Introduction
Sepsis, or septic shock, is a systemic response to
infection and causes excessive microvascular coagulation that leads
to disseminated intravascular coagulation (DIC). DIC is closely
associated with the development of multiple organ dysfunction
(1). Sepsis is caused by infectious
agents that are induced by lipopolysaccharides (LPS), viruses,
bacteria and a number of other infections. LPS injections have
commonly been used to generate experimental sepsis models (2). During sepsis, LPS causes an
inflammatory response that results in the release of a large number
of inflammatory cytokines, including tumor necrosis factor-α
(TNF-α) and interleukin-6 (IL-6) (3). The inflammatory responses may lead to
the activation of the coagulation system, the downregulation of
anticoagulant proteins and the inhibition of the fibrinolytic
system (4).
Coagulation abnormalities may occur in 50–70% of
patients with sepsis (4,5). Coagulation activation is one of the
most important characteristics in the pathophysiological of sepsis,
and thrombin generation serves a key role in the activation of
coagulation. Prothrombin fragment 1+2 (F1+2) is the earliest
molecular marker for prothrombin activation. In the present study,
F1+2 and thrombin-antithrombin complex (TAT) were selected to
detect the early stages of coagulation (6). The coagulation system is stimulated
during sepsis, and the corresponding anticoagulation system is
activated, which is followed by the profibrinolytic and
antifibrinolytic systems (4,5). Antithrombin (AT), which is also known
as AT-III, is a physiological inhibitor of serine proteases that
also exerts an inhibitory effect on other coagulation factors and
serine proteases (7). AT mitigates
vascular leakage by inhibiting neutrophil activation during acute
lung injury (7). The initiation of
fibrinolysis is mediated by plasminogen activators and is regulated
by plasminogen activator inhibitor-1 (PAI-1). PAI-1, which
indicates poor patient prognosis in sepsis-induced DIC, mediates
the inhibition of endogenous fibrinolysis during sepsis (8).
Endothelial activation and dysfunction are important
features of sepsis (3). The
endothelium responds to LPS or cytokines by exhibiting structural
changes and functional changes, including leukocyte infiltration,
vasodilation, increased vascular permeability and plasma protein
exudation and platelet adhesion (9).
The endothelial glycocalyx is a complex network of cell-bound
proteoglycans that are located on the surface of endothelial cells
and this network is one of the earliest sites that is associated
with sepsis (10,11). The endothelial glycocalyx contains
anchor protein syndecan-1 (SDC-1), heparan sulfate (HS) and other
glycosaminoglycans (GAGs), and is an anti-adhesive and
anticoagulant layer that protects vascular endothelial cells and
maintains vascular integrity (12).
Heparanase (HPA) is an endo-β-D-glucuronidase that degrades HS side
chains at specific intra-chain sites. Nadir et al (13) demonstrated that HPA increased
coagulation activity via the stimulation of tissue factor (TF)
expression in endothelial and cancer cells. Schmidt et al
(14) demonstrated that pulmonary
endothelial glycocalyx serves an important role in regulating
neutrophils adhesion. However, the modes of activation of HPA and
glycocalyx degradation products, and their association with
coagulation, remain largely undetermined.
Unfractionated heparin (UFH) is a glycosaminoglycan
that is largely used as anti-thrombotic and anticoagulant drug
since its identification over 100 years ago. The anti-inflammatory
properties and anticancer activity of UFH have been studied
extensively, it has been previously indicated that UFH inhibited
the activation of nuclear factor-κB (NF-κB) induced by LPS
(15). The efficacy and safety of
heparin use in patients with sepsis remains controversial, and
these patients have high risk of hemorrhage (16). NAH, a non-anticoagulant heparin
derivative, binds histones, prevents histone-mediated cytotoxicity
in vitro and has been demonstrated to improve mortality in
LPS/CLP induced sepsis mouse models (17). A previous study demonstrated that
heparin, as the competitive antagonist, inhibited the activity of
HPA, an endogenous HS-specific glucuronidase, and prevented
LPS-induced endothelial glycocalyx loss (14).
The present study aimed to explore the association
between the products of glycocalyx degradation and coagulation in a
sepsis rat model. Secondly, the present study aimed to evaluate the
effect of UFH and NAH, a non-anticoagulant heparin derivative, on
endothelial glycocalyx and coagulation function in an LPS-induced
sepsis rat model, and to compare the differences in coagulation
function between UFH and NAH.
Materials and methods
Animals
Male Sprague-Dawley rats (6–8 weeks; weight, 180–220
g) were obtained from the Model Animal Research Center of Nanjing
University. All animals were housed in standard conditions (22±2°C;
50±10% relative humidity; 12:12 h light: Dark cycle). The rats had
ad libitum access to food and water. The rats acclimated to
the environment for 3–5 days prior to the experiment.
The animal care and experimental procedures were
conducted in accordance with the Guide for the Care and Use of
Laboratory Animals, and the protocol was approved by the
Institutional Animal Care and Use Committee of Binzhou Medical
University Hospital.
Reagents and antibodies
LPS (Escherichia coli LPS 055:B5), UFH and
NAH were purchased from Sigma-Aldrich; Merck KGaA. ELISA kits for
rat TNF-α (cat. no. CSB-E11987r), IL-6 (cat. no. CSB-E04640r), F1+2
(cat. no. CSB-E13264r), TAT (cat. no. CSB-E08432r), AT (cat. no.
CSB-E13885r) and PAI-1 (cat. no. CSB-E07948r) were purchased from
Cusabio Technology LLC. SDC-1 (cat. no. E02S0301, Shanghai BlueGene
Biotech Co., Ltd.) level and Heparin sulfate (HS; cat. no.
DG94646Q; Beijing Donggeboye Biological Technology Co., Ltd.) were
determined using an ELISA according to the manufacturer's protocol.
Fluorescence decay-resistant medium (cat. no. S2100) was purchased
from Beijing Solarbio Science & Technology Co., Ltd.. Mouse
Anti-Heparan Sulfate (10E4 epitope) antibody (cat. no. 370255-1)
was purchased from United States Biological. The anti-SDC-1
antibody (cat. no. ab128936) was purchased from Abcam. The goat
polyclonal thrombomodulin (BDCA-3) antibody (cat. no. AF3894) was
purchased from R&D Systems, Inc. Rhodamine-conjugated anti-rat
IgG (cat. no. ZF-0318) was purchased from ZSGB-BIO (Beijing, China)
and fluorescein isothiocyanate (FITC)-conjugated anti-goat IgG
(cat. no. TA130029) was purchased from OriGene Technologies,
Inc.
Animal model of sepsis
Experimental rats were randomly allocated into the
control, LPS, UFH + LPS and NAH + LPS groups. The rats in the LPS
group received an intravenous (i.v.) injection of 10 mg/kg ml LPS.
The dose of LPS in rats was performed as previously described
(18) to establish the model of
sepsis. The rats in the UFH + LPS group and NAH + LPS group
received a tail intravenous injection of 100 U/kg body weight UFH
(19) or 1 mg/kg body weight NAH
(14) diluted in 1 ml normal saline
(NS), while the control rats received vehicle NS only. UFH or NAH
were administered concomitantly and this was followed ~1 min later
by the addition of LPS (10 mg/kg; 1 ml/kg of body weight; i.v.).
