Introduction
Gastric cancer is currently the third highest cause
of cancer mortality among all malignancies worldwide (1). In recent years, the overall incidence
and mortality of gastric cancer has been decreasing due to control
of risk factors and the development of gastric cancer screening
(2). However, the burden of gastric
cancer remains critical due to the large population base and an
increasingly aging population (3).
In China, the incidence of gastric cancer and associated deaths
accounts for ~50% of all gastric cancer cases in the world
(4). This may be due to the higher
proportion of advanced gastric cancer cases upon diagnosis.
Therefore, investigations into the pathogenesis of gastric cancer
may reveal biomarkers that can be used for early diagnosis and
treatment.
The Wnt signaling pathway, named after the wingless
and Int-1 genes (5), performs a
variety of different functions, including wide involvement in
embryonic development, tissue homeostasis and cell carcinogenesis
(6). It is typically divided into
the canonical and non-canonical signaling pathways, of which the
canonical pathway (Wnt/β-catenin) is the more well-known pathway
and is highly conserved throughout the evolutionary process
(7). Previous studies have
demonstrated that the Wnt/β-catenin signaling pathway serves a key
role in regulating cell proliferation, maintenance of stem cell
characteristics and homeostasis of normal gastric mucosa (8–10). In
particular, it has been reported that activation of the
Wnt/β-catenin signaling cascade is found in >30% of gastric
cancer cases (11,12). Indeed, evidence is accumulating
showing that Wnt3a, one member of the Wnt family, has the ability
to either promote or inhibit cancer progression via the canonical
Wnt/β-catenin signaling pathway, depending on the type of cancer
(13).
β-catenin is a multifunctional protein, which was
initially found to bind to E-cadherin-binding protein and
participate in cell adhesion (14).
E-cadherin is not only a key member of Wnt signaling pathway, but
also a critical epithelial marker of epithelial-mesenchymal
transition (EMT) (15). EMT refers
to the process whereby epithelial cells transform into mesenchymal
phenotype cells, resulting in enhanced cell motility and
invasiveness. EMT is one of the key initiation events for tumor
metastasis and has a crucial role in the invasion and metastasis of
cancers, including gastric cancer (16–18).
Therefore, increasing research has considered EMT as an effective
therapeutic target in cancer treatment (19). Research over the past decade has
mainly focused on searching for novel anticancer agents and
reassessing known drugs that have been used in other diseases to
uncover any potential anti-tumor effects (20). One of those, pizotifen, also known as
4-(1-methyl-4-piperidylidine)-9,10-dihydro-4H-benzo-(4,5)-cyclohepta(1,2)-thiophene,
is a potent serotonin or 5-hydroxytryptamine (HT)2
receptor antagonist (21). Pizotifen
is primarily indicated for the prevention of vascular headaches
(22). It has been previously
reported that serotonin 5-HT2A receptor antagonists,
such as ritanserin, can inhibit cell viability and promote cell
apoptosis in colorectal cancer cells (23). However, the effect of pizotifen on
cancer growth and metastasis remains unknown.
In the present study, it was found that pizotifen
inhibited the viability, migration and invasion of gastric cancer
whilst inducing cell apoptosis. The
Wnt/β-catenin-epithelial-mesenchymal transition (EMT) signaling
pathway may be involved in this process. These results suggested
that pizotifen could potentially be a new therapeutic agent for the
treatment of gastric cancer and warrants further assessment in
future clinical studies.
Materials and methods
Reagents and antibodies
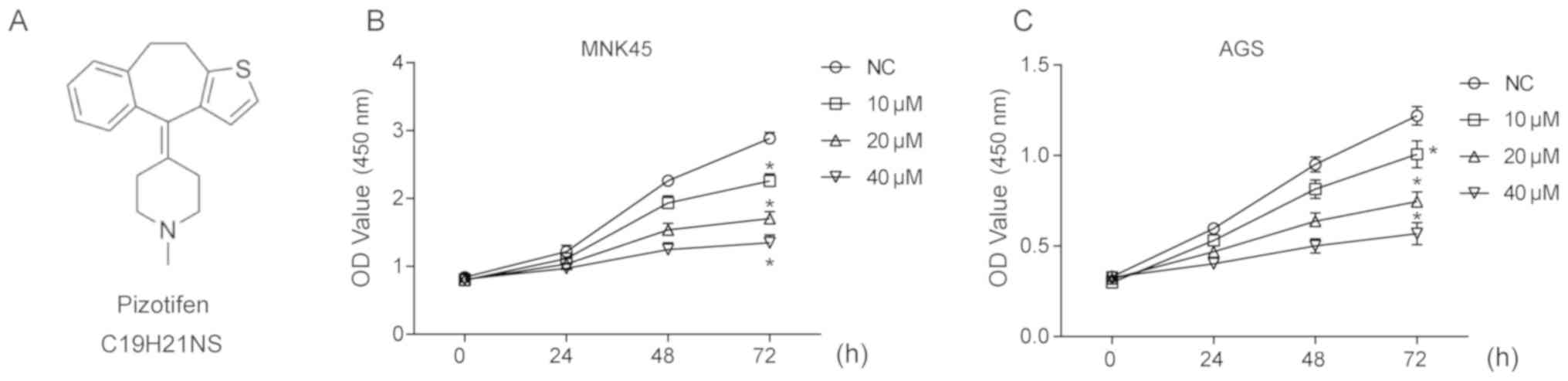
Pizotifen was purchased from MedChemExpress and its
molecular structure is shown in Fig.
1A. The primary antibodies against Wnt3a (cat. no. ab28472;
dilution, 1:1,000), active-caspase-3 (cat. no. ab2302; dilution,
1:1,000) were purchased from Abcam, and the primary antibodies
against Cyclin D-1 (cat. no. 60186-1-lg; dilution, 1:1,000), Bax
(cat. no. 50599-2-lg; dilution, 1:1,000), Bcl-2 (cat. no.
