Introduction
Post-operative cognitive dysfunction (POCD) is a
common syndrome in elderly patients, usually occurring in the first
several weeks or months after an operation. It results in
dysfunction of the central nervous system, including cognitive
impairment, declining learning and memory ability, information
processing disorder and delirium (1). Clinical methodological differences
between POCD studies, including the use of variable batteries of
tests, lack of control groups, loss of patients during follow-up
and inconsistent intervals between testing periods, limit their
usefulness (2). Due to the lack of
formal diagnostic criteria, as well as the subtlety of the
cognitive changes that occur, assessment and diagnosis of POCD is
difficult (3).
The causes of POCD in elderly patients are thought
to be multifactorial and complex. One potential risk factor for
POCD is the use of anesthetic agents (4). Widely used narcotics are currently
classified into inhalational and intravenous anesthetics (5). Inhalational general anesthetics such as
isoflurane and halothane have been demonstrated to increase the
risk of Alzheimer's disease (AD) in the aging brain (6), and exert a neurotoxic effect via
caspase-mediated apoptosis (7).
One proposed hypothesis is that epigenetic
regulation influenced by anesthetic may be a critical mechanism
underlying POCD (8). Despite the
similar pathological change in neurocytes with AD, the epigenetic
alteration in POCD predominantly results in memory and learning
disabilities (9). DNA
hydroxymethylation is a novel modification based on DNA methylation
catalyzed by dioxygenases. Hydroxymethylated cytosine (5hmC) is
identified as an intermediate of the active demethylation process
(10,11). 5hmC is highly distributed in the
early embryo, embryonic stem cells (12) and the nervous system (13,14).
Recent studies have revealed that the level of 5hmC is reduced by
10% after ten-eleven translocation methylcytosine dioxygenase 1
(TET1) knockdown, which can retard the proliferation of neural
progenitor cells and impair the abilities of spatial learning and
memory (15,16). Moreover, a reduction in 5hmC
modulates transcriptional activity of some genes involved in
neurogenesis in AD mice, which also indicates that 5hmC is closely
connected with memory maintenance (17,18).
Along with the TET family, ubiquitin-like with PHD and ring finger
domains 2 (Uhrf2) is also considered to be a novel regulator of
5hmC via its SET and ring finger associated domain (19,20).
Aberrant distribution of 5hmC may be one of the possible molecular
causes of POCD occurrence. The major enzymes that contribute to
5hmC metabolism affected by anesthetic in POCD are also poorly
understood. In this study DNA hydroxymethylation in the central
neural system of POCD mice was profiled in an attempt to reveal the
underlying pathogenesis of POCD caused by anesthetic.
Materials and methods
Animal study
All procedures were approved by the Institutional
Animal Care and Use Committee of Fudan University, Jinshan Hospital
(Animal protocol number 2017-32-166). A total of 170 18-month-old
outbred female C57BL/6 mice, purchased from Shanghai SLAC
Laboratory Animal Co., Ltd., were used in this study. The levels of
glucose and oxygen saturation (SpO2) in 50 µl blood isolated from
the caudal vein were determined using a biochemical analyzer
(Beckman Coulter Inc.). IL-1β levels were analyzed in a further 50
µl blood using ELISA kits (cat. no. PMLB00C; R&D Systems Inc.).
Animals were fed with standard food and water ad libitum. A
total of 150 randomly selected mice were treated with 2%
sevoflurane for 2 h (21–23) in an anesthesia chamber with a size of
25×13×13 cm. The remaining 20 mice were treated with normal air, as
a negative control. In a pilot study 2% sevoflurane treatment was
tested for 4 h, but >50% of mice died upon this condition (data
not shown).
The Morris water maze test (post-treatment 2–7 days)
and open field test (post-operative day 7) were performed as
previously described (24,25) in order to verify the POCD model. In
brief, for the Morris water maze test, a platform was set in the
center of a 0.5 m high and 1.2 m in diameter pool, 5 cm above the
water surface. Mice were trained at the same entrance for a total
of 2 min each time for a total of 6 times from the 2nd to 7th day
after sevoflurane treatment (one exposure). The motion trail, the
escape period and the distance travelled were recorded. For the
open field test, mice were put in a 30×72×72 cm box, with light
conditions that mimicked daytime. During the 5 min the mice spent
in the box, their average distance travelled, total grid crossings,
distance travelled around the central region and duration spent in
the central region were recorded. Both an escape period of <90
sec after the 7th day of sevoflurane treatment and an open field
score of <25% were considered as cognitive damage consistent
with a POCD model (26,27).
The mice were sacrificed on day 7 by cervical
dislocation. For each mouse, all brain tissues including
hippocampus, amygdaloid nucleus and cerebellum were harvested and
separated into two parts. One was fixed by 1% paraformaldehyde at
room temperature ≥24 h for DNA dot blot assay, Methylated DNA
immunoprecipitation (MeDIP) and immunofluorescence assay, while the
other is washed by cold PBS and treated with RIPA buffer (Solarbio
Inc.) or TRIzol® reagent (Invitrogen; Thermo Fisher
Scientific, Inc.) accordingly for western blotting and PCR.
DNA dot blot assay
A total of 2 µl genomic DNA was extracted from
tissues (Qiagen GmbH) and was dropped onto each nitrocellulose
membrane at a 2-fold serial dilution (0, 5, 10, 20 and 40 ng) for
dot blot assay. The spots were dried at room temperature and
incubated in TBST with antibodies against 5hmC (cat. no. ab214728;
Abcam) or 5-methylcytosine (5mC; cat. no. ab10805; Abcam) (1:500
dilution; 1 ng/ml) in 10 ml of TBS-T for 4 h overnight with gentle
shaking. The membranes were washed with TBS-T three times for 10
min each time at room temperature, followed by goat anti-mouse
IgG-HRP (1:10,000 dilution; 20 ng/ml; A0216, Beyotime Institute of
Biotechnology) or goat anti-rabbit IgG-HRP (1:10,000 dilution; 20
ng/ml; A0208, Beyotime Institute of Biotechnology) incubation in 10
ml of TBST for 1 h at room temperature with gentle shaking, and
washed again three times. BSA (New England Biolabs Inc.) was used
as a negative control. Membranes were subsequently incubated with 3
ml of ECL Western Blotting Substrate (Beyotime Institute of
Biotechnology) for 5 min in darkness at room temperature to develop
the bands by Tanon 5200 Chemiluminescence Imaging Analysis
System.
