Introduction
Garlic (Allium sativum L.) is a species of
the onion family, and has been widely used as a food and also as a
folk medicine. A previous study demonstrated that aged garlic
extract (AGE) exerts an anti-proliferative effect on a panel of
both sensitive [wild-type (WT)] and multidrug-resistant (MDR) human
cancer cells (1). Following
treatment of the cells with AGE, cytofluorimetric analysis revealed
the occurrence of dose-dependent mitochondrial membrane
depolarization (MMD) (1). A large
number of epidemiological investigations have suggested that garlic
is active in the prevention and treatment of various diseases with
multiple pharmacological functions, such as anticarcinogenic
(2), antithrombotic (3), hypolipidemic (4) and hepatoprotective (5) activities. It has been reported that
garlic is able to suppress carcinogenesis and to inhibit the
proliferation of cancer cells (e.g., esophageal, gastric,
colorectal, lung, skin and prostate cancer cells) in vivo
and in vitro (6). These
studies have yielded significant findings, since the number of
cancer patients worldwide has recently increased (1). Moreover, garlic has been reported to be
beneficial in preventing neurodegeneration due to its antioxidant
and amyloid β (Aβ)-lowering properties (7). Among a number of different natural
products, such as fruits, vegetables, herbs and other compounds,
garlic has represented one of the most important sources of dietary
supplements, not only for health advantages, but also for medicinal
purposes for centuries. Among a large diversity of commercially
available garlic supplements, AGE is more widely known and has been
carefully studied. It is prepared by immersing fresh garlic in 15%
aqueous ethanol solution over a prolonged period of time (up to 20
months) at room temperature (8). AGE
is a commercial odorless preparation with antioxidant properties
for scavenging reactive oxygen species (ROS) (9,10). This
natural product has been shown to possess immunomodulatory and
anticancer properties. These effects have been demonstrated by
in vitro and in vivo experiments; however, its
mechanisms of action remain to be fully elucidated (11). It was recently demonstrated by an MTT
assay that AGE induces an anti-proliferative effect on several
sensitive and MDR human cancer cells (1).
A number of the beneficial effects of garlic have
been shown to be attributed to various bioactive compounds isolated
from garlic, including the lipid-soluble allyl sulfur compounds
(e.g., diallyl sulfide, diallyl disulfide and diallyl trisulfide)
and water-soluble compounds, such as S-allyl-L-cysteine
(SAC) and S-allylmercaptocysteine (SAMC) (12–19).
Both the organosulfur compounds, SAC and SAMC, are the major
bioactive components identified in AGE (18). Recent studies have indicated that
numerous natural bioactive molecules, including organosulfur
compounds from Allium vegetables, have been reported to have
possible preventive and therapeutic properties against certain
types of cancer (20). SAC is the
most abundant organosulfur compound in aged-garlic extract (0.62 mg
SAC/g product) and has been used to standardize the commercial
products of AGE (12). Several
studies have demonstrated that SAC exhibits antioxidant properties
by scavenging ROS and reactive nitrogen species (RNS), regulating
oxidation-related pathways, as well as neuroprotective properties
and anticancer activities (21–23).
Several other studies have also demonstrated that SAC exerts
anticancer effects by suppressing the cellular proliferation and
metastasis, and induces apoptosis in a number of cancer models,
including ovarian and prostate cancer, and hepatocellular carcinoma
(20,21,24).
Moreover, SAC as a natural product with less side-effects, may be
considered as an excellent candidate for the treatment of
neuroinflammatory diseases, such as multiple sclerosis, a
deleterious autoimmune and demyelinating disorder of the central
nervous system (CNS) with debilitating sensory and motor
complications (25–27).
In this study, mitochondrial membrane alterations of
rat liver mitochondria (RLM) and the oxidation of both endogenous
glutathione and endogenous pyridine nucleotides following
incubation with AGE, were demonstrated. Moreover, to further
examine the anti-proliferative activity of SAC, the in vitro
effects of this compound were examined by an MTT assay using the
human NB cell lines, SJ-N-KP and the MYCN-amplified IMR5 cell line;
the cell cycle and apoptosis were examined using PI and Annexin
V-FITC labeling and flow cytometric assays. By labeling the NB
cells with the probe,
5,5′,6,6′-tetrachloro-1,1′,3,3′-tetraethylimidacarbocyanine iodide
(JC-1), cytofluorimetric analysis revealed that the target of SAC
is the mitochondria, the key bioenergetic intracellular organelles,
that contain a high large number of proteins macromolecules having
pivotal functions.
Materials and methods
Reagents
Thiazolyl blue Tetrazolium bromide (MTT), verapamil,
fetal bovine serum (FBS), 5,5′-dimethyl-oxazolidine-2,4-dione
(DMO), cyclosporin A (CsA), nicotinamide adenine dinucleotide
reduced form (NADH), nicotinamide adenine dinucleotide phosphate
reduced form (NADPH),
5,5′,6,6′-tetrachloro-1,1′,3,3′-tetraethyl-imidacarbocyanine iodide
(JC-1), 3,4-dimethoxybenzoic acid, hydrochloric acid, acetonitrile
and sodium bicarbonate were purchased from Sigma-Aldrich. All cell
culture flasks and dishes were obtained from Corning, Inc. AGE and
SAC provided by Wakunaga Pharmaceutical Co. Ltd. (Hiroshima, Japan)
were manufactured as follows: Garlic cloves were sliced, immersed
in a water-ethanol mixture solution and naturally extracted for
>10 months at room temperature, as previously described
(28). The AGE powder used in our
experiments was prepared by lyophilization. It contained
approximately 28.6% (w/v, 286 mg/ml) solid material, 0.63% (6.3
mg/ml) arginine and 0.1% SAC (calculated on a dry weight basis) as
a marker compound for standardization (29). Both SAC and the AGE powder was
freshly dissolved in complete Ham's F-12 or in RPMI-1640 medium
prior to each experiment. The analysis of the organosulfur compound
content in the AGE powder was performed using a Shimadzu LC-6A HPLC
system. The content of SAC was determined by ion-exchange
chromatography, using a column TSK-GEL Amino Pak, followed by
post-derivatization using o-phthalaldehyde (OPA), and a fluorescent
detection with excitation at 340 nm and emission at 455 nm. The
test samples for the analysis of SAC were prepared by the addition
of 5% trichloroacetic acid, as previously described (30). The SAC powder was freshly dissolved
in RPMI-1640 medium supplemented with 10% FBS, normally up to 50
mg/ml, prior to each experiment. During the experiment the
solution, which was kept in the dark, was for one use only.
Cells and cell culture
Two types of NB cells were used in this study. One
was the SJ-N-KP cells, a non-amplified NB cell line and the second
one was the IMR5 cells, a n-myc amplified NB cell line (31). Both cell lines were a kind gift from
Dr N. Crescenzio (Department of Paediatrics, University of Turin,
Regina Margherita Children's Hospital, Turin, Italy) and Dr F.