The rats in the control group were injected with NS instead of LPS
in the same manner. All rats remained alive after 0.5, 2 and 6 h of
LPS stimulation. Following this, the rats were sacrificed by
cervical dislocation following anesthesia by peritoneal injection
of 1% sodium pentobarbital (40 mg/kg body weight). Plasma and lung
tissue samples were then harvested.
ELISA for TNF-α, IL-6, SDC-1, HS,
F1+2, TAT, AT and PAI-1
Blood was collected from the harvested abdominal
aorta and stored in an anticoagulant tube and centrifuged at 4°C
for 20 min at 900 × g. Collected plasma samples were stored at
−80°C. TNF-α, IL-6, SDC-1, HS, F1+2, TAT, AT and PAI-1 were
evaluated with the corresponding ELISA kits in accordance with the
manufacturer's protocol.
HPA activity assay
Plasma samples were collected at 0.5, 2 and 6 h
after LPS administration and HPA activity was measured. The blood
was collected from the harvested abdominal aorta and stored in an
anticoagulant tube and centrifuged at 4°C for 20 min at 900 × g.
Collected plasma samples were stored at −80°C. The HPA activities
were measured using an Heparan Degrading Enzyme Assay kit (cat. no.
MK412; Takara Bio, Inc.) according to the manufacturer's
protocol.
Blood coagulation tests
Blood was collected and the plasma was separated as
aforementioned. The prothrombin time (PT), activated partial
thromboplastin time (APTT) and levels of fibrinogen (FIB) were
measured using a coagulometer (CS-5100; Sysmex Corporation).
According to the International Society of Thrombosis and Hemostasis
(ISTH) scoring system, the PT, APTT, FIB, AT and TAT have been used
to diagnostic screening of coagulation function for DIC (20).
Pulmonary histologic examination
The lower lobe, collected from the right lung, was
fixed in 4% paraformaldehyde for 48 h at 4°C and embedded in
paraffin. Sections (5 µm) were stained with hematoxylin for 5 min
at room temperature and then with eosin for 3 min at room
temperature. Tissue sections were observed under an optical
microscope. Hematoxylin and eosin staining was performed according
to the standard method used to assess lung injury. The lung injury
scores (LIS) were calculated as previously described by Aeffner
et al (21). ALI was scored
as follows: i) Alveolar congestion; ii) hemorrhage; iii)
infiltration or aggregation of neutrophils in the airspace or
vessel wall; and iv) thickness of alveolar wall/hyaline membrane
formation. Each item was scored on a 5-point scale as follows: 0,
minimal damage; 1, mild damage; 2, moderate damage; 3, severe
damage; and 4, maximal damage. In each stained sample, six
high-magnification fields were selected randomly, then graded for
average LIS. Repeated-measures data were statistically analyzed
using a repeated-measures analysis of variance (ANOVA).
Measurement of lung wet/dry (W/D)
weight ratio
To assess the magnitude of lung tissue edema, the
lung W/D weight ratio was calculated. A period of 6 h after LPS
administration, rats were euthanized and left lungs were excised
using blunt dissection, weighed to obtain the ‘wet’ weight. They
were subsequently placed in an oven at 60°C for 48 h to acquire the
‘dry’ weight.
Immunofluorescence
The lung tissues were fixed using 4%
paraformaldehyde for 48 h at 4°C, dehydrated with alcohol (100% for
20 min, 95% for 10 min, 85% for 10 min, 75% for 10 min and 50% for
10 min) and embedded with paraffin and cut into 4 µm sections. The
sections were then incubated with mouse anti-heparan sulfate (10E4
epitope; cat. no. 370255-1; 1:200) or anti-SDC-1 antibody (cat. no.
ab128936; 1:463) overnight at 4°C. Sequentially, the sections were
incubated for 1 h at 37°C, washed 3 times with PBS and incubated
with rhodamine-conjugated anti-rat IgG (cat. no. ZF-0318; 1:200)
for 1 h at room temperature. The sections were then washed 3 times
with PBS for 10 min each time and incubated with secondary antibody
goat polyclonal against BDCA-3 (cat. no. AF3894; 1:200) overnight
at 4°C. Sequentially, sections were washed again and incubated with
FITC-conjugated anti-goat IgG (cat. no. TA130029; 1:200) for 1 h at
room temperature. The sections were washed again and incubated with
DAPI (1 mg/ml, 1:300) for 8 min at room temperature. Finally,
sections were washed and blocked with fluorescence decay-resistant
medium for 1 min at room temperature. Sections were observed using
a fluorescence microscope (×200; Olympus BX53; Olympus
Corporation). Data were quantified using Image J 1.37v (National
Institutes of Health), and the results were statistically analyzed
using SPSS 25.0 (IBM Corp).
Statistical analysis
The data are expressed as the mean ± standard
deviation. The LIS, lung W/D weight ratio, IL-6, HPA, HS, SDC-1,
TAT and PAI-1 data were normally distributed, so statistical
comparisons were determined using a one-way ANOVA, followed by the
Student-Newman-Keuls (SNK) test for multiple group comparison. The
TNF-α, F1+2 and AT data were nonnormally distributed, so
statistical comparisons were determined using a Kruskal-Wallis test
along with Bonferroni correction. The associations between
inflammation parameters, glycocalyx degradation products, and
coagulation parameters were determined using a Spearman rank
correlation test. Statistical analyses was conducted using SPSS
v.25.0 (IBM Corp). P<0.05 was considered to indicate a
statistically significant difference.
Results
Preconditioning with heparin
attenuated LPS-induced lung injury
The primary organ affected during sepsis is the lung
(14). Previous studies have
demonstrated that UFH and NAH may decrease the severity of
sepsis-associated acute lung injury and lethality in LPS-induced
mice (17,19,22).
