Introduction
Globally, the number of cancer cases rose by 33%
between 2005 and 2015, and the morbidity and mortality of cancer
increases year on year (1). Data,
derived from the latest global cancer statistics, indicates lung
cancer is the most common diagnosis of cancer (11.6% of total
cases) and the main cause of cancer death (18.4% of total cancer
deaths). Colorectal cancer is also common (6.1% of total cases) and
is a major cause of cancer-related deaths (9.2% of total cases)
(2). Surgery is the mainstay
treatment strategy for patients with lung and colorectal cancer.
Unfortunately, metastatic recurrence after tumor resection remains
common and carries a high risk of mortality (3). Preventing tumor cells disseminating
during surgery still poses a major challenge for perioperative
clinicians.
Hypoxia is one of the typical characteristics of
solid tumors due to the excessive proliferation of cancer cells and
their increasing oxygen consumption (4). It is well established that hypoxia
promotes survival and increases the invasive ability of cancer
cells (5). Secondary hypoxia as a
result of treatment with anesthetic drugs or anesthetic techniques
is a common event during the perioperative period (6,7). Recent
findings have suggested that anesthetic drugs might have a
significant impact on cancer invasion and progression (8,9).
However, the effects of anesthetic agents on the aggressiveness of
cancer cells in hypoxia are still largely undefined.
Dexmedetomidine (DEX), a highly selective
α2-adrenergic receptor agonist, is used in clinical
anesthesia settings as both a sedative and analgesic agent.
Compelling evidence exists that demonstrates that DEX is able to
exert an anti-apoptotic and protective function on cells and organs
(10–13); however, the effects of DEX on the
invasiveness of cancer cells remains unknown. Several studies have
reported that DEX contributes to the growth and metastatic
potential of breast cancer cells (14–16). On
the contrary, Deng et al (17) found that DEX has no effect on the
migration of colorectal cancer cells. Atipamezole, a selective and
specific α2 receptor-antagonist, has been identified as
a drug capable of reversing DEX-induced effects (18). In the present study atipamezole was
used as a control for DEX during the observation of the effect of
DEX on hypoxia-induced growth and the metastatic potential of
cancer cells.
In the present study, the effects of DEX on the
progression of cancer cells were investigated in the context of
hypoxia. Our results revealed that DEX promotes hypoxia-induced
growth and may promote the metastasis of lung cancer cells and
colorectal cancer cells by regulating hypoxia inducible factor
(HIF)-1α signaling, which may be associated with the α2
adrenoceptor-dependent pathway.
Materials and methods
Cell culture
The human lung adenocarcinoma A549 cell line and
human colorectal cancer cell line HCT116 were obtained from the
Cell Bank of Type Culture Collection of the Chinese Academy of
Sciences. A549 cells and HCT116 cells were cultured in DMEM (Thermo
Fisher Scientific, Inc.) supplemented with 10% FBS (Thermo Fisher
Scientific, Inc.), 100 U/ml penicillin and 100 µg/ml streptomycin
under a humidified atmosphere of 5% CO2 at 37°C. The
media was replaced every 2 or 3 days. Cells were passaged three
times a week by treating with trypsin-EDTA. Cells in the
logarithmic growth phase were used for the experiments.
Cell treatment
For hypoxia incubation, cells were cultured in
airtight glass chambers that were infused with a mixture of 1%
O2, 5% CO2 and 94% N2 at 37°C as
described previously (19). A
concentration of 1% O2 was chosen because it is a
classical model for hypoxic insult to cells (20,21). The
re-oxygenation treatment after hypoxia was performed by incubating
the cells at 37°C in a humidified 5% CO2 atmosphere.
Cells in the exponential growth phase were plated, and were
cultured at 37°C in 5% CO2 for 24 h. Cells in the
normoxia (N) group were incubated at 37°C with 95% atmospheric
air/5% CO2 at 6 l/min for 4 h. Cells in the hypoxia (H)
group were incubated at 37°C in 94% N2/5%
CO2/1% O2 at 6 l/min for 4 h. Cells in the
hypoxic + DEX (HD) group were treated with 1 nM DEX
(Aibeining®; Jiangsu Hengrui Medicine Co., Ltd.) and
incubated at 37°C in 94%N2/5% CO2/1%
O2 at 6 l/min for 4 h. Cells in the hypoxia + DEX +
atipamezole (HDA) group were treated with 1 nM DEX, 10 nM
atipamezole (Sigma-Aldrich; Merck KGaA), at 37°C in 94%
N2/5% CO2/1% O2 at 6 l/min for 4
h. The duration of DEX treatment chosen was 4 h in the present
study to model surgeries, which based on observations at Affiliated
FoShan Hospital of Sun Yat-Sen University, China typically last 2–6
h.
MTT assay
MTT assay was used to examine cell viability. Cells
were incubated at 37°C in 96-well plates (1×104
cells/well) overnight. Following treatment, the cells were further
cultured at 37°C in DMEM supplemented with 10% FBS for 48 h prior
to the addition of MTT to each well. Thereafter, MTT was added to
each well and mixed. The cells were then incubated for an
additional 4 h. The media was removed and DMSO was added to each
well to fully dissolve the blue crystals. Absorbance was then
measured at 570 nm (optical density 570) using a
spectrophotometer.
Transwell assay
Transwell assays were used to evaluate the invasion
and migration ability of cells. The upper chambers of transwell
plates (24-well; Costar; Corning, Inc.) were coated with 100 µl of
0.2 mg/ml Matrigel (Corning, Inc.) and left to dry overnight at
room temperature. Following treatment, A549 cells and HCT116 cells
were seeded in the upper chamber (1×105 cells/well) of
these Matrigel-coated 24-well Transwell plate for 24 h. DMEM
containing 10% FBS was loaded into the lower well (600 µl/well) and
serum-free medium was loaded into the upper chamber. The cells were
cultured at 37°C in 5% CO2 for 24 h. Cells that did not
pass through the polycarbonate membrane at the bottom of the
chamber were scraped gently with a cotton swab. The membrane was
removed and fixed with methanol for 15 min and stained with 0.5%
crystal violet at room temperature for 20 min. The number of
invasive cells was counted from five fields of vision that were
selected randomly using an inverted light microscope (MSHOT/MC30
Digital imaging system; Guangzhou Micro-shot Technology Co., Ltd.)
under ×200 magnification. The migration ability of cells was
determined following the same approach but without the Matrigel
coating.