12789-1-AP; dilution, 1:1,000) and β-tubulin (cat. no. 10094-1-AP;
dilution, 1:1,000) were purchased from ProteinTech Group, Inc.
Primary antibodies against β-catenin (cat. no. 9562; dilution,
1:1,000), E-cadherin (cat. no. 14472; dilution, 1:1,000) and
N-cadherin (cat. no. 4061; dilution, 1:1,000) were purchased from
Cell Signaling Technology, Inc. Both secondary antibodies
(horseradish peroxidase conjugated anti-rabbit, cat. no.
SA00001-15; anti-mouse, cat. no. SA00001-1; both dilutions,
1:5,000) were purchased from ProteinTech Group, Inc. BML-284, a
selective Wnt activator, was purchased from MedChemExpress. Unless
specifically indicated, all other reagents were obtained from
Sigma-Aldrich (Merck KGaA).
Cell culture and treatment
Human gastric cancer cell lines MNK45 and AGS were
obtained from the Type Culture Collection of the Chinese Academy of
Sciences. The cell lines were cultured in DMEM containing 10% FBS
(Gibco; Thermo Fisher Scientific, Inc.), penicillin (100 U/ml), and
streptomycin (100 mg/ml), and were maintained in a humidified
atmosphere (95%) at 37°C under 5% CO2. Cells in the
logarithmic growth phase were treated with different concentrations
of pizotifen (10, 20 and 40 µM) or BML-284 (10 µM) alone or
co-treated with pizotifen (20 µM) and BML-284 (10 µM). The cells
were treated for varying time periods, including 0, 24, 48 and 72
h, 0.1% DMSO was used as negative control.
Cell viability assay
The Cell Counting Kit-8 (CCK-8) assay was performed
according to the manufacturer's protocol (Beijing Solarbio Science
& Technology Co., Ltd.). Cells were seeded into 96-well plates
at a density of 1×103 cells/well and cultured overnight
to allow for adherence. Cultures were subsequently treated with
different concentrations of pizotifen (10, 20 and 40 µM). DMSO
(0.1%) was used as the negative control. The cells were then
cultured for 3 days and measurements were obtained every 24 h. When
the cells were measured, CCK-8 solution (10 µl/well) was added to
each well and incubated for an additional 1.5 h at 37°C. The final
optical density was measured at a wavelength of 450 nm to estimate
cell viability in the different treatment groups. Each experiment
was performed in triplicate.
Wound healing assay
Cells were first seeded into six-well plates at a
density of 5×105 cells/well and incubated overnight at
37°C. The following day, when cells had reached ~95–100%confluency,
wounds were generated using pipette tips. After being scratched
with the pipettes, the cells were cultured in serum-free medium
supplemented with 10 or 20 µM pizotifen or DMSO for 24 h. The
wounds were photographed using a light microscope (magnification,
×40) and analyzed using ImageJ software v1.8.0 (National Institutes
of Health). The percentage wound closure was calculated using the
mathematical equation: Percentage wound closure (%) =
(A0-At)/A0 × 100%, where
A0 is the initial wound area, and At is the
wound area at 0, 24 h post-treatment.
Western blot analysis
Total protein was extracted from the cell cultures
according to instructions from radioimmunoprecipitation assay Lysis
Buffer (CWBIO Tech). Protein concentration was quantified using an
Enhanced Bicinchoninic Acid Protein Assay kit. A total of 20 µg
protein from each sample were separated by 10% SDS-PAGE and then
transferred to PVDF membranes (Bio-Rad Laboratories, Inc.). The
membranes were incubated in blocking buffer [5% non-fat milk
dissolved in TBS with Tween-20 (TBST)] at room temperature for 1-h
and then probed at 4°C overnight with primary antibodies (dilution,
1:1,000). The membranes were subsequently rinsed with TBST and
incubated with horseradish peroxidase-conjugated secondary
antibodies (1:5,000) for 1-h at room temperature. The blots were
visualized using the enhanced chemiluminescence detection kit
(Beyotime Institute of Biotechnology) and analyzed using the
Quantity One 1-D software v4.6.7 (Bio-Rad Laboratories, Inc.). Each
experiment was repeated at least three times.
Apoptosis assay
Cells (5×105) pretreated with 20 µM
pizotifen or 0.1% DMSO were double-stained with fluorescein
isothiocyanate-conjugated Annexin V (25 mg/ml) and propidium iodide
(PI; 50 mg/ml) at 4°C for 15 min according to the manufacturer's
protocol (4A Biotech Co., Ltd.), and then washed and analyzed using
a BD FACSCalibur flow cytometry system (BD Biosciences). The
obtained data was analyzed using FlowJo software v7.6.1 (FlowJo
LLC).
Transwell assays
For the Transwell migration assay, 5×103
cells pretreated with pizotifen or BML-284 were seeded into the
upper sections of the non-coated 96-well inserts (pore size, 8 µm;
Corning, Inc.). For the invasion assay, Matrigel (BD Biosciences)
was thawed at 4°C and diluted in serum-free medium at a ratio of
1:6, then added to the upper chamber of the Transwell chamber and
maintained at 37°C for 4–6 h. A total of 1×105 cells
were seeded into the top chamber of 24-well inserts with 8 µm pore
size and Matrigel-coated membranes. In both assays, cells were
diluted in medium without FBS and seeded into the top chamber,
whereas medium containing 10% FBS were introduced into the lower
chamber in order to establish a chemoattractant gradient. Cells
were incubated for 24 h, fixed with 4% paraformaldehyde at room
temperature for 30 min, and stained with 0.1% crystal violet
solution (Sigma-Aldrich; Merck KGaA) at room temperature for 20
min. Then, 5 microscopic fields were used to count and photograph
the stained cells under a light microscope (magnification,
×100).