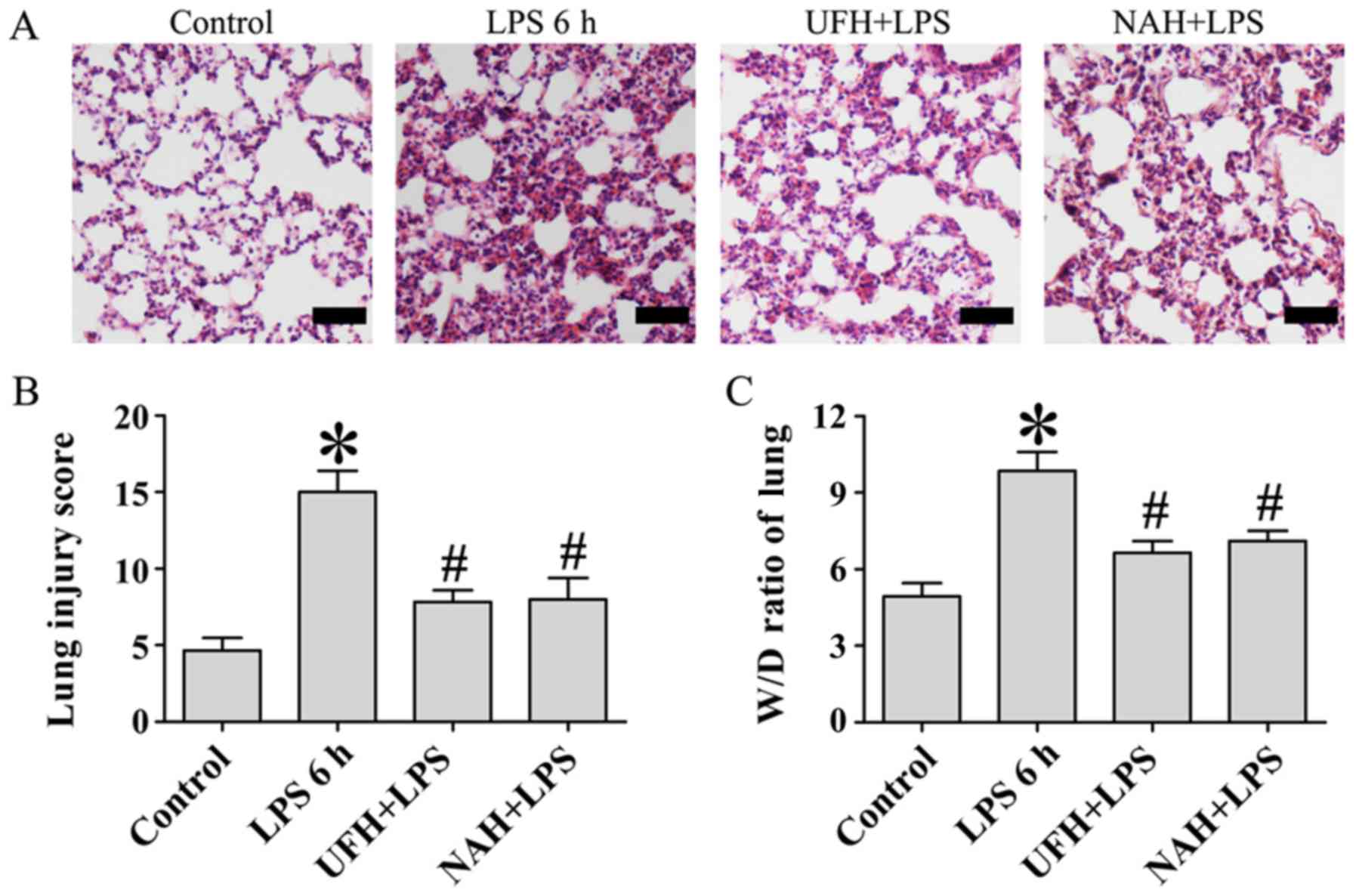
Therefore, pulmonary histopathology was examined using hematoxylin
and eosin staining and lung injury score to assess the protective
effect of UFH and NAH (Fig. 1A and
B). The lung tissues in the LPS group were characterized by
alveolar capillary barrier integrity, alveolar epithelial injury,
interstitial infiltration by neutrophils, alveolar edema, and
fibrin deposition. However, LPS-induced histopathological changes
were significantly alleviated by preconditioning with UFH and NAH
following LPS stimulation, which was indicated by decreased
neutrophil infiltration and protein leakage and the improved
integrity of alveolar capillary barrier.