Western blotting
Cells were washed with PBS, lysed on ice for 30 min
with lysis buffer [containing 20 mM Tris-HCl pH 7.5, 150 mM NaCl, 1
mM EDTA, 1 mM EGTA, 1% (v/v) Triton X-100, 0.5% (v/v) Nonidet P40,
2.5 mM sodium pyrophosphate, 1 mM sodium orthovanadate, 50 mM
sodium fluoride and 1 × protease inhibitor cocktail] and then
centrifuged at 18,894 × g for 15 min at 4°C. The supernatants were
collected and assessed for protein concentration using a BCA assay.
Proteins (50 µg per sample) were separated on 12% SDS-PAGE gels,
and then electrophoretically transferred onto a polyvinylidene
difluoride membrane (EMD Millipore). Membranes were blocked with 5%
BSA (Shanghai Rong Biological Technology Co., Ltd.) and with TBST
(0.05% Tween 20) at 4°C for 1 h. The membrane was then incubated
overnight at 4°C with anti-survivin antibodies [Santa Cruz
Biotechnology, Inc.; mouse monoclonal antibody (mAb); cat. no.
sc-17779; dilution 1:500], anti-HIF-1α antibody (Santa Cruz
Biotechnology, Inc.; mouse mAb; cat. no. sc-13515; dilution 1:500),
and anti-GAPDH antibody (Santa Cruz Biotechnology, Inc.; mouse mAb;
cat. no. sc-365062; dilution 1:1,000), followed by incubation at
4°C with fluorescence-conjugated secondary antibodies (Santa Cruz
Biotechnology, Inc.; normal anti-mouse IgG Alexa Fluor®
680; cat. no. sc-516610; dilution 1:400) for 2 h. The protein
signals were analyzed using an Odyssey IR scanner (LI-COR
Biosciences), and signal intensities were quantified using NIH
ImageJ software (v.16.0; National Institutes of Health). Expression
levels of each protein were measured relative to GAPDH.
Reverse transcription-quantitative
RT-qPCR
For the detection of the levels of matrix
metalloproteinase (MMP)-2 and MMP-9 expression in the two cells,
RT-qPCR was performed. Cell mRNA was isolated with
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.), and cDNA was synthesized with the M-MLV Reverse
Transcription kit (Promega Corporation). The tube was heated at
65°C for 10 min. Transcriptor reverse transcriptase reaction buffer
(4 µl), RNA inhibitor (0.5 µl), deoxynucleotide mix (10 mM; 2 µl),
and Transcriptor reverse transcriptase (0.5 µl) were added. The
reagents were mixed in the tube carefully. Tubes were put in the
PCR machine at 25°C for 10 min followed by 60 min at 50°C, then
85°C for 5 min. qPCR was performed using IQ SYBR Green Supermix
reagent (Bio-Rad Laboratories, Inc.) with a Bio-Rad real-time PCR
machine, according to the manufacturer's instructions. Denaturation
was performed at 95°C for 20 sec, followed by 38 cycles of 95°C for
5 sec, 50°C for 45 sec and 72°C for 30 sec. The data were analyzed
using the 2−ΔΔCq method (22). The expression levels of the target
genes were normalized to the GAPDH level in each sample. The primer
sequences were as follows: MMP-2, forward
5′-GCAGCCCATGAGTTCGGCCAT-3′ and reverse
5′-AGCATCAGGGGAGGGCCCATA-3′; MMP-9, forward
5′-GGAGACGGCAAACCCTGCGT-3′ and reverse
5′-TGACGTCGGCTCGAGTAGGACA-3′; and GAPDH, forward
5′-AGGTCGGTGTGAACGGATTTG-3′ and reverse
5′-TGTAGACCATGTAGTTGAGGTCA-3′.
Statistical analysis
Data analysis was performed using SPSS for Windows
(v.16.0; SPSS Inc.). Data are expressed as mean ± standard error of
the mean. Differences between the means of the groups were assessed
by one-way ANOVA followed by least significant difference tests (in
cases of equal variance) or Dunnett's test (when there was
heterogeneity in the variance) for post hoc comparisons. P<0.05
was considered to indicate a statistically significant
difference.
Results
DEX promotes hypoxia-induced
proliferation
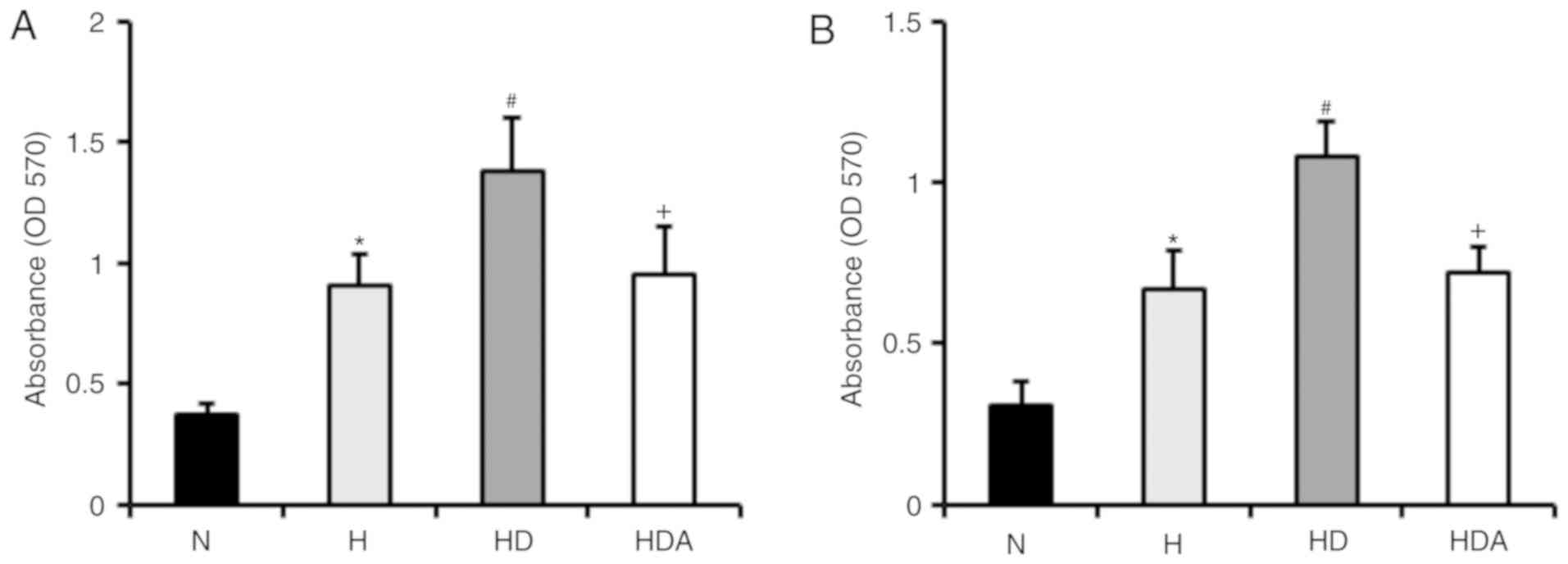
To examine whether DEX promotes hypoxia-induced
proliferation of A549 cells and HCT116 cells, cells were treated
with DEX and hypoxia for 4 h (Fig.