Statistical analysis
Experimental data are presented as the mean ± SD.
All statistical analysis was performed using SPSS 13.5 (SPSS,
Inc.). Student's t-test and one-way ANOVA followed by Tukey's
posthoc analysis were performed. P<0.05 was considered to
indicate a statistically significant difference.
Results
Pizotifen inhibits the viability of
gastric cancer MNK45 and AGS cells
A CCK8 assay was carried out to determine the effect
of pizotifen on the viability of gastric cancer cell lines MNK45
and AGS. As shown in Fig. 1B,
pizotifen treatment of 48h or 72 h significantly inhibited the
viability of MNK45 cells in a dose-dependent manner (P<0.05).
Similarly, pizotifen also inhibited the viability of AGS cells
(Fig. 1C).
Pizotifen inhibits MNK45 and AGS cells
migration and invasion
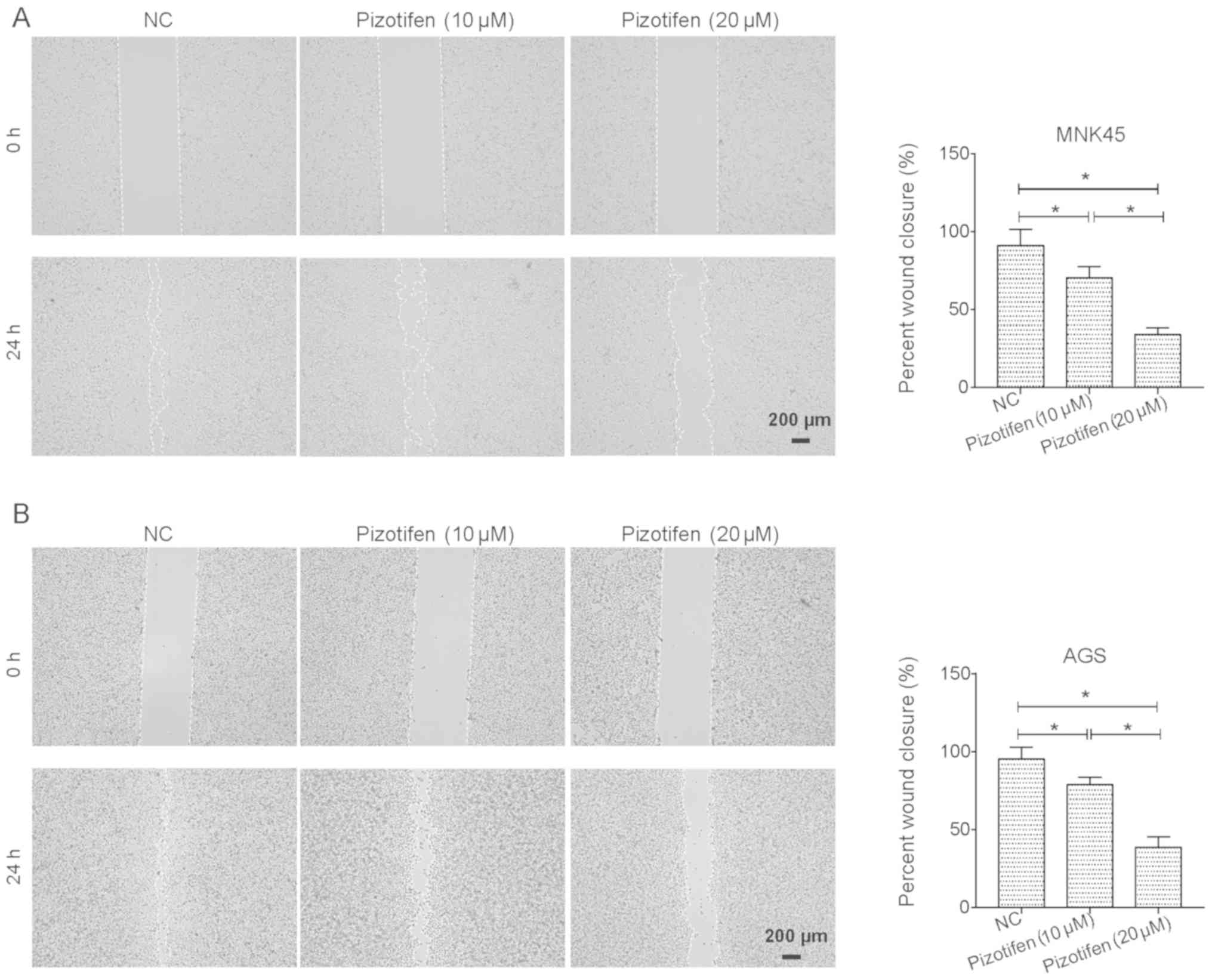
Wound-healing and Transwell assays were performed to
investigate whether pizotifen affects the migratory and invasive
properties of gastric cancer cells. Since 10 and 20 µM pizotifen
both caused appropriate inhibitory effect on MNK45 and AGS cells,
