Introduction
In less developed countries, cervical cancer is the
second most commonly diagnosed cancer, with a high mortality rate
(1). Early stage patients
[International Federation of Gynaecology and Obstetrics (FIGO)
stages I–IIA] may experience favourable outcomes by undergoing
radical surgery or radiotherapy, as indicated by an overall 5-year
survival rate of >65% (2,3). However, patients with later-stage
disease, including stage IIB-IV, require more severe therapeutic
strategies, including radiotherapy in addition to chemotherapy. The
5-year survival rate for patients with stage IIB-III cancer is
25–30% (2,3). For stage IV cancer, the survival rate
is <15% (2,3) due to chemoresistance resulting in local
recurrence or distant metastasis. Presently, chemotherapy is
clinically employed as one of the most efficient strategies in the
systematic treatment of cervical cancer. A combination of cisplatin
with other chemotherapeutic drugs has remained the dominant
systemic therapeutic modality for locally advanced and metastatic
cervical cancer for several decades (4). However, chemoresistance limits the
therapeutic effect of these chemoagent and frequently results in
poor prognosis. Therefore, there is an urgent need for novel
chemotherapeutic agents for use alone or in combination with a
primary chemotherapeutic agent.
The natural molecule angelicin,
2-oxo-(2H)-furo(2,3-h)-1-benzopyran, is one of the major active
compounds isolated from the traditional Chinese herb Angelica
archangelica. For decades, angelicin has been clinically used
to exert therapeutic effects on various skin diseases, such as
lichen planus, acting as a photosensitizer (5,6).
Angelicin exertsgenotoxic effects and thus induces cytotoxicity in
several types of tumour and non-tumour cells (7,8). Mira
and Shimizu (9) identified that
angelicin causes cytotoxicity by inhibiting tubulin polymerization
and histone deacetylase 8 activity in several types of tumour
cells, including human hepatocellular carcinoma, rhabdomyosarcoma
and colorectal carcinoma (9). To
investigate the potential mechanism for proliferation inhibition,
Wang et al (10) used liver
cancer for therapeutic research both in vitro and in
vivo. The study determined that the dose- and time-dependent
apoptotic effect of angelicin is caused by the regulation of
mitochondria, involving the P13K/AKT1 signalling pathway.
Accordingly, angelicin affects physiological processes in both
tumour and non-tumour cells.
Autophagy is a highly conserved multi-step lysosomal
degradation process. Cellular components are sequestered in
autophagosomes that subsequently fuse with lysosomes to degrade the
contents (11). Accumulating
evidence has established a close association between autophagy and
tumour progression, with autophagy having different functions
during tumour progression, including tumour suppression and
enhancement (5,6,12). Tsai
et al (5) reported that the
natural agent
1-(2-hydroxy-5-methylphenyl)-3-phenyl-1,3-propanedione, exerts
growth-inhibiting effects by promoting the autophagy of HeLa
cervical cancer cells. Li et al (6) determined that protein kinase C-β
inhibited autophagy and consequently sensitized HeLa cells to
chemotherapy. The study also reported that an increase in autophagy
inhibited cell growth and induced apoptotic cell death (12). The dynamic role of autophagy in
tumour progression has been the focus of research for potential
therapeutics. However, further studies into strategies for
controlling autophagy are required to increase understanding into
the association between autophagy and tumour progression.
The growth-inhibiting and apoptosis-promoting
effects of angelicin in several types of cancers have been
previously reported (13). However,
whether cervical cancer is chemosensitive to angelicin has not been
demonstrated. Therefore, the present study used the human cervical
carcinoma cell line, HeLa and the human cervical squamous cell
carcinoma cell line, SiHa as in vitro models to determine
the anticancer effects of angelicin. To evaluate its specific
activity on cervical cancer cells, the non-tumour cervical
epithelial cell line ECT1/E6E7 was also employed. The investigation
primarily focused on the regulation of malignant behaviours by
inducing or inhibiting autophagy in HeLa and SiHa. In addition, the
effects of angelicin on autophagy and the potentially relevant mTOR
signalling pathway were explored. The results of the present study
may reveal the novel effects of angelicin as a chemotherapeutic
strategy in certain types of cervical carcinomas.
Materials and methods
Cell culture and treatment
The human cervical carcinoma cell line, HeLa and the
cervical squamous cell carcinoma cell line, SiHa was obtained from
the American Type Culture Collection (accession no. HTB-35). The
cervical epithelial cell line, ECT1/E6E7 was purchased from Jennio
Biotech Co., Ltd. and used for identifying the difference of
chemosensitivity between cancer cell lines and a non-tumor cell
line. All cells were cultured at 37°C in a 5% CO2
incubator in DMEM (Gibco; Thermo Fisher Scientific, Inc.)
supplemented with 100 µg/ml streptomycin, 100 U/ml penicillin and
10% FBS (Gibco; Thermo Fisher Scientific, Inc.). Cells were
passaged every 3 days.
For identifying chemosensitivity, 0, 20, 40, 60, 80,
100, 120, 140, 160, 180 or 200 µM angelicin (cat. no. A0956-10MG;
Sigma-Aldrich; Merck KGaA) was added to the medium of HeLa or SiHa
for 24 h. For 5-ethynyl-2′-deoxyuridine (Edu) staining, cell cycle
distribution, colony formation, tumor formation in soft agar,
migration and invasion assays, the IC30 of angelicin (27.8 µM) was
employed to evaluate the effects of angelicin on malignant
behaviors. For carboxyfluorescein succinimidyl ester
(CFSE)/propidium iodide (PI) or Annexin V FITC/PI double staining,
the IC50 of angelicin was employed. For inhibiting the
degradation of microtubule associated protein 1 light chain 3-β
(LC3B)-II, cells were pretreated with 10 µM of chloroquine (cat.
no. C6628; Sigma-Aldrich; Merck KGaA) for 6 h. For rapamycin (cat.
no. V900930; Sigma-Aldrich; Merck KGaA) pretreatment, cells were
pretreated with 1 µM of rapamycin for 6 h. Mock group containing
vehicle only was considered as negative control in all the
experiments.
Cell counting kit-8 (CCK-8) assay
To determine HeLa or SiHa cell viability,
5×103 cells were plated in 96-well plates. The
aforementioned treatment was administered and 10 µl tetrazolium
salt WST-8 (KeyGen Biotech. Co. Ltd.) was added to each well for a
4 h incubation at 37°C. Optical density (OD) was measured at a
wavelength of 450 nm using a microplate reader (Synergy 2
Multi-Mode Microplate Reader; BioTek Instruments, Inc.).