1). The results revealed that hypoxia-treated cells developed a
significant elevation of proliferative rate compared with cells
treated with normoxia (P<0.05). The proliferation rate of
DEX-treated cells following hypoxia insult was markedly higher than
that of cells treated with hypoxia alone (P<0.05). These data
indicated that DEX promoted the hypoxia-induced proliferation of
the two cancer cell lines. Furthermore, atipamezole almost fully
abolished the effects of DEX on the proliferation of the cancer
lines following hypoxia-induced proliferation.
DEX increases the protein levels of
survivin in cells following hypoxia treatment
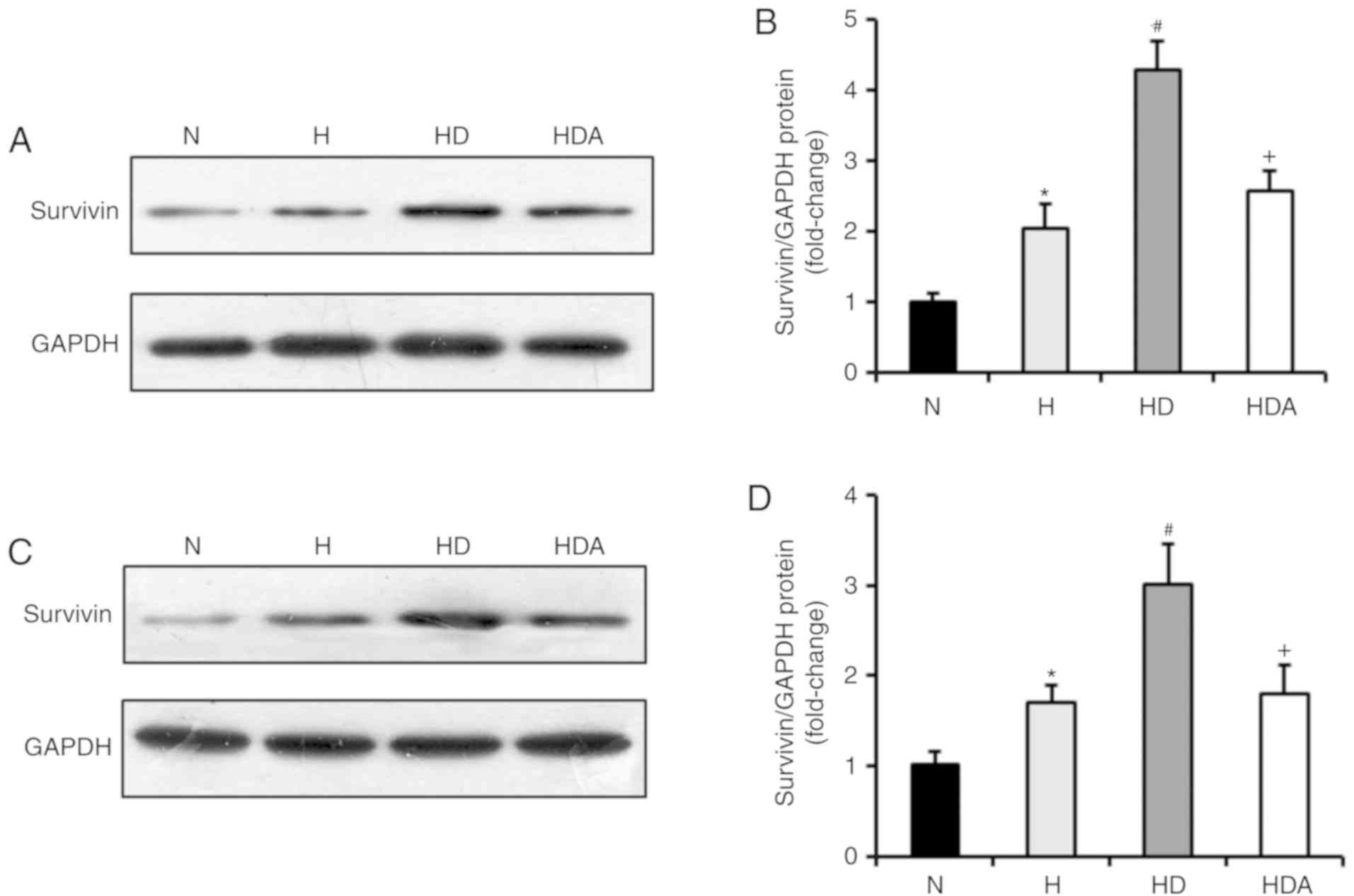
Whether DEX was able to increase the expression of
survivin was subsequently investigated. Through western blotting,
it was demonstrated that survivin was markedly induced in A549
cells and HCT116 cells in response to hypoxia treatment, compared
with normoxia treatment (P<0.05; Fig.
2). Moreover, DEX further upregulated the protein levels of
survivin in the two cancer cell lines after hypoxia (P<0.05
compared with the H group). The effects of DEX on the
hypoxia-induced expression of survivin were almost fully abrogated
by atipamezole treatment (P<0.05). These findings suggested that
DEX may increase the hypoxia-induced proliferation of the two
cancer cell lines by regulating survivin expression, which may be
associated with an α2 adrenoceptor mediated mechanism of
action.