whilst 40 µM pizotifen caused increased cell death, 10 and 20 µM of
pizotifen were used in the rest experiments. As shown in Fig. 2A, 10 and 20 µM pizotifen both led to
a significant decrease in the migratory abilities of MNK45 cells in
a dose-dependent manner (P<0.05). A similar inhibitory effect of
pizotifen was observed in AGS cells (P<0.05; Fig. 2B). Moreover, the results of the
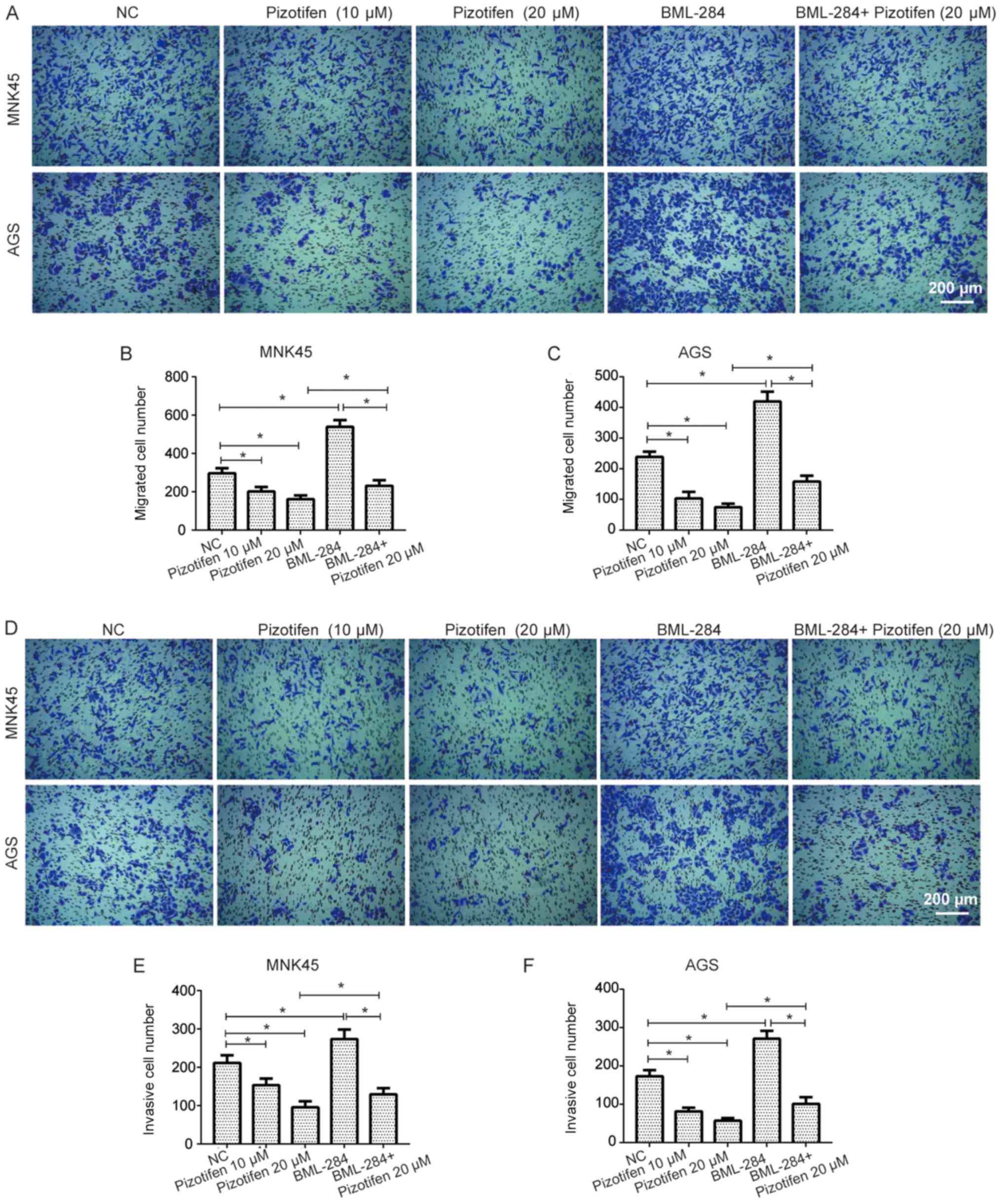
Transwell assay further suggested that the migratory abilities of
MNK45 and AGS cells were significantly inhibited following
treatment with pizotifen (P<0.05; Fig. 3A-C). In addition, the invasive
abilities of MNK45 and AGS cells treated with pizotifen were
significantly reduced (P<0.05; Fig.
3D-F). Furthermore, the data showed that BML-284, a selective
Wnt signaling activator that has been revealed to induce β-catenin
in vitro (24), significantly
increased the migration and invasion of both MNK45 and AGS cells
(both P<0.05; Fig. 3), and
partially restored the migratory and invasive abilities of MNK45
and AGS cells inhibited by pizotifen (P<0.05; Fig. 3).
