Inc.) in a 5% CO2 incubator at 37°C and
saturated humidity. The SCC9 cell line was incubated with RPMI-1640
(Gibco; Thermo Fisher Scientific, Inc.) containing 10% FBS in a 5%
CO2 incubator at 37°C.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted from the tissues or cell
lines using TRIzol® reagent according to manufacturer's
protocol. The cDNA was transcribed using a Prime Script™ RT Master
Mixture according to the manufacturer's protocol (Takara
Biotechnology Co., Ltd.). miR-21-5p in TSCC was detected using SYBR
Prime Script miRNA RT-PCR kit (Takara Biotechnology Co., Ltd.). The
thermocycling conditions were as follows: Pre-denaturation at 95°C
for 1 min, followed by denaturation at 95°C for 15 sec, annealing
at 60°C for 40 sec and extension at 72°C for 15 sec, for a total of
40 cycles. The primer sequences in the present study were as
follows: hsa-miR-21-5p forward, 5′-GGGGTAGCTTATCAGACTGATG-3′;
hsa-miR-21-5p reverse, 5′-TGTCGTGGAGCGGCAATTG-3′; U6: Forward,
5′-CGCTTCGGCACATATACTA-3′; U6 reverse,
5′-CGCTTCACGAATTTGCGTGTCA-3′; PDCD4 forward,
5′-TGTGCCAACCAGTCCA-3′; PDCD4 reverse, 5′-GATCCTAACTATGATGA-3′;
GAPDH forward, 5′-TGTTGCCATCAATGACCCCTT-3′; GAPDH reverse,
5′-CTCCACGACGTACTCAGCG-3′. The expression levels of miRNA-21-5p
were calculated using the 2−ΔΔCq method (10).
Transfection
miR-21-5p inhibitors were synthesized along with a
corresponding negative control by Shanghai GenePharma Co., Ltd.
Plasmid production and purification were performed by Shanghai
GenePharma Co., Ltd. miR-21-5p inhibitor sequences (forward,
5′-UAGCUUAUCAGACUGAUGUUGA-3′ and reverse,
5′-TCAACATCAGTCTGATAAGCTA-3′) were cloned into the lentivirus
without green fluorescence (Shanghai GeneChem Co., Ltd.). Polybrene
(6 µg/ml; Shanghai GeneChem Co., Ltd.) and an appropriate dose of
lentivirus (1×108) were added and incubated at 37°C for
24 h. Cells transfected with lentivirus were screened with
puromycin to increase transfection efficiency. The transfection
efficiency was additionally identified via PCR.
Cell Counting Kit-8 (CCK-8)
Next, 2,000 cells per well were seeded in the
96-well plate. Cell proliferation was detected using a CCK-8 assay
(MedChemExpress) according to the manufacturer's protocol. The
reproductive ability of cells was measured at 450 mm using a
microplate reader.
Transwell assay
SCC-9 and Cal 27 cells were re-suspended without
serum at a concentration of 1×105 cells/ml, and were
then (200 µl) seeded into the upper well of Matrigel-coated (Sigma
Aldrich; Merck KGaA) 8 µm pore Transwell inserts (Sigma Aldrich;
Merck KGaA). DMEM (or RPMI-1640; Gibco; Thermo Fisher Scientific,
Inc.) was added to the lower chamber containing 10% FBS. After 24 h
of incubation at 37°C, cells in the chamber were removed with a
cotton swab. Following fixed staining with 0.1% trypan blue for 20
min (Sigma Aldrich; Merck KGaA) at room temperature, images were
captured at randomly selected fields and cells were counted under a
light microscope (magnification, ×200).
Western blot analysis
Total protein was extracted from the cells using
radioimmunoprecipitation assay lysis buffer (Thermo Fisher
Scientific, Inc.). A BCA protein assay kit was used to quantify the
total protein. The proteins (30 µg) were separated via 10%
SDS-PAGE. The gels were transferred to polyvinylidene difluoride
(PVDF) membranes, which were blocked with 5% skimmed milk at room
temperature for 2 h and incubated with anti-Bax (1:1,000; cat. no.
ab32503), anti-Bcl2 (1:1,000; cat. no. ab182858), anti-PI3K
(1:1,000, ab127617), anti-AKT (1:1,000; cat. no. ab179463),
anti-phosphorylated Forkhead Box O1 (p-FOXO1; 1:800; cat. no.
ab52857) and anti-GAPDH (1:2,000; cat. no. ab181602) antibodies
(all from Abcam) overnight at 4°C. Following washing with PBS 3
times, the membranes were incubated with a secondary polyclonal
peroxidase-labeled antibody (Goat Anti-Rabbit IgG; 1:4,000; Abcam;
cat. no. ab205718) for 2 h, and detected using enhanced
chemiluminescence (Thermo Fisher Scientific, Inc.). Quantification
of the bands was performed using ImageJ software (Version d1.47;
National Institutes of Health).
Propidium iodide (PI) and Annexin V
staining
SCC9 and Cal 27 cells were collected, washed and
resuspended. Following the addition of 5 µl Annexin V (BD
Pharmingen; BD Biosciences) and 5 µl PI (BD Pharmingen; BD
Biosciences), cells were incubated at room temperature for 20 min
in the dark, washed with PBS and re-suspended with 300 ml of PBS.
Cell apoptosis rate was calculated using Flow Jo software (v10.1.1
FlowJo LLC).