DEX enhances hypoxia-induced invasion
ability
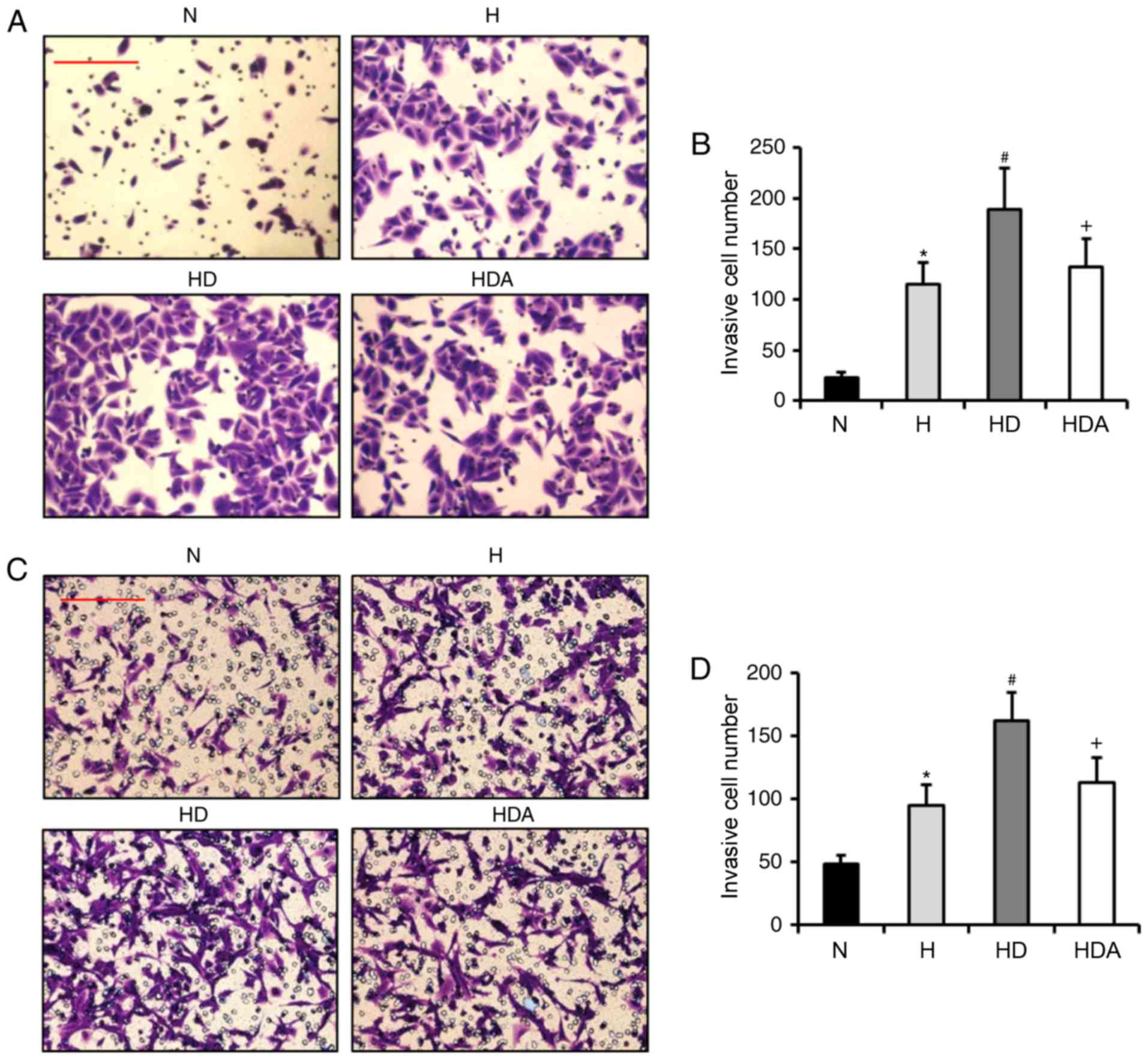
It has been reported that DEX contributes to the
invasion of breast cancer cells under normal condition (14,15).
Based on this information, it was hypothesized that DEX may also
enhance the hypoxia-induced invasion of A549 cells and HCT116
cells. Through Matrigel assays, it was revealed that the invasive
number of cells from the two cancer cell lines was increased
significantly after hypoxia treatment (P<0.05; Fig. 3). The invasive cell number in the HD
group was elevated substantially as compared with the H group for
both cancer cell lines (P<0.05). These data indicated that DEX
exacerbated the hypoxia-induced invasive potential of the two
cancer cell lines. The invasive cell number in the HDA group was
reduced significantly as compared with the HD group (P<0.05).
These results suggested that atipamezole reversed the effects of
DEX on the hypoxia-induced invasion of the two cancer cell
lines.
DEX aggravates hypoxia-induced
migration ability
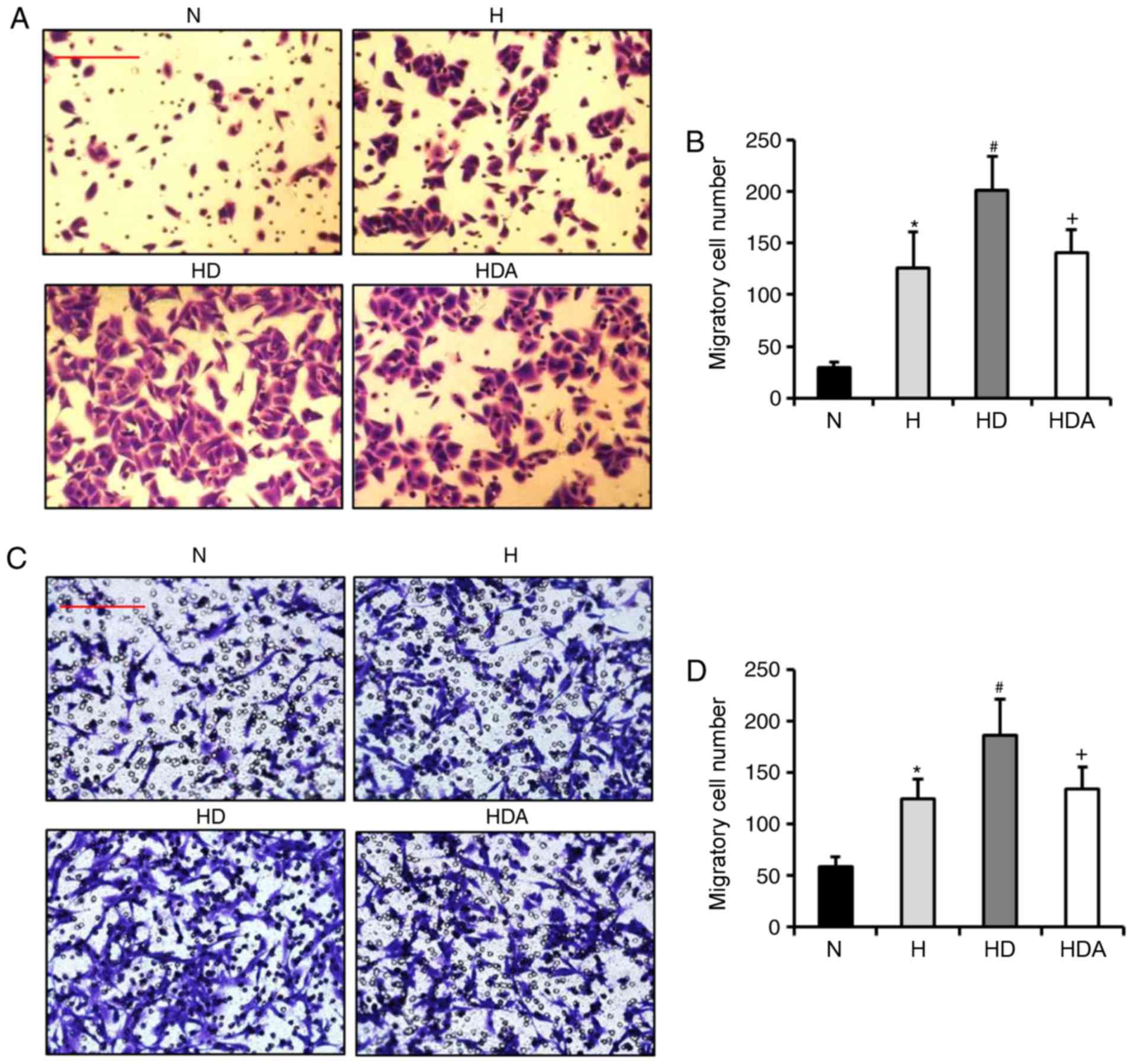
Subsequently, the present study assessed whether DEX
increases the hypoxia-induced migration of A549 cells and HCT116
cells. Using Transwell assays, the findings showed that the number