Pizotifen induces the
mitochondrial-mediated apoptosis of MNK45 and AGS cells
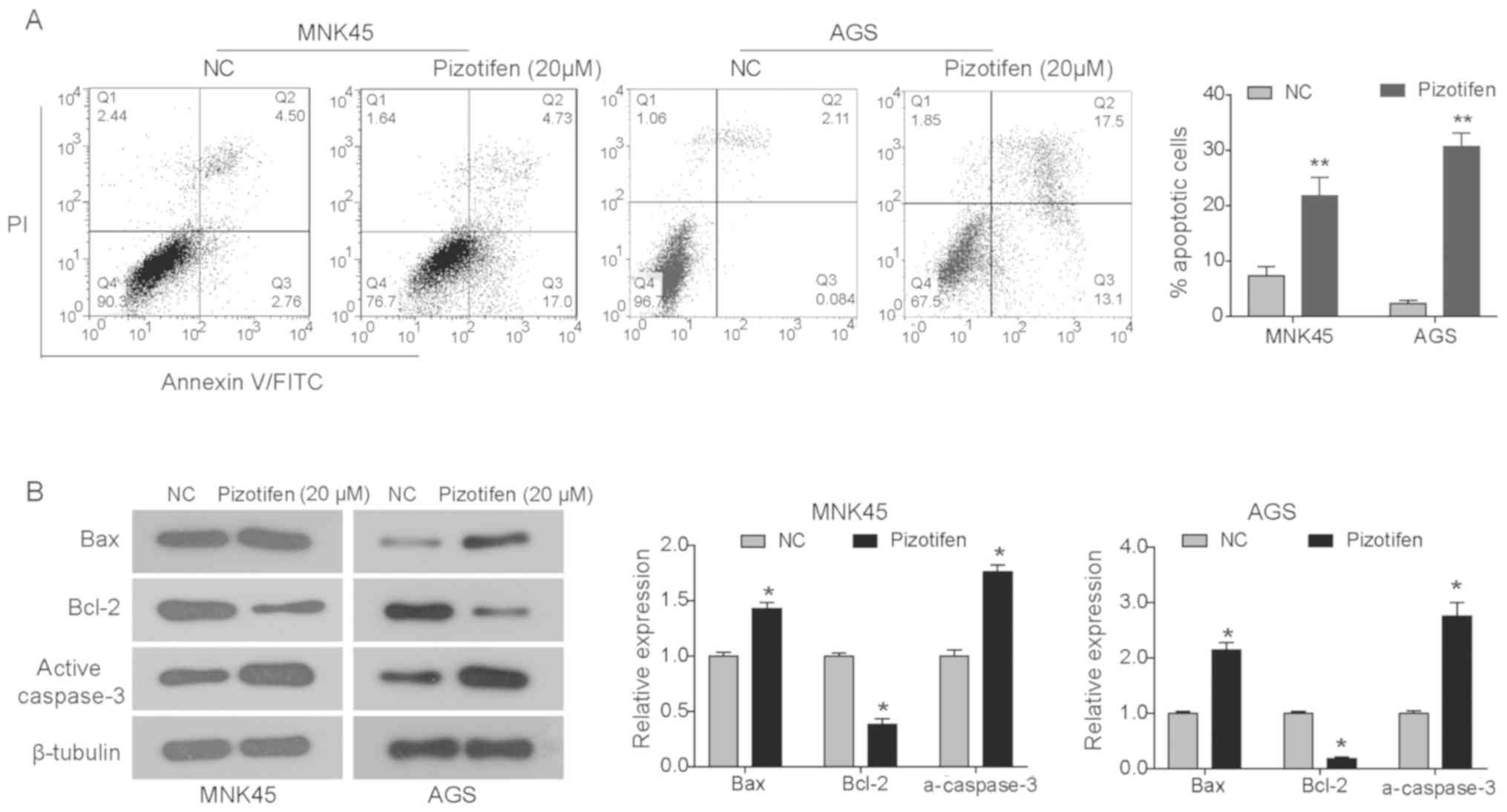
Annexin V-FITC/PI assay was performed to investigate
whether pizotifen inhibited the growth of gastric cancer cells by
triggering apoptosis. As shown in Fig.
4A, pizotifen (20 µM) significantly enhanced cell apoptosis in
both MNK45 and AGS cells (both P<0.01) compared with the NC
group (Fig. 4A). To further
determine the mechanism of pizotifen-induced apoptosis, western
blot analysis was performed to detect the expression of proteins
associated with the mitochondrial apoptotic pathway in MNK45 and
AGS cells treated with pizotifen for 48 h (Fig. 4B). The data demonstrated that the
expression of the anti-apoptotic protein Bcl-2 was significantly
reduced in pizotifen-treated cells, whereas the expression of
pro-apoptotic proteins Bax and active caspase-3 were significantly
increased (P<0.05; Fig. 4B).
These data indicated that pizotifen may induce the apoptosis of
gastric cancer cells via the intrinsic apoptotic pathway,
specifically the Bax/Bcl-2 and caspase cascade.
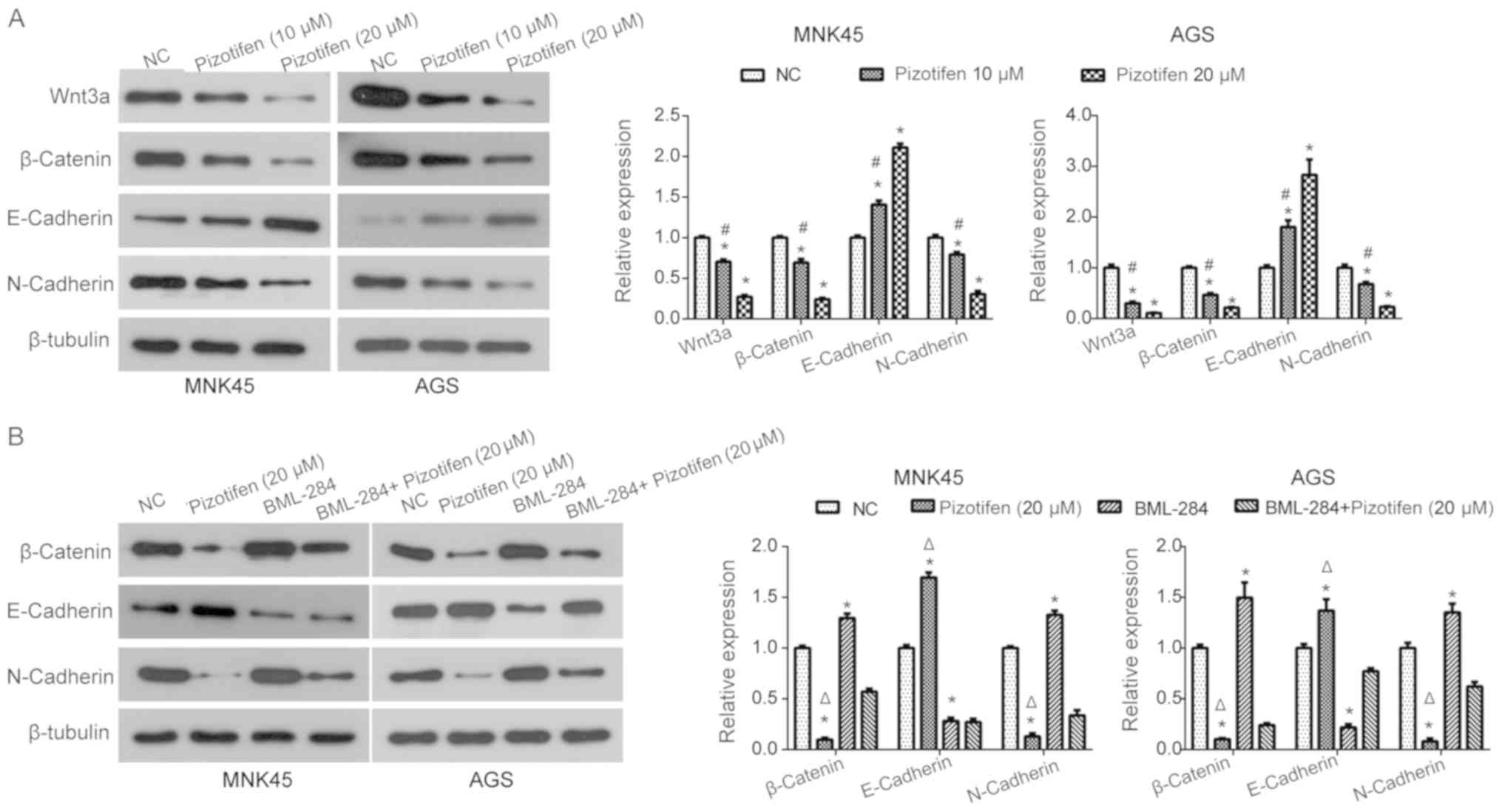
Pizotifen downregulates the
Wnt/β-catenin-EMT signaling pathway in MNK45 and AGS cells
The ability of pizotifen to inhibit the
Wnt/β-catenin signaling pathway in MNK and AGS cells was
investigated using western blot analysis. Pizotifen treatment
significantly reduced the expression of Wnt3a and β-catenin in a
dose-dependent manner in MNK45 and AGS cells (P<0.05; Fig. 5A). In addition, the expression of the
epithelial marker E-cadherin was significantly upregulated by
pizotifen treatment in MNK45 and AGS cells, while the expression of
the mesenchymal marker N-cadherin was significantly downregulated
(P<0.05; Fig 5A), suggesting that
pizotifen regulated the expression of EMT markers proteins in a
dose-dependent manner. This suggested that pizotifen can suppress
the Wnt/β-catenin-EMT signaling pathway in MNK45 and AGS cells.
Wnt/β-catenin-EMT signaling pathway
may play a role in the anti-invasive effect of pizotifen
To further investigate the mechanism of action of
pizotifen in the inhibition of gastric cancer cell invasion,
BML-284 was used. It was found that the expression of β-catenin was
significantly upregulated in MNK45 and AGS cells following BML-284
treatment compared with the NC group (Fig. 5B). In the presence of pizotifen,
BML-284 treatment also partially restored β-catenin expression
compared with the pizotifen-only group (Fig. 5B). In addition, BML-284 treatment
partially reversed the effects induced by pizotifen on E-cadherin