Western blot analysis
Brain tissues were homogenized in RIPA buffer and
then centrifuged at 4°C and 12,000 × g for 10 min. The protein
quantity in the supernatant was determined using a BCA protein
assay kit (E162-01; Fansbio). Equal amounts of total 40 µg protein
samples were separated by 10% sodium dodecyl sulfate-polyacrylamide
gel electrophoresis (SDS-PAGE) and transferred to polyvinylidene
fluoride membranes. The membranes were then blocked using 5%
non-fat milk in TBS at 4°C for 90 min and then incubated with the
respective primary antibodies against TET1 (1:2,000; cat. no.
ab191698; Abcam), TET2 (1:2,000; cat. no. ab124297; Abcam), Uhrf1
(1:2,000; cat no. 12387; Cell Signaling Technology, Inc.), Uhrf2
(1:2,000; cat. no. ab28673; Abcam) and β-actin (1:5,000; cat. no.
AA128; Beyotime Institute of Biotechnology) overnight at 4°C.
Membranes were washed with TBST and incubated with goat anti-mouse
and goat anti-rabbit IgG-HRP (1:10,000; 20 ng/ml, Beyotime
Institute of Biotechnology) at room temperature for 1 h. Membranes
were then treated with aBM Chemiluminescence Western Blotting Kit
(cat. no. 11520709001; EMD Millipore), and the bands were captured
to evaluate the difference of protein expression.
Immunofluorescence assay
In brief, mouse tissue was fixed using 4%
paraformaldehyde at room temperature for 24 h then dehydrated with
50–90% ethanol. Samples were permeabilized in 50% xylene-ethanol at
room temperature for 30 min then in 50% xylene-paraffin at 60°C for
15 min and embedded. Tissues were cut into 5 µm sections and washed
with 0.3% H2O2-methanol for 10 min then
heated to 92°C for 40 min for antigen retrieval. Following PBS
washing, sections were blocked with 5% BSA (Sigma-Aldrich; Merck
KGaA) for 30 min at 37°C. Subsequently, slices were incubated with
5hmC (1:200; cat. no. ab214728; Abcam) and Uhrf2 (1:200; cat no.
ab28673; Abcam) antibodies overnight at 4°C. After washing, tissues
were further incubated with the appropriate Alexa Fluor secondary
antibody (cat. no. A32732; Thermo Fisher Scientific, Inc.) at
1:20,000 dilution for 30 min at room temperature. After washing,
cells were mounted in mounting media with DAPI (Vector Laboratories
Inc.). The positive staining of 5hmC and Uhrf2 were digitally
captured by Olympus BX51 (Olympus Corporation) at ×400
magnification and analyzed using ImageJ software v1.8.0 (National
Institutes of Health).
MeDIP quantitative (q) PCR assay
MeDIP assay was performed as previously described
(28). Extracted genomic DNA was
sonicated (90 cycles of 30 sec on/30 sec off with high power;
UCD-300; Bioruptor) and incubated with 0.5 µg 5hmC (cat. no.
ab214728; Abcam) or 5mC (cat. no. ab10805; Abcam) antibody or IgG
(cat. no. A0208, Beyotime Institute of Biotechnology) as control
overnight to capture the DNA fragment with 5hmC or 5mC, then washed
and harvested for detection of the 5hmC or 5mC enrichment at
promoter regions of candidate genes, where the primers for qPCR
were designed to encompass ~200 bp (Table I). The qPCR reactions were done using
the Fast Universal SYBR Green Realtime PCR Master Mix (Roche
Diagnostics) and in triplicate under the following conditions:
95°Cv for 30 sec; 40 cycles of 95°C for 5 sec and 60°C for 30 sec.
Ct value was analyzed to calculate enrichment using the
2−ΔΔCq method (29).
 | Table I.Sequences of primers used. |
Table I.
Sequences of primers used.
| A, Med IP assay |
|---|
|
|---|
| Target | Primer sequence
(5′-3′) | Length of PCR
products |
|---|
| ACSS2 |
CCCAGACCATAACAGTACCGACTC | 294 |
|
|
CGCCTTTGCCATTCATAGAGC |
|
| BDNF |
TTAGAGGAGGTGTAGCCTTGTT | 223 |
|
|
TTGTCATCACAGTGGGAAGC |
|
| CCL2 |
AAAGTTTCCATTGCTGCTGCTC | 106 |
|
|
TCTGATGTAACGGGCTCTTGG |
|
| FAS |
ACTTCCCTACCCACCCATTC | 220 |
|
|
AAAGTACCCAAGGAGCTAAAGG |
|
| GCR |
ATTGCCCTGGATGCCTGTAA | 321 |
|
|
ATGACCATGAACCTCCTGAA |
|
| GDNF |
ATGGCTCTATGCTGCTTTGC | 163 |
|
|
TATCCCAGACGTGGACTTGC |
|
| GLUR2 |
GAGGGTGGAATGGGAAAGAG | 197 |
|
|
AGGCAGCTACCAAATGTCTCG |
|
| HMGB1 |
TTTTCCTTCTTTGGGTCTAA | 217 |
|
|
CCAGCCTAACTCTGCTTCCT |
|
| MMP9 |
TGGTTTCAGAAGAGGAGGACAGG | 248 |
|
|
GCAGCGAGGAACAGGGAGCA |
|
|
| B, qPCR
assay |
|
| Target | Primer sequence
(5′-3′) | Length of PCR
products |
|
| ACSS2 |
TCCCATTCTTCGGTGTAGCG | 376 |
|
|
GTAACAAAGCAGTAGAGGCATTCG |
|
| BNDF |
GTGGGTCACAGCGGCAGATA | 203 |
|
|
ACGATTGGGTAGTTCGGCATT |
|
| CCL2 |
AAGAAGGAATGGGTCCAGACA | 140 |
|
|
GCTTCAGATTTACGGGTCAACT |
|
| FAS |
TCTGGGCTGTCCTGCCTCTG | 111 |
|
|
CAGTTTCACGAACCCGCCTC |
|
| GCR |
TGGAATAGGTGCCAAGGGTC | 171 |
|
|
GCAGAGTTTGGGAGGTGGTC |
|
| GDNF |
GATGAAGTTATGGGATGTCGTGG | 175 |
|
|
TGCCGCTTGTTTATCTGGTGA |
|
| GLUR2 |
GAAGCCTCAGAAGTCCAAACC | 342 |
|
|
TTAGCCGTGTAGGAGGAGATG |
|
| HMGB1 |
ATCCTGGCTTATCCATTGGTG | 244 |
|
|
TCCTCATCCTCTTCATCCTCCT |
|
| MMP9 |
AAGGGTACAGCCTGTTCCTGGTG | 146 |
|
|
GATGCCGTCTATGTCGTCTTTATTCA |
|
Reverse transcription-quantitative
(RT-q)PCR
Total RNA was extracted using TRIzol®
reagent according to the manufacturer's instructions. Before
performing RT, RNA was treated with 5U DNase I (Beyotime Institute
of Biotechnology) on ice for 10 min to remove bacterial genomic
DNA, and purified using isopropanol and 3M sodium acetate, before
washing with 75% ice ethanol. Reverse transcription was performed
as the conditions of 42°C for 15 min and 95°C for 3 min using the
QuantiTect Reverse Transcription kit (Qiagen GmbH). qPCR were
performed using Fast Universal SYBR Green Realtime PCR Master
(Roche Diagnostics) with the conditions followed were 95°C for 30
sec; 40 cycles of 95°C for 5 sec and 60°C for 30 sec using the
primers listed in Table I.