EdU staining
HeLa or SiHa cells were seeded at a density of
2×105 cells per well in 6-well plates supplemented with
DMEM containing 50 µM EdU (RiboBio Co. Ltd.). Following 2 h
incubation at room temperature, cells were washed with ice-cold PBS
and fixed with 4% paraformaldehyde for 10 min at room temperature.
EdU immunostaining was performed with Apollo staining reaction
buffer followed by nuclei staining with Hoechst 33342 (cat. no.
B2261; Sigma-Aldrich; Merck KGaA) at final concentration of 10
µg/ml at room temperature for 10 min. Stained cells were imaged
under a X71 (U-RFL-T) fluorescence microscope (Olympus Corporation;
magnification, ×40).
PI staining
HeLa or SiHa cells were dissociated using 0.25%
trypsin (Thermo Fisher Scientific, Inc.) and three time washed with
PBS. Following the last wash, the cell pellet, which was
centrifuged at 400 × g for 10 min at room temperature, was
suspended and fixed in 70% ice-cold alcohol overnight at 4°C. Cells
were then washed in triplicate with ice-cold PBS and suspended in
400 µl PI solution (5 µg/ml) for 30 min in the dark. Apoptotic
cells were analyzed via flow cytometry using a 3 laser Navios flow
cytometer (Beckman Coulter, Inc.) and analyzed using FlowJo
software (FlowJo LLC; version 9).
Colony formation
HeLa or SiHa cells were seeded in 6-well plates at a
density of 1,000 cells/well. Cells were then cultured at 37°C for
10 days until visible colonies appeared. Colonies were stained with
500 µl Giemsa solution (Nanjing KeyGen Biotech Co., Ltd.) and
incubated for 30 min at 37°C. Colonies were then imaged using a X71
(U-RFL-T) fluorescence microscope (Olympus Corporation;
magnification, ×40).
Tumor formation in soft agar
To assess tumor formation in vitro, soft agar
clonogenic assays were performed. Each well of a 6-well plate was
coated with 2 ml of 0.5% (w/v) low-melting agar (Sigma-Aldrich;
Merck KGaA) in DMEM with 10% FBS. Cells were mixed and
5×103 cells in 2 ml 0.3% low-melting agar with 10% FBS
were added above the polymerized base solution. Plates were
incubated (37°C; 5% CO2) for 14 days before colony
number and diameter were quantified microscopically using a X71
(U-RFL-T) fluorescence microscope (Olympus Corporation;
magnification, ×40).
Scratch wound healing assay
HeLa or SiHa cells were seeded at a density of
1×106 in 6-well plates and allowed to attach for 24 h.
When cell confluence reached ~100%, a scratch wound was
subsequently introduced by scraping the cell monolayer with a 10 µl
sterile micropipette tip. Cells were then washed with PBS to remove
unattached cells and incubated at 37°C for 24 h in medium
containing 1% FBS. Cells were then imaged at the same site at 0 and
24 h following induction of the scratch using a X71 (U-RFL-T)
fluorescence microscope (Olympus Corporation; magnification,
×40).
Transwell invasion assay
HeLa or SiHa cells were dissociated with 0.25%
trypsin and washed three times with ice-cold PBS. In the lower
chamber, 500 µl of DMEM supplemented with 10% FBS was added. A
total of 200 µl of cells at the concentration of
2×105/ml were seeded into the top chamber of transwell
inserts containing 8 µM pore polycarbonate filters (Corning Inc.)
that had been precoated with Matrigel for 2 h at room temperature
(BD Biosciences). The plate was incubated at 37°C for 24 h.
Experiments were performed in triplicate. Following 24 h of
incubation, the cells on the upper membrane were removed and the
invaded cells were stained with 0.25% crystal violet (Beyotime
Institute of Biotechnology) at room temperature for 10 min then
counted using a X71 (U-RFL-T) fluorescence microscope (Olympus
Corporation; magnification, ×40).
CFSE/PI double staining
A total of 1×106 HeLa or SiHa cells were
seeded in 6-well plates and allowed to attach overnight. Then 100
µl of CFSE fluorescent dye (Sigma-Aldrich; Merck KGaA) was added
and incubated at 37°C for 15 min. Supernatant was removed and cells
were washed with DMEM without FBS. Following the aforementioned
treatments, cells were incubated with PI to a final concentration
of 5 µg/ml at room temperature for 10 min. Cells were imaged using
a X71 (U-RFL-T) fluorescence microscope (Olympus Corporation;
magnification, ×40).
Annexin V/PI double staining
HeLa or SiHa cells were dissociated using 0.25%
Trypsin and washed three times with ice-cold PBS. Following the
last wash, cells were suspended in PBS and the cell concentration
was adjusted to 1×106 cells/ml. Cells were
simultaneously stained with Annexin V-FITC (green fluorescence) and
PI (red fluorescence), which allowed for the identification of
intact cells (FITC−/PI−), early apoptotic
cells (FITC+/PI−) and late apoptotic cells
(FITC+/PI+). Samples were analyzed using the
FACS LSRII flow cytometer (BD Biosciences) with FlowJo software
(FlowJo LLC; version 9).
Immunofluorescence microscopy
HeLa or SiHa cells were plated in 6-well plates on
coverslips and allowed to attach for 24 h. Cells were fixed with 4%
paraformaldehyde for 20 min and permeabilized with 0.2% Triton
X-100 (Sigma-Aldrich; Merck KGaA) for 10 min at room temperature.
Normal goat serum (5%; Sigma-Aldrich; Merck KGaA) in PBS was used
for unspecific blocking at room temperature for 30 min. Cells were
incubated with primary antibodies against LC3B (1:2,000; cat. no.
ab48394; Abcam) at room temperature for 2 h. Cells were then rinsed
four times with PBS-Tween 20 and incubated with secondary
antibodies produced in rabbit (1:500 in 0.5% normal goat serum)
conjugated with Alexa Fluor 488 for 1 h at room temperature. Cell
nuclei were stained with DAPI (Sigma-Aldrich; Merck KGaA) at room
temperature for 10 min. Images were captured with a X71 (U-RFL-T)
fluorescence microscope (Olympus Corporation) at a magnification of
×200.