Pulmonary edema is one of the primary features of
acute respiratory distress syndrome (ARDS) (14). In the present study, lung W/D weight
ratios were used to independently evaluate LPS-induced changes in
pulmonary vascular permeability to water. A period of 6 h after LPS
injection, the lung W/D ratio was significantly decreased compared
with the control group. Preconditioning with UFH or NAH inhibited
this increase in the lung W/D ratio compared with the LPS group
(Fig. 1C). These results
demonstrated that UFH and NAH demonstrated a protective effect on
LPS-induced acute lung injury in rats.
Plasma inflammatory cytokines, HPA
activity, glycocalyx components and coagulation/fibrinolysis
markers in rats with LPS-induced sepsis
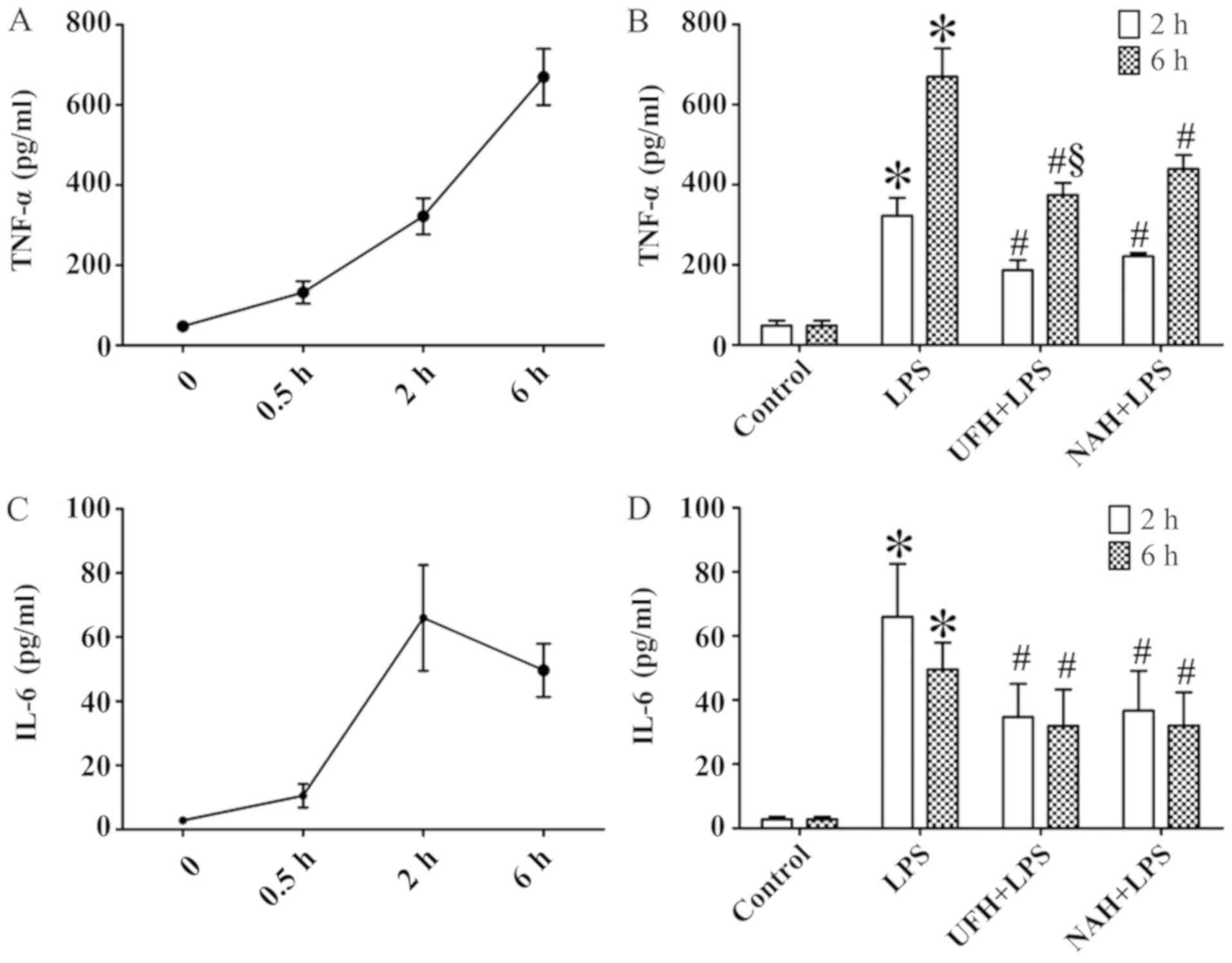
In the LPS group, the concentration of TNF-α was
increased significantly from 2 to 6 h. The IL-6 level markedly
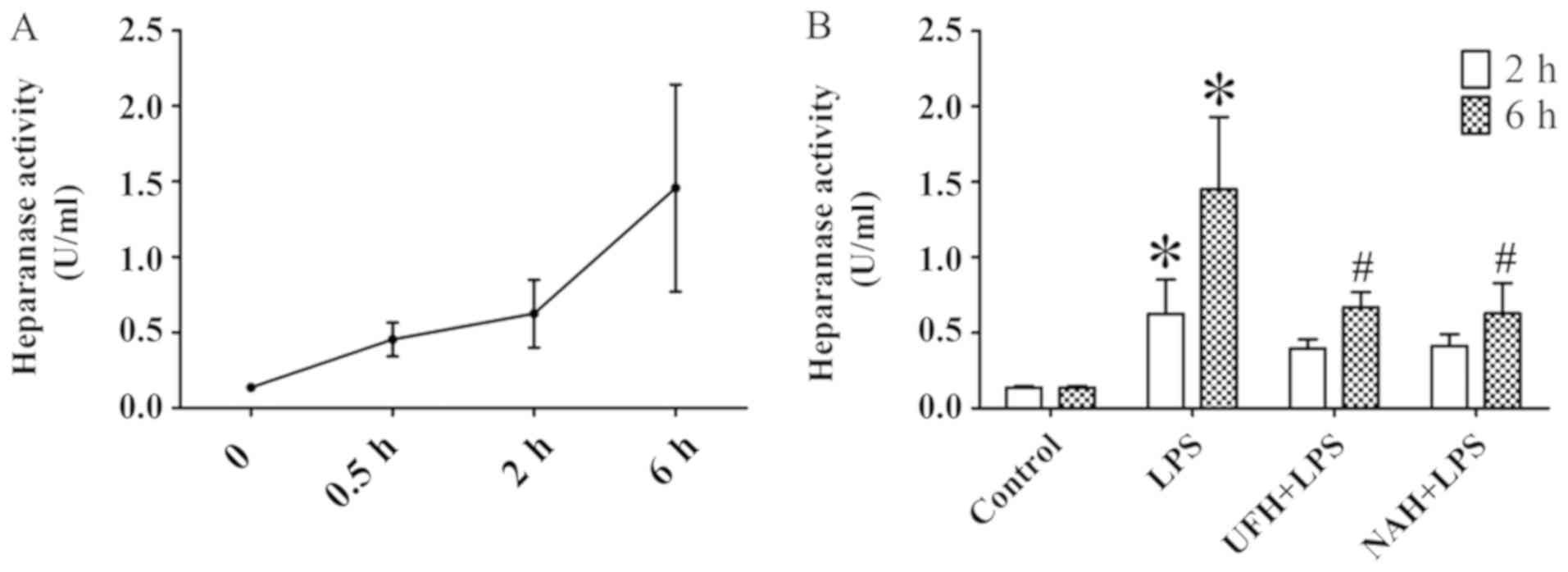
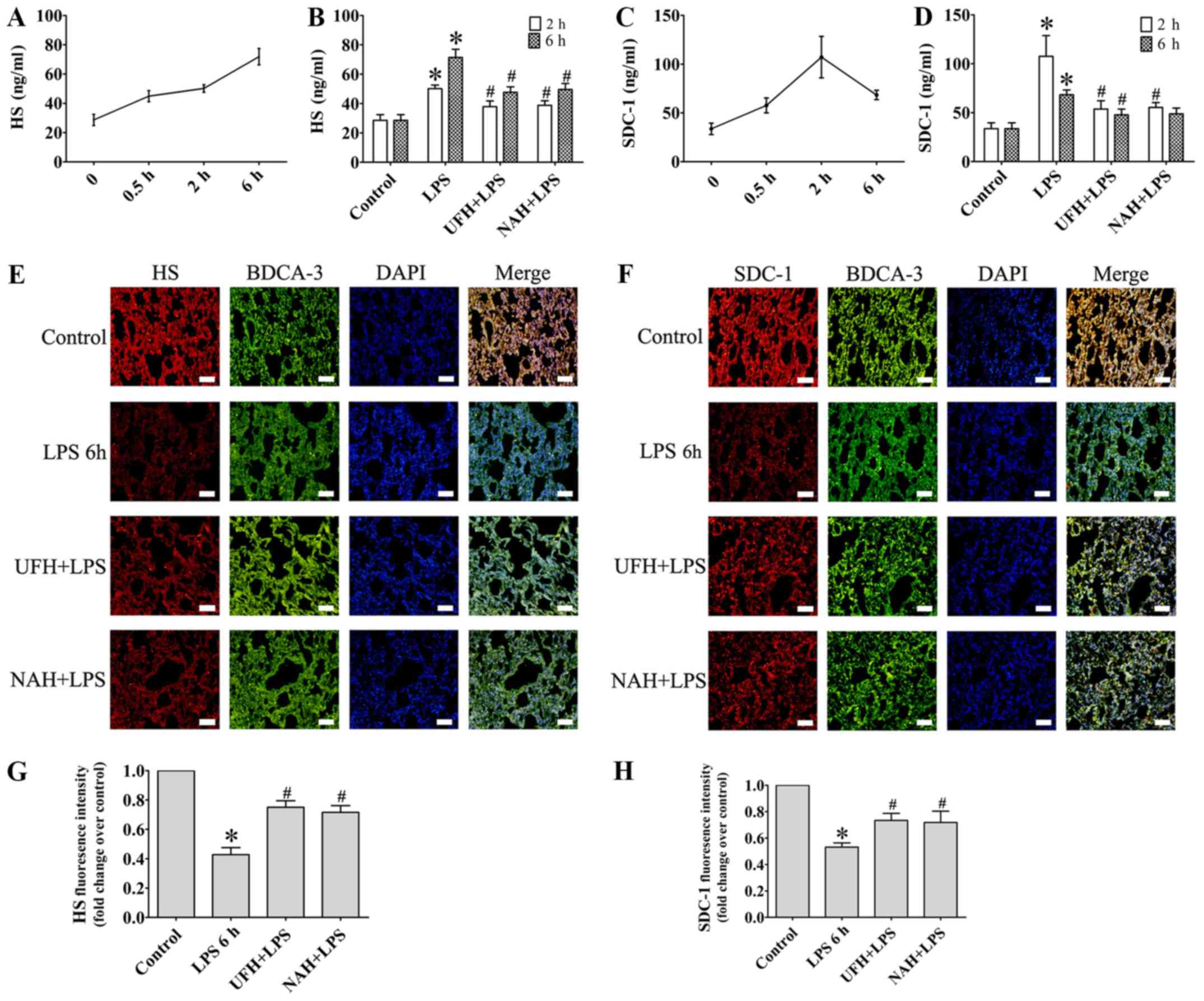
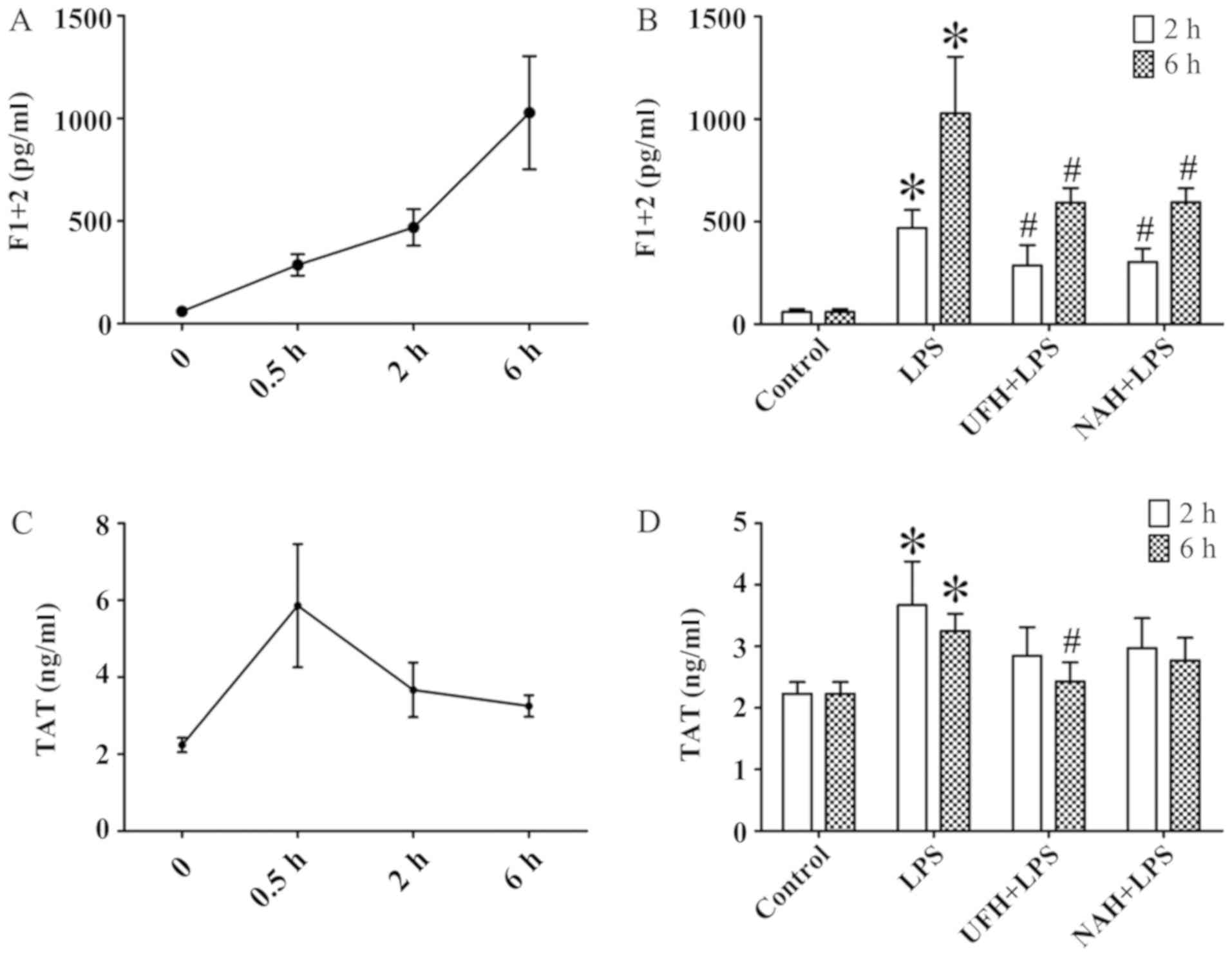
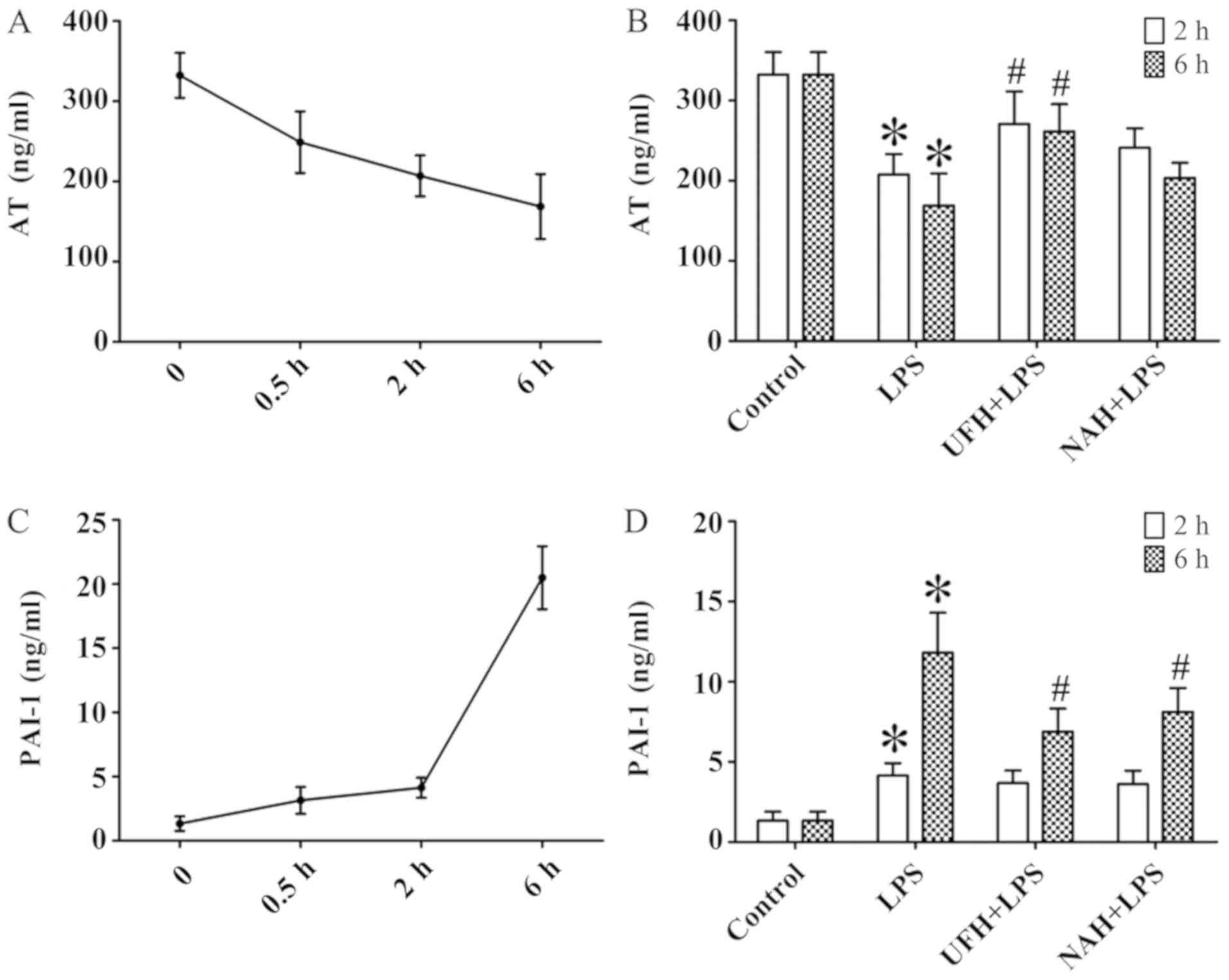
increased from 0.5 to 2 h and then decreased slowly (Fig. 2A and C). HPA activity (Fig. 3A) and HS levels (Fig. 4A) increased slowly from 0.5 to 2 h,
and then increased markedly from 2 to 6 h. The plasma level of
SDC-1 (Fig. 4C) increased sharply at
2 h and subsequently decreased slowly. The F1+2 level increased
significantly from 2 to 6 h (Fig.