Statistical analysis
To distinguish between the non-POCD and POCD groups,
the escape latency values of control group at post-operative day 7
were collected and fitted a normal distribution as µ ± σ. Normal
samples were distributed within the range from µ-3σ to µ+3σ by
99.73% probability. Thus, the values included or excluded in µ ± 3σ
were considered as non-POCD or POCD respectively. Likewise, the
values of distance travelled as well as the data of open field test
including average distance, grid crossings numbers, distance among
central region and staying time among central region were all
analyzed in the same way. Finally, the intersection of the
potential POCD from each type of parameters was confirmed as the
POCD group. Data are presented as the mean ± standard deviation for
multiple independent experiments. The multiple comparisons of the
difference between values were analyzed using one-way ANOVA. The
pairwise comparisons of control group and POCD/non-POCD groups were
analyzed using Fisher's Least Significant Difference post hoc test
following ANOVA. Pearson's correlation analysis was used to
evaluate the association between 5mC and 5hmC and the mRNA level.
P<0.05 was considered to indicate a statistically significant
difference.
Results
5hmC expression is altered in mice
with cognitive dysfunction
Initial levels of blood glucose, SpO2 and IL-1β were
analyzed in order to rule out respiratory depression, infection or
hypoglycemia, because these physical signs were subject to POCD
occurrence (Table II). A total of
150 mice were treated with 2% sevoflurane in order to establish a
