Introduction
Cervical cancer has become the fourth most common
cause of cancer-related death and the second most common malignancy
in women worldwide (1). Despite
improved treatment, the number of women succumbing to cervical
cancer is increasing. The prognosis of patients with
advanced/recurrent cervical cancer is poor and 5-year of survival
is only 10–20% (2). Tumor metastasis
and recurrence are the main cause of the poor outcome and high
mortality. The current standardized treatment for
advanced/recurrent cervical cancer is cisplatin-based chemotherapy
(3,4). However, numerous patients do not
benefit from chemotherapy and tumors often develop chemoresistance
(5). Therefore, there is an urgent
requirement to uncover the mechanisms of progression and
chemoresistance in cervical carcinoma. Notably, recent studies have
demonstrated that microRNAs (miRNAs/miRs) may be involved in drug
resistance (6–8).
miRNAs are small single stranded non-coding RNA
molecules of 17–24 nucleotides, which regulate targeted genes by
inducing post-transcriptional degradation or suppressing
translation (9). Aberrant miRNA
expression has been found to play a vital role in the progression
of cervical cancer. It has been reported that miR-150 is
overexpressed in patient tissues and promotes cervical cancer cell
viability, migration and invasion (10). miR-506 suppresses tumor growth and
enhances apoptosis and chemosensitivity in cervical cancer
(11). miR-138 has been reported to
function as a tumor suppressor, since it plays critical role in
certain pathological processes, including tumor progression,
metastasis, invasion and apoptosis (12–14).
However, the function and chemoresistance mechanism of miR-138 in
cervical cancer are poorly understood.
In the current study, the expression and function of
miR-138 in cervical cancer were investigated in vitro and
in vivo. Furthermore, the target of miR-138 was identified.
The results revealed that miR-138 was downregulated in cervical
cancer cells. In vitro and in vivo, miR-138
suppressed progression and enhanced chemosensitivity.
Dual-luciferase reporter assays and rescue experiments revealed
that histone H2AX phosphorylation (H2AX) was the functional target
gene of miR-138. The results of the present study indicate that
miR-138 may be a novel biomarker of progression and chemoresistance
in cervical cancer.
Materials and methods
Cell lines
Cervical cancer cells SiHa and C33A and immortalized
normal cervical epithelial squamous cell line H8 were obtained from
the Cell Bank of the Chinese Academy of Sciences. SiHa and H8 cells
were cultured in Dulbecco's modified Eagle's medium (Thermo Fisher
Scientific, Inc.) and C33A cells were cultured in minimum Eagle's
medium (Thermo Fisher Scientific, Inc.) each supplemented with 10%
fetal bovine serum (Thermo Fisher Scientific, Inc.), in a 37°C
incubator with 5% CO2.
Transfection
In total, 50 nM miR-138 mimic
(5′-AGCUGGUGUUGUAAUCAGGCCGGCCUGAUUCACAACACCAGCUUU-3′), 100 nM
miR-138 inhibitor (5′-CGGCCUGAUUCACAACACCAGCU-3′) or 50 nM
miR-negative control (NC; sense, 5′-UUCUCCGAACGUGUCACGUTT-3′ and
antisense, 5′-ACGUGACACGUUCGGAGAATT-3′) were transfected into the
cells using Lipofectamine® 3000 (Thermo Fisher
Scientific, Inc.) according to the manufacturer's protocol.
Following 48 h transfection, the cells were subjected to further
experimentation. The miR-138 mimic, miR-138 inhibitor and miR-NC
were designed by Shanghai GenePharma Co., Ltd. The H2AX expression
vector, carrying the human H2AX-coding DNA sequence, was cloned
into a pcDNA3.1 vector (Thermo Fisher Scientific, Inc.).
RNA detection
miRNA and total RNA extraction were extracted from
cells using miRcute miRNA Isolation Kit (Tiangen Biotech Co., Ltd.)
or TRIzol® reagent (Thermo Fisher Scientific, Inc.)
according to the manufacturers' protocols. cDNA was synthesized
using the miRcute Plus miRNA First-Strand cDNA Kit (Tiangen Biotech
Co., Ltd.) according to the manufacturer's protocol. The miRNA cDNA
reverse transcription conditions were as follows: 42°C for 60 min
and 95°C for 3 min. Total RNA was reverse transcribed into cDNA
with the following conditions: 42°C for 60 min and 70°C for 5 min.
miRNA and mRNA expression levels were determined using miRcute Plus
miRNA qPCR Kit (SYBR® Green; Tiangen Biotech Co., Ltd.)
and SYBR® Green Real-Time PCR System (Applied
Biosystems; Thermo Fisher Scientific, Inc.), respectively. The
thermocycling conditions for miRNA were: Initial denaturation at
95°C for 15 min, followed by 45 cycles of denaturation at 94°C for
20 sec and anneal/extend at 60°C for 34 sec. The thermocycling
conditions for mRNA were: Initial denaturation at 95°C for 2 min,
followed by 40 cycles of denaturation at 95°C for 15 sec and
anneal/extend at 60°C for 1 min. The internal controls were U6 and
β-actin. Relative expression was calculated and normalized using
the 2−ΔΔCq method (15).
The primer sequences are listed in Table
I.
 | Table I.Reverse transcription-PCR Primer
Sequences. |
Table I.
Reverse transcription-PCR Primer
Sequences.
| Primer | Sequence |
|---|
| β-actin |
5′-TGTCCACCTTCCAGCAGATGT-3′ (Forward) |
|
|
5′-GCTCAGTAACAGTCCGCCTAGA-3′
(Reverse) |
| H2AX |
5′-CGGCAGTGCTGGAGTACCTCA-3′ (Forward) |
|
|
5′-AGCTCCTCGTCGTTGCGGATG-3′ (Reverse) |
| GAPDH |
5′-AGAAGGCTGGGGCTCATTTG −3′ (Forward) |
|
|
5′-AGGGGCCATCCACAGTCTTC −3′ (Reverse) |
| miR-138 |
5′-AGCUGGUGUUGUGAAUCAGGCCG-3′
(Forward) |
|
|
5′-TGGTGTCGTGGAGTCG-3′ (Reverse) |
Cell proliferation and
chemosensitivity assay
To analyze cell proliferation, a Cell Counting Kit-8
assay (CCK-8; Dojindo Molecular Technologies, Inc.) was used
according to the manufacturer's protocol. Cells were plated in
96-well plates (2,000 cells/well) following a 24 h transfection.
After overnight incubation, the cells were incubated with CCK-8
solution at 37°C for 2 h. Next, viable cell numbers were measured
by absorbance at 450 nm using a spectrophotometer. Cells were
plated in 96-well plates (4,000 cells/well) and treated with
different doses of cisplatin (Sigma-Aldrich; Merck KGaA) following
transfection for 24 h. Subsequently, the toxicity of DDP in the
cells was determined using a CCK-8 assay.
EdU staining
EdU assays (Guangzhou RiboBio Co., Ltd.) were
performed according to the manufacturer's protocol. Transfected
cells were incubated with 50 µM of EdU labeling medium for 2 h at
37°C. Then, 4% paraformaldehyde (50 µl/well) was used to fix cells
for 20 min at 37°C, prior to incubation with glycine (2 mg/ml).