and N-cadherin expression in MNK45 and AGS cells compared with the
pizotifen-treated group (Fig. 5B).
Taken together, the results of the present study indicated that the
pizotifen-induced inhibitory effects on gastric cancer cell
migration and invasion may be due to the inhibition of the
Wnt/β-catenin-EMT signaling pathway.
Discussion
Gastric cancer is one of the most serious malignant
tumors; its mortality rate is lower only than that of lung cancer
and liver cancer worldwide (1).
Therefore, it has become a priority to explore the mechanism of the
occurrence and development of gastric cancer in order to develop
potential novel anti-cancer agents. In the present study, it was
found that pizotifen could inhibit the viability, migration and
invasion of MNK45 and AGS cells, as well as inducing apoptosis.
Pizotifen is a serotonin antagonist that is primarily indicated for
the prevention of recurrent migraine headaches. A number of studies
have reported that serotonin can promote the growth of several
types of cancer, including breast (25), prostate (26), colorectal (27), hepatocellular (28) and small-cell lung cancer (29). In addition, it has been reported that
serotonin is involved in cancer cell migration, metastasis and
angiogenesis (30). In some cases,
antagonists of serotonin receptors have been successfully used to
inhibit cancer cell growth (31,32). The
present study is the first to report the anti-tumor properties of
pizotifen against gastric cancer cells. However, the effects of
pizotifen on cell morphology and non-cancerous cells remain
unclear, which is a limitation of the present study. In future
studies, any potential anti-tumor properties of pizotifen should be
further investigated in vivo and in clinical studies.
Aberrant activation of the Wnt signaling pathway is
associated with the occurrence and development of tumors, and it
has become the focus of studies seeking to therapeutic targets
(33,34). Therefore, drug development has been
mainly focused on inhibiting the Wnt signaling cascade, and some of
these drugs have since entered clinical trials (35,36).
Genome sequencing studies have revealed that the mutation of many
components of the Wnt pathway are inherited in patients with
gastric cancer, such as AXIN1, AXIN2 and β-catenin (37), suggesting that targeting Wnt has
potential in the treatment of gastric cancer. These data were
verified in a mouse model of gastric cancer (38–40).
This suggests that Wnt signaling is likely to be a driving factor
in the occurrence of gastric cancer.