Timeus (Paediatric Haematology-Oncology, Regina Margherita
Children's Hospital) (31). The NB
cell lines, SJ-N-KP and IMR5 were maintained in monolayer cultures
in RPMI-1640 medium supplemented with 10% FBS, 2 mM L-glutamine,
100 µg/ml streptomycin and 100 IU/ml penicillin (31). Both cell lines were incubated in a
humidified atmosphere of 5% CO2 in a water-jacketed
incubator at 37°C. For each passage, exponentially growing SJ-N-KP
and IMR5 cells were harvested with 10 mM EDTA and by the further
addition of 0.10% trypsin solution. The trypsin activity was
quenched by the addition of complete RPMI-1640 medium (31).
Determination of apoptotic cell death
by Annexin V-FITC staining
To detect phosphatidylserine residue exposure on the
surface of the plasma membrane of the tumor cells in the initial
step of apoptosis, an Annexin V-FITC apoptosis detection kit was
used as previously described by Van Engeland et al (32). The NB cell lines, SJ-N-KP and IMR5
(9.1×104 cells/ml), were seeded in a 6-well plate,
containing complete RPMI-1640 medium and incubated for 24 h at 37°C
to allow for the the complete reattachment of the cells to the
plates. After exchanging the medium with fresh medium, the cells
were incubated at 37°C in the presence of 0, 10 and 20 mM SAC for
48 h. Following incubation at 37°C, the cells were detached, washed
with PBS, centrifuged at 400 × g for 2 min at 25°C and then stained
with 1 µg/ml of Annexin V-FITC and with 1 µg/ml of PI for 10 min at
room temperature in the dark. Annexin V-FITC and PI fluorescence
were measured on the FL-1 channel (533/30 nm) and the FL-3 channel
(>670 nm), respectively, with excitation at 488 nm. A minimum of
10,000 events/sample was acquired (32).
Cell cycle analysis
Cell cycle distribution was analyzed by labeling the
cells with PI. The assays were carried out as previously described
by Nicoletti et al (33).
S-allyl-L-cysteine-treated and the untreated NB cell lines, SJ-N-KP
and IMR5 (9.1×104 cells/ml), as described above in
Annexin V-FITC labeling, were harvested, washed twice with cold PBS
and centrifuged at 400 × g for 2 min at 4°C. The pellet was fixed
in 70% ethanol at −20°C for 1 h. After washing twice with PBS, the
cells were resuspended in PBS containing 100 µg/ml RNase A and 40
µg/ml PI. Following incubation at 37°C for 1 h, the cells were
subsequently analyzed by flow cytometry using the FL-3 channel
(>670 nm) with the acquisition of 10,000 events/sample.
SAC dose response assay
The NB cell lines, SJ-N-KP and IMR5, were seeded in
a 96-well plate and incubated for 24 h to allow for the complete
reattachment of the cells to the plates. After the medium was
replaced with fresh medium, the cells were incubated in the
presence of 0, 1, 5, 10, 20 and 30 mM SAC for 48 h. The
anti-proliferative effects of SAC on the human NB cells were
examined by MTT assay. Briefly, MTT was added to each well at a
final concentration of 0.5 mg/ml. Following 3 h of incubation at
37°C, dimethyl sulfoxide was added to dissolve the crystals. The
absorbance was determined at 577 and 660 nm using a
spectrophotometer multi-mode plate reader [Synergy HT BioTek,
serial no. 270204; BioTek, Bernareggio (MB)].
Measurements ‘in situ’ of
mitochondrial membrane potential (Δψm)
The changes in Δψm in whole cells were assayed using
the lipophilic cationic probe, JC-1 dye. The NB cell lines, SJ-N-KP
and IMR5, were seeded in a 12-well plate and incubated for 24 h at
37°C to allow the cells to adhere to the plates. After the medium
was replaced with fresh medium, the cells were incubated with 30
and 50 mM of SAC for 24 h at 37°C. Subsequently, the cells were
stained with 2.5 µg/ml of JC-1 for 20 min at 37°C. The detached
cells were washed with PBS and then resuspended in PBS. The samples
were then analyzed using a BD Accuri C6 flow cytometer (BD
Biosciences). JC-1 was excited using an argon laser at a wavelength
of 488 nm (using a BD Accuri C6 flow cytometer). The emitted green
(JC-1 monomer) and red (JC-1 aggregate) fluorescence were detected
at the FL-1 channel (533/30 nm) and FL-2 channel (585/40 nm),
respectively. At least 10,000 events/sample were acquired in log
mode. The ratio of red (FL2)/green (FL1) fluorescence intensity was
used to represent the Δψm.
Gas chromatography-mass spectrometry
(GC-MS) analysis of AGE and SAC
SAC was analyzed by GC-MS as its
tert-butyldimethylsilyl (TBDMS) derivative according to
Jiménez-Martín et al (34).
Briefly, a 20 µl aliquot of a solution containing SAC at 1 mg/ml in
0.1 N HCl spiked with 15 µl internal standard (3,4-dimethoxybenzoic
acid, 0.1 mg/ml) was dried, and 50 µl of neat
N-tert-butyldimethylsilyl-N-methyltrifluoroacetamide
(MTBSTFA), followed by 50 µl of acetonitrile, were added. The
mixture was heated at 100°C for 2 h. The sample was then
neutralized with sodium bicarbonate and subjected to GC-MS
analysis. The same derivatization protocol was used for AGE power
(1 mg/0.1 M HCl). GC-MS analyses were performed with an Agilent
7890B gas chromatograph coupled to a 5977B quadrupole mass
selective detector (Agilent Technologies). Chromatographic
separations were carried out with an Agilent HP5 ms fused-silica
capillary column (30 m × 0.25 mm i.d.) coated with
5%-phenyl-95%-dimethylpolysiloxane (film thickness, 0.25 µm) as a
stationary phase. The injection mode was splitless at a temperature
of 280°C, the column temperature program was 70°C (1 min) then to
300°C at a rate of 20°C/min and held for 10 min. The carrier gas
was helium at a constant flow of 1.0 ml/min. The spectra were
obtained in the electron impact mode at 70 eV ionization energy;
ion source 280°C; ion source vacuum 10−5 Torr. MS
analysis was performed simultaneously in TIC (mass range scan from
m/z 50 to 600 at a rate of 0.42 scans/sec) and SIM mode. GC-SIM-MS
analysis was performed selecting the following ions: m/z 302 and
m/z 332 for SAC and m/z 239 for 3,4-dimethoxybenzoic acid (internal
standard).