After ~5 min, 200 µl 1X Apollo solution was used to stain cells for
30 min at 37°C in the dark. Triton X-100 (0.5%) in PBS was used to
wash cells for 5 min, then three random fields of view per slide
were examined using a fluorescence microscope (Olympus
Corporation). The number of proliferative cells (EdU positive) was
counted.
Apoptosis assay
SiHa and C33A cells (1×106) were seeded
in 6-well plates and treated with cisplatin (10 µM/ml) for 24 h
prior to harvesting. An Annexin V-FITC Apoptosis Detection kit (BD
Biosciences) was used to perform flow cytometry analysis. Cell
apoptosis was measured using BD FACSCanto™ II flow cytometer (BD
Biosciences) and analyzed using the FlowJo software (version
10.0.7; FlowJo LLC).
Cell migration assay
A 24-well Transwell chamber assay (pore size, 8 µm)
was used to perform migration experiments (BD Biosciences).
Transfected cervical cancer cells (2×105 cells/well)
were placed into the upper chambers. After 24 h of incubation, the
cells that had migrated through the membrane were fixed in 4%
methanol for 30 min and stained with 0.05% crystal violet for 30
min, both at room temperature. Then, the invaded cells were
quantified under a microscope (Olympus Corporation).
Wound-healing assay
Cells (1×106 cells/well) were incubated
in a 24-well plate; the cell monolayer was then wounded with a 100
µl pipette tip and cultured in serum-free medium. Cell migration
images were captured at 0 and 24 h.
Prediction of miR-138 targets using
TargetScan
Potential targets of miR-138 were searched using the
TargetScan software (release 7.2; http://www.targetscan.org/vert_72/), and the resulting
search resulted in H2AX being a candidate.
Luciferase reporter assay
The wild-type 3′-untranslated region (UTR) sequences
of H2AX cDNA (WT-H2AX) were synthesized into the luciferase
reporter vector [Obio Technology (Shanghai) Corps., Ltd.]. A mutant
H2AX 3′-UTR vector (MUT-H2AX) was also constructed, which contained
a mutation in the predicted H2AX-binding sequence. The WT-H2AX or
MUT-H2AX and the miR-138 mimic or miR-138 NC, were transfected into
cervical cells using Lipofectamine® 3000 reagent (Thermo
Fisher Scientific, Inc.). After 48 h transfection, a
Luciferase-Reporter Assay System kit (Promega Corporation) was used
to measure luciferase activity using Renilla luciferase
activity as internal reference.
Western blotting analysis
Total protein was obtained following cell lysis,
using RIPA buffer (Beyotime Institute of Biotechnology)
supplemented with 1 nM phenylmethylsulfonyl fluoride. Total protein
was collected and the concentration was determined with an Enhanced
Bicinchoninic Acid Protein Assay kit (Beyotime Institute of
Biotechnology). Proteins (30 µg) were subjected to denaturing 10%
SDS-polyacrylamide gel electrophoresis and transferred onto a PVDF
membrane (EMD Millipore). The following antibodies were used
overnight at 4°C: Caspase-3 (1:1,000; Abcam; cat. no. ab13585),
bcl-2 (1:1,000; Abcam; cat. no. ab117115), bax (1:1,000; Abcam;
cat. no. ab32503), matrix metalloproteinase (MMP) 9 (1:1,000;
Abcam; cat. no. ab38898), H2AX (1:1,000; Abcam; cat. no. ab81299)
and β-actin (1:1,000; Proteintech Group, Inc.; cat. no.
66009-1-Ig). The membrane was incubated with horseradish
peroxidase-conjugated anti-rabbit (1:4,000; cat. no. A0208;
Beyotime Institute of Biotechnology) or anti-mouse (1:4,000; cat.
no. A0216; Beyotime Institute of Biotechnology) for 2 h at room
temperature. An enhanced chemiluminescence system (Thermo Fisher
Scientific, Inc.) was used was used to produce signals and Quantity
One software (version 3.0; Bio-Rad Laboratories, Inc.) was used for
signal detection.
Animal experiment
A total of 20 (4-week old, 15–20 g) female BALB/c
nude mice were purchased from the Shanghai Laboratory Animal
Center. The animals were housed at 21–24°C with 50–60% relative
humidity and a 12-h light/dark cycle. They had free access to food
and water. All experiments were approved by the Animal Management
Rule of the Chinese Ministry of Health (document 55, 2001) and
approved by the Tongji University Animal Ethics Committee.
Lentivirus miR-138 vector or empty vector (Obio Technology) was
transfected into SiHa cells. A total of 5×106 cells
stably transfected with empty vector or miR-138 vector were
injected subcutaneously into the right flank region of each mouse.
Tumor length and width were measured every 2 days using Vernier
calipers, and tumor volume was calculated as: Volume = 0.5 × length
× width2. When the tumor became palpable, the mice
bearing either miR-138-expressing tumors or empty vector tumors
were randomized into 4 groups (5 mice/group) and were
intraperitoneally injected with PBS or cisplatin (10 mg/Kg). The
mice were humanely sacrificed at 23 days after implantation and the
xenograft tumors were collected for weighing and subsequent
analysis.
Immunohistochemical (IHC)
analysis
For histopathology, paraffin-embedded tissues
obtained from xenograft mice were sectioned to a thickness of 4 µm.
The expression of caspase-3, cleaved-caspase3, MMP9, H2AX and MDR1
was evaluated, as previously described (16).
Statistical analysis
All experiments were performed at least three times
with duplicate or triplicate samples in each assay. Statistical
analysis was performed using SPSS 17.0 software (SPSS, Inc.) and
GraphPad Prism software 7.0 (GraphPad Software, Inc.). The
Student's t-test and one-way analysis of variance followed by
Tukey's or Student's t-test were performed to calculate the
differences between groups. P<0.05 was considered to indicate a
statistically significant difference. Data are presented as the
mean ± standard deviation.