Western blot analysis
Following the aforementioned treatments, HeLa or
SiHa cells were lysed in chilled lysis buffer containing 50 mM
Tris-HCl (pH 7.4), 150 mM NaCl, 1% Triton X-100, 5 mM EDTA, 1 mM
Na3VO4, 1 mM NaF and 10 µM PMSF on ice for 10
min. Supernatants were collected via centrifugation at 12,000 × g
for 10 min at 4°C. The extracted protein concentration was measured
using Bicinochoninic Acid kit (Sigma-Aldrich; Merck KGaA) for
Protein Determination according to the manufacturer's protocol. For
each sample, 20 µg of total protein was loaded per lane and
separated via SDS-PAGE on a 12.5% gel, then transferred onto a
nitrocellulose membrane (EMD Millipore) followed by blocking using
5% BSA (Sigma–Aldrich; Merck KGaA). The membrane was subsequently
incubated with the following primary antibodies at a dilution of
1:1,000 overnight at 4°C: Rabbit anti-LC3B (cat. no. ab48394;
Abcam), rabbit anti-β-actin (cat. no. ab8227; Abcam), rabbit
anti-autophagy related protein (Atg)-3 (cat. no. ab108251; Abcam),
rabbit anti-Atg7 (cat. no. ab133528; Abcam), rabbit anti-β-actin
(cat. no. ab8227; Abcam), rabbit anti-Atg12-Atg5 (cat. no.
orb375397; Biorbyt Ltd.), rabbit anti-mTOR (cat. no. ab2732; Abcam)
and rabbit anti-mTOR (phospho S2448, cat. no. ab109268, Abcam).
Membranes were subsequently incubated with horseradish
peroxidase-conjugated secondary antibodies (goat anti-rabbit IgG
H&L antibody; cat. no. ab7090; 1:5,000; Abcam) for 2 h at room
temperature. Enhanced chemiluminescence reagents (cat. no. PRN2232;
GE Healthcare Bio-Sciences) were used to visualize protein
bands.
Statistical analysis
Statistical analysis was performed with SPSS version
13.0 (SPSS, Inc.). Data were expressed as the mean ± standard
deviation. All experiments were repeated three times,
independently. Comparisons between groups were assessed using a
Student's t-test for two groups and one-way analysis of variance
followed by Bonferroni post hoc analysis for multiple groups.
P<0.05 was considered to indicate statistical significance.
Results
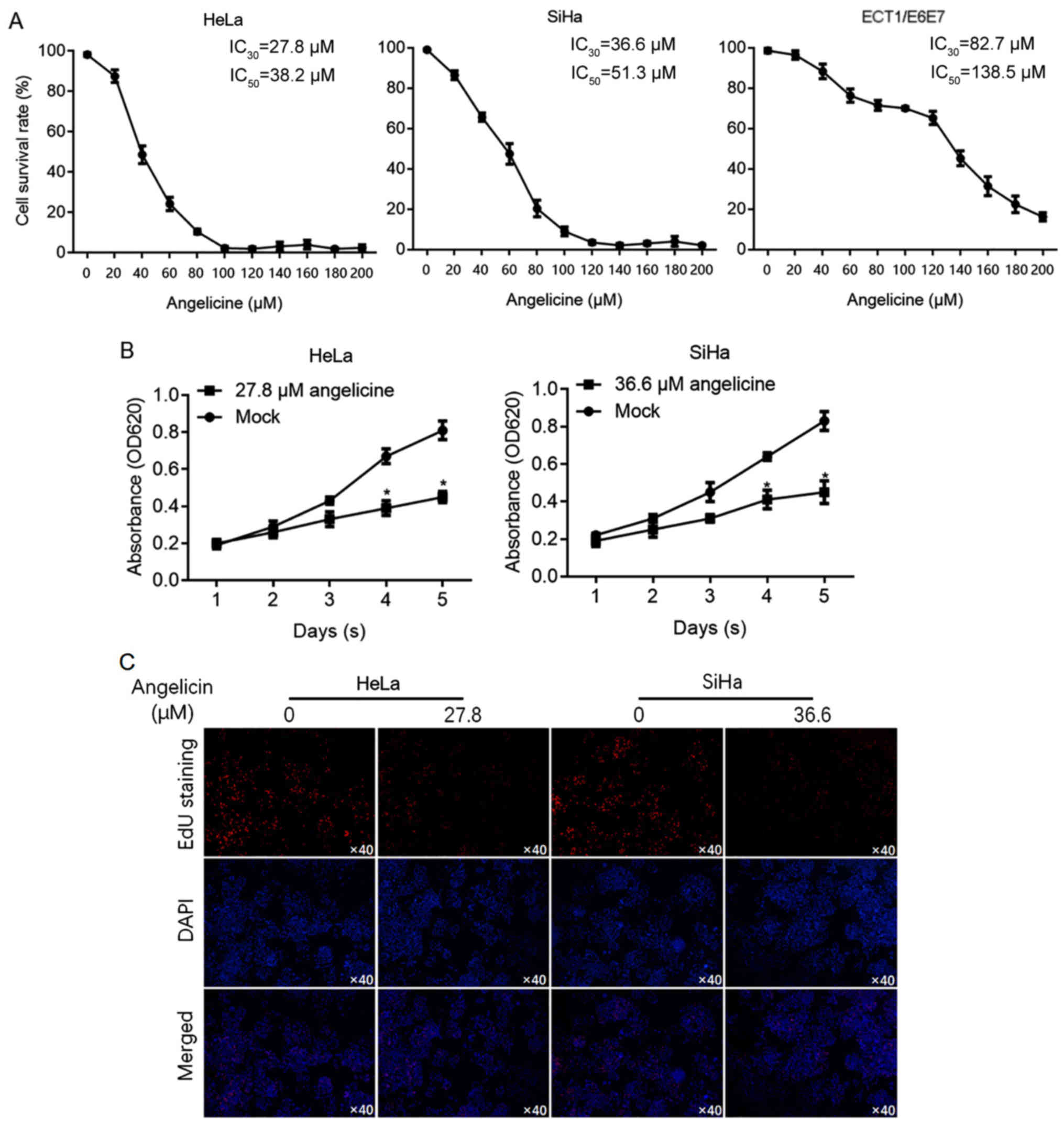
Cervical cancer cells are more
sensitive to angelicin than cervical epithelial cells
It has been reported that angelicin is cytotoxic to
hepatic cancer cells (10). To
investigate the effects of angelicin in cervical cancer cells (HeLa
and SiHa), cell viability was determined using CCK-8 assay
following exposure to a range of angelicin concentrations for 24 h.