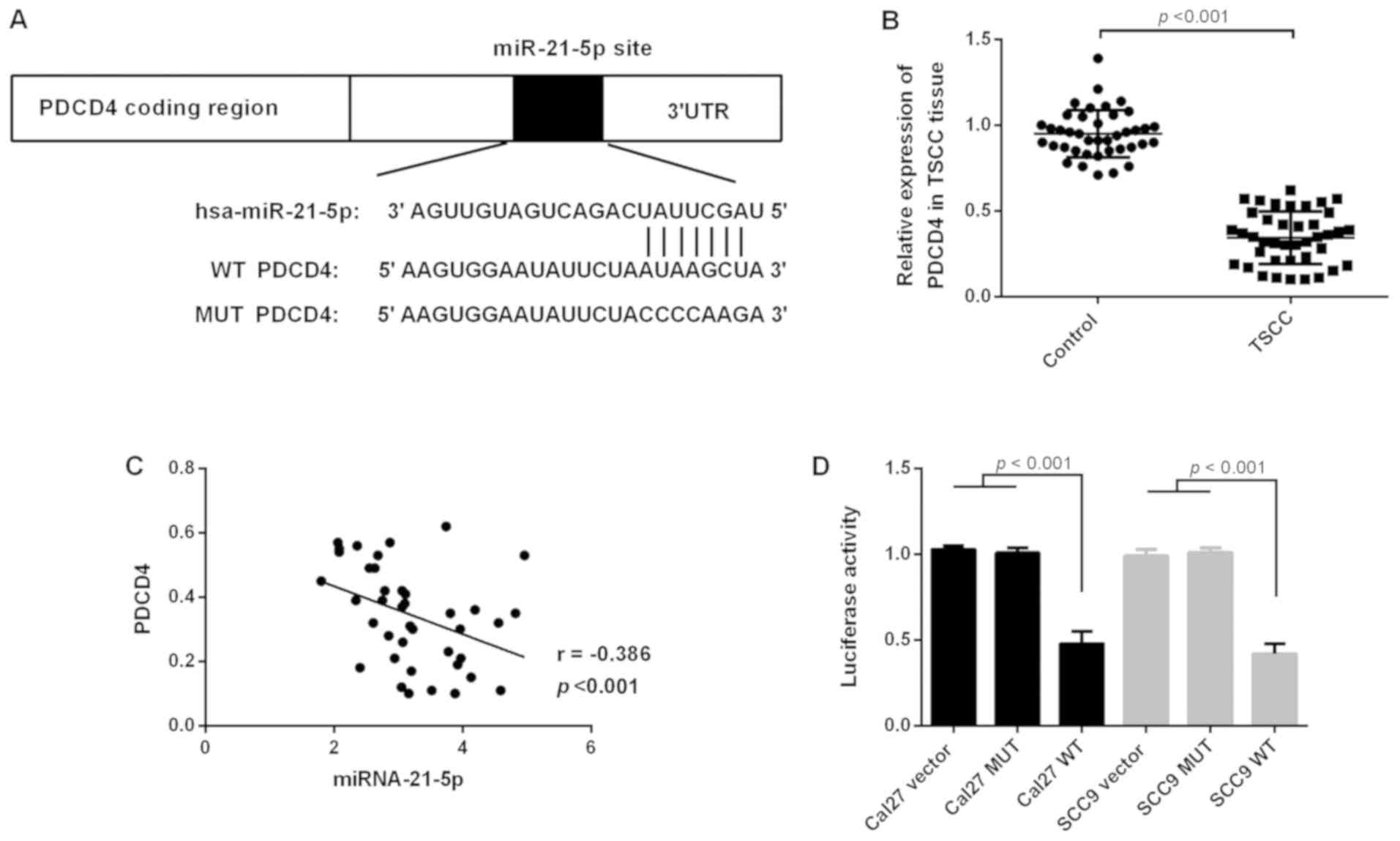
Target gene PDCD4 of miR-21-5p
predicted by TargetScan
Targets of miR-21-5p were searched on TargetScan
(www.targetscan.org/vert_71/)
(11), and the results were further
confirmed by PicTar29 (pictar.mdc-berlin.de/) and microRNA.org 30 (www.microrna.org) (12), suggesting PDCD4 is a potential target
of miR-21-5p.
Luciferase reporter assay
The PDCD4 gene 3′-untranslated region (3′-UTR)
sequence was cloned into a pMIR-REPORT vector (Ambion; Thermo
Fisher Scientific, Inc.). Luciferase reporter plasmids of wild-type
(WT)-PDCD4 mRNA and mutant (Mu)-PDCD4 mRNA were constructed using
the clones. The cells were cultured for 24 h, while the
PDCD4-UTR-pMIR plasmid and miR-21-5p inhibitors or
Mu-PDCD4-UTR-pMIR plasmid and miR-21-5p inhibitors were
co-transfected into cells using Lipofectamine™ 2000
transfection reagent (Thermo Fisher Scientific, Inc.) according to
the manufacturer's protocol. The relative luciferase activity of
PDCD4 was detected using a Dual-Luciferase Reporter assay after
transfection for 48 h. The Renilla luciferase values were
normalized to that of firefly luciferase.
Statistical analysis
All data were presented as the mean ± standard
deviation. Statistical analysis was performed using GraphPad Prism
(v6.0; GraphPad Software Inc.). The data were assessed using the
unpaired two-tailed Student's t-test for comparisons between two
groups, or one-way analysis of variance followed by Kruskal-Wallis
test and Dunn's multiple comparison post-hoc test for comparisons
between >2 groups. Correlations between the miR-21-5p and PDCD4
gene were evaluated using Spearman's coefficient of correlation.
P<0.05 was considered to indicate a statistically significant
difference.
Results
Clinical characteristics of patients
with TSCC
The clinical characteristics of the patients with
TSCC are presented in Table I.