of migrated cells from the two cell cancer lines was elevated
significantly after hypoxia treatment (Fig. 4). In comparison, treatment with DEX
further significantly increased the number of migratory cells
(P<0.05). These results suggested that DEX aggravated
hypoxia-induced migratory ability. The migratory cell number in the
HDA group decreased substantially as compared with the HD group
(P<0.05). These data indicated that atipamezole abolished the
effect of DEX on the hypoxia-induced migration of the two cancer
cell lines.
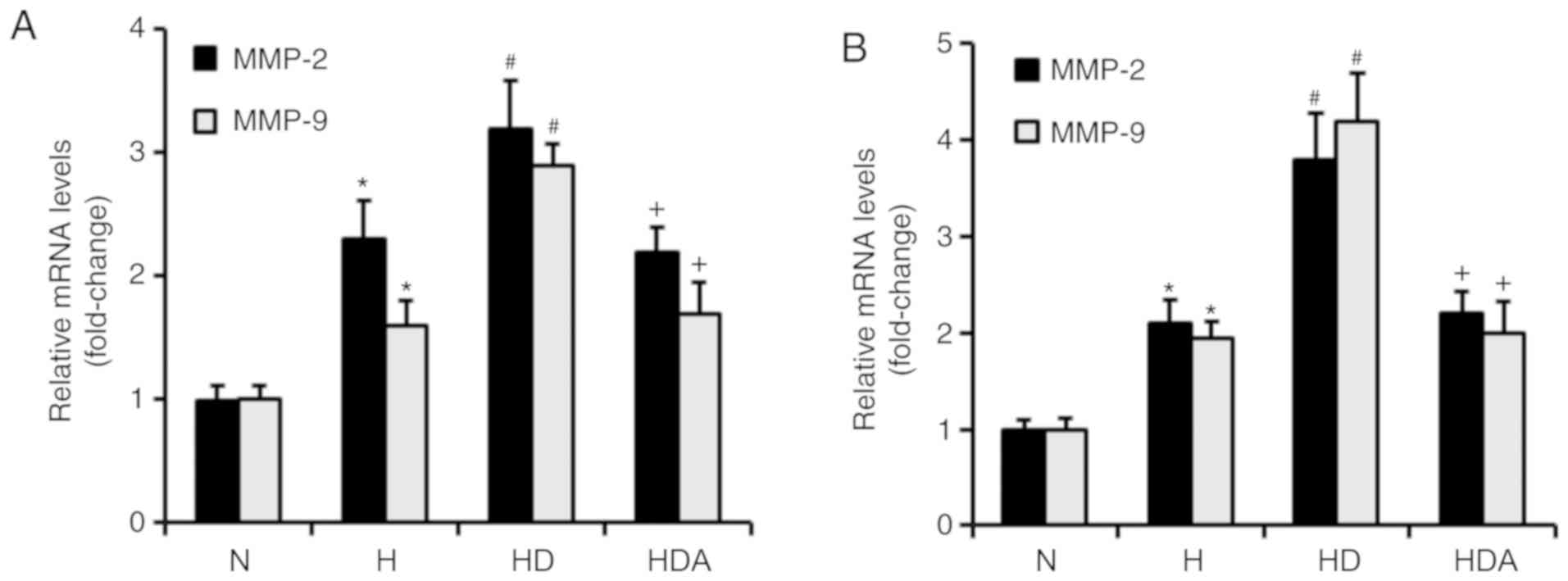
DEX upregulates the mRNA levels of
MMP-2 and MMP-9 in cancer cells in response to hypoxia
MMP-2 and MMP-9 both have critical roles in the
metastasis of cancer cells (23).
RT-qPCR was used to investigate whether DEX increased the mRNA
levels of MMP-2 and MMP-9 (Fig. 5).
It was found that hypoxia treatment significantly upregulated the
mRNA levels of both MMP-2 and MMP-9 in the two cancer cell lines
(P<0.05). The mRNA levels of MMP-2 and MMP-9 were increased
significantly in the HD group compared with the H group
(P<0.05). Compared with the HD group, the mRNA levels of MMP-2
and MMP-9 in group HDA were significantly elevated (P<0.05).