Results
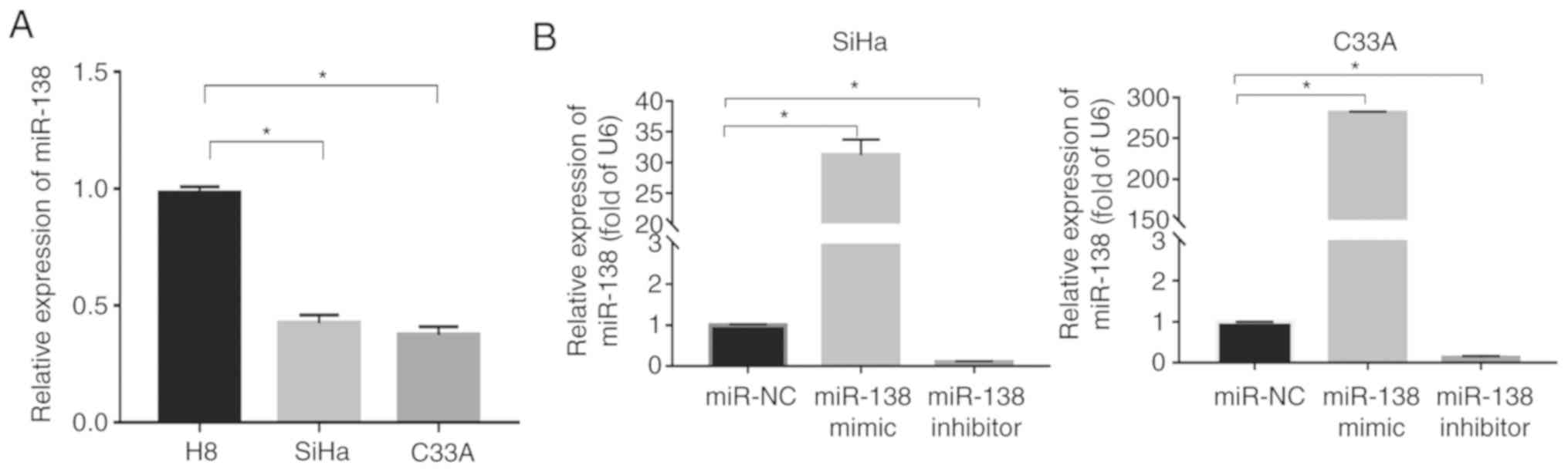
miR-138 expression is downregulated in
human cervical cancer cell lines
Reverse transcription (RT)-PCR was used to evaluate
miR-138 expression in immortalized normal cervical epithelial
squamous cell line H8 and cervical squamous cancer cell lines (SiHa
and C33A). The results revealed that the miR-138 expression in the
SiHa and C33A cells was significantly downregulated compared with
in the H8 cells (P<0.05; Fig.
1A).
miR-138 inhibits cell proliferation
and induces apoptosis in vitro
To investigate the role of miR-138 in cervical
cancer, the miR-138 mimic, miR-138 inhibitor and miR-NC were first
transfected into SiHa and C33A cells. RT-PCR was used to detect the
transfection efficiency. As shown in Fig. 1B, the level of miR-138 significantly
increased in the cells transfected with miR-138 mimic (P<0.05),
but the miR-138 inhibitor exhibited the opposite effect in both
SiHa and C33A cells.
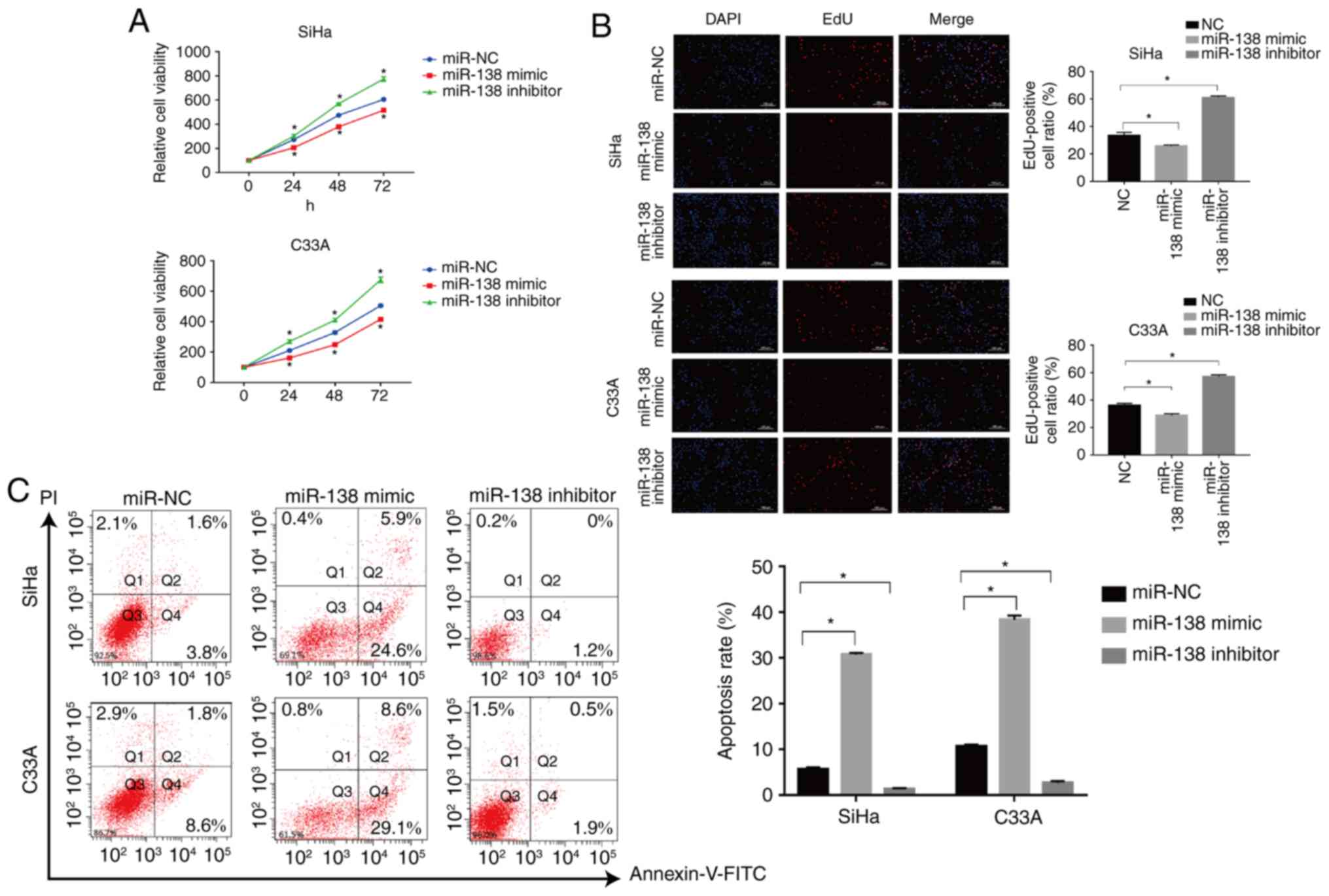
The CCK-8 and EdU assays were used to assess the
proliferation of transfected SiHa and C33A cells for the indicated
times. The miR-138 mimic significantly suppressed the proliferation
of SiHa cells compared with the miR-NC-transfected cells, but
miR-138 inhibitor significantly enhanced the proliferation of the
cells (P<0.05; Fig. 2A). The
effects of miR-138 were also validated with EdU staining. The
number of EdU positive cells was significantly decreased in the
miR-138 mimic-transfected group (P<0.05), while the number of
EdU positive cells in miR-138 inhibitor-transfected group was
significantly increased when compared with the miR-NC group in the
SiHa cells, as shown in Fig. 2B
(P<0.05). Similar results were also found in C33A cells. Flow
cytometric analysis was used to assess the apoptosis of SiHa and
C33A cells. As shown in Fig. 2C, the
miR-138 mimic significantly induced apoptosis of SiHa cells when
compared with the miR-NC group (P<0.05), but the miR-138
inhibitor significantly inhibited apoptosis (P<0.05). Similar
results were also found in C33A cells. In addition, the expression
of important proteins involved in apoptosis were also examined. The
miR-138 mimic increased the expression of caspase-3 and Bax, and
suppressed the expression of Bcl-2. By contrast, the miR-138
inhibitor significantly inhibited the expression of caspase-3 and
Bax but promoted the expression of Bcl-2 (P<0.05; Fig. 3C). Taken together, these findings
indicated that miR-138 inhibited proliferation and induced
apoptosis in cervical cancer cells.
