Antioxidant effects of aged garlic
extract
The intake of raw garlic or its preparations has
been shown to mitigate multiple risk factors associated with
cardiovascular diseases (1). In
clinical studies, aged garlic extract (AGE) has been shown to
improve endothelial dysfunction that is considered as an early
marker of atherosclerosis (2,3). One
possible mechanism underlying endothelial dysfunction is an
increase in reactive oxygen species (ROS) generated by an enhanced
energy metabolism or chronic inflammation. The antioxidant
properties of AGE have been proposed to play a role in preventing
endothelial dysfunction (4). AGE has
been shown to contain organosulfur compounds, polyphenols and
Maillard reaction products, such as
Nα-(1-deoxy-D-fructos-1-yl)-L-arginine. These chemicals have
radical scavenging properties in vitro; however, the
mechanisms through which these chemicals exert their antioxidant
effects in vivo remain unclear (5,6). The
activation of nuclear factor erythroid 2 like 2 (Nrf2)-dependent
gene expression has also been proposed as a mechanism for the
maintenance of vascular homeostasis via the enhancement of the
cellular defense mechanism against oxidative stresses (7). A recent study using canines
demonstrated that the administration of AGE upregulated the gene
expression levels of canine Nrf2 and phase II antioxidant enzymes
(8).
Sulfur amino acids are known to increase
resistance to oxidative stress by modulating the Nrf2/SKN-1
pathway
Garlic is rich in organosulfur compounds which are
believed to be responsible for most of its pharmacological
properties. S−1-propenylcysteine (S1PC) and
S-allylcysteine (SAC) are two sulfur amino acids
predominantly found in AGE (5). In
oxidative stress models using Caenorhabditis elegans, SAC
has been shown to increase stress resistance and reduce the
accumulation of ROS. These antioxidant effects have been shown to
require the transcription factor, SKN-1, that is the Nrf2
orthologue in mammals (9). Nrf2 is a
transcription factor that regulates key antioxidant genes and phase
II detoxification genes in mammals. Although the activation of
Nrf2-dependent gene expression has been proposed as a mechanism for
maintaining vascular homeostasis, it remains unclear as to whether
SAC and S1PC activate Nrf2 in endothelial cells.
Known synergistic effect of S1PC and
NO-donors on antioxidant gene expression
Nitric oxide (NO) is an important signaling molecule
involved in maintaining vascular homeostasis and is also one of the
activators of the Nrf2 pathway (10). Therefore, in a previous study, we
examined the effects of SAC and S1PC on the signaling mechanism of
NO in modulating the Nrf2-dependent antioxidant response in
endothelial cells (11). Neither
S1PC nor SAC, were found to independently affect the expression of
Nrf2-regulated genes, such as heme oxygenase-1 (HMOX1) and
glutamate-cysteine ligase modifier subunit (GCLM) in human
umbilical vein endothelial cells (HUVECs). However, S1PC was found
to augment the expression of HMOX1 and GCLM induced
by NO donors, such as NOR3. In that study, SAC did not exert such
synergistic effects with NO donors (11). In that previous study, we also
confirmed the synergistic effect of S1PC with another NO donor,
S-nitrosoglutathione (GSNO), on the expression of
HMOX1 in human aortic endothelial cells (HAECs) (11).