For comparison, the sensitivity of cervical epithelial cells
(ECT1/E6E7) to angelicin was also measured to evaluate the 30%
inhibitory concentration (IC30) and 50% inhibitory
concentration (IC50). The results revealed that HeLa
(IC30, 27.8 µM; IC50, 38.2 µM) and SiHa
(IC30, 36.6 µM; IC50, 51.3 µM) cells were
more sensitive to angelicin than ECT1/E6E7 cells (IC30,
82.7 µM; IC50, 138.5 µM; Fig.
1A). Cell viability was assessed on days 1–5 following
treatment with angelicin at the IC30. The results
revealed that Angelicin treatment significantly inhibited HeLa and
SiHa cell proliferation (P<0.05 vs. mock group containing
vehicle only; Fig. 1B). To confirm
that the decrease in cell viability was due to a change in cell
proliferation, EdU labelling of proliferating cells was performed.
The results revealed that HeLa and SiHa cell treatment with
angelicin at the IC30 substantially decreased the number
of proliferating cells compared with the control (Fig. 1C).
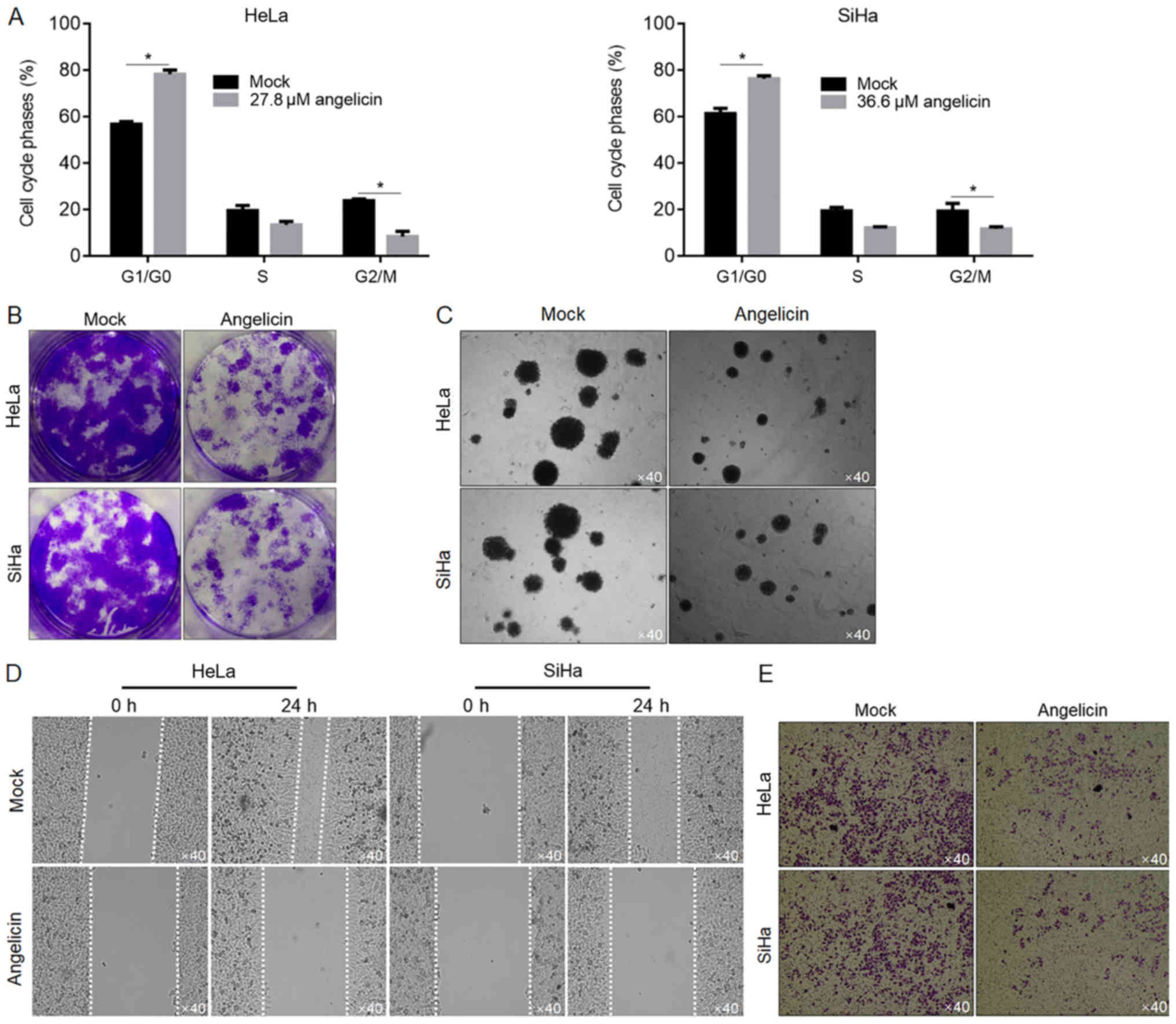
Angelicin treatment inhibits the
migration and invasion of cervical cancer cells
To determine whether angelicin inhibited cell
proliferation by regulating cell cycle phase distribution, flow
cytometry of PI-stained cells was performed to detect cell cycle
entry following angelicin treatment. The ratio of cells in the G1
and G0 phases following angelicin treatment increased significantly
compared with the mock group (P<0.05; Fig. 2A), whilst the ratio of cells in the
G2/M phases decreased substantially (P<0.05; Fig. 2A). To investigate the effects of
angelicin IC30 treatment on HeLa or SiHa cells, colony
formation was assessed by seeding cells at low density to obtain
single-cell-derived colonies as previously described (14). Mock-treated HeLa and SiHa cells
formed single-cell-derived colonies (Fig. 2B). By contrast, angelicin treatment
substantially decreased the colony formation ability of both HeLa
and SiHa cells (Fig. 2B). The effect
of angelicin on tumour formation in soft agar was investigated.
Consistent with the effects exerted on colony formation (>50 µM
in diameter), angelicin treatment markedly decreased the tumour
formation ability of the cells in soft agar compared with the
control (Fig. 2C). Angelicin
treatment substantially inhibited the migration and invasion of
cells compared with mock treated cells (Fig. 2D and E).