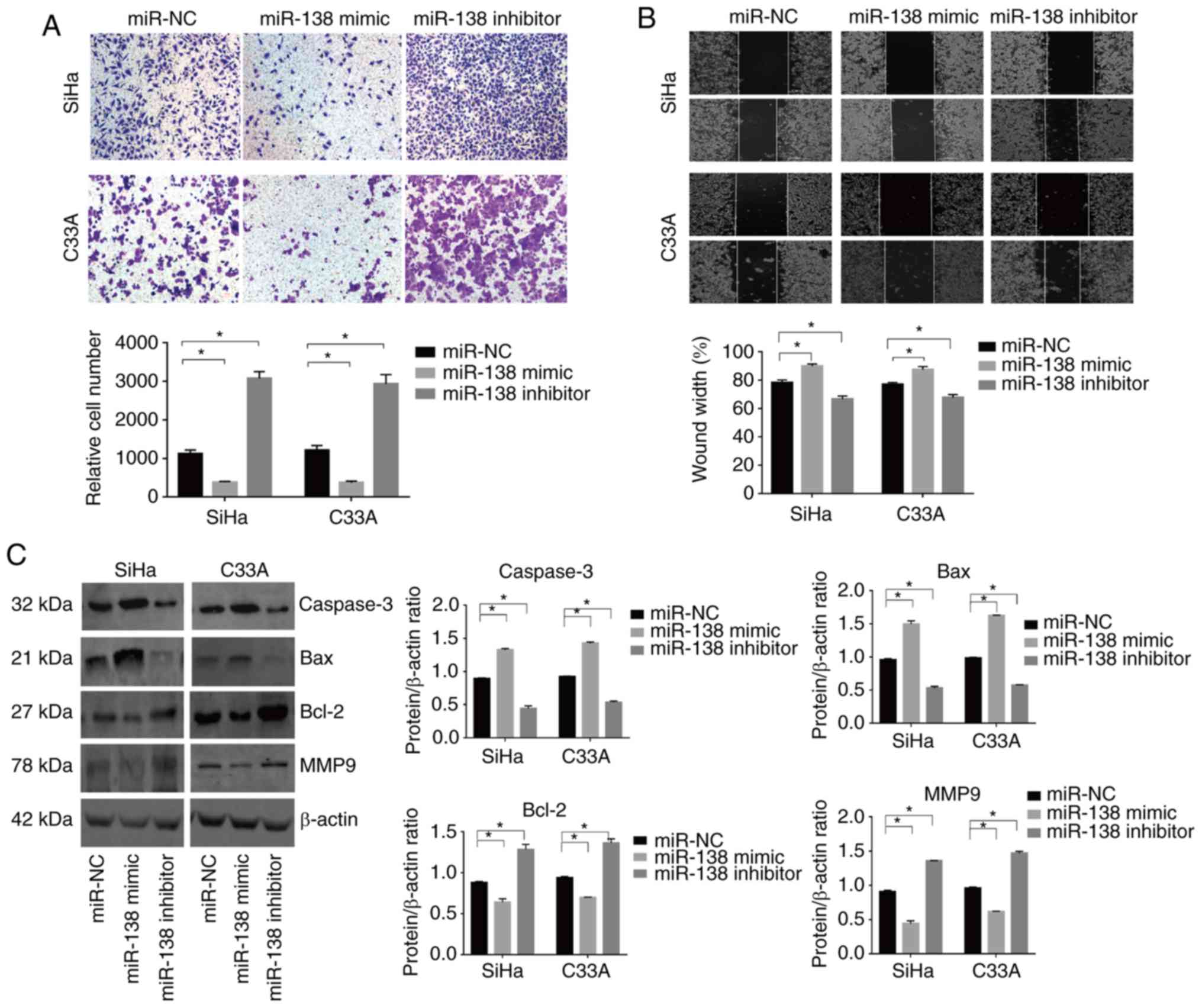
miR-138 inhibits cell mobility in
vitro
Transwell and wound-healing assays were used to
evaluate the effect of miR-138 on the mobility of cervical cancer
cells. The miR-138 mimic significantly decreased the number of
invading cells in the SiHa and C33A lines. By contrast, the miR-138
inhibitor exhibited the opposite effect (Fig. 3A and B). In addition, the expression
of important proteins involved in metastasis were examined. The
expression of MMP9 was decreased in the miR-138 mimic-transfected
cells, though the miR-138 inhibitor induced the opposite effect
(Fig. 3D).
miR-138 inhibits cell proliferation,
invasion and induces apoptosis in vivo
To evaluate whether the inhibitory effect of miR-138
on cell proliferation was also present in vivo, a tumor
xenograft study was performed. A lentiviral miR-138 vector or empty
vector was transfected into SiHa cells. The tumor volumes in the
miR-138 overexpression group were significantly reduced compared
with those in the empty vector group (P<0.05; Fig. 4A and B). Tumor weights in the miR-138
overexpression group were also reduced compared with in the empty
vector group (Fig. 4C). Furthermore,
miR-138-overexpressing tumors showed increased expression levels of
caspase-3 and cleaved caspase-3 compared with the empty vector, as
determined by IHC. By contrast, the expression of MMP9 was
significantly inhibited in tumors with miR-138 overexpression,
suggesting that the effect was still present in vivo
(P<0.05; Fig. 4D).