POCD model. A total of 18 of these mice were identified as having
POCD via both Morris water maze test and an open field test daily
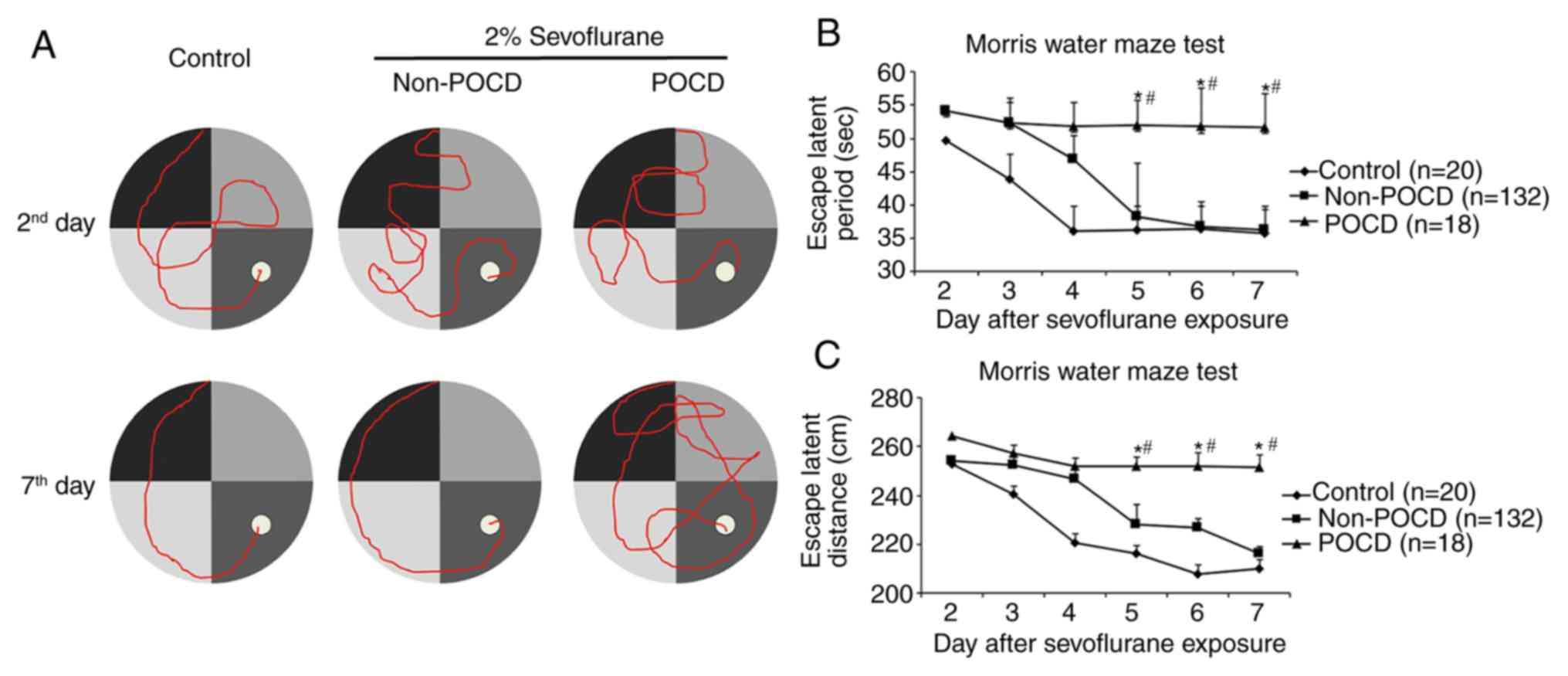
between 2 and 7 days post-operatively (Fig. 1A; Table
II). Unlike POCD in humans, which happens most conspicuously on
post-operative day 7 and at post-operative month 3 (30), POCD mice started to display
increasing escape latency and travelled distance from the 5th day
when compared to control and non-POCD mice (Fig. 1B and C). The mice were sacrificed at
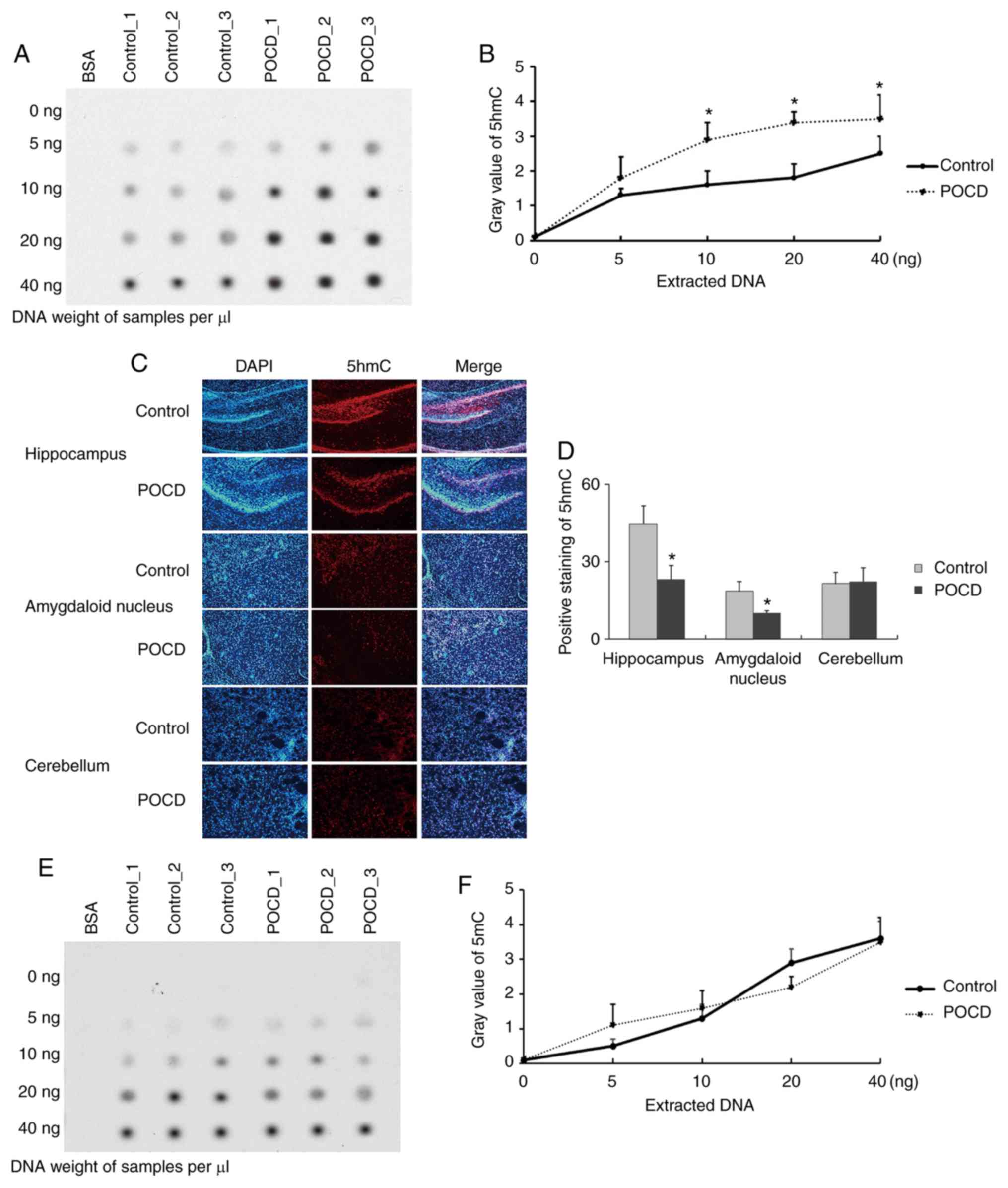
day 7 and the brain tissues harvested to detect global 5hmC
expression via dot blot assay. The presence of differential change
of global 5hmC expression in whole brain lysate before and after
POCD was observed (n=3; Fig. 2A and
B). Furthermore, the hippocampus, amygdaloid nucleus and
cerebellum were also harvested on day 7 and it was observed that
5hmC was significantly lower in POCD mice compared to control in
the hippocampus and amygdaloid nucleus, but no obvious change was
seen in the cerebellum (n=5; Fig. 2C and
D). There were no differences in 5mC level between POCD and
control (n=5; Fig. 2E and F). Taken
together, the loss of 5hmC in both the hippocampus and amygdaloid
nucleus was observed in POCD mice and it may be responsible for the
cognitive impairment seen, including the loss of abilities of
memory, spatial learning, and new environmental adaptation.
| Table II.Principle indexes of an open field
test. |
Table II.
Principle indexes of an open field
test.