Furthermore, atipamezole reversed the effects of DEX on the
hypoxia-induced mRNA expression of MMP-2 and MMP-9 in the two
cancer cell lines (Fig. 5). These
results indicated that DEX may enhance the hypoxia-induced
metastatic potential of the two cancer cell lines by regulating
MMP-2 and MMP-9 expression, which may be dependent on the
α2 adrenoceptor.
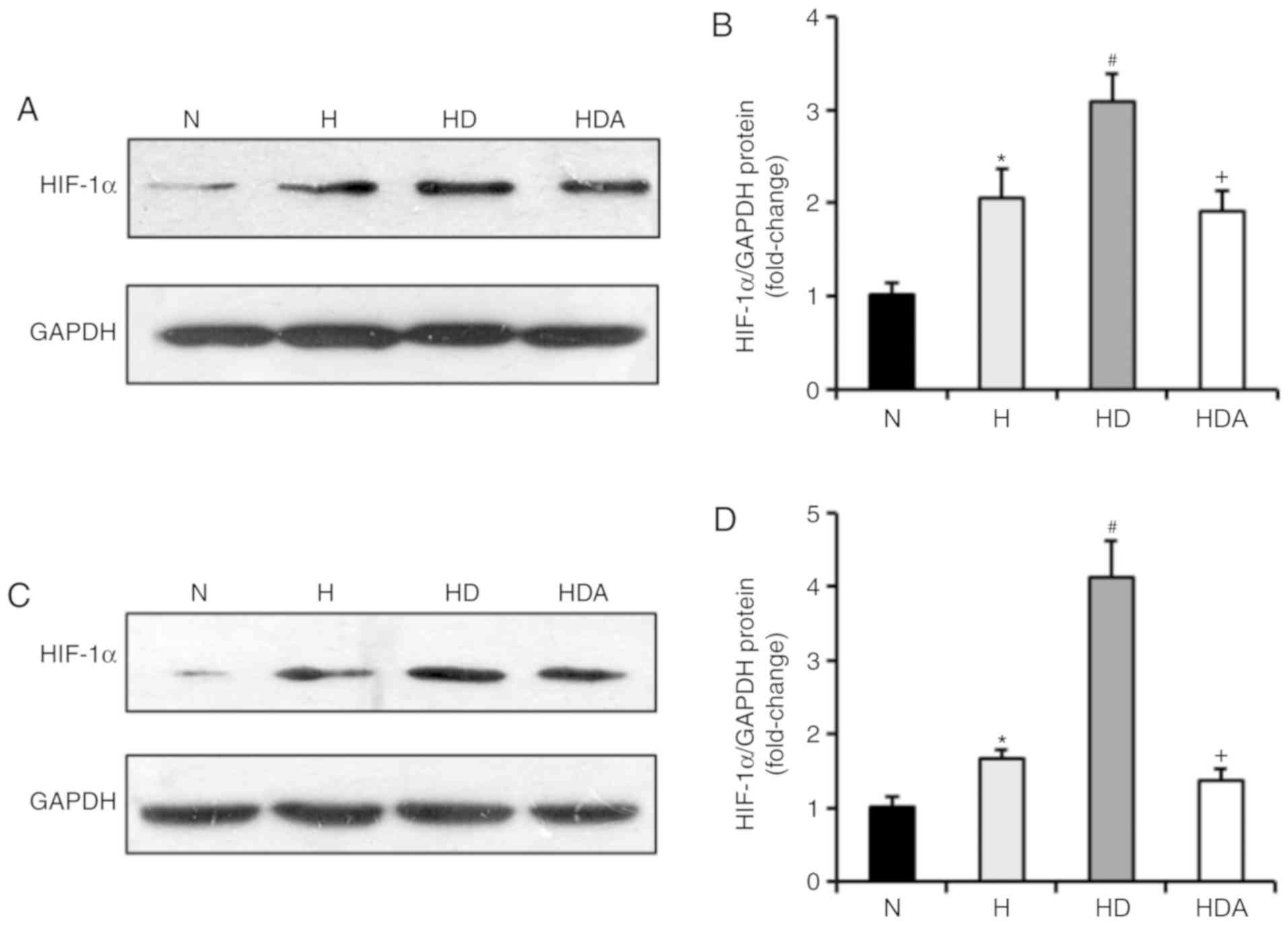
DEX drives upregulation of HIF-1α
protein in cells following hypoxic stress
It has been documented that DEX has a protective
influence on organs by upregulating HIF-1α (24). Therefore, the present study examined
whether DEX affected the protein expression of HIF-1α in the two
cancer cell lines (Fig. 6). The
results of the western blotting showed that the HIF-1α protein
levels of the two cells were elevated markedly in response to
hypoxia (P<0.05). Cells treated with DEX and hypoxia displayed a
significant elevation of HIF-1α protein compared with cells exposed
to hypoxia alone (P<0.05). These data suggested that the effects
of DEX on hypoxia-induced growth and metastasis of the cancer cells
may be related to upregulation of HIF-1α signaling. Furthermore,
treatment with atipamezole abolished the effects of DEX on
hypoxia-induced expression of HIF-1α (P<0.05), suggesting the
effect of DEX on HIF-1α expression may be mediated by α2
adrenoceptor signaling.
DEX enhances the progression of cancer
cells under normoxic conditions
The effects of DEX on the biological functions of
A549 cells and HCT116 cells under normoxic condition were also
examined. The results demonstrated that DEX may enhance
proliferation and metastasis of these two cancer cells treated with
normoxia (data not shown). These results suggested that DEX
promotes cancer cell progression under normoxic condition.
Discussion
The perioperative period is a critical time of
metastatic vulnerability as dissemination of tumor cells often
occurs during surgery (25).
Evidence shows that anesthetic agents have an important impact on
the biological behaviors of tumor cells and may influence the
recurrence or the progression of cancer (26,27).
Thus, the choice of anesthetic drugs during the perioperative
period of cancer surgery should be considered seriously. Hypoxia is
one of the most important features of solid tumors, and has been
shown to contribute to the proliferation and metastasis of cancer
cells (28). Unfortunately, hypoxia
is a common complication during the perioperative period. DEX is
increasingly used as an adjunct to anesthesia. Therefore, a better
understanding of the effects of DEX on the hypoxia-induced
progression of cancer cells is urgently needed.