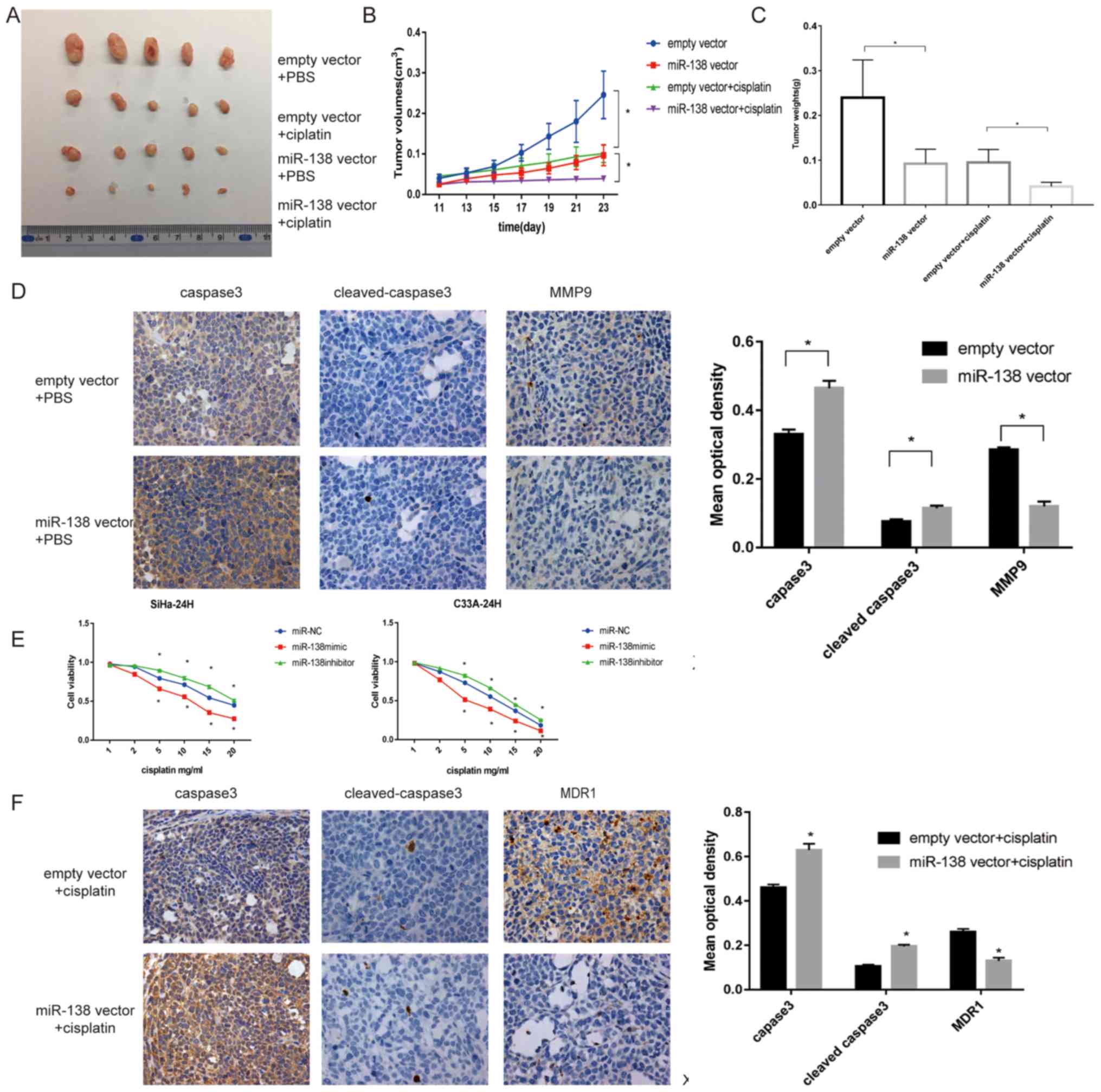 | Figure 4.miR-138 suppresses cell proliferation
and invasion and increases apoptosis in vivo and promoted
the sensitivity of cervical cancer cells in vitro and in
vivo. (A) Representative images of tumors isolated from nude
mice (n=20). (B) Weight of tumors treated with empty vector or
miR-138 vector injected with PBS and cisplatin were recorded and
presented as the mean ± standard deviation (n=20). (C) Tumor
volumes were measured every 2 days, 7 days after injection. (D)
Representative images of IHC-stained tumor specimens for apoptotic
proteins caspase-3, cleaved caspase-3 and bax, which were markedly
upregulated in the miR-138-SiHa group. MMP9, an invasive protein,
was markedly decreased in miR-138 vector tumors. Magnification,
×200. (E) Proliferation of transfected cells following treatment
with different concentrations of cisplatin, as examined by Cell
Counting Kit-8 assay. (F) Representative expression of caspase-3,
cleaved caspase-3 and MDR1 in the three groups of tumor xenografts,
as determined by IHC. Magnification, ×200. Data are presented as
the mean ± standard deviation. *P<0.05. miR, microRNA; MMP,
matrix metalloproteinase; IHC, immunohistochemistry. |
miR-138 promotes the sensitivity in
vitro and in vivo
To investigate whether miR-138 affects the
sensitivity of cervical cancer cells, cells were subjected to
different doses of cisplatin and a CCK-8 assay was used to measure
cell viability at the indicated times. As shown in Fig. 4E, the miR-138 mimic significantly
improved the toxicity of cisplatin compared with the miR-NC, while
the miR-138 inhibitor exhibited the opposite effect
(P<0.05).
Furthermore, whether this effect could also be
observed in vivo was investigated. As shown in Fig. 4B and C, following cisplatin
treatment, tumor growth derived from miR-138 vector cells was
significantly delayed compared with the group that received
cisplatin only (P<0.05). IHC revealed that miR-138 increased
caspase-3 and cleaved caspase-3 staining, but deceased MDR1
staining, which was consistent with the results in vitro
(Fig. 4F).
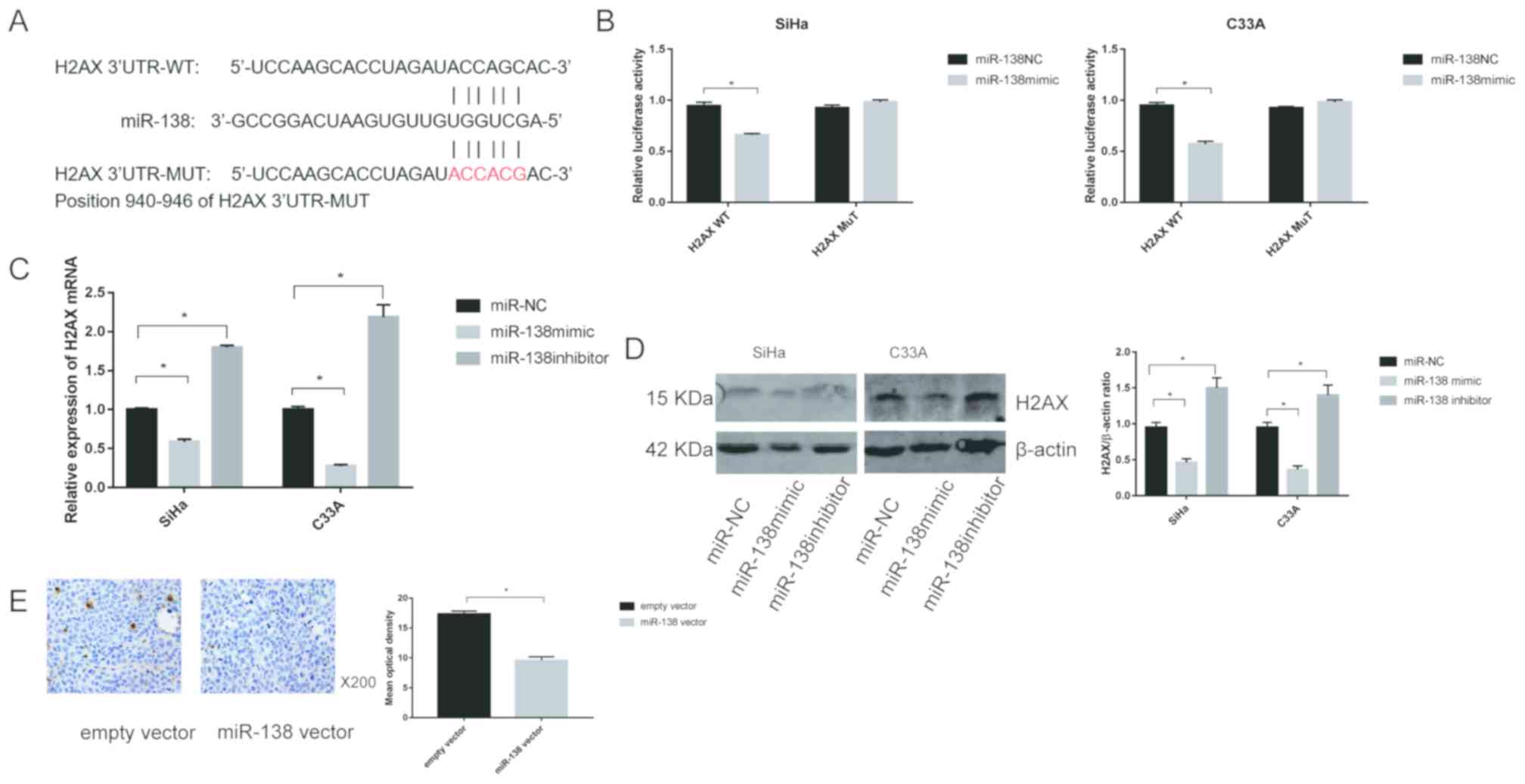
H2AX is a direct target of
miR-138
To further clarify the mechanisms of miR-138 in
cervical cancer, the miRNA prediction websites, TargetScan software
was used to screen for potential target of miR-138, and the H2AX
gene was selected as a potential candidate (Fig. 5A). Then, two types of plasmid
containing luciferase reporter was constructed to examine whether
miR-138 directly binds to H2AX. miR-138 significantly reduced the
luciferase activity of the plasmid with WT-H2AX (P<0.05), but
did not affect MUT-H2AX in SiHa and C33A cells, suggesting that
miR-138 could target H2AX in cervical cancer (Fig. 5B).