| Group | Sample size | Weight (g) | Blood glucose
(mmol/l) | SpO2 (%) | IL-1β (pg/m) | Average distance
(cm) | Total across
grids | Distance among
central region (cm) | Staying time among
central region (s) |
|---|
| Control | 20 | 23.5±3.2 | 9.4±2.1 | 96.13±1.36 | 14.87±2.09 | 13.389±3.8 | 84±12 | 1.895±0.44 | 2.926±0.89 |
| Non-POCD | 132 | 23.7±1.8 | 10.1±3.3 | 95.87±2.14 | 15.39±1.56 | 12.354±4.9 | 81±20 | 1.754±0.33 | 3.179±1.04 |
| POCD | 18 | 23.8±2.5 | 9.8±2.8 | 95.61±1.08 | 15.14±1.91 |
8.94±2.2a,b | 54±14a,b |
1.246±0.38a,b |
2.67±2.44a,b |
Loss of Uhrf2 is responsible for 5hmC
alteration in hippocampus of POCD mice
To elucidate the hydroxymethylation impacted by
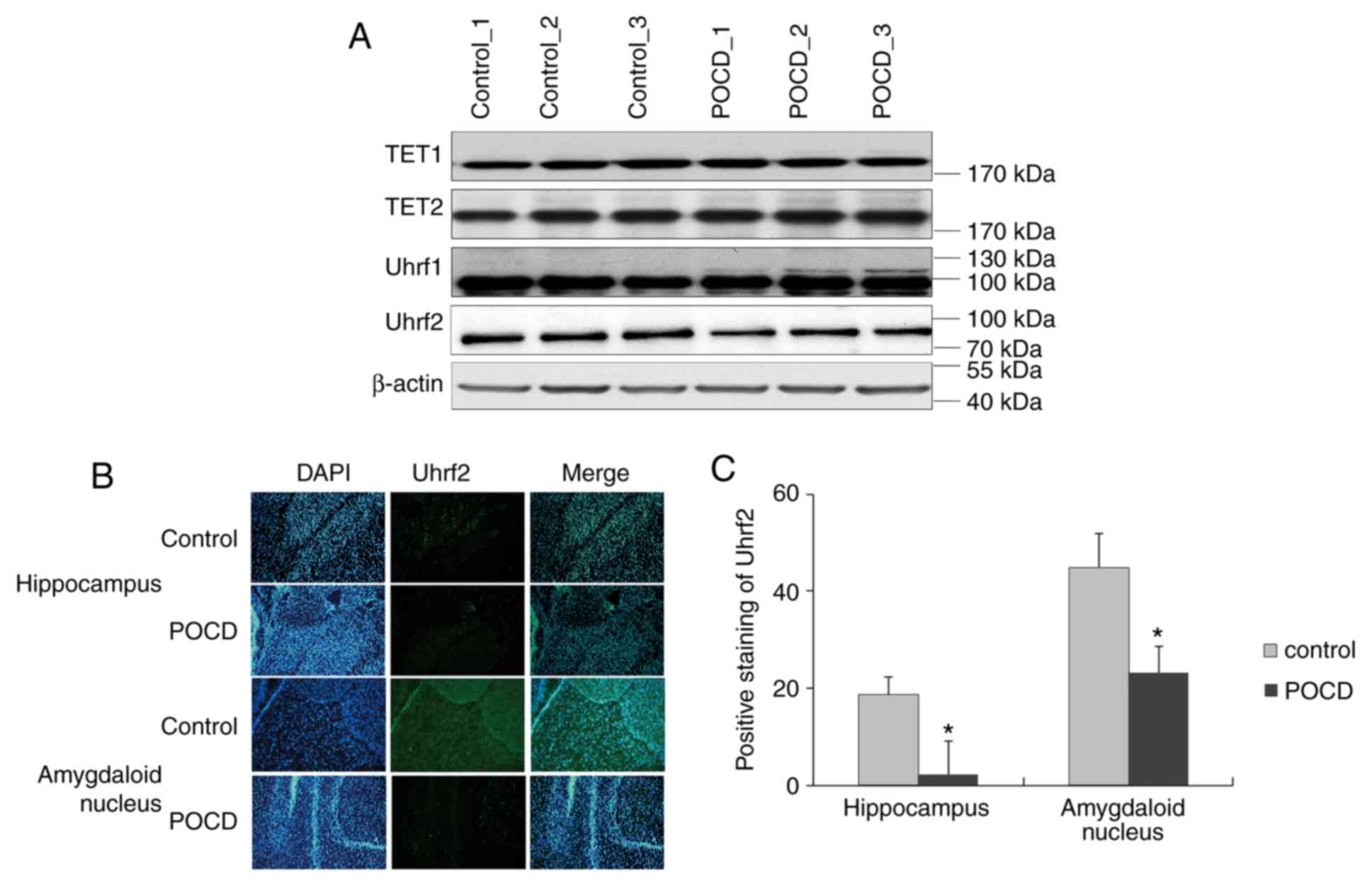
POCD, the enzymes Uhrf1, Uhrf2, TET1 and TET2 for 5hmC were
investigated via western blotting. The protein levels of Uhrf1,
TET1 and TET2 exhibited no significant differences between control
and POCD in whole brain, whilst Uhrf2 displayed a slight
downregulation in POCD (n=5; Fig.
3A). When considering the background noise from the whole
brain, Uhrf2 was further investigated in specific regions of brain
and observed a suppression in hippocampus and amygdaloid nucleus in
POCD compared with control (n=5; Fig. 3B
and C), coinciding with the change of 5hmC. Collectively, Uhrf2
was suppressed by sevoflurane for hindrance of 5hmC in POCD.
POCD results in 5hmC reduction at the
promoter regions of genes associated with neurodevelopment
The present study hypothesized that Uhrf2 may be
responsible for 5hmC maintenance in hippocampus and amygdaloid
nucleus. To further validate the role of 5hmC in POCD, the local
5mC and 5hmC enrichment at promoter regions of genes associated
with neurodevelopment was detected through MedIP-qPCR. Glial
cell-derived neurotropic factor (GDNF), brain-derived neurotrophic
factor (BDNF), glucocorticoid receptor (GCR) and acyl-CoA
synthetase short chain family member 2 (ACSS2) displayed a reduced
5hmC (Fig. 4A) and unvaried 5mC
(Fig. 4B) level on their promoters
in POCD compared with control (control n=5; POCD n=6). Compared
with their transcriptional levels (Fig.
4C), 5hmC levels on the promoters of GDNF, BDNF, GCR and ACSS2
could reflect the transcriptional activation of these genes
(Figs. 4D and S1). While 5mC levels on the promoters of
MMP9, HGMB1 and GLUR2 showed an inverse tendency with transcription
of these genes (Fig. 4B and D).
Additionally, neither 5hmC nor 5mC of FAS and CCL2 had any
correlation with their mRNA change in POCD compared to control.