According to the Tumor-Node-Metastasis classification for TSCC
(Union for International Cancer Control, UICC 2010, 7th edition)
(13), the patients were divided
into 4 stages: Stage I (n=4), stage II (n=12), stage III (n=17) and
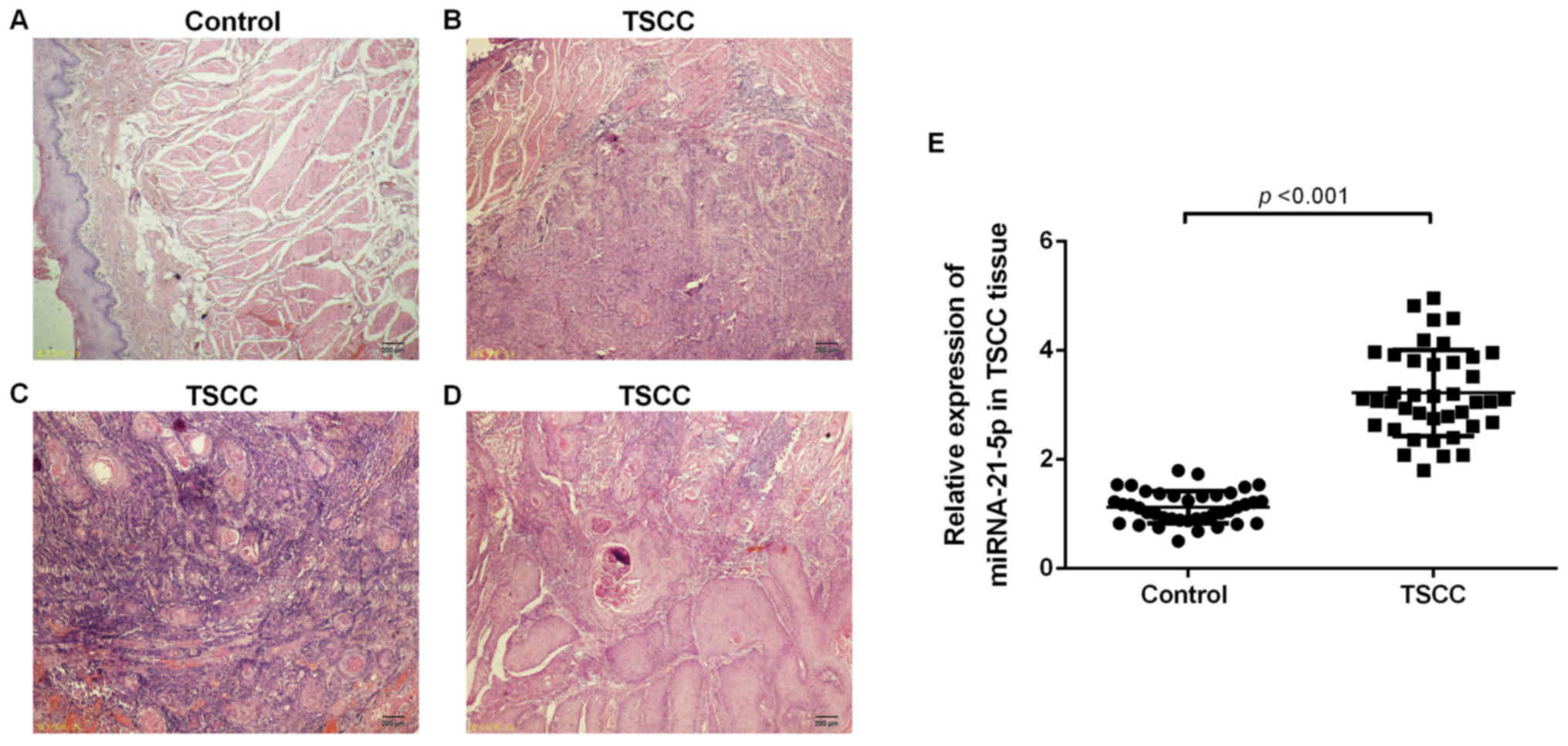
stage IV (n=7). Hematoxylin and eosin staining demonstrated that
all patients had squamous cell carcinoma (Fig. 1A-D).
 | Table I.Characteristic features of study
subjects. |
Table I.
Characteristic features of study
subjects.
| Characteristics | Patients with
TSCC | Non-cancerous
tissues |
|---|
| Age, years |
|
|
|
Range | 38–76 | 35–71 |
| Mean ±
SD | 57±9.25 | 59±11.38 |
| Smoking | 21 | 20 |
| Non-smoking | 19 | 20 |
| Drinking | 15 | 20 |
| Non-drinking | 25 | 20 |
| Local
stimulation | 5 | – |
| Residual
roots and crowns of teeth | 3 |
|
| Bad
prosthesis | 2 |
|
| Tumor location |
| – |
| Lingual
margin | 26 |
|
| Lingual
root | 8 |
|
| Ventral
of tongue | 6 |
|
| Tumor size |
| – |
| T1 | 4 |
|
| T2 | 21 |
|
| T3 | 10 |
|
| T4 | 5 |
|
| Lymph node
involvement |
| – |
| N0 | 23 |
|
| N+ | 17 |
|
| Pathological
classification |
| – |
|
Squamous cell carcinoma | 40 |
|
| Histological
classification |
| – |
| Well
differentiated | 11 |
|
|
Moderately differentiated | 24 |
|
| Poorly
differentiated | 5 |
|
| Clinical stage |
| – |
| I | 4 |
|
| II | 12 |
|
|
III | 17 |
|
| IV | 7 |
|
miR-21-5p is upregulated in TSCC
tissues
miR-21-5p was detected in TSCC tissues via RT-qPCR.
As presented in Fig. 1B, the
expression levels of miR-21-5p were significantly upregulated in
TSCC tissue when compared with that of the control group (Fig. 1E).