To evaluate the association between miR-138 and
H2AX, the expression of H2AX was detected by transfection with the
miR-138 mimic or inhibitor. The results showed that the mRNA levels
of H2AX were downregulated when miR-138 mimic was transfected into
both SiHa and C33A cells. In contrast, transfection with miR-138
inhibitor markedly enhanced the expression of H2AX mRNA in SiHa and
C33A cells (Fig. 5C). Similar
results were also observed for H2AX protein expression (Fig. 5D). In addition, the expression of
H2AX in vivo was assessed. As shown in Fig. 5E, IHC revealed that miR-138
significantly decreased the expression of H2AX compared with the
empty vector group.
H2AX is a functional target of
miR-138
To assess whether H2AX has functional relevance for
miR-138 expression, a rescue assay was generated. PcDNA 3.1-H2AX or
pcDNA3.1 and miR-138 mimic or NC were co-introduced into SiHa and
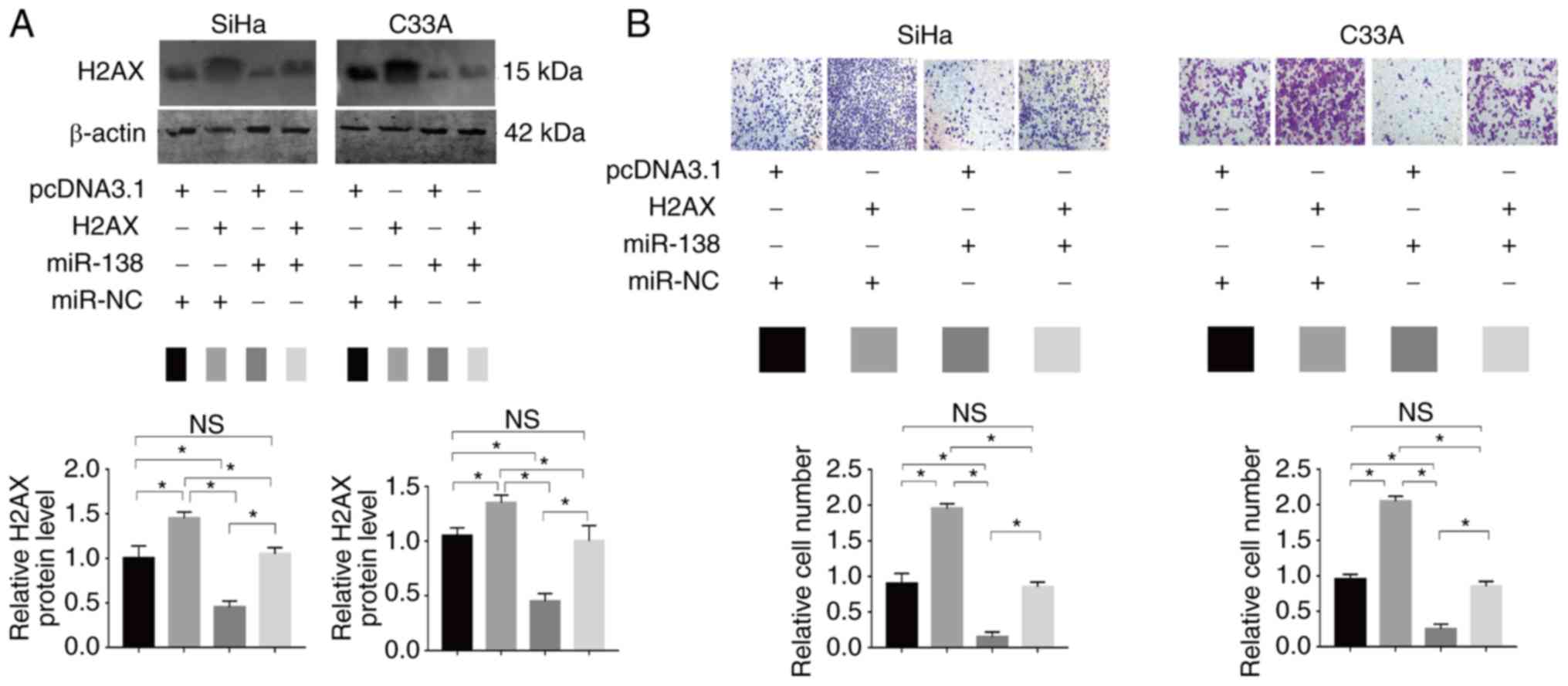
C33A cells (Fig. 6A). A Transwell
assay revealed that the enforced expression of H2AX partially
restored the migratory abilities of SiHa and C33A cells (Fig. 6B). The results suggested that H2AX is
a functional target of miR-138 in human cervical cancer.