5A). Additionally, TAT level increased markedly at 0.5 h and
subsequently decreased significantly and progressively decreased
from 2 to 6 h (Fig. 5C). The level
of AT gradually decreased, and PAI level gradually increased from
0.5 to 6 h (Fig. 6A and C). In the
present study, glycocalyx components (HS and SDC-1) and HPA
activity were positively correlated with inflammatory cytokines
(TNF-α and IL-6) and coagulation/fibrinolysis markers (F1+2 and
PAI-1), and were negatively correlated with AT anticoagulant
protein (Table I).
 | Table I.Correlations between glycocalyx
components and inflammatory cytokines and coagulation/fibrinolysis
markers. |
Table I.
Correlations between glycocalyx
components and inflammatory cytokines and coagulation/fibrinolysis
markers.
|
| HPA | HS | SDC-1 |
|---|
|
|
|
|
|
|---|
|
| rho | P-value | rho | P-value | rho | P-value |
|---|
| Inflammatory
cytokines |
|
|
|
|
|
|
|
TNF-α | 0.94 | <0.001 | 0.95 | <0.001 | 0.47 | 0.066 |
|
IL-6 | 0.83 | <0.001 | 0.47 | 0.066 | 0.72 | 0.002 |
|
Coagulation/fibrinolysis markers |
|
|
|
|
|
|
|
F1+2 | 0.92 | <0.001 | 0.88 | <0.001 | 0.45 | 0.08 |
|
TAT | 0.238 | 0.374 | 0.28 | 0.284 | 0.61 | 0.012 |
| AT | −0.847 | <0.001 | −0.78 | <0.001 | −0.447 | 0.083 |
|
PAI-1 | 0.918 | <0.001 | 0.79 | <0.001 | 0.294 | 0.269 |
Effect of heparin on plasma of
inflammatory cytokines in rats with LPS-induced sepsis
Preconditioning rats with UFH or NAH decreased
LPS-induced TNF-α and IL-6 expression levels after 2 and 6 h. The
effect of UFH on TNF-α levels was improved compared with that of
NAH (Fig. 2B and D).
Effect of heparin on HPA activity and
endothelial glycocalyx integrity in an LPS-induced sepsis rat
model
Compared with the control group, HPA activity in the
LPS group was significantly increased, whereas preconditioning with
UFH or NAH inhibited HPA activity compared with the LPS group
(Fig. 3B). The integrity of
endothelial glycocalyx was indicated by plasma HS and SDC-1 levels,
and the expression of HS and SDC-1 in the pulmonary vascular
endothelial cells. Preconditioning with UFH or NAH significantly
suppressed the shedding of HS and SDC-1 and decreased the plasma
levels of HS and SDC-1 in rats with sepsis. No statistically
significant difference was observed between the UFH + LPS and NAH +
LPS groups (Fig. 4B and D).
Effect of heparin on
coagulation/fibrinolysis markers in rats with LPS-induced
sepsis
The levels of F1+2 and TAT were markedly increased
in the LPS group compared with the control group. UFH and NAH
treatment also decreased LPS-induced F1+2 levels in the plasma at 2
and 6 h. UFH treatment decreased LPS-induced TAT deposition in the
plasma of sepsis rat models at 6 h. Preconditioning with NAH
decreased TAT level compared with the LPS group. However, the
effect was not statistically significant. No statistically
significant difference was observed between the UFH + LPS and NAH +
LPS groups (Fig. 5B and D).
Preconditioning with UFH increased the AT level compared with the
LPS group and NAH + LPS group. Preconditioning with NAH increased
the AT level compared with the LPS group. However, the effect was
not statistically significant. Preconditioning with UFH or NAH
decreased the PAI-1 level compared with the LPS group at 6 h after
LPS injection, and there was no statistically significant
difference observed between the UFH + LPS and NAH + LPS groups
(Fig. 6B and D).
Effect of heparin on APTT, PT and FIB
level in an LPS-induced sepsis rat model
The APTT was 20.1 sec (mean) in the control animals
and was significantly increased to 60.9 sec (mean) 6 h after the
LPS injection. Preconditioning with UFH significantly inhibited the
LPS-induced prolongation of the APTT to 49.6 sec (mean) at 6 h.
Preconditioning with NAH inhibited the LPS-induced prolongation of
the APPT to 55.6 sec (mean) at 6 h. However, the effect was not
statistically significant (Table
II).
| Table II.Effect of UFH or NAH on APTT, PT and
FIB level in LPS-induced sepsis rat model. |
Table II.
Effect of UFH or NAH on APTT, PT and
FIB level in LPS-induced sepsis rat model.
| Group | APTT, sec | PT, sec | FIB levels,
g/l |
|---|
| Control | 20.5±1.4 | 15.6±0.4 | 2.5±0.11 |
| LPS 6h |
60.9±5.8a | 18.3±1.9 |
1.3±0.23a |
| UFH + LPS |
49.6±4.6b,c | 18.3±1.0 |
1.9±0.34b |
| NAH + LPS | 55.6±5.9 | 18.7±1.2 |
1.7±0.24b |
The PT was 15.8 sec (mean) in the control animals
and was significantly increased to 18.3 sec (mean) 6 h following
the LPS injection. Preconditioning with UFH or NAH did not affect
the PT (Table II).
LPS injection decreased the plasma fibrinogen level,
which decreased to 54% of the normal level after 6 h.
Preconditioning with UFH or NAH significantly inhibited the
LPS-induced decrease in plasma fibrinogen level. However, the
effect was not statistically significant between the two groups
(Table II).
Discussion
Sepsis is a systemic inflammatory response syndrome
that is caused by infectious agents. A large number of mediators
can lead to the activation of the coagulation system and the
inhibition of anticoagulant mechanisms and fibrinolysis.
Additionally, the coagulation system may in turn affect the
inflammatory response (3,4). In the present study, in rats with
LPS-induced sepsis, plasma inflammation cytokines indicated a
positive correlation with coagulation parameters. Previous studies
have demonstrated that heparin decreased the levels of the
inflammatory cytokines TNF-α, IL-6, IL-8 and IL-1β in an
LPS-induced sepsis model (15,19,22).
Similar results were indicated in the present study, which
demonstrated that preconditioning with NAH or UFH inhibited the
LPS-induced cytokines of TNF-α and IL-6. The results also indicated
that UFH was more effective compared with NAH, and this may be due
to an increased level of coagulation activation, contributing to an
increase in the level of inflammation.
The glycocalyx is a negatively charged network that
is composed of core proteins and side chains that modulate the
microvascular environment, coagulation, thrombosis and vascular
permeability (23). Edema and
increased vascular permeability are the major characteristics of
ARDS, which is a common manifestation of sepsis-associated organ
dysfunction (24,25). The results of the present study
demonstrated that UFH and NAH significantly alleviated lung edema
and decreased the shedding of HS and SDC-1. Wildhagen et al
(17) and Li et al (19) demonstrated that UFH and NAH may
alleviate LPS induced lung pathological changes and lung edema
through downregulating the NF-κB signaling pathway and binding
histones. These results were consistent with those of previous
studies (17,19), which demonstrated that UFH and NAH
attenuated LPS-induced acute lung injury in rats. HPA is an
endo-β-glucuronidase that is involved in the cleavage of heparan
sulfate side chains, and serves an important role in inflammation
and mediates acute pulmonary and renal injuries during sepsis
(14,26). Heparan sulfates are the most common
endothelial cell surface GAG, comprising 50–90% of the GAG pool.