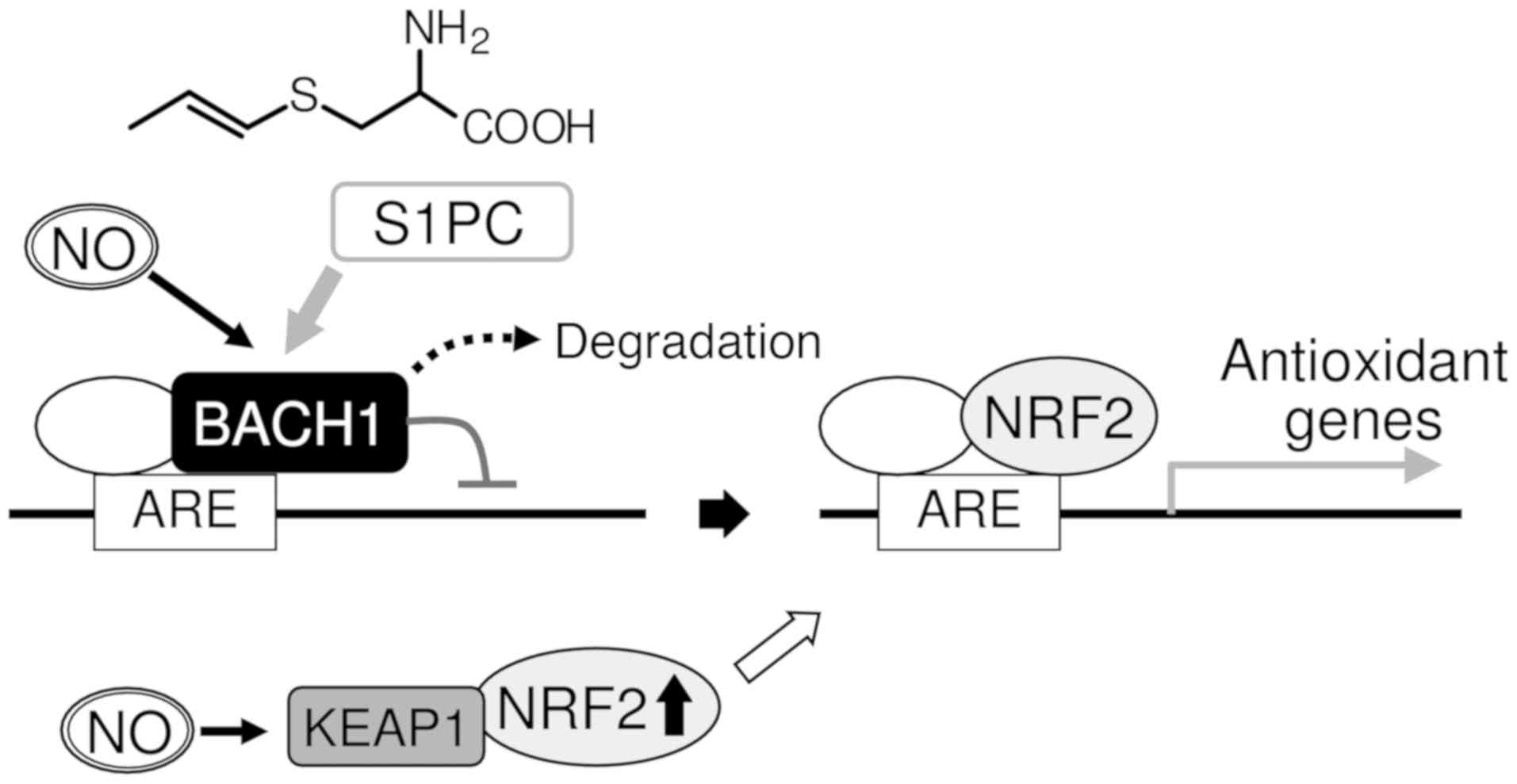
Evidence of the induction of BACH1
downregulation by S1PC in a NO-dependent manner
Under basal conditions, Nrf2 binds to Kelch-like
ECH-associated protein 1 (Keap1) resulting in proteasomal
degradation of Nrf2 in the cytoplasm. In the presence of oxidative
stress, Nrf2 is released from the Keap1-dependent complex and
accumulates in the nucleus. Nrf2 binds to antioxidant response
elements (AREs) that are cis-elements essential for the expression
of various antioxidant genes, including HMOX1 and
GCLM, whereas the BTB domain and CNC homolog 1 (BACH1)
interacts with the AREs of the corresponding genes to prevent Nrf2
binding, thus inhibiting gene expression (12). In a previous study, in order to
examine the synergistic effects of S1PC and NO in the context of
antioxidant gene expression, we analyzed the protein expression of
NRF2 and BACH1. A significant accumulation of NRF2 was not observed
in HUVECs treated with S1PC alone. NOR3 was found to induce NRF2
accumulation; however, cells co-treated with NOR3 and S1PC did not
exhibit any significant differences in NRF2 protein levels compared
to cells treated only with NOR3 (11). NOR3 was also found to induce a
reduction in BACH1 protein levels in HUVECs, whereas S1PC did not
affect BACH1 levels. However, in that study, it was found that S1PC
enhances BACH1 downregulation upon co-treatment with NOR3 (11). In that same study, to further
evaluate the role of BACH1 in the synergistic effects of S1PC and
NO-donors, we used HUVECs transfected with siRNA targeting
BACH1. Compared to cells treated with NOR3 only, the
suppression of BACH1 did not result in any significant
enhancement in HMOX1 and GCLM expression upon
co-treatment with S1PC and NOR3 (11). These results indicated that BACH1
plays a pivotal role in the synergistic effects of S1PC and NO
donors on the expression of antioxidant genes.