Animals
A total of 50 male Wistar rats, 2 months old,
weighing approximately 150 g, were used in our experiments. The
rats, housed in the animal facility of the Department of Biomedical
Sciences, University of Padova, were maintained under controlled
conditions (temperature 20–22°C, relative humidity 48–50%, water
with antibacterial control and a 12:12 h light/dark cycle) and
provided with water and a standard diet (4RF25) purchased by
Mucedola s.r.l., Settimo Milanese (MI) according to Ohkubo et
al (1). The experimental
procedures were approved by the local Ethics Committee for Animal
Experimentation (CEASA) (protocol no. 3619, 15.1.2014) and
performed in agreement with the international guidelines, as well
as European Communities Council Directive and National Regulations
(CEE Council 86/609 and DL 116/92).
RLM isolation and purification
Rat liver mitochondria (RLM) were isolated by the
differential centrifugation method according to Ohkubo et al
(1). In detail, the rats were
starved overnight and sacrificed by cervical dislocation. The
livers were rapidly explanted, immersed in ice-cold isolation
medium containing 250 mM sucrose, 5 mM HEPES (pH 7.4), 0.5 mM EGTA
and washed 4/5 times with the same medium. The livers were minced
into small sections and washed with ice-cold fresh medium without
EGTA. The suspension was transferred to a glass potter and
homogenized using a Teflon pestle operating at 1,600 rpm, by 3–4
strokes. The homogenate was centrifuged at 700 × g for 5 min at 4°C
and the obtained supernatant was centrifuged at 10,800 × g for 10
min at 4°C. The pellet was washed with isolation medium,
resuspended and centrifuged at 15,900 × g for 10 min at 4°C.
Finally, the obtained pellet, containing the mitochondria, was
suspended in the standard medium (see the incubation procedure)
(35). Mitochondrial proteins were
measured by the biuret method with BSA, as a standard (36). The mitochondria (1 mg protein/ml)
were incubated in a water-jacketed cell at 25°C under continuous
stirring. The standard medium contained 250 mM sucrose, 10 mM HEPES
(pH 7.4), 5 mM Na-succinate, 1.25 µm rotenone and 1 mM
Na-phosphate. Variations and/or other additions are provided with
each experiment.
Determination of mitochondrial
functions
Mitochondrial membrane potential (ΔΨm)
was calculated by detecting the distribution of the lipid-soluble