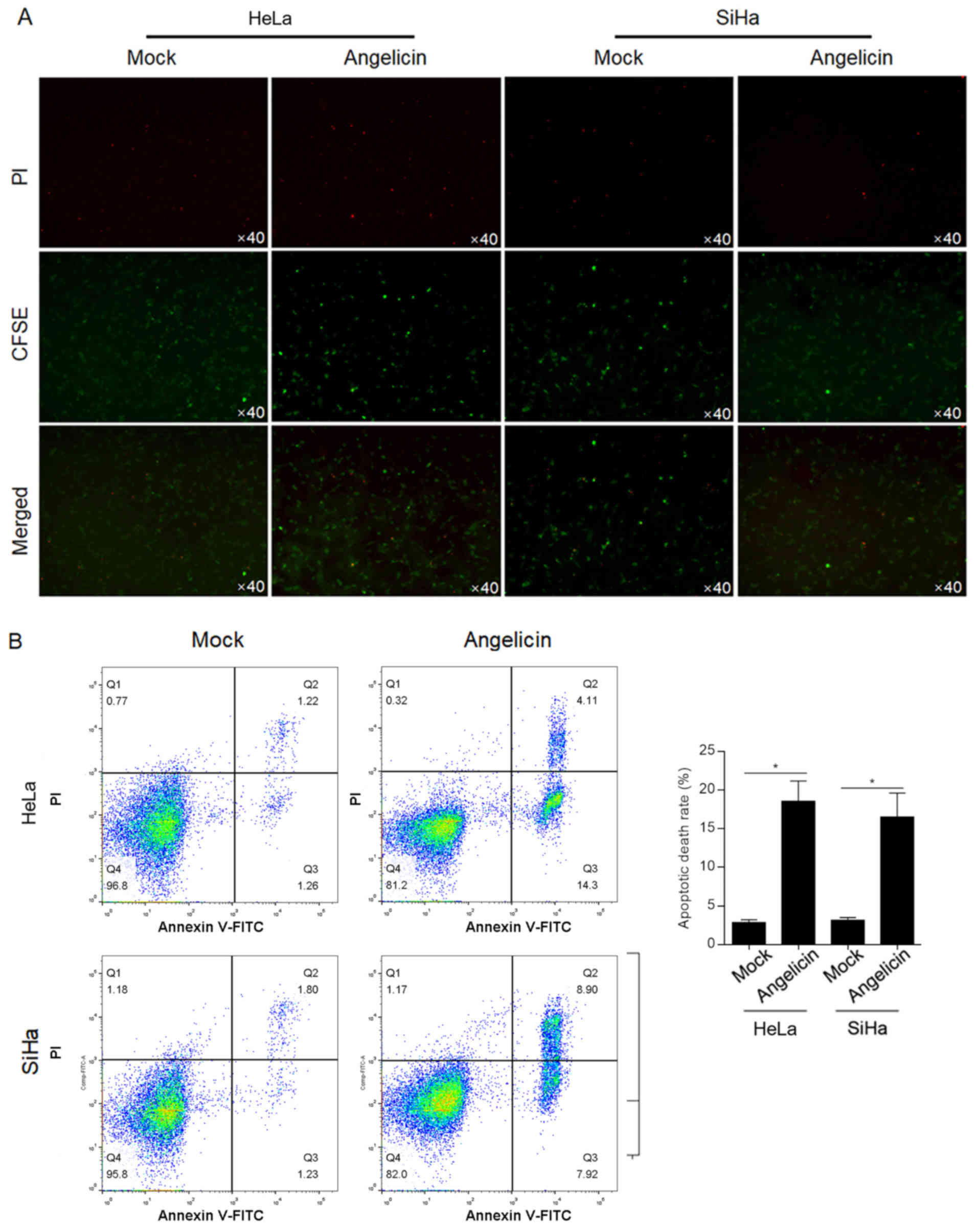
Angelicin induces apoptotic cell death
in HeLa and SiHa cells
The cytotoxic effect of angelicin was verified by a
cytotoxicity assay with CFSE-labelled HeLa or SiHa cells.
CFSE-positive cells demonstrated no significant change following
angelicin treatment compared with mock treated cells (Fig. 3A). Following PI staining, it was
observed that angelicin treatment markedly increased the number of
PI-positive cells compared with mock treated cells, indicating that
angelicin treatment increased the cell death rate (Fig. 3A). To further confirm that the
increased cell death by angelicin treatment was due to induction of
apoptosis, Annexin V/PI double-staining was performed. Angelicin
treatment increased the apoptotic cell death rate (Annexin
V+/PI− and Annexin
V+/PI+) in both HeLa (18.7±2.4%) and SiHa
(16.9±3.1%) cells (P<0.05; Fig.
3B). Taken together, the results indicated that angelicin
treatment inhibited the malignant behaviours of HeLa and SiHa
cervical cancer cells and induced apoptotic cell death.