Bach1-deficient mice have been
reported to exhibit a resistance phenotype in disease models
Bach1-deficient mice are viable and fertile
with no obvious phenotypic abnormalities under normal conditions,
but compared to normal mice, these mice exhibit an enhanced
expression of Hmox1 in diverse tissues (13). As the enhanced expression of
Hmox1 is expected to exert antioxidant effects,
Bach1-deficient mice have been studied in several disease
models considered to involve oxidative stress. In apolipoprotein
E-deficient mice that function as an atherosclerosis-prone model,
Bach1-deficiency has been shown to reduce the plaque area
and the excretion of 8-iso-PG F2α, a marker of the systemic
oxidative stress level (14). In
several disease models listed in Table
I, Bach1 deficiency has been shown to exhibit a
resistance phenotype. These results indicate that BACH1
downregulation might have potential therapeutic applications.
However, promising lead compounds with a potential to reduce Bach1
levels have not yet been found, apart from heme-related compounds
that also have the potential to produce ROS (15).
 | Table I.Effects of Bach1 deficiency in
mouse disease models. |
Table I.
Effects of Bach1 deficiency in
mouse disease models.
| Disease models | Results of
Bach1 (−/-) mice (Refs.) |
|---|
| Atherosclerosis,
apolipoprotein E knockout mice fed a high-fat diet | Total atherosclerotic
plaque area was reduced (14). |
| Myocardial
infarction, ischemia/reperfusion injury | Myocardial infarction
was reduced (16). |
| Heart failure,
pressure overload induced hypertrophy | Left ventricular
hypertrophy was inhibited (17). |
| Diabetes,
alloxan-induced oxidative stress model | Oxidative
stress-induced apoptosis was reduced in pancreatic β-cells
(18). |
| Inflammatory bowel
diseases, trinitrobenzene sulfonic acid (TNBS)-induced colitis | TNBS-induced colitis
was ameliorated (19). |
| Steatohepatitis,
methionine-choline deficient diet model | Hepatic triglyceride
and malondialdehyde was reduced (20). |
Conclusions and future perspectives
S1PC, a sulfur amino acid present in AGE, has the
unique property of downregulating BACH1 in a NO-dependent manner
and enhancing the expression of antioxidant genes reciprocally
regulated by NRF2 and BACH1 (Fig.
1). Therefore, the interaction of S1PC and BACH1 could provide
insight into the mechanisms through which AGE exerts its
antioxidant effects in vivo.
Acknowledgements
The author would like to thank Dr Takami Oka and Dr
Koichi Tamura (Wakunaga Pharmaceutical Co. Ltd.) for providing
insight and valuable discussions pertaining to this review.
Funding
No funding was received.
Availability of data and materials
Not applicable.
Authors' contributions
TT designed the review and wrote the manuscript. The
author has read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The author declares that they have no competing
interests.
References
|
1
|
Banerjee SK and Maulik SK: Effect of
garlic on cardiovascular disorders: A review. Nutr J. 1:42002.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Budoff MJ, Ahmadi N, Gul KM, Liu ST,
Flores FR, Tiano J, Takasu J, Miller E and Tsimikas S: Aged garlic
extract supplemented with B vitamins, folic acid and L-arginine
retards the progression of subclinical atherosclerosis: A
randomized clinical trial. Prev Med. 49:101–107. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Weiss N, Ide N, Abahji T, Nill L, Keller C
and Hoffmann U: Aged garlic extract improves homocysteine-induced
endothelial dysfunction in macro- and microcirculation. J Nutr. 136
(Suppl 3):750S–754S. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Weiss N, Papatheodorou L, Morihara N,
Hilge R and Ide N: Aged garlic extract restores nitric oxide
bioavailability in cultured human endothelial cells even under
conditions of homocysteine elevation. J Ethnopharmacol.
145:162–167. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kodera Y, Ushijima M, Amano H, Suzuki JI
and Matsutomo T: Chemical and biological properties of
S−1-propenyl-l-cysteine in aged garlic extract. Molecules.