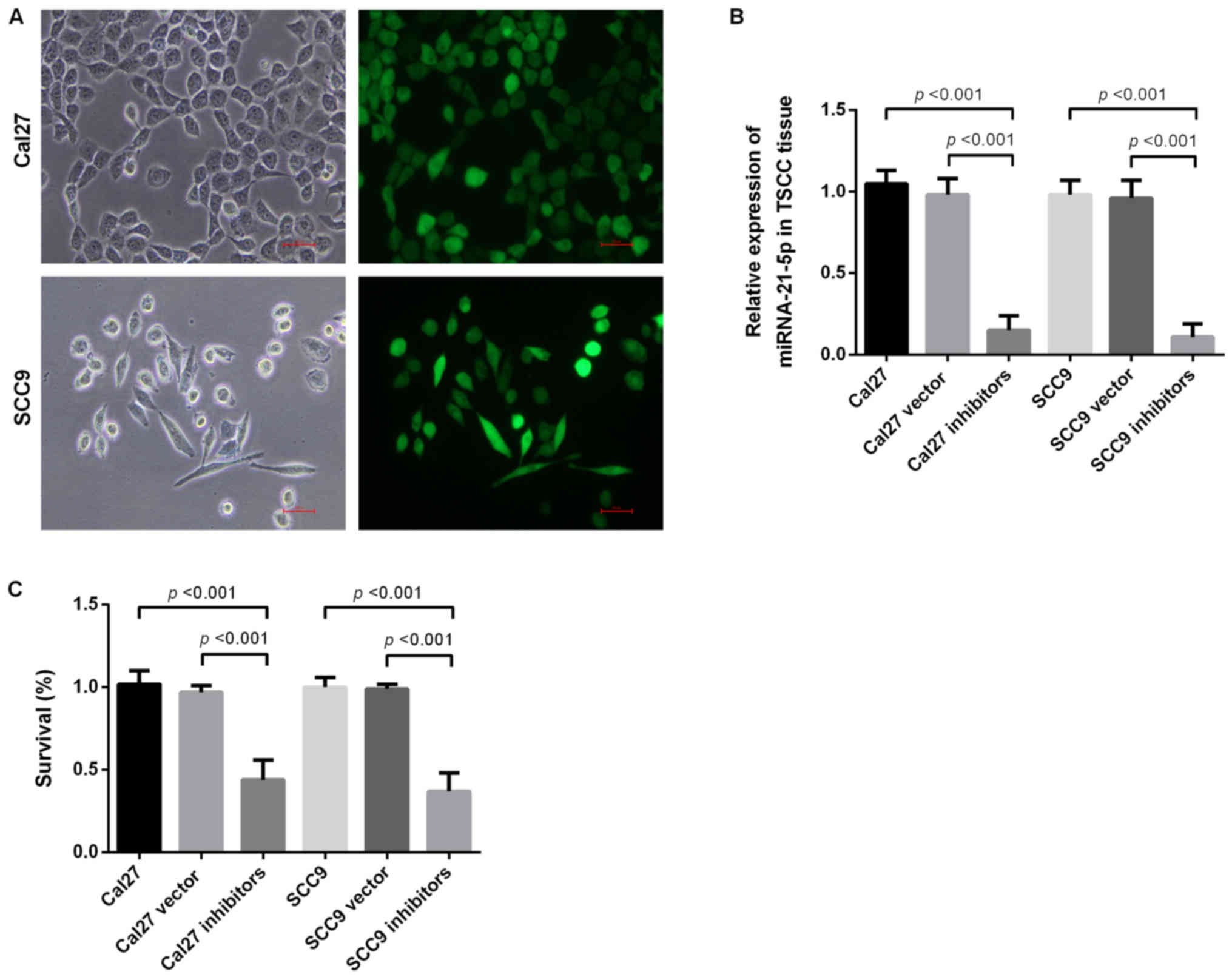
Inhibition of miR-21-5p suppresses
proliferation of Cal 27 cells and SCC9 cells
To determine the function of miR-21-5p, miR-21-5p
was knocked down in the Cal 27 and SCC9 cell lines. Following
miR-21-5p knockdown, the expression levels of miR-21-5p in Cal 27
and SCC9 cells were significantly decreased when assessed via
RT-qPCR (Fig. 2A and B). The CCK-8
assay subsequently determined the proliferation ability of Cal 27
and SCC9 cells following miR-21-5p knockdown. The results revealed
that miR-21-5p knockdown markedly inhibited the proliferation
levels of Cal 27 and SCC9 cells (Fig.
2C).