It was demonstrated in the present study that
pizotifen could significantly downregulate the expression of Wnt3a
and β-catenin in MNK45 and AGS cells. β-catenin is a
multifunctional protein, which was initially found to bind to
E-cadherin-binding protein and participate in cell adhesion
(14). E-cadherin has been reported
to be a key member of the Wnt signaling pathway, with
transcriptional regulatory activity (15). It is hypothesized that the disruption
of E-cadherin/β-catenin complexes is involved in the malignant
transformation of epithelial tumors (41). E-cadherin and N-cadherin are key
markers for the EMT process, both of which serve essential roles in
tumor metastasis. It was previously demonstrated that the
downregulation of E-cadherin in gastric cancer could reduce the
expression of the cadherin/catenin complex to increase cancer cell
invasiveness (42). In the present
study, it was found that pizotifen treatment in gastric cancer cell
lines upregulated the expression of E-cadherin and downregulated
the expression of N-cadherin and β-catenin, an effect that was
reversed by concomitant treatment with BML-284, a selective Wnt
signaling activator. It was also observed in the present study that
MNK45 and AGS cell migration and invasion, which were inhibited by
pizotifen, were restored by simultaneous BML-284 treatment. Taken
together, these observations suggested that inhibition of the
Wnt/β-catenin-EMT signaling pathway may be responsible for the
anti-invasion properties of pizotifen.
In summary, the present study showed that pizotifen
could inhibit the growth, migration and invasion of gastric cancer
cells and induce cell apoptosis. The anti-cancer activity of
pizotifen may be achieved by blocking the Wnt/β-catenin-EMT
signaling pathway. Based on these findings, pizotifen could
potentially be used in a novel therapeutic approach for the
treatment of gastric cancer, a notion which warrants further
clinical evaluation. However, the precise mechanism of action of
pizotifen will be explored further in a future study.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
YJ, WW and XW designed the study. YJ and WW
performed the experiments, and XW and JS analyzed the data. All
authors interpreted the results, and produced and approved the
final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Carlomagno N, Santangelo ML, Amato B,
Calogero A, Saracco M, Cremone C, Miranda A, Dodaro C and Renda A:
Total colectomy for cancer: Analysis of factors linked to patients'
age. Int J Surg. 12 (Suppl 2):S135–S139. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Strand MS, Lockhart AC and Fields R:
Genetics of gastric cancer. Surg Clin North Am. 97:345–370. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Hamashima C: Current issues and future
perspectives of gastric cancer screening. World J Gastroenterol.
20:13767–13774. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Chen W, Zheng R, Baade PD, Zhang S, Zeng
H, Bray F, Jemal A, Yu XQ and He J: Cancer statistics in China,
2015. CA Cancer J Clin. 66:115–132. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Baker NE: Transcription of the
segment-polarity gene wingless in the imaginal discs of drosophila,
and the phenotype of a pupal-lethal wg mutation. Development.
102:489–497. 1988.PubMed/NCBI
|
|
6
|
Croce JC and McClay DR: Evolution of the
Wnt pathways. Methods Mol Biol. 469:3–18. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Clevers H and Nusse R: Wnt/β-catenin
signaling and disease. Cell. 149:1192–1205. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Rapp J, Jaromi L, Kvell K, Miskei G and
Pongracz JE: WNT signaling - lung cancer is no exception. Respir
Res. 18:1672017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Byun T, Karimi M, Marsh JL, Milovanovic T,
Lin F and Holcombe RF: Expression of secreted Wnt antagonists in
gastrointestinal tissues: Potential role in stem cell homeostasis.
J Clin Pathol. 58:515–519. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Barker N, Huch M, Kujala P, van de
Wetering M, Snippert HJ, van Es JH, Sato T, Stange DE, Begthel H,
van den Born M, et al: Lgr5(+ve) stem cells drive self-renewal in
the stomach and build long-lived gastric units in vitro. Cell Stem
Cell. 6:25–36. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wang K, Yuen ST, Xu J, Lee SP, Yan HH, Shi
ST, Siu HC, Deng S, Chu KM, Law S, et al: Whole-genome sequencing
and comprehensive molecular profiling identify new driver mutations
in gastric cancer. Nat Genet. 46:573–582. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Cristescu R, Lee J, Nebozhyn M, Kim KM,
Ting JC, Wong SS, Liu J, Yue YG, Wang J, Yu K, et al: Molecular
analysis of gastric cancer identifies subtypes associated with
distinct clinical outcomes. Nat Med. 21:449–456. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Nagaoka T, Karasawa H, Turbyville T,
Rangel MC, Castro NP, Gonzales M, Baker A, Seno M, Lockett S, Greer
YE, et al: Cripto-1 enhances the canonical Wnt/β-catenin signaling
pathway by binding to LRP5 and LRP6 co-receptors. Cell Signal.
25:178–189. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Garinis GA, Spanakis NE, Menounos PG,
Manolis EN and Peros G: Transcriptional impairment of
beta-catenin/E-cadherin complex is not associated with beta-catenin
mutations in colorectal carcinomas. Br J Cancer. 88:206–209. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kikuchi A, Yamamoto H, Sato A and
Matsumoto S: New insights into the mechanism of Wnt signaling
pathway activation. Int Rev Cell Mol Biol. 291:21–71. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Micalizzi DS, Farabaugh SM and Ford HL:
Epithelial-mesenchymal transition in cancer: Parallels between
normal development and tumor progression. J Mammary Gland Biol
Neoplasia. 15:117–134. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Peng Z, Wang CX, Fang EH, Wang GB and Tong
Q: Role of epithelial-mesenchymal transition in gastric cancer
initiation and progression. World J Gastroenterol. 20:5403–5410.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yue H, Tang B, Zhao Y, Niu Y, Yin P, Yang
W, Zhang Z and Yu P: MIR-519d suppresses the gastric cancer
epithelial-mesenchymal transition via Twist1 and inhibits
Wnt/β-catenin signaling pathway. Am J Transl Res. 9:3654–3664.
2017.PubMed/NCBI
|
|
19
|
Cho ES, Kang HE, Kim NH and Yook JI:
Therapeutic implications of cancer epithelial-mesenchymal
transition (EMT). Arch Pharm Res. 42:14–24. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Fuchs R, Schwach G, Stracke A,
Meier-Allard N, Absenger M, Ingolic E, Haas HS, Pfragner R and
Sadjak A: The anti-hypertensive drug prazosin induces apoptosis in
the medullary thyroid carcinoma cell line TT. Anticancer Res.