In the present study, the following points were
demonstrated: i) DEX may enhance the hypoxia-induced growth and
metastatic potential of A549 cells and HCT116 cells; ii) DEX
increased the protein expression of survivin for both of the two
cancer cell lines in response to hypoxic stress; iii) DEX further
promoted the hypoxia-induced invasive and migratory ability of the
two cancer cell lines; iv) DEX promoted the upregulation of MMP-2,
MMP-9 and HIF-1α mRNA expression of the two cancer cell lines after
hypoxia; and v) the effects of DEX on hypoxia-induced progression
of cancer cells may be brought about in an α2
adrenoreceptor-dependent manner. These data suggested that DEX
enhanced the hypoxia-induced progression of lung cancer cells and
colorectal cancer cells via the regulation of HIF-1α signaling,
which may be functioning in an α2 adrenoceptor-dependent
pathway.
Survivin is a member of the inhibitor of apoptosis
protein family, which is expressed in a wide variety of human
tumors. Survivin orchestrates several important mechanisms to
promote the proliferation of cancer cells, including anti-apoptotic
activity and regulation of the cell cycle (29). Overexpression of survivin in cancer
cells is associated with massive proliferation of cancer cells,
resulting in a poor prognosis (30).
In this present study, it was demonstrated that the expression of
survivin in the two cancer cell lines was substantially elevated
after hypoxic treatment. Furthermore, treatment with DEX increased
the protein levels of survivin in A549 cells and HCT cells in
response to hypoxic stress. These results indicated that DEX may
contribute to hypoxia-induced proliferation of cancer cells by
regulating the expression of survivin.
MMP-2 and MMP-9 are both zinc-dependent proteolytic
enzymes that are capable of cleaving extracellular matrix and
basement membrane components. The two enzymes directly facilitate
tumor invasion and migration, and play a prominent role in
metastasis (23). Overexpression of
MMP-2 and MMP-9 is consistently related to cancer metastasis and
poorer prognosis (23). In the
present study, it was observed that the mRNA expression of both
MMP-2 and MMP-9 was increased significantly in the two cancer cell
lines following hypoxia. Moreover, DEX treatment promoted the
hypoxia-induced raised expression of MMP-2 and MMP-9. These data
suggested that DEX may promote the invasion and migration of cancer
cells induced by hypoxia, by upregulating MMP-2 and MMP-9.
HIF-1 is composed of HIF-1α and HIF-1β. As an oxygen
sensor in hypoxic microenvironments, HIF-1α regulates cancer cell
metabolism, proliferation, apoptosis and metastasis (31). Under hypoxic conditions, HIF-1α binds
to HIF-1β to form the HIF-1 transcription complex that activates
the transcription of many target genes, including survivin and MMPs
(32–35). In the present study, the findings
showed that hypoxia treatment upregulated the HIF-1α levels of the
two cancer cell lines. Recently, a study reported that DEX protects
against lung ischemia-reperfusion injury via upregulation of HIF-1α
(24). The findings of this present
study also showed that DEX may drive the upregulation of HIF-1α
expression in cancer cells following hypoxic insult. These data
indicated that DEX may enhance hypoxia-induced cancer cell
progression by modulating the HIF-1α signaling pathway.
It is reported that the organ-protective effects of
DEX are regulated by α2 adrenoceptors (36,37).
This present study investigated whether α2 adrenoceptors
play a role in the effect of DEX on the progression of the two
cancer cell lines. It was demonstrated that the effect of DEX on
the hypoxia-induced proliferation and metastatic ability of cancer
cells may be abolished by atipamezole. In addition, the effects of
DEX on the hypoxia-induced expression of survivin, MMP-2, MMP-9 and
HIF-1α were removed by atipamezole. The present data suggested that
the effect of DEX on hypoxia-induced cancer cell progression may
occur in an α2 adrenoceptor mediated manner.
This present study also investigated the effects of
DEX on the biological function of A549 cells and HCT cells under
normoxic conditions. It was revealed that DEX may enhance
proliferation and the metastasis of these two cancer cell lines
under normoxic conditions (data not shown), suggesting that DEX
promotes cancer cell progression under normoxic conditions.
In conclusion, dexmedetomidine enhances the
hypoxia-induced progression of lung cancer cells and colorectal
cancer cells by regulating HIF-1α signaling, which may function
though the α2 adrenoceptor pathway.
Acknowledgements
Not applicable.
Funding
This study was supported by Guangdong Province
Science and Technology Planning Project, Guangdong, China (grant
no. 2014A020212467).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
HYC performed the experiments and revised this
manuscript. JYZ performed the western blotting and data analysis.
GHL performed the Transwell assays and data analysis. GCT and XHL
were responsible for proliferation evaluation, western blotting and
RT-qPCR. HL and QH conceived and designed this investigation. All
authors read and approved the final version of the manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Hopewood P and Milroy MJ and Milroy MJ:
Cancer Statistics: Global and NationalQuality Cancer Care.
Springer; New York, NY, USA: pp. 29–35. 2018
|
|
2
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Hiller JG, Perry NJ, Poulogiannis G,
Riedel B and Sloan EK: Perioperative events influence cancer
recurrence risk after surgery. Nat Rev Clin Oncol. 15:205–218.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Olcina MM, Kim RK, Melemenidis S, Graves
EE and Giaccia AJ: The tumour microenvironment links complement
system dysregulation and hypoxic signalling. Br J Radiol.
15:201800692018.(Epub ahead of print). View Article : Google Scholar
|
|
5
|
Wigerup C, Påhlman S and Bexell D:
Therapeutic targeting of hypoxia and hypoxia-inducible factors in
cancer. Pharmacol Ther. 164:152–169. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Gupta K, Prasad A, Nagappa M, Wong J,
Abrahamyan L and Chung FF: Risk factors for opioid-induced
respiratory depression and failure to rescue: A review. Curr Opin
Anaesthesiol. 31:110–119. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Campos JH and Feider A: Hypoxia during
one-lung ventilation-a review and update. J Cardiothorac Vasc
Anesth. 32:2330–2338. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Soltanizadeh S, Degett TH and Gogenur I:
Outcomes of cancer surgery after inhalational and intravenous
anesthesia: A systematic review. J Clin Anesth. 42:19–25. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Cassinello F, Prieto I, del Olmo M, Rivas
S and Strichartz GR: Cancer surgery: How may anesthesia influence
outcome? J Clin Anesth. 27:262–272. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wu GJ, Chen JT, Tsai HC, Chen TL, Liu SH
and Chen RM: Protection of dexmedetomidine against
ischemia/reperfusion-induced apoptotic insults to neuronal cells
occurs via an intrinsic mitochondria-dependent pathway. J Cell
Biochem. 118:2635–2644. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wang SL, Duan L, Xia B, Liu Z, Wang Y and
Wang GM: Dexmedetomidine preconditioning plays a neuroprotective
role and suppresses TLR4/NF-kB pathways model of cerebral ischemia
reperfusion. Biomed Pharmacother. 93:1337–1342. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Cui J, Zhao H, Wang C, Sun JJ, Lu K and Ma
D: Dexmedetomidine attenuates oxidative stress induced lung
alveolar epithelial cell apoptosis in vitro. Oxid Med Cell Longev.