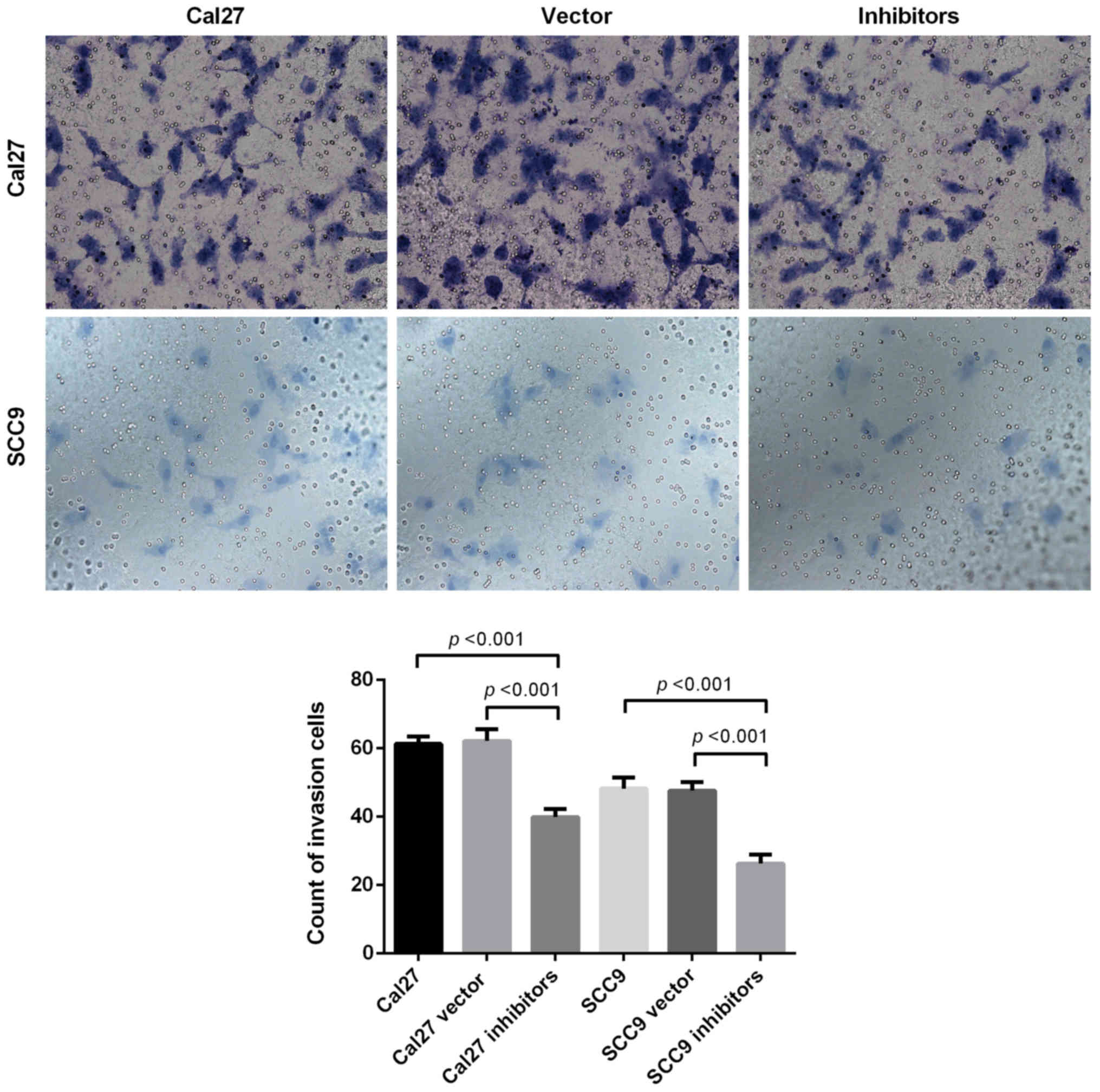
Inhibition of miR-21-5p suppresses the
invasion ability of Cal 27 and SCC9 cells
A Transwell assay was used to detect the invasive
capabilities of Cal 27 and SCC9 cells. The results revealed that
following miR-21-5p knockdown, the invasive capabilities of the Cal
27 and SCC9 cells decreased. The invasion assay demonstrated that
the downregulation of miR-21-5p significantly inhibited the
invasion capacity of Cal 27 and SCC9 cells (Fig. 3).
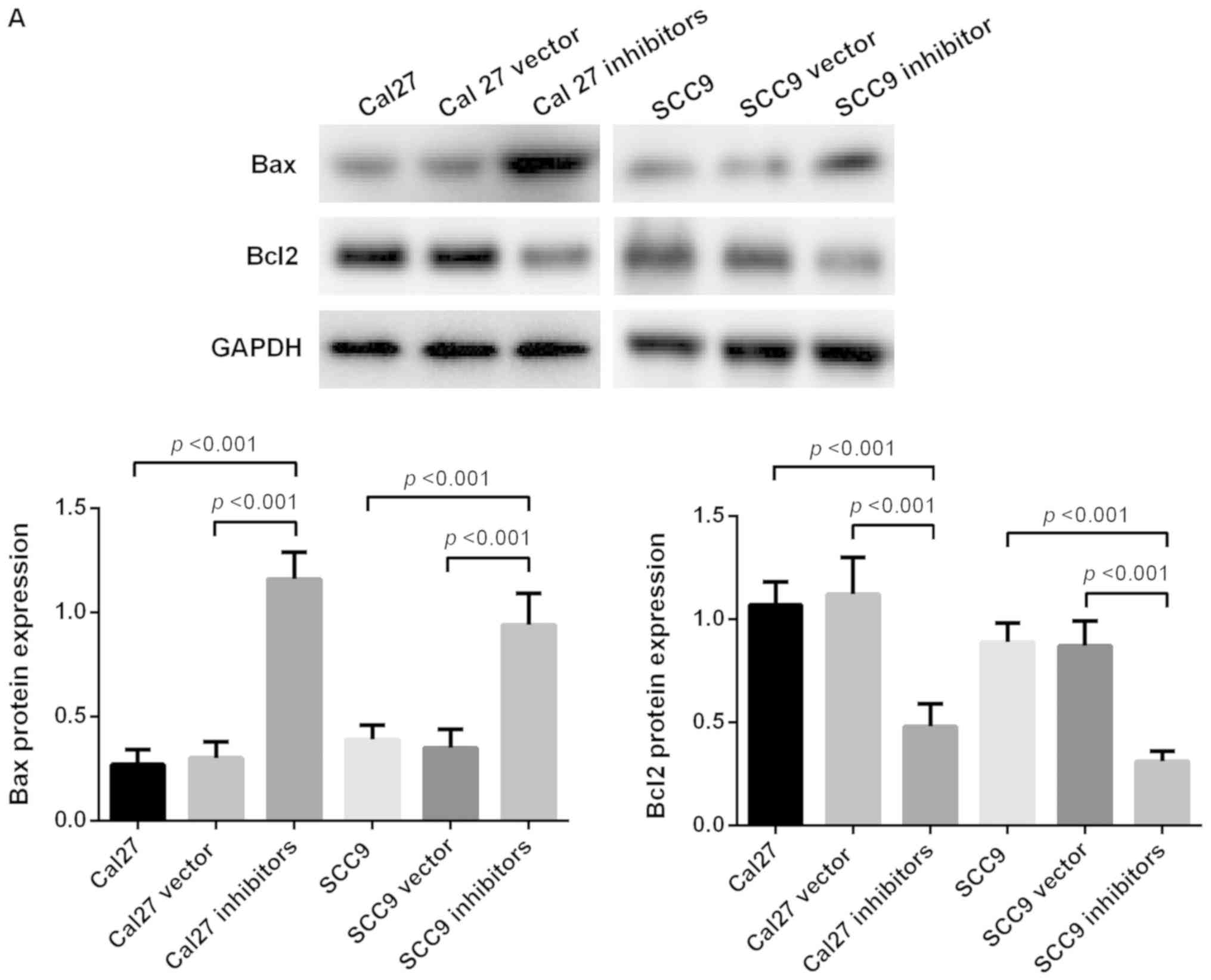
Inhibition of miR-21-5p promotes
apoptosis of Cal 27 and SCC9 cells
To investigate the effect of miR-21-5p on Cal 27 and
SCC9 cells, western blot analysis was used to detect apoptosis. The
results revealed that the expression levels of the pro-apoptotic
protein Bax increased significantly following miR-21-5p knockdown,
while the expression level of anti-apoptosis protein Bcl2 was
significantly decreased following knockdown of miR-21-5p. PI and
Annexin V staining revealed that the early apoptosis of SCC9 and
Cal 27 cells significantly increased following miR-21-5p knockdown
(P<0.05). These results demonstrate that the downregulation of
miR-21-5p expression promotes the apoptosis of Cal 27 and SCC9
cells (Fig. 4).
PDCD4 is the target of miR-21-5p
miRNAs affect the expression levels of mRNA by
binding to their 3′-UTR, and the role of miR-21-5p is closely
associated with the function of the target gene [for example
phosphatase and tensin homolog (PTEN) and Smad7]. Target gene
prediction indicated that PDCD4 may be a potential target of
miR-21-5p (Fig. 5A). RT-qPCR was
used to detect the expression levels of PDCD4 in TSCC tissues. The
results revealed that the expression levels of PDCD4 decreased
significantly in TSCC tissues (Fig.
5B), and the expression levels of miR-21-5p were negatively
correlated with PDCD4 (Fig. 5C). To
determine whether miRNA-21-5p directly targeted PDCD4, the present
study cloned the 3′-UTR of PDCD4 downstream from the pMIR-CMV
luciferase reporter. The results revealed that miRNA-21 decreased
luciferase activity in WT-PDCD4-transfected cells (Fig. 5D). The luciferase assay demonstrated