35:31–38. 2015.PubMed/NCBI
|
|
21
|
Lin OA, Karim ZA, Vemana HP, Espinosa EV
and Khasawneh FT: The antidepressant 5-HT2A receptor
antagonists pizotifen and cyproheptadine inhibit serotonin-enhanced
platelet function. PLoS One. 9:e870262014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Speight TM and Avery GS:
Pizotifen(BC-105): A review of its pharmacological properties and
its therapeutic efficacy in vascular headaches. Drugs. 3:159–203.
1972. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ahmadi AA, Shadifar M, Ataee R,
Vaillancourt C, Ataee A, Oufkir T and Jafari-Sabet M: The serotonin
5-HT2A receptor antagonist ritanserin induces apoptosis
in human colorectal cancer and acts in synergy with curcumin. Int
Biol Biomed J. 1:56–65. 2015.
|
|
24
|
Liu J, Wu X, Mitchell B, Kintner C, Ding S
and Schultz PG: A small-molecule agonist of the Wnt signaling
pathway. Angew Chem Int Ed Engl. 44:1987–1990. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Cheng YY, Yu J, Wong YP, Man EP, To KF,
Jin VX, Li J, Tao Q, Sung JJ, Chan FK and Leung WK: Frequent
epigenetic inactivation of secreted frizzled-related protein 2
(SFRP2) by promoter methylation in human gastric cancer. Br J
Cancer. 97:895–901. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Hanaki H, Yamamoto H, Sakane H, Matsumoto
S, Ohdan H, Sato A and Kikuchi A: An anti-Wnt5a antibody suppresses
metastasis of gastric cancer cells in vivo by inhibiting
receptor-mediated endocytosis. Mol Cancer Ther. 11:298–307. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ataee R, Ajdary S, Zarrindast M, Rezayat M
and Hayatbakhsh MR: Anti-mitogenic and apoptotic effects of 5-HT1B
receptor antagonist on HT29 colorectal cancer cell line. J Cancer
Res Clin Oncol. 136:1461–1469. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Soll C, Riener MO, Oberkofler CE,
Hellerbrand C, Wild PJ, DeOliveira ML and Clavien PA: Expression of
serotonin receptors in human hepatocellular cancer. Clin Cancer
Res. 18:5902–5910. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Vicentini LM, Cattaneo MG and Fesce R:
Evidence for receptor subtype cross-talk in the mitogenic action of
serotonin on human small-cell lung carcinoma cells. Eur J
Pharmacol. 318:497–504. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Soll C, Jang JH, Riener MO, Moritz W, Wild
PJ, Graf R and Clavien PA: Serotonin promotes tumor growth in human
hepatocellular cancer. Hepatology. 51:1244–1254. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Dizeyi N, Hedlund P, Bjartell A, Tinzl M,
Austild-Taskén K and Abrahamsson PA: Serotonin activates MAP kinase
and PI3K/Akt signaling pathways in prostate cancer cell lines. Urol
Oncol. 29:436–445. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Siddiqui EJ, Shabbir MA, Mikhailidis DP,
Mumtaz FH and Thompson CS: The effect of serotonin and serotonin
antagonists on bladder cancer cell proliferation. BJU Int.
97:634–639. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Hirokawa Y, Yip KH, Tan CW and Burgess AW:
Colonic myofibroblast cell line stimulates colonoid formation. Am J
Physiol Gastrointest Liver Physiol. 306:G547–G556. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Byun MR, Hwang JH, Kim AR, Kim KM, Hwang
ES, Yaffe MB and Hong JH: Canonical Wnt signalling activates TAZ
through PP1A during osteogenic differentiation. Cell Death Differ.
21:854–863. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Bu G, Lu W, Liu CC, Selander K, Yoneda T,
Hall C, Keller ET and Li Y: Breast cancer-derived Dickkopf1
inhibits osteoblast differentiation and osteoprotegerin expression:
Implication for breast cancer osteolytic bone metastases. Int J
Cancer. 123:1034–1042. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
He S, Lu Y, Liu X, Huang X, Keller ET,
Qian CN and Zhang J: Wnt3a: Functions and implications in cancer.
Chin J Cancer. 34:554–562. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Pan KF, Liu WG, Zhang L, You WC and Lu YY:
Mutations in components of the Wnt signaling pathway in gastric
cancer. World J Gastroenterol. 14:1570–1574. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Kahn M: Can we safely target the WNT
pathway? Nat Rev Drug Discov. 13:513–532. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Mook RA Jr, Wang J, Ren XR, Chen M,
Spasojevic I, Barak LS, Lyerly HK and Chen W: Structure-activity
studies of Wnt/β-catenin inhibition in the niclosamide chemotype:
Identification of derivatives with improved drug exposure. Bioorg
Med Chem. 23:5829–5838. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Steinhart Z, Pavlovic Z, Chandrashekhar M,
Hart T, Wang X, Zhang X, Robitaille M, Brown KR, Jaksani S,
Overmeer R, et al: Corrigendum: Genome-wide CRISPR screens reveal a
Wnt-FZD5 signaling circuit as a druggable vulnerability of
RNF43-mutant pancreatic tumors. Nat Med. 23:13842017. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Kim H, Yoo SB, Sun P, Jin Y, Jheon S, Lee
CT and Chung JH: Alteration of the E-cadherin/β-catenin complex is
an independent poor prognostic factor in lung adenocarcinoma.
Korean J Pathol. 47:44–51. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Cheng XX, Wang ZC, Chen XY, Sun Y, Kong
QY, Liu J, Gao X, Guan HW and Li H: Frequent loss of membranous
E-cadherin in gastric cancers: A cross-talk with Wnt in determining
the fate of beta-catenin. Clin Exp Metastasis. 22:85–93. 2005.
View Article : Google Scholar : PubMed/NCBI
|