Schmidt et al (14) revealed
that HPA inhibition prevented LPS-associated glycocalyx shedding
and neutrophil adhesion and attenuated sepsis-induced acute lung
injury, and these data support the results of the present study.
Therefore, UFH and NAH may decrease shedding of HS due to the
inhibition of HPA activity. Syndecans serve important regulatory
roles in a number of biological processes, including inflammation,
calcium metabolism and wound healing, and the shedding of syndecans
is regulated by matrix metalloproteinases (MMPs) (27). Circulating levels of SDC-1 have been
indicated to represent the extent of endothelial damage and
glycocalyx degradation. A marked correlation was observed between
the levels of IL-6 and SDC-1 in the sepsis and surgery groups of a
previous study (28). In the present
study, glycocalyx degradation products in plasma revealed a
positive correlation with inflammatory factors, and the protective
effects of UFH and NAH were observed on SDC-1 shedding. This
protective effect may be due to the anti-inflammatory effects of
UFH or NAH and the subsequent inhibition MMPs activity. However,
the specific mechanisms governing this are yet to be
determined.
During sepsis, pathogenic agents and inflammatory
mediators regulate coagulation through at least 3 simultaneous
pathways: The activation of pro-coagulation pathways, the
downregulation of physiological anticoagulant production and the
inhibition of fibrinolysis (29).
F1+2 and TAT are regarded as the sensitive indicators of thrombin
generation (6). Consistent with
previous studies, an immediate increase in F1+2, TAT, APTT, PT and
PAI-1, and a decrease in levels of AT and FIB were observed in rat
plasma following LPS administration in the present study.
Additionally, preconditioning with UFH was indicated to decrease
the levels of F1+2, TAT and APTT and increase the levels of FIB and
AT in plasma. UFH has been indicated to exert an anticoagulant
effect by binding to the lysine site on AT, which is also modulated
by platelets, fibrin, thrombin, factors Xa, IXa, XIa and XIIa and
TF (30,31). Tipoe (8) demonstrated that high levels of PAI-1
may predict an adverse outcome in severe sepsis, and suppressed
fibrinolysis has been suggested to be one the most important
predictors of multiple organ dysfunction during DIC. The results of
the present study identified that preconditioning UFH attenuated
the level of PAI-1. However, NAH does not contain a binding site
for AT, but may improve the coagulopathy in LPS-induced sepsis.
This result may be attributed to NAH inhibiting of HPA activity and
decreasing the loss of glycocalyx. Sieve et al (12) indicated that HPA upregulated the
expression of blood coagulation initiator TF in endothelial and
tumor cells, resulting in increased cell surface coagulation
activity. Vascular endothelial glycocalyx includes anticoagulant
heparan sulfate, which is a small subpopulation (0.5–10%) of HS and
contains a specific pentasaccharide motif with high affinity for
plasma AT. Molecules of heparan sulfate combined with AT have been
demonstrated to prevent microvascular thrombosis and contribute to
the maintenance of microvascular patency (32). Vink et al (33) indicated that the specific disruption
of glycocalyx promoted thrombin generation and platelet adhesion
within a short time period. Chappell et al (34) indicated that protection of the
endothelial glycocalyx decreased platelet adhesion in an
ischemia/reperfusion pig model. Ikeda et al (35) revealed that SDC-1 levels were
associated with the severity of illness and mortality and with DIC
development in sepsis. These results support the suggestion that
glycocalyx damage may aggravate coagulation disorder, and the
inhibition of glycocalyx shedding may further improve
coagulation.
No specific pharmacotherapy is available for septic
coagulopathy, the efficacy of heparin for sepsis has not been well
established, and the use of heparin for sepsis may increase
hemorrhage. The present study indicated that NAH protected the
vascular endothelial glycocalyx and concomitantly improved partial
coagulation disorders. These results suggest that the inhibition of
glycocalyx shedding may be used for improving coagulation disorders
in sepsis. However, the present study lacks direct evidence to
identify the effect of the destruction of glycocalyx induced by HPA
on coagulation function. Additionally, the present study observed
the sepsis model over a short time period of 6 h. Therefore, future
studies should investigate the effects of glycocalyx components on
the coagulation/fibrinolytic system, and this will aid in the
determination of the pathogenesis of coagulation dysfunction in
sepsis.
In conclusion, the results of the present study
demonstrated that UFH and NAH alleviated coagulopathy and
glycocalyx loss, and that glycocalyx component (HS and SDC-1)
levels were significantly correlated with coagulation/fibrinolysis
markers in LPS-induced sepsis. These results indicated that NAH
treatment may alleviate coagulation disorders through the
inhibition of HPA activity and protection of the integrity of the
glycocalyx. It may be hypothesized that the degradation of
endothelial glycocalyx may be one of the important causes of
coagulation dysfunction in sepsis.
Acknowledgements
Not applicable.
Funding
This work has received funding from the Medicine and
Health Science Technology Development Plan of Shandong Province
(grant no. 2018WS549), National Natural Science Foundation of China
(grant no. 81670078), Taishan Scholar Project of Shandong Province,
Key Clinical Specialty Project of Shandong Province, and Department
of Clinical Laboratory, Yantai Affiliated Hospital of Binzhou
Medical University.
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
XH performed the majority of the experiments and
prepared the manuscript. SH, XL and TW performed the ELISAs. HX and
JL performed the immunofluorescence analyses. GK and HH analyzed
the data. BX performed the blood coagulation tests and participated
in experimental design. DH and XW designed the experiments. WZ
analyzed and interpreted the data and participated in language
editing. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
The animal care and experimental procedures were
conducted in accordance with the Guide for the Care and Use of
Laboratory Animals, and the protocol was approved by the
Institutional Animal Care and Use Committee of Binzhou Medical
University Hospital.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Rhodes A, Evans LE, Alhazzani W, Levy MM,
Antonelli M, Ferrer R, Kumar A, Sevransky JE, Sprung CL, Nunnally
ME, et al: Surviving sepsis campaign: International guidelines for
management of sepsis and septic shock: 2016. Intensive Care Med.
43:304–377. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Berthelsen LO, Kristensen AT and Tranholm
M: Animal models of DIC and their relevance to human DIC: A
systematic review. Thromb Res. 128:103–116. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Schouten M, Wiersinga WJ, Levi M and van
der Poll T: Inflammation, endothelium, and coagulation in sepsis. J
Leukoc Biol. 83:536–545. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Levi M and van der Poll T: Coagulation and
sepsis. Thromb Res. 149:38–44. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Simmons J and Pittet JF: The coagulopathy
of acute sepsis. Curr Opin Anaesthesiol. 28:227–236. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Liu XL, Wang XZ, Liu XX, Hao D, Jaladat Y,
Lu F, Sun T and Lv CJ: Low-dose heparin as treatment for early
disseminated intravascular coagulation during sepsis: A prospective
clinical study. Exp Ther Med. 7:604–608. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Rehberg S, Yamamoto Y, Sousse LE, Jonkam
C, Zhu Y, Traber LD, Cox RA, Prough DS, Traber DL and Enkhbaatar P:
Antithrombin attenuates vascular leakage via inhibiting neutrophil
activation in acute lung injury. Crit Care Med. 41:e439–e446. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Tipoe TL, Wu WK, Chung L, Gong M, Dong M,
Liu T, Roever L, Ho J, Wong MC, Chan MT, et al: Plasminogen
activator inhibitor 1 for predicting sepsis severity and mortality
outcomes: A systematic review and meta-analysis. Front Immunol.