that PDCD4 was the target of miR-21-5p.
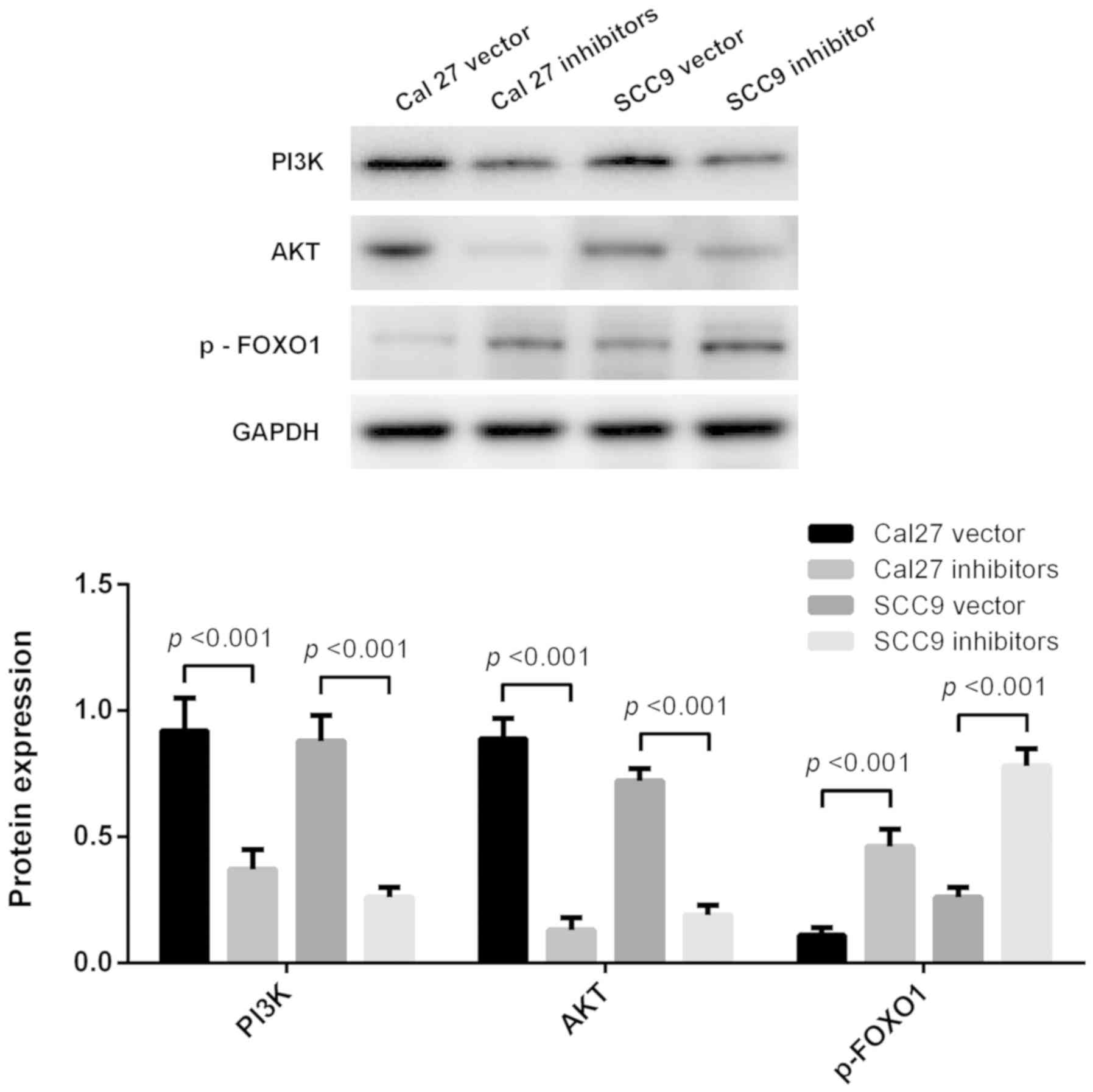
Apoptosis effect of miRNA-21-5p is
determined via regulating the PI3K/Akt/FOXO1 signaling pathway
PI3K, AKT and p-FOXO1 protein expression levels were
measured using western blot analysis (Fig. 6). The results revealed that the PI3K,
AKT and p-FOXO1 protein expression levels were significantly
decreased following miR-21-5p knockdown both in Cal27 and SCC9
cells (Fig. 6). Inhibition of
miR-21-5p promoted apoptosis of Cal27 and SCC9 cells via regulating
the PI3K/AKT/FOXO1 signaling pathway.
Discussion
miRNAs are factors that regulates biological
responses including apoptosis, proliferation, differentiation and
invasion. miRNAs account for ~1% of the genome, but they regulates
~60% of gene activity (14). Under
the catalysis of RNA polymerase II, miRNA is transformed into
primitive miRNAs with a cap structure and polyA tail, then the
primitive RNA forms a stem-loop structure, which is transferred
from the nucleus to the cytoplasm with the help of transporter
exportin-5. This stem-loop structure is cut to form mature miRNAs.
Finally, this stem-loop structure is cleaved to form mature miRNAs
(15). The mature miRNAs bind to the
RNA-induced silencing complex (RISC) to form an asymmetric RISC
complex, which binds to the 3′-UTR on the mRNA of the target gene
to regulate the expression of that target gene (15).
miR-21-5p is a member of the miRNA family, which is
highly expressed in the serum of patients with a number of
different types of tumors and serves an anti-apoptotic role
(9). miR-21-5p is closely associated
with the molecular mechanisms underlying drug resistance, cell
invasion and metastasis of tumors, among other processes (16–18). The
results of the present study revealed that the expression levels of
miRNA21-5p were upregulated in human TSCC, which was involved in
the occurrence and development of tumors. CCK-8 and Transwell assay
results demonstrated that the proliferative and invasive
capabilities of the Cal 27 and SCC9 cell lines decreased following
downregulation of miR-21-5p expression. Western blot analysis was
used to detect the apoptotic and anti-apoptotic factors in the