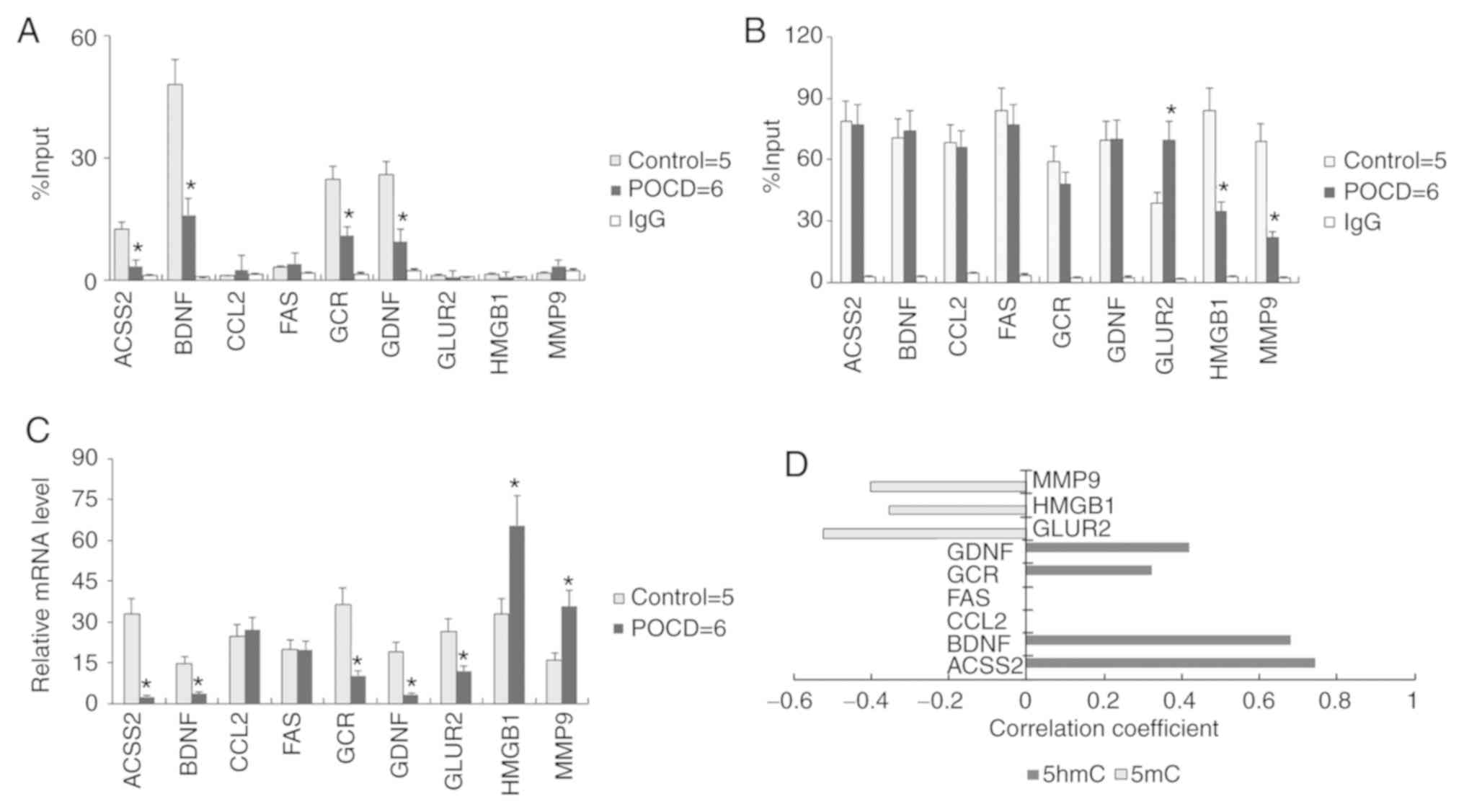 | Figure 4.Association between 5hmC and gene
transcription. Local (A) 5hmC and (B) 5mC enrichment at promoter
regions of genes associated with neurodevelopment. (C)
Transcriptional level of nine genes in mouse brain. (D) The
significant difference P<0.05 between 5mC or 5hmC enrichment and
mRNA level are listed. All data are presented as the mean ±
standard deviation of the mean of individual experiments (control
for five; POCD for six duplications). *P<0.05 vs. control group.
5hmC, 5′hydroxymethylcytosine; 5mC, 5′methylcytosine; ACSS2,
acyl-CoA synthetase short chain family member 2; BDNF, brain
derived neurotrophic factor; CCL2, C-C motif chemokine ligand 2;
FAS, fas cell surface death receptor; GCR, glucocorticoid receptor;
GDNF, glial cell-derived neurotrophic factor; GluR2, glutamate
receptor 2 precursor; HMGB1, high mobility group protein B1; MMP9,
matrixmetallopeptidase-9. |
Discussion
In the present study, 2% sevoflurane was used to
establish a POCD model. Mouse models of POCD displayed signs of
behavioral and memory problems (27,31), as
indicated by the results of water maze and open field tests. By
contrast, the results for the non-POCD group were similar to the
normal control. Although certain phenotypes have been studied to
classify early POCD, such as serum proteomics (32), cerebrospinal fluid (33), and cerebral oxygen saturation
(34), no clear test for the
susceptibility of individuals to POCD is available.
Due to a lack of potent drug for the treatment of
POCD in clinical practice, a previous study has prompted the use of
DNA methyltransferase inhibitors, which can restore
memory-associated transcriptional regulation and improve behavioral
memory function in elderly animals (8). However, the role of epigenetic factors
in POCD has not been previously investigated to the best of our
knowledge. The current study demonstrated an association between
5hmC and POCD, and aimed to clarify the role of hydroxymethylation
in POCD for the regulation of memory and learning ability. The
global 5hmC level in brain was reduced in POCD. However, of the
different brain regions, 5hmC levels in hippocampus and amygdaloid
nucleus declined the most markedly in the POCD group, which implies
that 5hmC may contribute to memory and learning ability in the
hippocampus (35) as well as fear
emotion control in the amygdaloid nucleus (36). However, the presence of unaltered
5hmC levels in the cerebellum before and after POCD, suggests that
the basic associative learning and memory from cerebellum impaired
by POCD (37) may be independent of
accumulation or loss of hydroxymethylation (37). The expression levels of
hydroxymethylation associated enzymes such as Uhrf1 and TETs
remained unchanged, while the decreased protein levels of Uhrf2 in
POCD were consistent with 5hmC levels both in the hippocampus and
the amygdaloid nucleus. A previous study reported that TET1
knockout did not affect overall brain morphology in mice, and
concluded that TET1 deletion could enhance the consolidation and
storage of threat recognition (cued and contextual fear
conditioning) and object location memories (38), which is apparently reciprocal with
the phenotype of POCD. It is therefore unlikely that TET levels
exert an effect in POCD. A recent study reported that loss of Uhrf2
reduced 5hmC in the brain, including the cortex and hippocampus,
but did not change 5mC level, and that mice exhibited a partial
impairment in spatial memory acquisition and retention (20), which was further confirmed by the
current results. The results of the present study suggest that
Uhrf2 may be a target responding to 5hmC regulation in POCD.