cation tetraphenylphosphonium (TPP+, Sigma-Aldrich)
through the inner membrane, measured by a TPP+ selective
electrode, according to Kamo et al (37). The mitochondrial electrochemical
gradient (58ΔpHm) was calculated from the distribution
of DMO ([14C]5,5′-dimethyl-oxazolidine-2,4-dione) across
the mitochondrial membrane, according to Rottemberg (38). 58 is the value in mV of RT/zF for
giving the ΔpH value in mV. The redox state of mitochondrial
glutathione was detected by using the Ellman reagent, according to
Tietze (39). The change with time
of the redox state of mitochondrial pyridine nucleotides was
followed fluorometrically in an Aminco Bowman 4–8202
spectrofluometer with excitation at 354 nm and emission at 462
nm.
Statistical analysis
The data were presented as the means ± SEM or means
± SD. Statistical analysis was performed using one-way ANOVA with
the Dunnett's post hoc test, in EZR (Kanda) (40). A P-value <0.05 was considered to
indicate a statistically significant difference.
Results
Mitochondrial membrane alterations of
RLM following incubation with AGE
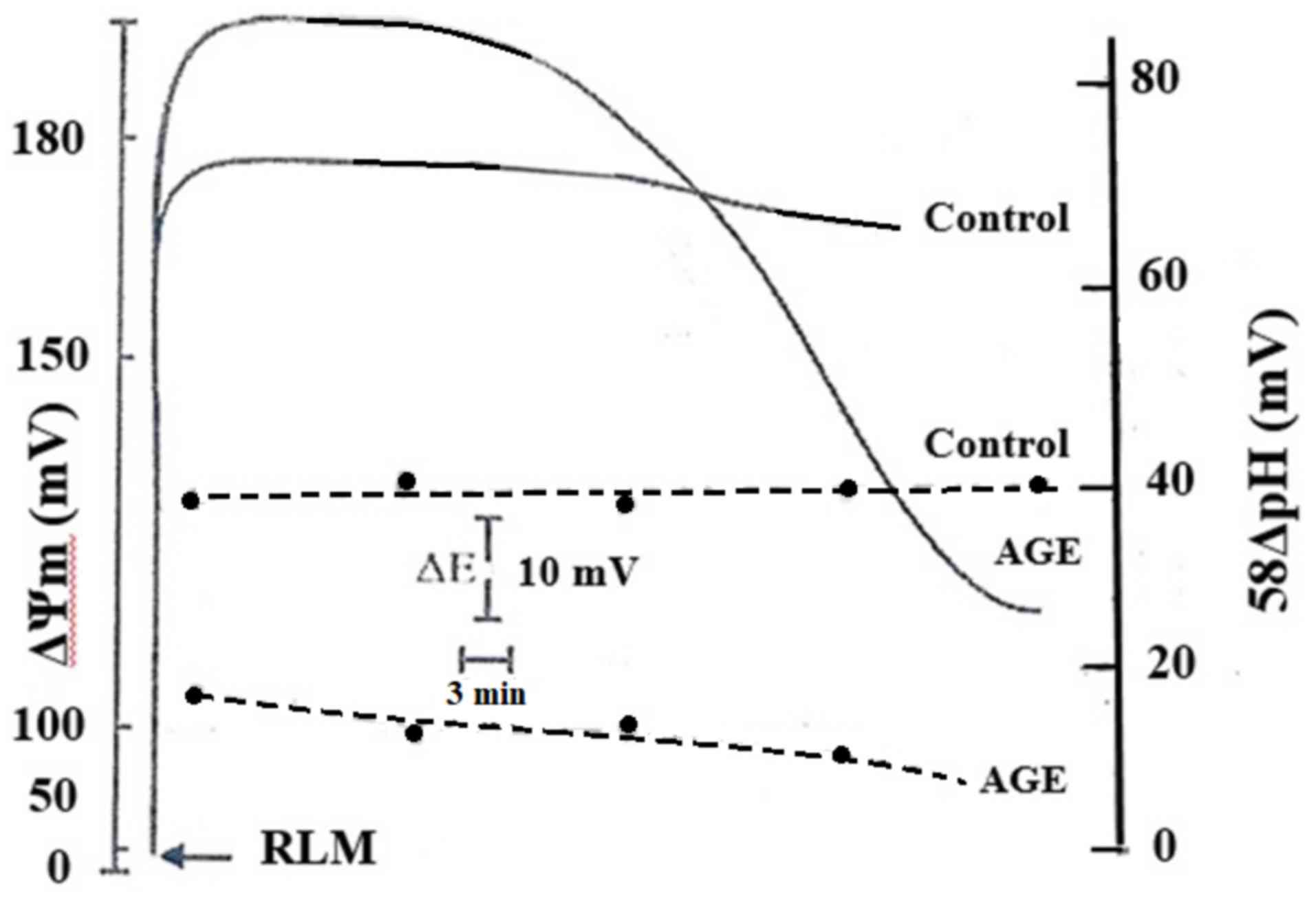
Fig. 1 shows the
effects of AGE on the components of the electrochemical gradient
(ΔµH+), membrane potential, ΔΨm,
and the chemical gradient, ΔpHm, expressed in mV. The
results revealed that under normal conditions, both the
ΔΨm and 58ΔpHm exhibited the physiological
values of 170 and 40 mV, respectively. The presence of AGE induced
an initial increase in ΔΨm of approximately 30 mV and a
corresponding identical decrease in 58ΔpHm. However,
after approximately 15 min, the ΔΨm gradually decreased
until 120 mV. In addition, the 58ΔpHm value underwent a
parallel drop of approximately 10 mV. The membrane alterations of
RLM by AGE are shown in Fig. 1. The
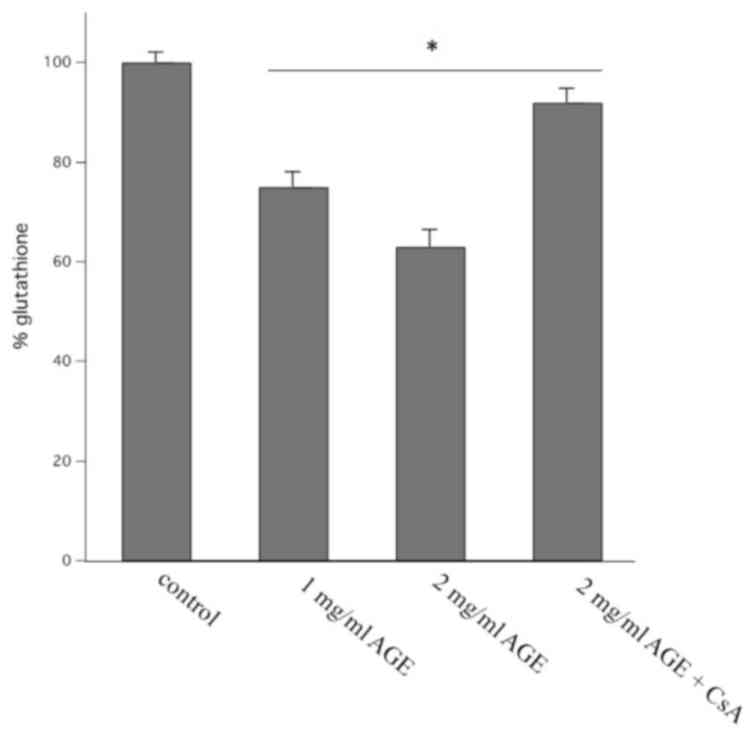
decrease in both ΔΨm and 58ΔpHm was
accompanied by an increase in the oxidation of endogenous
glutathione content. This is evidenced by the decrease in the
reduced content of glutathione induced by different concentration
of AGE as reported in Fig. 2. AGE
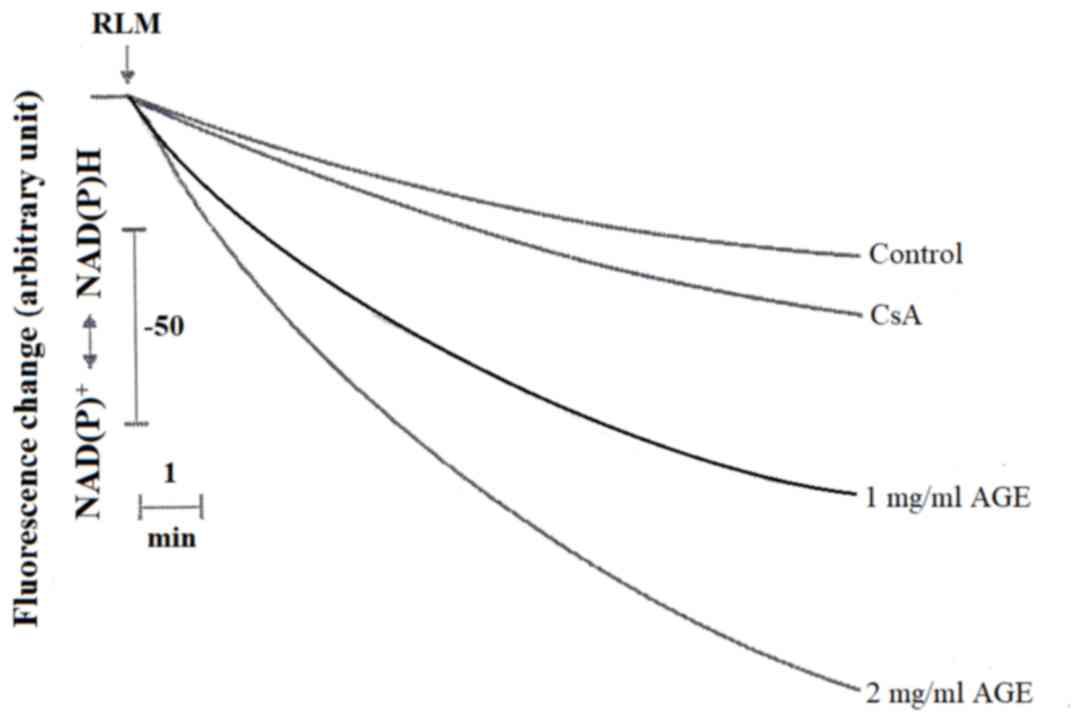
also caused the oxidation of endogenous pyridine nucleotides, NADH
and NADPH, as observed by the downward deflection of the curves
detecting the reduced state of these nucleotides in Fig. 3. It is also to evidence that
cyclosporine A (CsA) strongly inhibits the oxidation of both
demetabolites.