present study. The results revealed that downregulation of
miR-21-5p promoted the apoptosis of Cal 27 and SC9 cells.
miR-21-5p directly or indirectly promotes the
pathophysiological process of cells by regulating the expression of
PDCD4, PTEN and tissue inhibitor of metalloproteinase-3 (19,20).
Usually one miRNA targets multiple genes. As PDCD4 has been
frequently reported as a target of miR-21-5p, the role of miR-21-5p
in PDCD4 expression in TSCC requires additional study (21). In the present study, the luciferase
reporter assay revealed that PDCD4-UTR-pMIR (WT) was significantly
increased by miR-21-5p inhibitors. This indicated that PDCD4 was
the target of microRNA-21-5p, but its mechanism requires additional
experimental confirmation.
PI3K/AKT/FOXO1 is an important signaling pathway,
which regulates progression in numerous different types of tumors.
The PI3K/AKT pathway is closely associated with apoptosis. This
pathway controls cell proliferation, growth, translation, migration
and survival, and overactivation of this signaling pathway is
associated with poor prognosis (21,22).
Activated PI3K may activate downstream protein kinase AKT;
Activated AKT is able to phosphorylate Bax, inactivate Bax and
inhibit Bax and Bcl-2 to form dimers, thereby resulting in Bcl-2
dissociation and an anti-apoptotic effect (23,24). In
the present study, the expression levels of PI3K/AKT signaling
pathway-associated proteins were detected. It was demonstrated that
inhibition of miR21-5p suppressed the PI3K/AKT/FOXO1 signaling
pathway, suggesting that inhibiting the activation of the
PI3K/AKT/FOXO1 signaling pathway may be a potential strategy for
the treatment of TSCC.
In conclusion, downregulation of miR21-5p may
effectively inhibit the proliferation and invasion of cancer cells
and also promote the apoptosis of cancer cells. These data provide
the basis for novel strategies for gene therapy of TSCC.
Acknowledgements
Not applicable.
Funding
The present study was funded by the Nature Science
Foundation of Liaoning Province (grant no. 2015020326).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author upon reasonable
request.