The relationship between 5hmC enrichment and gene
transcription in POCD was further investigated. Two epigenetic
means of gene transcriptional regulation closely associated with
neurodevelopment, DNA methylation-mediated gene silencing and loss
of DNA hydroxymethylation-mediated gene silencing were identified
in the current study. 5hmC was reduced in ACSS2, BDNF, GCR and
GDNF, while 5mC levels for these genes remained unaltered, which
indicated that sevoflurane can suppress Uhrf2 to compromise the
5hmC modification on the promoter regions of these genes associated
with neuroprotection and proliferation and thereby repress the
transcriptional activity. Moreover, the change of expression of
GLUR2, HMGB1 and MMP9 were significantly negative correlated with
their DNA methylation change at promoter level. This may be due to
a change in DNA methylation patterns in their promoters.
The molecular mechanism underlying the differences
between non-POCD and POCD remains to be elucidated. We hypothesized
that it depends on individual factors, such as neuroplasticity,
immunity and the sensitivity to sevoflurane. However, it is
difficult to distinguish differences in the epigenetic patterns of
brain tissues from a group of animals as variations exist between
each individual. Once POCD develops, samples of the diseased brain
in its pre-diseased normal state cannot be obtained. A further
limitation is that incidence of POCD in both humans and mouse
models is low. In addition, sevoflurane treatment may not be an
optimal model for POCD induction. In future studies a rapid method
for delirium induction by scopolamine (39) or another sedative may be a better way
to mimic and further study POCD. An additional limitation of this
study is that genetic editing (over-expression or knockdown in
vivo) operation of the 5hmC metabolism associated enzymes such
as Tet1/2/3 or Uhrf1/2 was not performed to further investigate the
sevoflurane-induced effect of learning and memory impairment as
well as 5hmC loss in POCD model.
Overall, the current data suggest that sevoflurane
may lead to the suppression of Uhrf2 and induce the loss of global
5hmC in the hippocampus and amygdaloid nucleus, thereby impairing
the learning and memory ability of mice with POCD. The present
study revealed a novel connection between 5hmC, which is an
important biomarker of memory and POCD. The current findings may
provide a new biomarker to target or inform the development of a
new anesthetic to reduce the incidence of POCD.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
This study was supported by the Science &
Technology Commission of Jinshan District, Shanghai (grant no.
2017-3-09).
Availability of data and materials
The data sets used and/or analyzed during the
current study are available from the corresponding author on
reasonable request.
Authors' contributions
JZ performed experiments and analyzed the data. WX
designed the project and drafted the paper.
Ethics approval and consent to
participate
This study was approved by the Institutional Animal
Care and Use Committee of Fudan University, Shanghai (animal
protocol number 2017-32-166) and all animal protocols were
conducted following the guidelines accordingly.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Pappa M, Theodosiadis N, Tsounis A and
Sarafis P: Pathogenesis and treatment of post-operative cognitive
dysfunction. Electron Physician. 9:3768–3775. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Funder KS, Steinmetz J and Rasmussen LS:
Methodological issues of postoperative cognitive dysfunction
research. Semin Cardiothorac Vasc Anesth. 14:119–122. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Monk TG and Price CC: Postoperative
cognitive disorders. Curr Opin Crit Care. 17:376–381. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Tzimas P, Samara E, Petrou A, Korompilias
A, Chalkias A and Papadopoulos G: The influence of anesthetic
techniques on postoperative cognitive function in elderly patients
undergoing hip fracture surgery: General vs spinal anesthesia.
Injury. 49:2221–2226. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Amare M, McEvoy M and Smith A: The effect
of intravenous and inhalational maintenance of anaesthesia on
postoperative cognitive outcomes in elderly people. Anaesthesia.
74:10682019. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Eckenhoff RG, Johansson JS, Wei H, Carnini
A, Kang B, Wei W, Pidikiti R, Keller JM and Eckenhoff MF: Inhaled
anesthetic enhancement of amyloid-beta oligomerization and
cytotoxicity. Anesthesiology. 101:703–709. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Xie Z, Culley DJ, Dong Y, Zhang G, Zhang
B, Moir RD, Frosch MP, Crosby G and Tanzi RE: The common inhalation
anesthetic isoflurane induces caspase activation and increases
amyloid beta-protein level in vivo. Ann Neurol. 64:618–627. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wang Y, Chen Z, Zhao Y, Shi R, Wang Y, Xu
J, Wu A, Johns RA and Yue Y: Epigenetics as a new therapeutic
target for postoperative cognitive dysfunction. Med Hypotheses.
80:249–251. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Sweatt JD: Neuroscience. Epigenetics and
cognitive aging. Science. 328:701–702. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wu X and Zhang Y: TET-mediated active DNA
demethylation: Mechanism, function and beyond. Nat Rev Genet.
18:517–534. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ito S, Shen L, Dai Q, Wu SC, Collins LB,
Swenberg JA, He C and Zhang Y: Tet proteins can convert
5-methylcytosine to 5-formylcytosine and 5-carboxylcytosine.
Science. 333:1300–1303. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ito S, D'Alessio AC, Taranova OV, Hong K,
Sowers LC and Zhang Y: Role of Tet proteins in 5mC to 5hmC
conversion, ES-cell self-renewal and inner cell mass specification.