22:222017. View Article : Google Scholar
|
|
6
|
Morihara N, Hayama M and Fujii H: Aged
garlic extract scavenges superoxide radicals. Plant Foods Hum Nutr.
66:17–21. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Takabe W, Warabi E and Noguchi N:
Anti-atherogenic effect of laminar shear stress via Nrf2
activation. Antioxid Redox Signal. 15:1415–1426. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Yamato O, Tsuneyoshi T, Ushijima M,
Jikihara H and Yabuki A: Safety and efficacy of aged garlic extract
in dogs: Upregulation of the nuclear factor erythroid 2-related
factor 2 (Nrf2) signaling pathway and Nrf2-regulated phase II
antioxidant enzymes. BMC Vet Res. 14:3732018. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ogawa T, Kodera Y, Hirata D, Blackwell TK
and Mizunuma M: Natural thioallyl compounds increase oxidative
stress resistance and lifespan in Caenorhabditis elegans by
modulating SKN-1/Nrf. Sci Rep. 6:216112016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Liu XM, Peyton KJ, Ensenat D, Wang H,
Hannink M, Alam J and Durante W: Nitric oxide stimulates heme
oxygenase-1 gene transcription via the Nrf2/ARE complex to promote
vascular smooth muscle cell survival. Cardiovasc Res. 75:381–389.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Tsuneyoshi T, Kunimura K and Morihara N:
S−1-Propenylcysteine augments BACH1 degradation and heme
oxygenase 1 expression in a nitric oxide-dependent manner in
endothelial cells. Nitric Oxide. 84:22–29. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Reichard JF, Motz GT and Puga A: Heme
oxygenase-1 induction by NRF2 requires inactivation of the
transcriptional repressor BACH1. Nucleic Acids Res. 35:7074–7086.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Sun J, Hoshino H, Takaku K, Nakajima O,
Muto A, Suzuki H, Tashiro S, Takahashi S, Shibahara S, Alam J, et
al: Hemoprotein Bach1 regulates enhancer availability of heme
oxygenase-1 gene. EMBO J. 21:5216–5224. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Watari Y, Yamamoto Y, Brydun A, Ishida T,
Mito S, Yoshizumi M, Igarashi K, Chayama K, Ohshima T and Ozono R:
Ablation of the bach1 gene leads to the suppression of
atherosclerosis in bach1 and apolipoprotein E double knockout mice.
Hypertens Res. 31:783–792. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Igarashi K and Watanabe-Matsui M: Wearing
red for signaling: The heme-bach axis in heme metabolism, oxidative
stress response and iron immunology. Tohoku J Exp Med. 232:229–253.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yano Y, Ozono R, Oishi Y, Kambe M,
Yoshizumi M, Ishida T, Omura S, Oshima T and Igarashi K: Genetic
ablation of the transcription repressor Bach1 leads to myocardial
protection against ischemia/reperfusion in mice. Genes Cells.
11:791–803. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Mito S, Ozono R, Oshima T, Yano Y, Watari
Y, Yamamoto Y, Brydun A, Igarashi K and Yoshizumi M: Myocardial
protection against pressure overload in mice lacking Bach1, a
transcriptional repressor of heme oxygenase-1. Hypertension.
51:1570–1577. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kondo K, Ishigaki Y, Gao J, Yamada T, Imai
J, Sawada S, Muto A, Oka Y, Igarashi K and Katagiri H: Bach1
deficiency protects pancreatic β-cells from oxidative stress
injury. Am J Physiol Endocrinol Metab. 305:E641–E648. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Harusato A, Naito Y, Takagi T, Uchiyama K,
Mizushima K, Hirai Y, Higashimura Y, Katada K, Handa O, Ishikawa T,
et al: BTB and CNC homolog 1 (Bach1) deficiency ameliorates TNBS
colitis in mice: Role of M2 macrophages and heme oxygenase-1.
Inflamm Bowel Dis. 19:740–753. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Inoue M, Tazuma S, Kanno K, Hyogo H,
Igarashi K and Chayama K: Bach1 gene ablation reduces
steatohepatitis in mouse MCD diet model. J Clin Biochem Nutr.
48:161–166. 2011. View Article : Google Scholar : PubMed/NCBI
|