9:12182018. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ince C, Mayeux PR, Nguyen T, Gomez H,
Kellum JA, Ospina-Tascón GA, Hernandez G, Murray P and De Backer D;
ADQI XIV Workgroup, : The endothelium in sepsis. Shock. 45:259–270.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Martin L, Koczera P, Zechendorf E and
Schuerholz T: The endothelial glycocalyx: New diagnostic and
therapeutic approaches in sepsis. Biomed Res Int. 2016:37582782016.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Uchimido R, Schmidt EP and Shapiro NI: The
glycocalyx: A novel diagnostic and therapeutic target in sepsis.
Crit Care. 23:162019. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Sieve I, Münster-Kühnel AK and
Hilfiker-Kleiner D: Regulation and function of endothelial
glycocalyx layer in vascular diseases. Vascul Pharmacol. 100:26–33.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Nadir Y, Brenner B, Zetser A, Ilan N,
Shafat I, Zcharia E, Goldshmidt O and Vlodavsky I: Heparanase
induces tissue factor expression in vascular endothelial and cancer
cells. J Thromb Haemost. 4:2443–2451. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Schmidt EP, Yang Y, Janssen WJ, Gandjeva
A, Perez MJ, Barthel L, Zemans RL, Bowman JC, Koyanagi DE, Yunt ZX,
et al: The pulmonary endothelial glycocalyx regulates neutrophil
adhesion and lung injury during experimental sepsis. Nat Med.
18:1217–1223. 2012. View
Article : Google Scholar : PubMed/NCBI
|
|
15
|
Li X and Ma X: The role of heparin in
sepsis: Much more than just an anticoagulant. Br J Haematol.
179:389–398. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wang C, Chi C, Guo L, Wang X, Guo L, Sun
J, Sun B, Liu S, Chang X and Li E: Heparin therapy reduces 28-day
mortality in adult severe sepsis patients: A systematic review and
meta-analysis. Crit Care. 18:5632014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wildhagen KC, García de Frutos P,
Reutelingsperger CP, Schrijver R, Aresté C, Ortega-Gómez A, Deckers
NM, Hemker HC, Soehnlein O and Nicolaes GA: Nonanticoagulant
heparin prevents histone-mediated cytotoxicity in vitro and
improves survival in sepsis. Blood. 123:1098–1101. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Fodor RŞ, Georgescu AM, Cioc AD,
Grigorescu BL, Cotoi OS, Fodor P, Copotoiu SM and Azamfirei L:
Time- and dose-dependent severity of lung injury in a rat model of
sepsis. Rom J Morphol Embryol. 56:1329–1337. 2015.PubMed/NCBI
|
|
19
|
Li X, Li Z, Zheng Z, Liu Y and Ma X:
Unfractionated heparin ameliorates lipopolysaccharide-induced lung
inflammation by downregulating nuclear factor-κB signaling pathway.
Inflammation Dec. 36:1201–1208. 2013. View Article : Google Scholar
|
|
20
|
Taylor FB Jr, Toh CH, Hoots WK, Wada H and
Levi M; Scientific Subcommittee on Disseminated Intravascular
Coagulation (DIC) of the International Society on Thrombosis and
Haemostasis (ISTH), : Towards definition, clinical and laboratory
criteria, and a scoring system for disseminated intravascular
coagulation. Thromb Haemost. 86:1327–1330. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Aeffner F, Bolon B and Davis IC: Mouse
models of acute respiratory distress syndrome: A review of
analytical approaches, pathologic features, and common
measurements. Toxicol Pathol. 43:1074–1092. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhao D, Ding R, Liu Y, Yin X, Zhang Z and
Ma X: Unfractionated heparin protects the protein C system against
lipopolysaccharide-induced damage in vivo and in
vitro. Exp Ther Med. 14:5515–5522. 2017.PubMed/NCBI
|
|
23
|
Rizzo AN and Dudek SM: Endothelial
glycocalyx repair: Building a wall to protect the lung during
sepsis. Am J Respir Cell Mol Biol. 56:687–688. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Huang X, Kong G, Li Y, Zhu W, Xu H, Zhang
X, Li J, Wang L, Zhang Z, Wu Y, et al: Decitabine and 5-azacitidine
both alleviate LPS induced ARDS through
anti-inflammatory/antioxidant activity and protection of glycocalyx
and inhibition of MAPK pathways in mice. Biomed Pharmacother.
84:447–453. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Confalonieri M, Salton F and Fabiano F:
Acute respiratory distress syndrome. Eur Respir Rev. 26:2017.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Chen S, He Y, Hu Z, Lu S, Yin X, Ma X, Lv
C and Jin G: Heparanase mediates intestinal inflammation and injury
in a mouse model of sepsis. J Histochem Cytochem. 65:241–249. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Afratis NA, Nikitovic D, Multhaupt HA,
Theocharis AD, Couchman JR and Karamanos NK: Syndecans-key
regulators of cell signaling and biological functions. FEBS J.
284:27–41. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Steppan J, Hofer S, Funke B, Brenner T,
Henrich M, Martin E, Weitz J, Hofmann U and Weigand MA: Sepsis and
major abdominal surgery lead to flaking of the endothelial
glycocalix. J Surg Res. 165:136–141. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Tsao CM, Ho ST and Wu CC: Coagulation
abnormalities in sepsis. Acta Anaesthesiol Taiwan. 53:16–22. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kakkar VV, Bentley PG, Scully MF,
MacGregor IR, Jones NA and Webb PJ: Antithrombin III and heparin.
Lancet. 1:103–104. 1980. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Onishi A, St Ange K, Dordick JS and
Linhardt RJ: Heparin and anticoagulation. Front Biosci (Landmark
Ed). 21:1372–1392. 2016. View
Article : Google Scholar : PubMed/NCBI
|
|
32
|
Shworak NW, Kobayashi T, de Agostini A and
Smits NC: Anticoagulant heparan sulfate to not clot-or not? Prog
Mol Biol Transl Sci. 93:153–178. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Vink H, Constantinescu AA and Spaan JA:
Oxidized lipoproteins degrade the endothelial surface layer:
Implications for platelet-endothelial cell adhesion. Circulation.
101:1500–1502. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Chappell D, Brettner F, Doerfler N, Jacob
M, Rehm M, Bruegger D, Conzen P, Jacob B and Becker BF: Protection
of glycocalyx decreases platelet adhesion after
ischaemia/reperfusion: An animal study. Eur J Anaesthesiol.
31:474–481. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Ikeda M, Matsumoto H, Ogura H, Hirose T,
Shimizu K, Yamamoto K, Maruyama I and Shimazu T: Circulating
syndecan-1 predicts the development of disseminated intravascular
coagulation in patients with sepsis. J Crit Care. 43:48–53. 2018.
View Article : Google Scholar : PubMed/NCBI
|