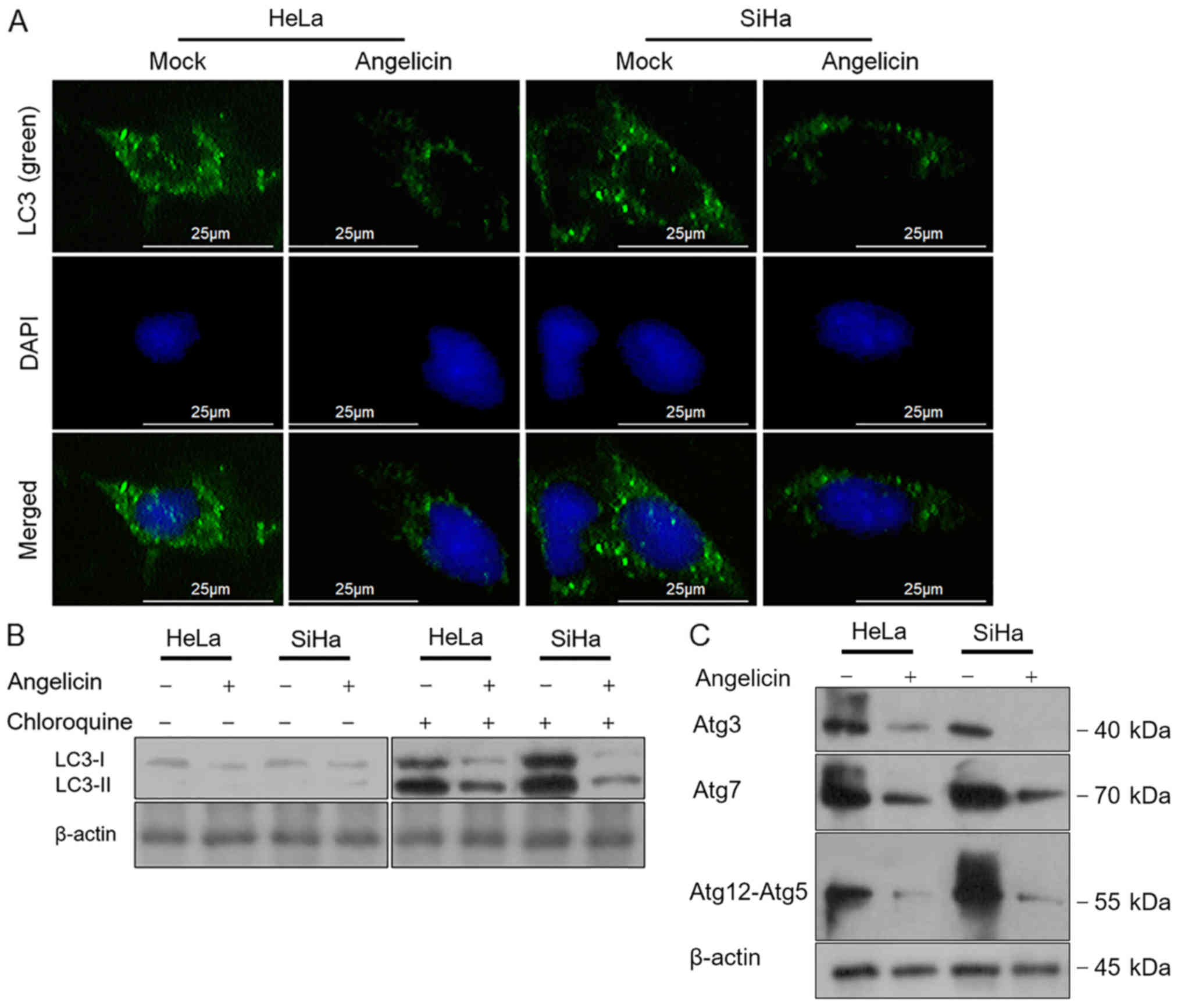
Angelicin inhibits autophagy in
cervical cancer cells
The chemotherapy-induced inhibition of cell
viability, proliferation or cell death leading to the demise of
cancer cells is mediated by autophagic pathways (15,16).
Therefore, the effects of angelicin treatment on autophagy were
investigated in the present study. The regulatory effects of
angelicin on autophagy were assessed by performing LC3B
immunostaining. The results revealed that the LC3B-stained signal
was greatly decreased following angelicin treatment for 24 h
compared with the mock group in HeLa and SiHa cells (Fig. 4A). Chloroquine, a lysosome inhibitor
inhibiting the fusion of autophagosomes and lysosomes and/or the
activity of autolysosomes (17) was
employed to accumulate LC3B-I and -II. Following the inhibition of
LC3B degradation, the results revealed that angelicin treatment
decreased the quantity of LC3B and cleaved LC3B-II compared with
mock cells (Fig. 4B). The formation
of an autophagosome involves the coordinated action of several Atg
protein complexes (18–20). Thus, the expression of certain Atg
proteins, including Atg3, Atg7 and Atg12-5, was determined via
western blot analysis. Consistent with the change in LC3B, Atg3,
Atg7 and Atg12-5 protein levels decreased following angelicin
treatment compared with mock treatment (Fig. 4C).
Angelicin activates mTOR
phosphorylation and potentially regulates malignant behaviours by
modulating autophagy in cervical cancer cells
mTOR is a central regulator of several physiological
processes, including autophagy. Therefore, the current study
assessed whether angelicin treatment regulated mTOR and thus
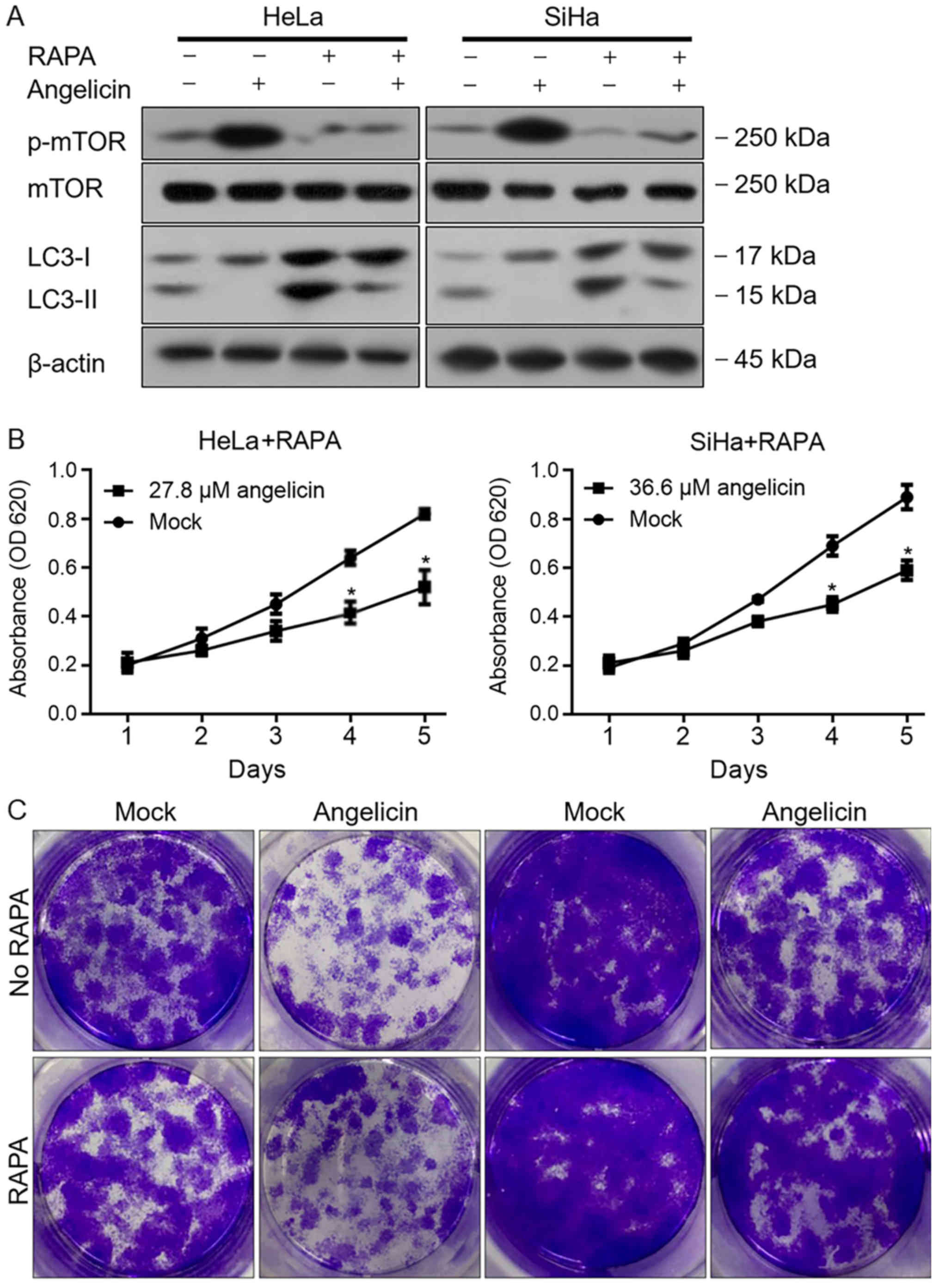
affected autophagy. The results revealed that Angelicin treatment
markedly increased the phosphorylation of mTOR in HeLa and SiHa
cells, and decreased LC3B-II when compared with mock treated cells
(Fig. 5A). Following the addition of
rapamycin, angelicin-induced mTOR phosphorylation was decreased,
but the inhibitory effects of angelicin on autophagy were not fully
reversed, indicating that mTOR might not be a direct target of
angelicin, at least in part. To further confirm these results, cell
viability and colony formation were measured following
co-incubation with rapamycin and angelicin or rapamycin alone. As
presented in Fig. 5B and C,
angelicin treatment decreased the malignant behaviors of HeLa and
SiHa cells, and indicated that rapamycin may exert its inhibitory
effect in an mTOR-independent manner. Taken together, the results
indicated that angelicin treatment regulated the phosphorylation of
mTOR; however, this was not the main mechanism for affecting
autophagy, which remains unknown.
Discussion
Angelicin has been previously reported to possess
anticancer properties. In liver cancer, by activating the PI3K/AKT1
signalling pathway, angelicin treatment induced
mitochondrial-dependent apoptotic cell death (10). Angelicin transcriptionally regulates
members of the Bcl-2 family of proteins that serve key roles in the
regulation of the mitochondrial apoptotic pathway (21). The Bax/Bcl-2 ratio is therefore
altered to cause mitochondrial destabilization, which leads to the
release of proapoptotic factors (22). In the non-small cell lung cancer cell
line and its sub-line (A549 and A549/D16, respectively) that
exhibits multidrug resistance, angelicin treatment promoted
chemotherapy-induced apoptotic cell death and sensitized A549/D16
cells to chemotherapy (23).
However, the antitumor effects of angelicin in human cervical
carcinoma, as well as the mechanisms underlying its actions, are
largely unknown. D'Anqiolillo et al (24) reported that angelicin exerts