Discussion
Uncontrolled tumor progression and chemoresistance
are the leading causes of poor treatment outcomes in cervical
cancer. Clarifying the mechanisms involved in cervical cancer cell
proliferation and chemoresistance is urgently required in order to
improve the survival rates of patients with cervical cancer.
miR-138 was previously reported to be dysregulated in various
malignancies, including non-small cell lung cancer (17) and colorectal cancer (18). In the present study, the results
revealed that miR-138 was downregulated in cervical cancer cell
lines. To investigate what role the downregulation of miR-138 plays
in cervical cancer, miR-138 mimic or inhibitor was transfected into
the cells. miR-138 suppressed cell viability, metastasis and
invasion, and enhanced apoptosis in cervical cancer cells. By
contrast, downregulation of miR-138 increased proliferation,
metastasis and invasion, and inhibited apoptosis. Xenograft
experiments also confirmed these findings in vivo. The
results were consistent with a previous report, which stated that
miR-138 functions as a tumor suppressor (19).
Downregulation of miR-138 has been demonstrated to
be involved in the chemoresistance of several cancer types
(20). Prior studies have
demonstrated that miR-138 modulate sensitization in prostate and
non-small cell lung cancer, multiple myeloma, renal cell carcinoma,
and osteosarcoma (14,21–25).
Consistent with previous studies, the results of the present study
showed that miR-138 enhanced the cisplatin-induced decrease in
cervical cancer cell viability. To characterize the effect of
miR-138 more precisely, xenograft models transfected with
lentiviral miR-138 vector or empty vector (negative control group)
were used. Tumor sizes and volumes were decreased following miR-138
overexpression and cisplatin treatment, as compared with in the
group that received cisplatin only, indicating that upregulation of
miR-138 sensitized cervical cancer cells to cisplatin. MDR1, which
has been investigated closely in relation to drug resistance, was
used to measure chemosensitivity (26). IHC was used to analyze apoptosis and
drug resistance. These results revealed that miR-138 induced
apoptosis and reversed drug resistance.
Identification of miRNA gene targets helps to
further understand the function of miRNA in carcinogenesis. Several
target genes of miR-138 had been revealed, including GIT1, PDK1,
Survivin and SIRT1 (27–30). The present data demonstrated that
H2AX acted as a functional target of miR-138 in cervical cancer.
H2AX, histone H2A variant phosphorylation, is an early marker of
DNA double-strand break (DSB) formation and regulates molecules
needed for DNA repair (31,32). DSBs signify genomic instability and
contribute to cancer initiation and progression. DNA damage occurs
in a variety of events, such as treatment with radiomimetic agents,
drugs. DNA damage can active three phosphatidylinositol
3-kinase-like kinases, which in turn phosphorylated H2AX at Ser
139. H2AX phosphorylated at the sites of DNA damage and facilitated
repair proteins, including the MRN complex, 53BP1, MDC1 and RAD51.
In addition to this function role as response to DSBs, H2AX also
play a structural role, by regulating cell proliferation (33), angiogenesis (34) and other biological processes. H2AX
can serve as an indicator of clonogenic survival and induction of
apoptosis. Radio-resistant tumor cells can be made sensitive to
radiation through blocking H2AX (35,36).
Previous studies had revealed that miR-138
negatively regulates the expression of H2AX in osteosarcoma
(37) and in small cell lung cancer
(38). miR-138 enhances cellular
sensitivity to DNA damaging agents by inhibiting H2AX. However, the
specific molecular mechanisms in cervical cancer have been unclear
and few studies have explored the relationship between H2AX and
chemosensitivity in cervical cancer. Consistent with previous
results, the present study revealed that miR-138 targets H2AX in
cervical cancer. Furthermore, a xenograft model was used to confirm
these findings. These results provide further evidence that H2AX is
a target of miR-138 in cervical cancer, not just in osteosarcoma
and small lung cancer.
In the current study, a luciferase reporter assay,
mRNA quantification, western blot analysis and animal experiments
demonstrated that miR-138 targets H2AX, and that an inverse
relationship exists between miR-138 and H2AX. Furthermore, a
functional study showed that H2AX expression was directly inhibited
by miR-138. Ectogenic overexpression of H2AX was able to rescue the
invasive abilities induced by miR-138, confirming that miR-138
targets H2AX. Moreover, functional studies revealed that H2AX
silencing induced miR-138 overexpression in cervical cancer,
resulting in the suppression of cell viability and migration. These
results suggested that H2AX is a functional mediator of
miR-138.
In conclusion, the present data revealed that
miR-138 was downregulated in cervical cancer. miR-138 inhibited
cell proliferation, invasion and migration, and enhanced drug
sensitivity by targeting H2AX in cervical cancer. The present
results not only provide insight into the molecular mechanisms that
regulate human cervical cancer, but also offer an explanation for
the challenges associated with chemoresistance. The study provides
evidence that miR-138 may have potential as a novel therapeutic
target in cervical cancer.
Acknowledgements
Not applicable.
Funding
The present study was supported by National Natural
Science Foundation of China (grant number 81360380).
Authors' contributions
QW and JC conceived and designed the experiments. MY
and SZ performed the experiments and constructed the table and
figures. RC, GW, YB contributed reagents/materials/analysis tools.
SZ wrote the paper. All authors reviewed the manuscript.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Ethics approval and consent to
participate
All experiments were approved by the Animal
Management Rule of the Chinese Ministry of Health (document 55,
2001) and approved by the Tongji University Animal Ethics Committee
(Shanghai, China). No human data were applicable in the present
manuscript.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Diaz-Padilla I, Monk BJ, Mackay HJ and
Oaknin A: Treatment of metastatic cervical cancer: Future
directions involving targeted agents. Crit Rev Oncol Hematol.
85:303–314. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Lukka H and Johnston M: Concurrent
cisplatin-based chemotherapy plus radiotherapy for cervical cancer:
A meta-analysis. Clin Oncol (R Coll Radiol). 16:160–161. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Pfaendler KS and Tewari KS: Changing
paradigms in the systemic treatment of advanced cervical cancer. Am
J Obstet Gynecol. 214:22–30. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Dasari S and Tchounwou PB: Cisplatin in
cancer therapy: Molecular mechanisms of action. Eur J Pharmacol.
740:364–378. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bayraktar R and Van Roosbroeck K: miR-155
in cancer drug resistance and as target for miRNA-based
therapeutics. Cancer Metastasis Rev. 37:33–44. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Huang YA, Hu P, Chan KCC and You ZH: Graph
convolution for predicting associations between miRNA and drug
resistance. Bioinformatics:. (pii): btz6212019. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Tung SL, Huang WC, Hsu FC, Yang ZP, Jang
TH, Chang JW, Chuang CM, Lai CR and Wang LH: miRNA-34c-5p inhibits
amphiregulin-induced ovarian cancer stemness and drug resistance
via downregulation of the AREG-EGFR-ERK pathway. Oncogenesis.
6:e3262017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ambros V: The functions of animal
microRNAs. Nature. 431:350–355. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhang Z, Wang J, Li J, Wang X and Song W:
MicroRNA-150 promotes cell proliferation, migration and invasion of
cervical cancer through targeting PDCD4. Biomed Pharmacother.
97:511–517. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wen SY, Lin Y, Yu YQ, Cao SJ, Zhang R,
Yang XM, Li J, Zhang YL, Wang YH, Ma MZ, et al: miR-506 acts as a
tumor suppressor by directly targeting the hedgehog pathway
transcription factor Gli3 in human cervical cancer. Oncogene.