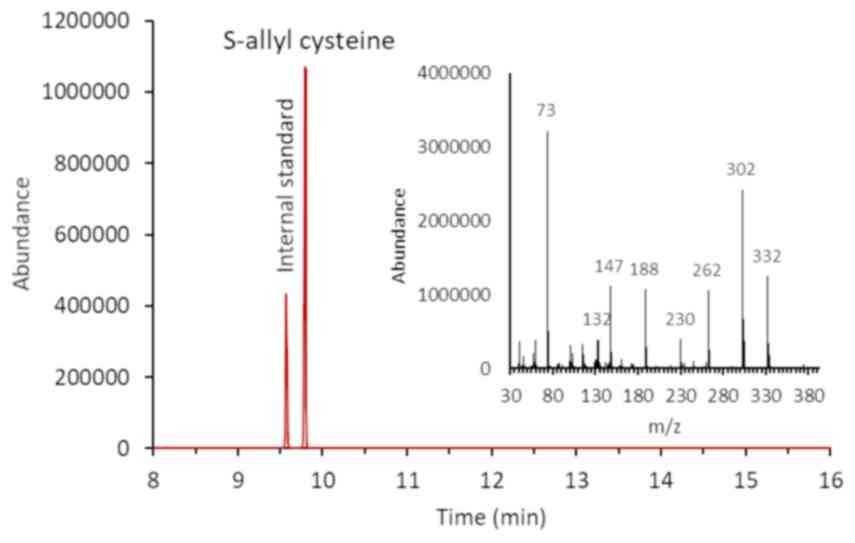
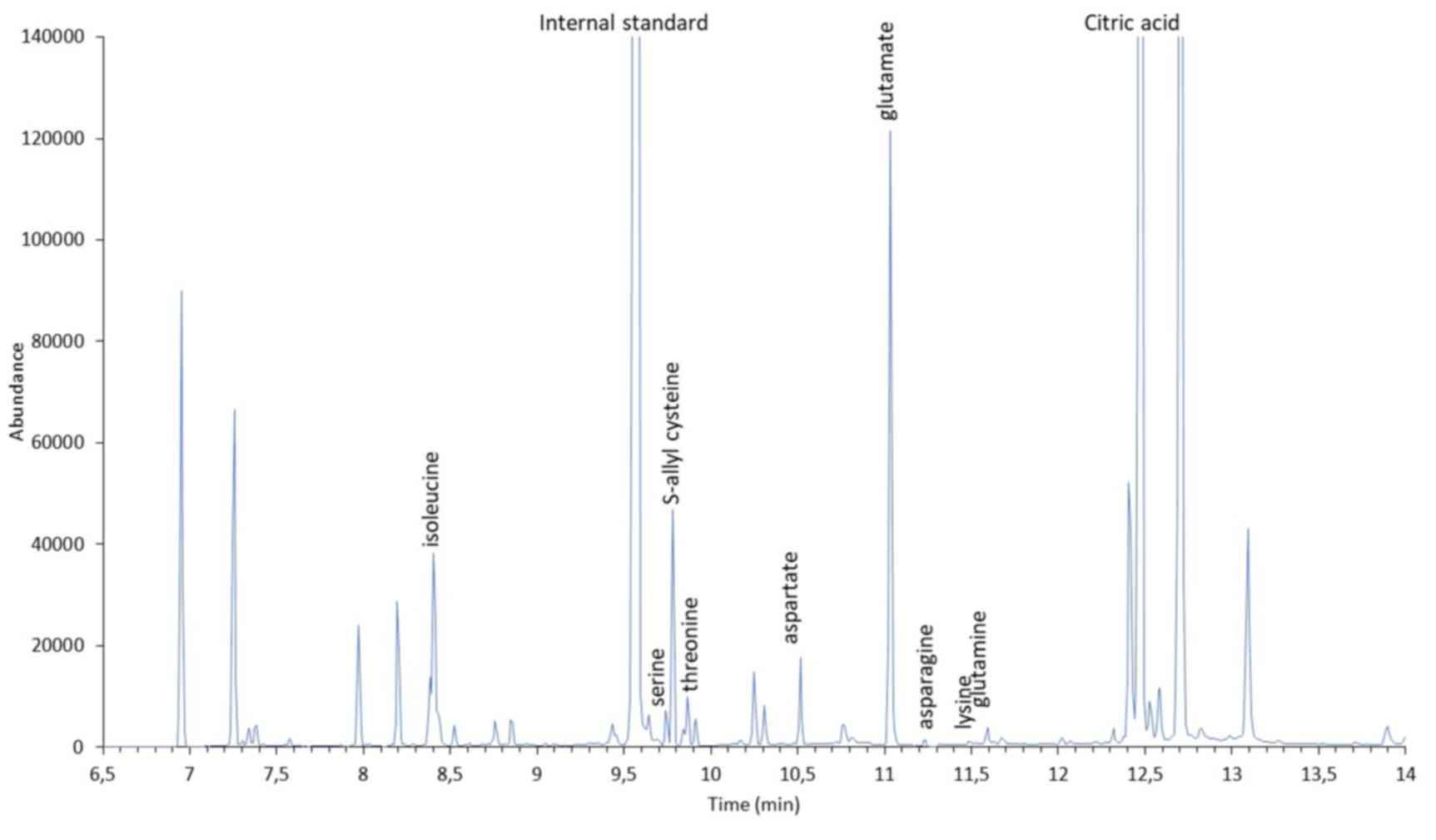
GC-MS analysis of AGE and SAC
In this study, we evaluated the use of the
silylation reagent, MTBSTFA, for the derivatization of SAC. MTBSTFA
derivatives are more stable and less moisture-sensitive than those
formed using lower molecular weight reagents. A chromatogram of SAC
is presented in Fig. 4. In the
chromatographic condition described above, SAC has a retention time
of 9,8 min and an electron impact mass spectrum (Fig. 4, inset) showing a typical fragment
(m/z 302) corresponding to the molecular weight of the derivative
less C3H5S (M-73). In Fig. 5, a typical GC-MS chromatogram of AGE
is presented. The chromatogram shows the presence of a peak having
the same retention time and mass spectrum of SAC. In addition,
TBDMS derivatization allows for the identification of in AGE a
number of polar compounds, including amino acids and carboxylic
acids. Amino acid derivatives were identified by comparing spectral
data with those reported in the NIST2018 library. Electron impact
spectra of these derivatives contain typical fragments
corresponding to the molecular weight of the derivative less
CH3 (M-15), C4H9 (M-57),
C4H9 + CO (M-85) and CO-O-TBDMS (M-159).