2015:3583962015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Cho JS, Shim JK, Soh S, Kim MK and Kwak
YL: Perioperative dexmedetomidine reduces the incidence and
severity of acute kidney injury following valvular heart surgery.
Kidney Int. 89:693–700. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Szpunar MJ, Burke KA, Dawes RP, Brown EB
and Madden KS: The antidepressant desipramine and alpha2-adrenergic
receptor activation promote breast tumor progression in association
with altered collagen structure. Cancer Prev Res (Phila).
6:1262–1272. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Bruzzone A, Pinero CP, Castillo LF,
Sarappa MG, Rojas P, Lanari C and Luthy IA: Alpha2-adrenoceptor
action on cell proliferation and mammary tumour growth in mice. Br
J Pharmacol. 155:494–504. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Castillo LF, Rivero EM, Goffin V and Luthy
IA: Alpha2-adrenoceptor agonists trigger prolactin signaling in
breast cancer cells. Cell Signal. 34:76–85. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Deng F, Ouyang M, Wang X, Yao X, Chen Y,
Tao T, Sun X, Xu L, Tang J and Zhao L: Differential role of
intravenous anesthetics in colorectal cancer progression:
Implications for clinical application. Oncotarget. 7:77087–77095.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Virtanen R: Pharmacological profiles of
medetomidine and its antagonists, atipamezole. Acta Vet Scand
Suppl. 85:29–37. 1989.PubMed/NCBI
|
|
19
|
Liang H, Yang CX, Zhang B, Wang HB, Liu
HZ, Lai XH, Liao MJ and Zhang T: Sevoflurane suppresses
hypoxia-induced growth and metastasis of lung cancer cells via
inhibiting hypoxia-inducible factor-1α. J Anesth. 29:821–830. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Peng JK, Shen SQ, Wang J, Jiang HW and
Wang YQ: Etaypoxia-inducible factor 1-α promotes colon cell
proliferation and migration by upregulating AMPK-related protein
kinase 5 under hypoxic conditions. Oncol Lett. 15:3639–3645.
2018.PubMed/NCBI
|
|
21
|
Li H, Wang X, Wen C, Huo Z, Wang W, Zhan
Q, Cheng D, Chen H, Deng X, Peng C and Shen B: Long noncoding RNA
NORAD, a novel competing endogenous RNA, enhances the
hypoxia-induced epithelial-mesenchymal transition to promote
metastasis in pancreatic cancer. Mol Cancer. 16:1692017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Alaseem A, Alhazzani K, Dondapati P,
Alobid S, Bishayee A and Rathinavelu A: Matrix metalloproteinases:
A challenging paradigm of cancer management. Semin Cancer Biol.
56:100–115. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhang W, Zhang JQ, Meng FM and Xue FS:
Dexmedetomidine protects against lung ischemia-reperfusion injury
by the PI3K/Akt/HIF-1α signaling pathway. J Anesth. 30:826–833.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Piegeler T and Beck-Schimmer B: Anesthesia
and colorectal cancer-The perioperative period as a window of
opportunity? Eur J Surg Oncol. 42:1286–1295. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wigmore TJ, Mohammed K and Jhanji S:
Long-term survival for patients undergoing volatile versus IV
anesthesia for cancer surgery: A retrospective analysis.
Anesthesiology. 124:69–79. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Xie Z: Cancer prognosis: Can anesthesia
play a role? Anesthesiology. 119:501–503. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Terry S, Buart S and Chouaib S: Hypoxic
stress-induced tumor and immune plasticity, suppression, and impact
on tumor heterogeneity. Front Immunol. 8:16252017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Athanasoula KCh, Gogas H, Polonifi K,
Vaiopoulos AG, Polyzos A and Mantzourani M: Survivin beyond
physiology: Orchestration of multistep carcinogenesis and
therapeutic potentials. Cancer Lett. 347:175–182. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Mobahat M, Narendran A and Riabowol K:
Survivin as a preferential target for cancer therapy. Int J Mol
Sci. 15:2494–2516. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Rankin EB and Giaccia AJ: Hypoxic control
of metastasis. Science. 352:175–180. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Li X, Liu X, Xu Y, Liu J, Xie M, Ni W and
Chen S: KLF5 promotes hypoxia-induced survival and inhibits
apoptosis in non-small cell lung cancer cells via HIF-1α. Int J
Oncol. 45:1507–1514. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Sun XP, Dong X, Lin L, Jiang X, Wei Z,
Zhai B, Sun B, Zhang Q, Wang X, Jiang H, et al: Up-regulation of
survivin by AKT and hypoxia-inducible factor 1α contributes to
cisplatin resistance in gastric cancer. FEBS J. 281:115–128. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Ai P, Shen B, Pan H, Chen K, Zheng J and
Liu F: MiR-411 suppressed vein wall fibrosis by downregulating
MMP-2 via targeting HIF-1α. J Thromb Thrombolysis. 45:264–273.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Yang L, Shen L, Li G, Yuan H, Jin X and Wu
X: Silencing of hypoxia inducible factor-1α gene attenuated
angiotensin-induced abdominal aortic aneurysm in apolipoprotein
E-deficient mice. Atherosclerosis. 252:40–49. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Gu J, Sun P, Zhao H, Watts HR, Sanders RD,
Terrando N, Xia P, Maze M and Ma D: Dexmedetomidine provides
renoprotection against ischemia-reperfusion injury in mice. Crit
Care. 15:R1532011. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zhang XY, Liu ZM, Wen SH, Li YS, Li Y, Yao
X, Huang WQ and Liu KX: Dexmedetomidine administration before, but
not after, ischemia attenuates intestinal injury induced by
intestinal ischemia-reperfusion in rats. Anesthesiology.
116:1035–1046. 2012. View Article : Google Scholar : PubMed/NCBI
|