cytotoxic activity on HeLa cells, but did not elucidate the exact
mechanism by which this occurs (24). Therefore, the aim of the present
study was to assess the effects of angelicin on the human cervical
carcinoma cell lines, HeLa and SiHa, and to investigate the
molecular mechanisms underlying its action. To determine whether
cervical carcinomas were sensitive to angelicin, the non-tumour
cervical epithelial cell line ECT1/E6E7 was also analyzed.
The present study determined that angelicin exerted
antitumor effects on HeLa and SiHa cells but demonstrated no
detectable cytotoxity to ECT1/E6E7 cells. Treatment of both HeLa
and SiHa cells with angelicin at the IC30 suppressed
malignant behaviours, including proliferation, colony formation,
tumour formation in soft agar, migration and invasion. When
evaluating migrating ability, medium containing 1% FBS was used
instead of serum-free medium, which may be a limitation to the
present study. Treatment with the IC50 of angelicin
significantly induced cell death via apoptosis. Flow cytometry was
employed to determine angelicin-induced apoptosis, however, a study
limitation was that the detection of apoptosis markers, such as
caspase-3, was not performed. The present study identified that
angelicin treatment greatly inhibited autophagy by measuring
hallmarks of autophagy, including LC3BI, LC3BII, Atg3, Atg7 and
Atg12-5. Emerging evidence has indicated that interactions between
autophagy and apoptosis occur via crucial proteins, including mTOR
and Atgs (25). Through these
regulatory mediators of crosstalk, cooperation between autophagy
and apoptosis has been established. However, the present in
vitro study requires in vivo research to further confirm
the results gained.
mTOR is a critical regulator of autophagy that
integrates nutrient signals and cytokines from different pathways,
inhibiting autophagy and promoting cell growth (26). Signal starvation inhibits the
phosphorylation of mTOR and initiates autophagy by forming
theunc-51-like kinase complex, which comprises Atg13 and a protein
tyrosine kinase 2-family interacting protein of 200 kDa (27,28). The
present study determined that angelicin treatment induced marked
phosphorylation of mTOR without altering the total amount of mTOR.
However, mTOR signalling has also been revealed to have no
significant role in controlling autophagic flux (29), which may explain why rapamycin
treatment failed to inhibit the effects of angelicin on autophagy
regulation.
In conclusion, the present study demonstrated that
angelicin treatment significantly inhibited malignant behaviours,
including proliferation, colony formation, tumour formation,
migration and invasion, in cervical cancer cells, potentially by
inhibiting autophagy. Although angelicin treatment induced the
phosphorylation of mTOR, its regulatory roles on autophagy and
malignant behaviours were identified to be independent of mTOR
signalling. Further studies are required to elucidate the exact
molecular mechanisms underlying the regulatory role of angelicin on
cervical cancer malignant behaviours. The results of the current
study indicated that angelicin may have potential as a
chemotherapeutic agent against cervical cancer.
Acknowledgements
The author would like to thank Mrs. Yun Bai (Third
Military Medical University, Chongqing) for language editing.
Funding
No funding was received.
Availability of data and materials
The datasets generated and/or analyzed during the
current study are available from the corresponding author on
reasonable request.
Authors' contributions
YW, ZL and YC designed the experiments. XC, YL and
DY performed cell culture and data analysis. YW wrote the
manuscript. JD collected data and performed statistical analysis.