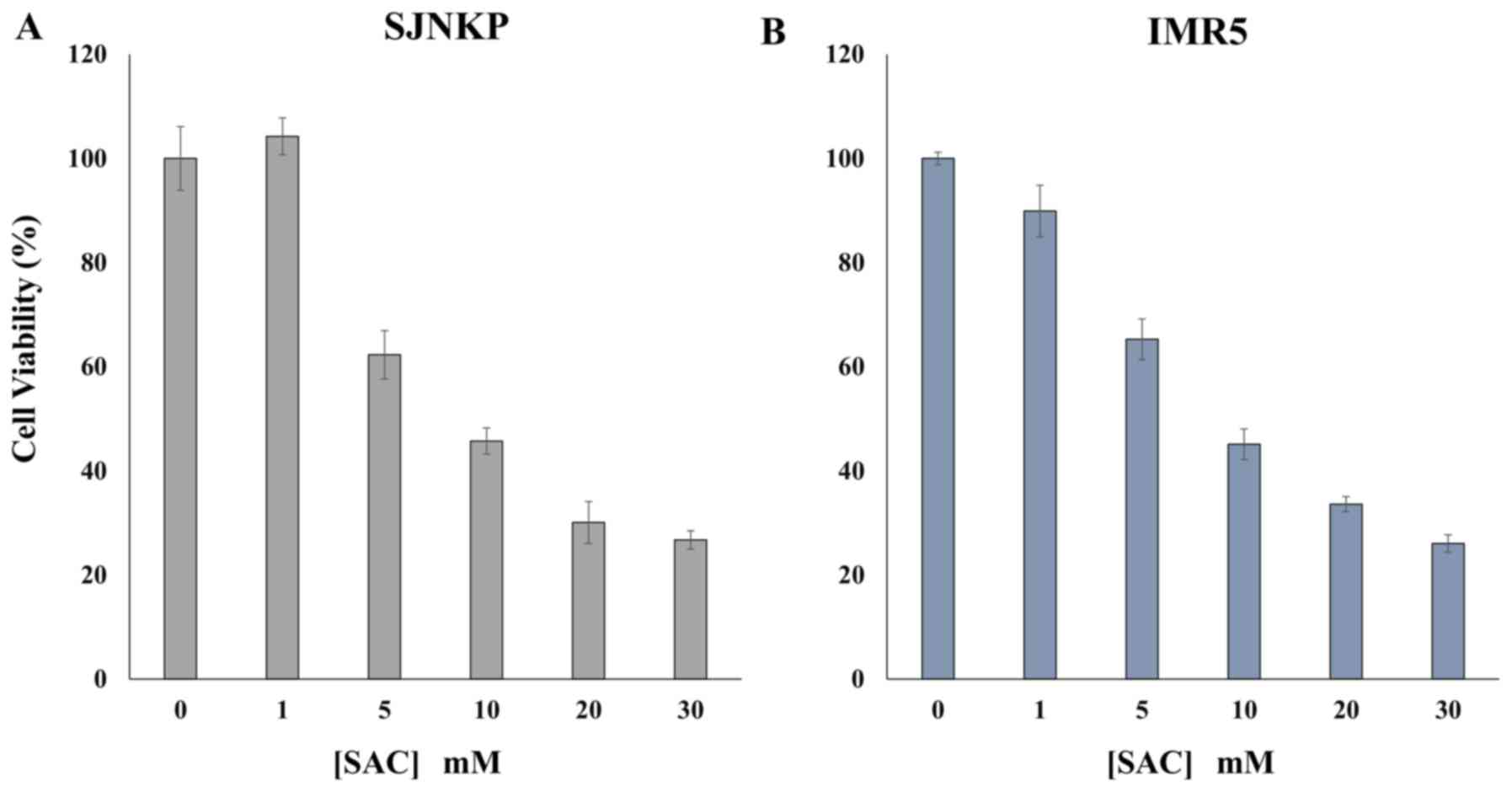
Anti-proliferative effects of SAC on
the human NB cell lines, SJ-N-KP and IMR-5
The results of MTT assays revealed that treatment
with SAC exerted anti-proliferative effects on the NB cancer cells
examined in a dose-dependent manner. Treatment with 20 mM of SAC
for 48 h decreased the viability of both the SJ-N-KP and IMR-5
cells to 30.05 and 33.58%, respectively, when compared with the
untreated control cells (Fig. 6).
The proliferation of both cell lines was already markedly decreased
to approximately 63.77% of the control level following treatment
with 5 mM SAC for the same incubation time of 48 h (Fig. 6). A further extension of the
incubation time at 72 h, did not considerably affect the viability
of the cells (data not shown). A concentration of <5 mM SAC
exerted a modest effect of about 10% only on the IMR-5 cells.
Analysis of the induction of apoptosis
by flow cytometry
The loss of phospholipid asymmetry and the
appearance of phosphatidylserine residues on the outer layer of the
plasma membrane is an early signal of cell death, namely apoptosis.
Annexin V is a Ca2+-dependent phospholipid-binding
protein with a high affinity for phosphatidylserine. In this study,
to detect externalized phosphatidylserine, Annexin V-FITC/PI
staining was performed in the NB cell lines, SJ-N-KP and IMR5. The
results of flow cytometric analysis are presented in Fig. 7. Treatment with 10 and 20 mM of SAC
increased the percentage of the total apoptotic cells (Fig. 7A, middle and upper right panels) to
18.1 and 42.6%, respectively, compared with the untreated SJ-N-KP
cells (5.2%) (Fig. 7A, upper left
panel). On the contrary, the percentage of apoptotic cells was
higher in the IMR5 cells than the SJ-N-KP cells following treatment
with 10 and 20 mM of SAC (Fig. 7A,
lower panels, 33.0 and 51.7%, respectively, and Fig. 7B), compared with the untreated IMR5
cells (6.2%) (Fig. 7A, lower left
panel).
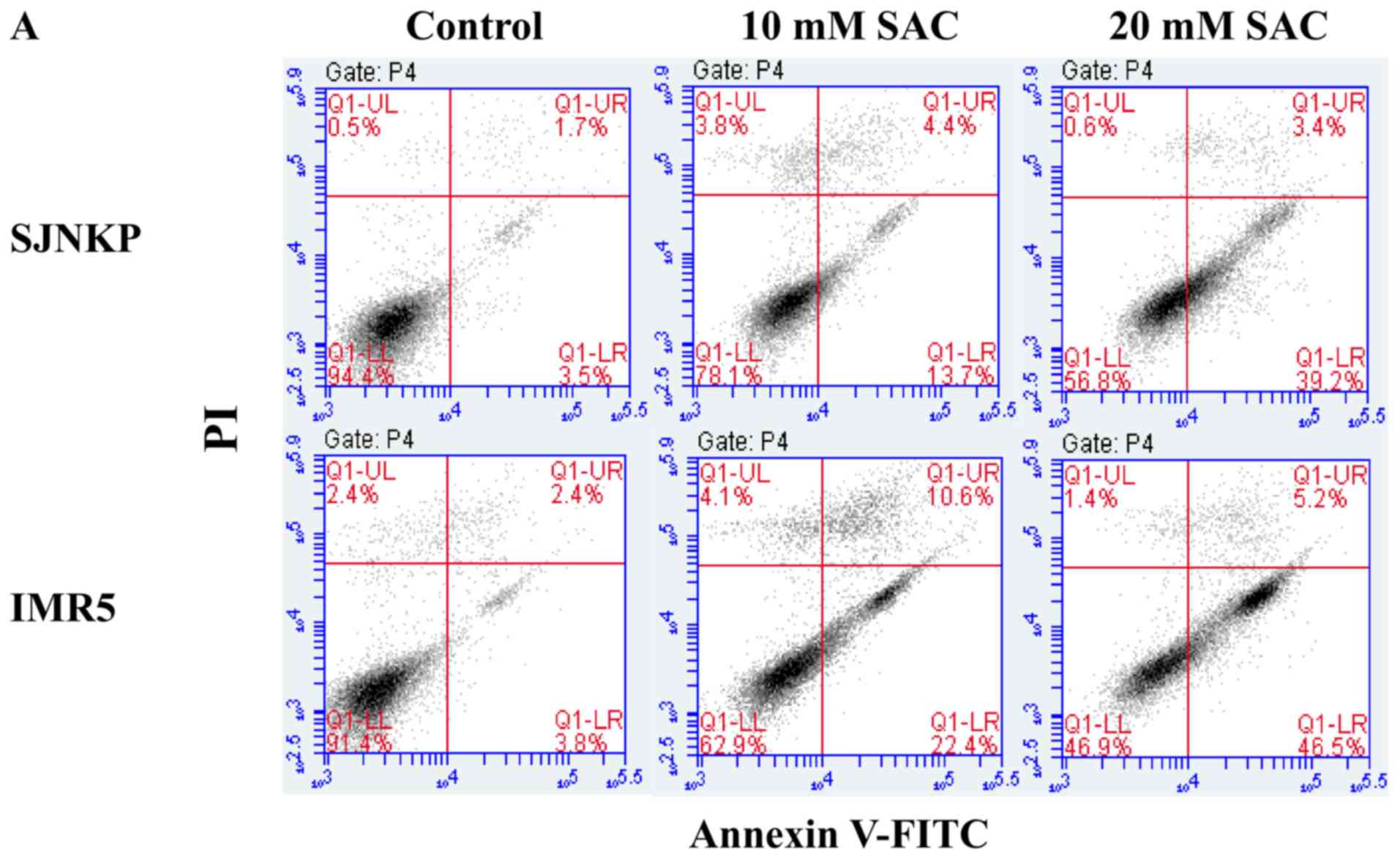 | Figure 7.Annexin V/PI assay: Effect of SAC on
SJ-N-KP and IMR5 cells. Flow cytometric analysis of apoptosis of