34:717–725. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Xu R, Zeng G, Gao J, Ren Y, Zhang Z, Zhang
Q, Zhao J, Tao H and Li D: miR-138 suppresses the proliferation of
oral squamous cell carcinoma cells by targeting Yes-associated
protein 1. Oncol Rep. 34:2171–2178. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ye XW, Yu H, Jin YK, Jing XT, Xu M, Wan ZF
and Zhang XY: miR-138 inhibits proliferation by targeting
3-phosphoinositide-dependent protein kinase-1 in non-small cell
lung cancer cells. Clin Respir J. 9:27–33. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Stojcheva N, Schechtmann G, Sass S, Roth
P, Florea AM, Stefanski A, Stühler K, Wolter M, Müller NS, Theis
FJ, et al: MicroRNA-138 promotes acquired alkylator resistance in
glioblastoma by targeting the Bcl-2-interacting mediator BIM.
Oncotarget. 7:12937–12950. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C (T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Nie W, Huang W, Zhang W, Xu J, Song W,
Wang Y, Zhu A, Luo J, Huang G, Wang Y and Guan X: TXNIP interaction
with the Her-1/2 pathway contributes to overall survival in breast
cancer. Oncotarget. 6:3003–3012. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Gao Y, Fan X, Li W, Ping W, Deng Y and Fu
X: miR-138-5p reverses gefitinib resistance in non-small cell lung
cancer cells via negatively regulating G protein-coupled receptor
124. Biochem Biophys Res Commun. 446:179–186. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhao L, Yu H, Yi S, Peng X, Su P, Xiao Z,
Liu R, Tang A, Li X, Liu F and Shen S: The tumor suppressor
miR-138-5p targets PD-L1 in colorectal cancer. Oncotarget.
7:45370–45384. 2016.PubMed/NCBI
|
|
19
|
Zhu D, Gu L, Li Z, Jin W, Lu Q and Ren T:
MiR-138-5p suppresses lung adenocarcinoma cell
epithelial-mesenchymal transition, proliferation and metastasis by
targeting ZEB2. Pathol Res Pract. 215:861–872. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yu C, Wang M, Chen M, Huang Y and Jiang J:
Upregulation of microRNA1385p inhibits pancreatic cancer cell
migration and increases chemotherapy sensitivity. Mol Med Rep.
12:5135–5140. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Sossey-Alaoui K and Plow EF:
miR-138-mediated regulation of KINDLIN-2 expression modulates
sensitivity to chemotherapeutics. Mol Cancer Res. 14:228–238. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Tang X, Jiang J, Zhu J, He N and Tan J:
HOXA4-regulated miR-138 suppresses proliferation and gefitinib
resistance in non-small cell lung cancer. Mol Genet Genomics.
294:85–93. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Rastgoo N, Pourabdollah M, Abdi J, Reece D
and Chang H: Dysregulation of EZH2/miR-138 axis contributes to drug
resistance in multiple myeloma by downregulating RBPMS. Leukemia.
32:2471–2482. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yun EJ, Zhou J, Lin CJ, Xu S, Santoyo J,
Hernandez E, Lai CH, Lin H, He D and Hsieh JT: The network of
DAB2IP-miR-138 in regulating drug resistance of renal cell
carcinoma associated with stem-like phenotypes. Oncotarget.
8:66975–66986. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhu Z, Tang J, Wang J, Duan G, Zhou L and
Zhou X: MiR-138 acts as a tumor suppressor by targeting EZH2 and
enhances cisplatin-induced apoptosis in osteosarcoma cells. PLoS
One. 11:e1500262016.
|
|
26
|
Chen KG and Sikic BI: Molecular pathways:
Regulation and therapeutic implications of multidrug resistance.
Clin Cancer Res. 18:1863–1869. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wang B, Wang D, Yan T and Yuan H:
MiR-138-5p promotes TNF-α-induced apoptosis in human intervertebral
disc degeneration by targeting SIRT1 through PTEN/PI3K/Akt
signaling. Exp Cell Res. 345:199–205. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Yang R, Liu M, Liang H, Guo S, Guo X, Yuan
M, Lian H, Yan X, Zhang S, Chen X, et al: miR-138-5p contributes to
cell proliferation and invasion by targeting Survivin in bladder
cancer cells. Mol Cancer. 15:822016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Liu Y, Yang K, Sun X, Fang P, Shi H, Xu J,
Xie M and Li M: MiR-138 suppresses airway smooth muscle cell
proliferation through the PI3K/AKT signaling pathway by targeting
PDK1. Exp Lung Res. 41:363–369. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Li J, Wang Q, Wen R, Liang J, Zhong X,
Yang W, Su D and Tang J: MiR-138 inhibits cell proliferation and
reverses epithelial-mesenchymal transition in non-small cell lung
cancer cells by targeting GIT1 and SEMA4C. J Cell Mol Med.
19:2793–2805. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Siddiqui MS, Francois M, Fenech MF and
Leifert WR: Persistent γH2AX: A promising molecular marker of DNA
damage and aging. Mutat Res Rev Mutat Res. 766:1–19. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Turinetto V and Giachino C: Multiple
facets of histone variant H2AX: A DNA double-strand-break marker
with several biological functions. Nucleic Acids Res. 43:2489–2498.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Seo J, Kim K, Chang DY, Kang HB, Shin EC,
Kwon J and Choi JK: Genome-wide reorganization of histone H2AX
toward particular fragile sites on cell activation. Nucleic Acids
Res. 42:1016–1025. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Xiao H, Tong R, Ding C, Lv Z, Du C, Peng
C, Cheng S, Xie H, Zhou L, Wu J and Zheng S: γ-H2AX promotes
hepatocellular carcinoma angiogenesis via EGFR/HIF-1α-/VEGF
pathways under hypoxic condition. Oncotarget. 6:2180–2192. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Bassing CH, Chua KF, Sekiguchi J, Suh H,
Whitlow SR, Fleming JC, Monroe BC, Ciccone DN, Yan C, Vlasakova K,
et al: Increased ionizing radiation sensitivity and genomic
instability in the absence of histone H2AX. Proc Natl Acad Sci USA.
99:8173–8178. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Banáth JP, Macphail SH and Olive PL:
Radiation sensitivity, H2AX phosphorylation and kinetics of repair
of DNA strand breaks in irradiated cervical cancer cell lines.
Cancer Res. 64:7144–7149. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wang Y, Huang JW, Li M, Cavenee WK,
Mitchell PS, Zhou X, Tewari M, Furnari FB and Taniguchi T:
MicroRNA-138 modulates DNA damage response by repressing histone
H2AX expression. Mol Cancer Res. 9:1100–1111. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Yang H, Luo J, Liu Z, Zhou R and Luo H:
MicroRNA-138 regulates DNA damage response in small cell lung
cancer cells by directly targeting H2AX. Cancer Invest. 33:126–136.
2015. View Article : Google Scholar : PubMed/NCBI
|