NY is responsible for data collection. All authors read and
approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Burger H, Loos WJ, Eechoute K, Verweij J,
Mathijssen RH and Wiemer EA: Drug transporters of platinum-based
anticancer agents and their clinical significance. Drug Resist
Updat. 14:22–34. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Lilic V, Lilic G, Filipovic S, Milosevic
J, Tasic M and Stojiljkovic M: Modern treatment of invasive
carcinoma of the uterine cervix. J BUON. 14:587–592.
2009.PubMed/NCBI
|
|
4
|
Rodríguez Villalba S, Díaz-CanejaPlanell C
and Cervera Grau JM: Current opinion in cervix carcinoma. Clin
Transl Oncol. 13:378–384. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Tsai JH, Hsu LS, Huang HC, Lin CL, Pan MH,
Hong HM and Chen WJ:
1-(2-Hydroxy-5-methylphenyl)-3-phenyl-1,3-propanedione induces g1
cell cycle arrest and autophagy in HeLa cervical cancer cells. Int
J Mol Sci. 17:E12742016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Li N and Zhang W: Protein kinase C β
inhibits autophagy and sensitizes cervical cancer HeLa cells to
cisplatin. Biosci Rep. 37:BSR201604452017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kavli G, Midelfart K, Raa J and Volden G:
Phototoxicity from furocoumarins (psoralens) of Heracleum
laciniatum in a patient with vitiligo. Action spectrum studies on
bergapten, pimpinellin, angelicin and sphondin. Contact Dermatitis.
9:364–336. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lampronti I, Bianchi N, Borgatti M, Fibach
E, Prus E and Gambari R: Accumulation of gamma-globin mRNA in human
erythroid cells treated with angelicin. Eur J Haematol. 71:189–195.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Mira A and Shimizu K: In vitro cytotoxic
activities and molecular mechanisms of angelica shikokiana extract
and its isolated compounds. Pharmacogn Mag. 11 (Suppl 4):S564–S569.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wang F, Li J, Li R, Pan G, Bai M and Huang
Q: Angelicin inhibits liver cancer growth in vitro and in
vivo. Mol Med Rep. 16:5441–5449. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Fang W, Shu S, Yongmei L, Endong Z, Lirong
Y and Bei S: miR-224-3p inhibits autophagy in cervical cancer cells
by targeting FIP200. Sci Rep. 6:332292016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Spanò V, Parrino B, Carbone A, Montalbano
A, Salvador A, Brun P, Vedaldi D, Diana P, Cirrincione G and
Barraja P: Pyrazolo[3,4-h]quinolines promising photosensitizing
agents in the treatment of cancer. Eur J Med Chem. 102:334–351.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Bruni R, Barreca D, Protti M, Brighenti V,
Righetti L, Anceschi L, Mercolini L, Benvenuti S, Gattuso G and
Pellati F: Botanical sources, chemistry, analysis, and biological
activity of furanocoumarins of pharmaceutical interest. Molecules.
24:E21632019. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Colter DC, Class R, DiGirolamo CM and
Prockop DJ: Rapid expansion of recycling stem cells in cultures of
plastic-adherent cells from human bone marrow. Proc Natl Acad Sci
USA. 97:3213–3218. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Hu L, Sun S, Wang T, Li Y, Jiang K, Lin G,
Ma Y, Barr MP, Song F, Zhang G and Meng S: Oncolytic newcastle
disease virus triggers cell death of lung cancer spheroids and is
enhanced by pharmacological inhibition of autophagy. Am J Cancer
Res. 5:3612–3623. 2015.PubMed/NCBI
|
|
16
|
Hanahan D and Weinberg RA: Hallmarks of
cancer: The next generation. Cell. 144:646–674. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Trout JJ, Stauber WT and Schottelius BA:
Increased autophagy in chloroquine-treated tonic and phasic
muscles: An alternative view. Tissue Cell. 13:393–401. 1981.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yang Z and Klionsky DJ: Mammalian
autophagy: Core molecular machinery and signaling regulation.
CurrOpin Cell Biol. 22:124–131. 2010.
|
|
19
|
Feng Y, He D, Yao Z and Klionsky DJ: The
machinery of macroautophagy. Cell Res. 24:24–41. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Tooze SA and Yoshimori T: The origin of
the autophagosomal membrane. Nat Cell Biol. 12:831–835. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Adem J, Ropponen A, Eeva J, Eray M,
Nuutinen U and Pelkonen J: Differential expression of Bcl-2 family
proteins determines the sensitivity of human follicular lymphoma
cells to dexamethasone-mediated and anti-BCR-mediated apoptosis. J
Immunother. 39:8–14. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Chen TC, Yu MC, Chien CC, Wu MS, Lee YC
and Chen YC: Nilotinib reduced the viability of human ovarian
cancer cells via mitochondria-dependent apoptosis, independent of
JNK activation. Toxicol In Vitro. 31:1–11. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Hsieh MJ, Chen MK, Yu YY, Sheu GT and
Chiou HL: Psoralen reverses docetaxel-induced multidrug resistance
in A549/D16 human lung cancer cells lines. Phytomedicine.
21:970–977. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
D'Anqiolillo F, Pistellia L, Noccioli C,
Ruffoni B, Piaqqi S, Scarpato R and Pistelli L: In vitro cultures
of Bituminariabituminosa: Pterocarpan, furanocoumarin and
isoflavone production and cytotoxic activity evaluation. Nat Prod
Commun. 9:477–480. 2014.PubMed/NCBI
|
|
25
|
Li M, Gao P and Zhang J: Crosstalk between
autophagy and apoptosis: Potential and emerging therapeutic targets
for cardiac diseases. Int J Mol Sci. 17:3322016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kamada Y, Yoshino K, Kondo C, Kawamata T,
Oshiro N, Yonezawa K and Ohsumi Y: Tor directly controls the Atg1
kinase complex to regulate autophagy. Mol Cell Biol. 30:1049–1058.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Hara T and Mizushima N: Role of ULK-FIP200
complex in mammalian autophagy: FIP200, a counterpart of yeast
Atg17? Autophagy. 5:85–87. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Mizushima N: The role of the Atg1/ULK1
complex in autophagy regulation. CurrOpin Cell Biol. 22:132–139.
2010.
|
|
29
|
Choi H, Merceron C, Mangiavini L, Seifert
EL, Schipani E, Shapiro IM and Risbud MV: Hypoxia promotes
noncanonical autophagy in nucleus pulposus cells independent of
MTOR and HIF1A signaling. Autophagy. 12:1631–1646. 2016. View Article : Google Scholar : PubMed/NCBI
|