neuroblastoma cells following double labeling with Annexin V-FITC
and PI. Neuroblastoma cells were treated with increasing
concentrations (0, 10 and 20 mM) of SAC. At 48 h after the end of
the treatment, and incubation at 37°C, cells were analyzed by flow
cytometry. (A) Representative Annexin V-FITC and PI flow cytometry
dot plots of SJ-N-KP and IMR5 cells are shown. The x-axis
represents FITC staining, and the y-axis represents PI staining.
The percentage of cells displaying Annexin V-FITC
positive/PI-negative (early apoptosis), Annexin V-FITC
positive/PI-positive (late apoptotic or dead), Annexin V-FITC
negative/PI-positive (necrotic) and double negative cells (viable
cells) is indicated. The dot plots profile of cells were obtained
from 1 out of 2 independent experiments, performed in the same
experimental conditions, which yielded similar results. (B) Each
bar represents the mean ± SD of normal or total apoptotic cells of
2 independent experiments. Data were analyzed by one-way ANOVA,
followed by a Dunnett's post hoc test. (A) *P<0.05 vs. control
SJ-N-KP cells; and (B) *P<0.05 and ***P<0.001 vs. control
IMR5 cells. SAC, S-allyl-L-cysteine. |
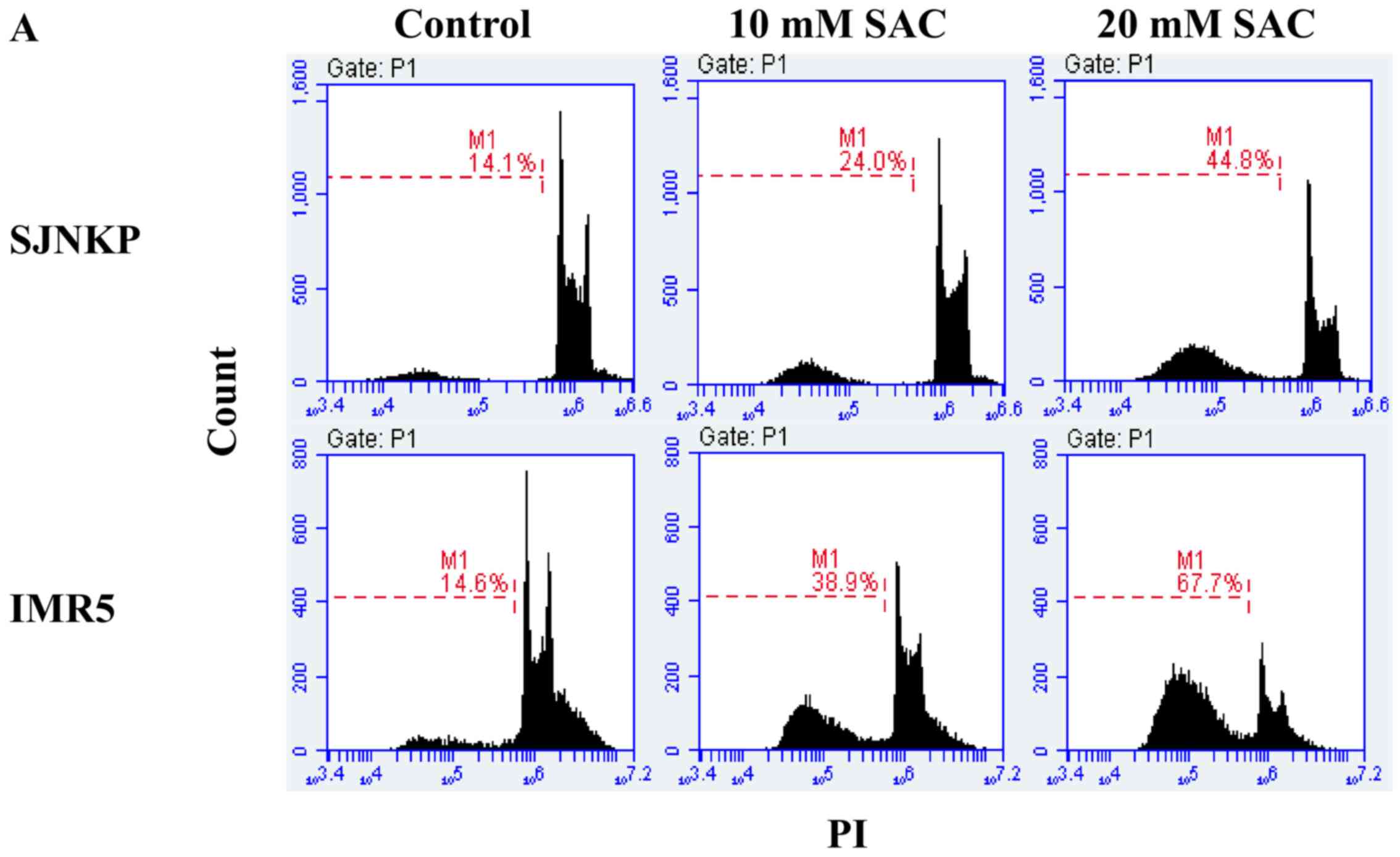
To confirm the involvement of apoptotic cell death
induced by SAC, flow cytometric analysis using PI staining was
performed to analyze the cell cycle status. The apoptotic cells
that undergo DNA fragmentation exhibit sub-G1 DNA contents. The
exposure of the NB cell lines, SJ-N-KP and IMR5, to 10 and 20 mM of
SAC for 48 h induced a significant increase in the sub-G1 apoptotic
cell population, compared to that observed in the untreated control
cells (Fig. 8). The percentage of
treated IMR5 cells in the sub-G1 phase (38.9 and 67,7%) was higher
than the percentage of SJ-N-KP cells in the same phase (24.0 and
44.8%), at both concentrations of SAC, which was consistent with
the results of the Annexin V-FITC/PI staining assay (Fig. 7). These results confirm the
cytotoxicity induced by both concentrations (10 and 20 mM) of SAC
and suggest the involvement of an apoptotic mechanism.