Nature. 466:1129–1133. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Mellén M, Ayata P, Dewell S, Kriaucionis S
and Heintz N: MeCP2 binds to 5hmC enriched within active genes and
accessible chromatin in the nervous system. Cell. 151:1417–1430.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kriaucionis S and Heintz N: The nuclear
DNA base 5-hydroxymethylcytosine is present in Purkinje neurons and
the brain. Science. 324:929–930. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhang RR, Cui QY, Murai K, Lim YC, Smith
ZD, Jin S, Ye P, Rosa L, Lee YK, Wu HP, et al: Tet1 regulates adult
hippocampal neurogenesis and cognition. Cell Stem Cell. 13:237–245.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Rudenko A, Dawlaty MM, Seo J, Cheng AW,
Meng J, Le T, Faull KF, Jaenisch R and Tsai LH: Tet1 is critical
for neuronal activity-regulated gene expression and memory
extinction. Neuron. 79:1109–1122. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Shu L, Sun W, Li L, Xu Z, Lin L, Xie P,
Shen H, Huang L, Xu Q, Jin P and Li X: Genome-wide alteration of
5-hydroxymenthylcytosine in a mouse model of Alzheimer's disease.
BMC Genomics. 17:3812016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Bernstein AI, Lin Y, Street RC, Lin L, Dai
Q, Yu L, Bao H, Gearing M, Lah JJ, Nelson PT, et al:
5-Hydroxymethylation-associated epigenetic modifiers of Alzheimer's
disease modulate Tau-induced neurotoxicity. Hum Mol Genet.
25:2437–2450. 2016.PubMed/NCBI
|
|
19
|
Zhou T, Xiong J, Wang M, Yang N, Wong J,
Zhu B and Xu RM: Structural basis for hydroxymethylcytosine
recognition by the SRA domain of UHRF2. Mol Cell. 54:879–886. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Chen R, Zhang Q, Duan X, York P, Chen GD,
Yin P, Zhu H, Xu M, Chen P, Wu Q, et al: The
5-Hydroxymethylcytosine (5hmC) reader UHRF2 is required for normal
levels of 5hmC in mouse adult brain and spatial learning and
memory. J Biol Chem. 292:4533–4543. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Fang F, Lin W, Ling X, Song R, Liu Q, Lai
B and Cang J: The hippocampal cyclin D1 expression is involved in
postoperative cognitive dysfunction after sevoflurane exposure in
aged mice. Life Sci. 160:34–40. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Murphy KL, McGaughy J, Croxson PL and
Baxter MG: Exposure to sevoflurane anesthesia during development
does not impair aspects of attention during adulthood in rats.
Neurotoxicol Teratol. 60:87–94. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Guo S, Liu L, Wang C, Jiang Q, Dong Y and
Tian Y: Repeated exposure to sevoflurane impairs the learning and
memory of older male rats. Life Sci. 192:75–83. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Rosczyk HA, Sparkman NL and Johnson RW:
Neuroinflammation and cognitive function in aged mice following
minor surgery. Exp Gerontol. 43:840–846. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Hovens IB, Schoemaker RG, van der Zee EA,
Absalom AR, Heineman E and van Leeuwen BL: Postoperative cognitive
dysfunction: Involvement of neuroinflammation and neuronal
functioning. Brain Behav Immun. 38:202–210. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Stanford SC: The open field test:
Reinventing the wheel. J Psychopharmacol. 21:134–135. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Tang Y, Wang X, Zhang S, Duan S, Qing W,
Chen G, Ye F, Le Y and Ouyang W: Pre-existing weakness is critical
for the occurrence of postoperative cognitive dysfunction in mice
of the same age. PLoS One. 12:e01824712017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhou S, Shen Y, Zheng M, Wang L, Che R, Hu
W and Li P: DNA methylation of METTL7A gene body regulates its
transcriptional level in thyroid cancer. Oncotarget. 8:34652–34660.
2017.PubMed/NCBI
|
|
29
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Johnson T, Monk T, Rasmussen LS,
Abildstrom H, Houx P, Korttila K, Kuipers HM, Hanning CD, Siersma
VD, Kristensen D, et al: Postoperative cognitive dysfunction in
middle-aged patients. Anesthesiology. 96:1351–1357. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Almahozi A, Radhi M, Alzayer S and Kamal
A: Effects of memantine in a mouse model of postoperative cognitive
dysfunction. Behav Sci (Basel). 9(pii): E242019. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhang Q, Li SZ, Feng CS, Qu XD, Wang H,
Zhang XN, Liu Y, Wang Y, Wu AS and Yue Y: Serum proteomics of early
postoperative cognitive dysfunction in elderly patients. Chin Med J
(Engl). 125:2455–2461. 2012.PubMed/NCBI
|
|
33
|
Ji MH, Yuan HM, Zhang GF, Li XM, Dong L,
Li WY, Zhou ZQ and Yang JJ: Changes in plasma and cerebrospinal
fluid biomarkers in aged patients with early postoperative
cognitive dysfunction following total hip-replacement surgery. J
Anesth. 27:236–242. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Lin R, Zhang F, Xue Q and Yu B: Accuracy
of regional cerebral oxygen saturation in predicting postoperative
cognitive dysfunction after total hip arthroplasty: Regional
cerebral oxygen saturation predicts POCD. J Arthroplasty.
28:494–497. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Webb WM, Sanchez RG, Perez G, Butler AA,
Hauser RM, Rich MC, O'Bierne AL, Jarome TJ and Lubin FD: Dynamic
association of epigenetic H3K4me3 and DNA 5hmC marks in the dorsal
hippocampus and anterior cingulate cortex following reactivation of
a fear memory. Neurobiol Learn Mem. 142:66–78. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Martin-Fernandez M, Jamison S, Robin LM,
Zhao Z, Martin ED, Aguilar J, Benneyworth MA, Marsicano G and
Araque A: Synapse-specific astrocyte gating of amygdala-related
behavior. Nat Neurosci. 20:1540–1548. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Jungwirth B, Zieglgänsberger W, Kochs E
and Rammes G: Anesthesia and postoperative cognitive dysfunction
(POCD). Mini Rev Med Chem. 9:1568–1579. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Kumar D, Aggarwal M, Kaas GA, Lewis J,
Wang J, Ross DL, Zhong C, Kennedy A, Song H and Sweatt JD: Tet1
oxidase regulates neuronal gene transcription, active DNA
hydroxymethylation, object location memory and threat recognition
memory. Neuroepigenetics. 4:12–27. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Qiu Y, Chen D, Huang X, Huang L, Tang L,
Jiang J, Chen L and Li S: Neuroprotective effects of HTR1A
antagonist WAY-100635 on scopolamine-induced delirium in rats and
underlying molecular mechanisms. BMC Neurosci. 17:662016.
View Article : Google Scholar : PubMed/NCBI
|