Authors' contributions
ZW and LT conceived and designed the study. CL and
ZT drafted the manuscript. Cell cultures were completed by ZT. The
RT-qPCR protocol was performed by JT. Data analysis was performed
by CL and ZX. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
The present study was approved by the Ethical
Committees of Jinzhou Medical University on October 26, 2016
(approval no. JZH2016052). Informed consent was obtained from all
patients included in the present study.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
TSCC
|
tongue squamous cell carcinoma
|
|
miRNA
|
microRNA
|
|
PDCD4
|
programmed cell death 4
|
|
WT
|
wild-type
|
References
|
1
|
Weatherspoon DJ, Chattopadhyay A,
Boroumand S and Garcia I: Oral cavity and oropharyngeal cancer
incidence trends and disparities in the United States: 2000–2010.
Cancer Epidemiol. 39:497–504. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ramqvist T, Grün N and Dalianis T: Human
papillomavirus and tonsillar and base of tongue cancer. Viruses.
7:1332–1343. 2015. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Karatas OF, Oner M, Abay A and Diyapoglu
A: MicroRNAs in human tongue squamous cell carcinoma: From
pathogenesis to therapeutic implications. Oral Oncol. 67:124–130.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Sun L, Liang J, Wang Q, Li Z, Du Y and Xu
X: MicroRNA-137 suppresses tongue squamous carcinoma cell
proliferation, migration and invasion. Cell Prolif. 49:628–635.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Yuen PW, Lam KY, Chan AC, Wei WI and Lam
LK: Clinicopathological analysis of local spread of carcinoma of
the tongue. Am J Surg. 175:242–244. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Rusthoven K, Ballonoff A, Raben D and Chen
C: Poor prognosis in patients with stage I and II oral tongue
squamous cell carcinoma. Cancer. 112:345–351. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Chu H, Jia B, Qiu X, Pan J, Sun X, Wang Z
and Zhao J: Investigation of proliferation and migration of tongue
squamous cell carcinoma promoted by three chemokines, MIP-3α,
MIP-1β, and IP-10. Onco Targets Ther. 10:4193–4203. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Yan D, Cai X and Feng Y: miR-183 modulates
cell apoptosis and proliferation in tongue squamous cell carcinoma
SCC25 cell line. Oncol Res. 24:399–404. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Jiang Y, Zhang M, Guo T, Yang C, Zhang C
and Hao J: MicroRNA-21-5p promotes proliferation of gastric cancer
cells through targeting SMAD7. Onco Targets Ther. 11:4901–4911.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Agarwal V, Bell GW, Nam JW and Bartel DP:
Predicting effective microRNA target sites in mammalian mRNAs.
eLife. 4:e050052015. View Article : Google Scholar
|
|
12
|
Betel D, Wilson M, Gabow A, Marks DS and
Sander C: microRNA target predictions: The microRNA.org resource:
Targets and expression. Nucleic Acids Res. 36:D149–D153. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Sobin L, Gospodarowicz M and Wittekind C:
UICC International Union Against Cancer TNM Classification of
Malignant Tumors7th. West Sussex, Wiley-Blackwell; United Kingdom:
2009
|
|
14
|
Fang H, Xie J, Zhang M, Zhao Z, Wan Y and
Yao Y: miRNA-21 promotes proliferation and invasion of
triple-negative breast cancer cells through targeting PTEN. Am J
Transl Res. 9:953–961. 2017.PubMed/NCBI
|
|
15
|
Zhang Z, Li Z, Gao C, Chen P, Chen J, Liu
W, Xiao S and Lu H: miR-21 plays a pivotal role in gastric cancer
pathogenesis and progression. Lab Invest. 88:1358–1366. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Mohr AM and Mott JL: Overview of microRNA
biology. Semin Liver Dis. 35:3–11. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ricci C, Marzocchi C and Battistini S:
MicroRNAs as biomarkers in amyotrophic lateral sclerosis. Cells.
7(pii): E2192018. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Bourguignon LY: Matrix hyaluronan promotes
specific MicroRNA upregulation leading to drug resistance and tumor
progression. Int J Mol Sci. 17:5172016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhao Y, Zhao L, Ischenko I, Bao Q, Schwarz
B, Nieß H, Wang Y, Renner A, Mysliwietz J, Jauch KW, et al:
Antisense inhibition of microRNA-21 and microRNA-221 in
tumor-initiating stem-like cells modulates tumorigenesis,
metastasis, and chemotherapy resistance in pancreatic cancer.
Target Oncol. 10:535–548. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Liu ZL, Wang H, Liu J and Wang ZX:
MicroRNA-21 (miR-21) expression promotes growth, metastasis, and
chemo- or radioresistance in non-small cell lung cancer cells by
targeting PTEN. Mol Cell Biochem. 372:35–45. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wang Z, Yao W, Li K, Zheng N, Zheng C,
Zhao X and Zheng S: Reduction of miR-21 induces SK-N-SH cell
apoptosis and inhibits proliferation via PTEN/PDCD4. Oncol Lett.
13:4727–4733. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Pardini B, De Maria D, Francavilla A, Di
Gaetano C, Ronco G and Naccarati A: MicroRNAs as markers of
progression in cervical cancer: A systematic review. BMC Cancer.
18:6962018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Syed DN, Afaq F, Sarfaraz S, Khan N,
Kedlaya R, Setaluri V and Mukhtar H: Delphinidin inhibits cell
proliferation and invasion via modulation of Met receptor
phosphorylation. Toxicol Appl Pharmacol. 231:52–60. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Hao Y, Huang J, Ma Y, Chen W, Fan Q, Sun
X, Shao M and Cai H: Asiatic acid inhibits proliferation, migration
and induces apoptosis by regulating Pdcd4 via the
PI3K/Akt/mTOR/p70S6K signaling pathway in human colon carcinoma
cells. Oncol Lett. 15:8223–8230. 2018.PubMed/NCBI
|