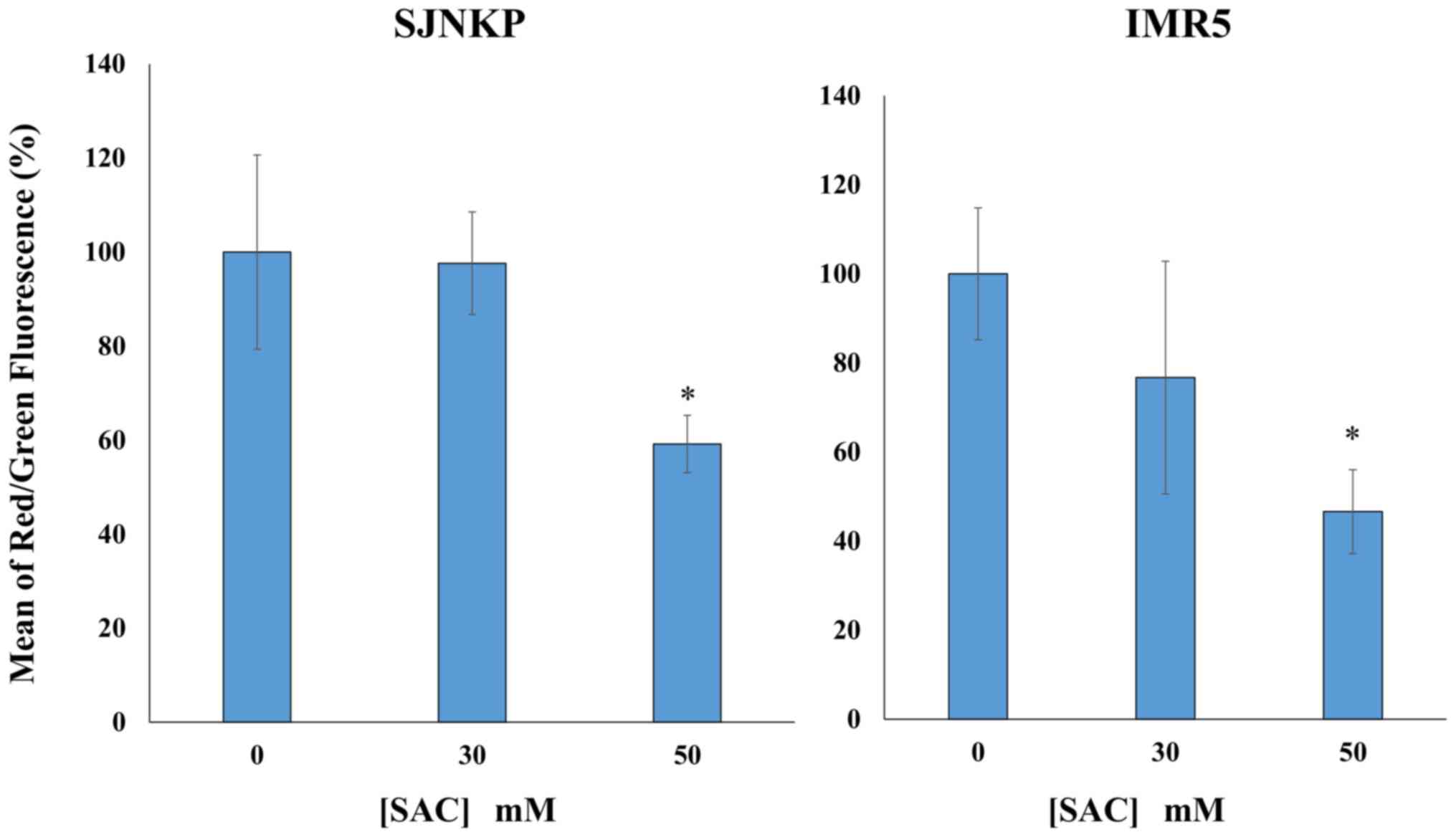
Depolarization of Δψm induced by SAC
in the NB SJ-N-KP and IMR5 cell lines
To investigate the mechanisms through which SAC
induces cell death, we examined the loss of Δψm using a flow
cytometric analysis of the control and treated cells loaded with
the mitochondrial probe, JC-1. In healthy cells, membrane-permeable
JC-1 dye spontaneously accumulates in the mitochondria and forms
aggregates known as J-aggregates that emit red fluorescence after
excitation. By contrast, in apoptotic cells, with a low Δψm, JC-1
remains in the cytoplasm as monomers, which emit green
fluorescence. Therefore, the loss of Δψm is indicated by a decrease
in the ratio of red/green fluorescence intensity. As shown in
Fig. 9, in both the NB cell lines
SJ-N-KP and IMR5, exposure to SAC induced an evident MMD. Treatment
with SAC increased the ratio green/red fluorescence intensity and
concomitantly decreased the red fluorescence intensity in a
dose-dependent manner. The SAC-induced Δψm dissipation was more
evident in the IMR5 cells than the SJ-N-KP cells following
treatment with 50 mM SAC solutions for 48 h at 37°C (average 46.6
and 59.1%, respectively). Moreover, a decrease in MMD was already
observed in the IMR5 cells at lower concentrations of SAC, 30 mM,
whereas the same concentration of SAC did not induce any variations
in the SJ-N-KP cells, as shown in Fig.
9.
Discussion
Therapeutic approaches based on isolated compounds
obtained from natural products to handle central and peripheral
disorders involving oxidative stress and inflammation are more
common nowadays. Among the natural compound, garlic, a species of
the Allium genus, has been used throughout recorded history
for both culinary and medicinal purposes (20,21,24). In
our previous study, as well as in numerous other reports, it was
observed that AGE induced a decrease in the viability of several
human cancer cell lines (1,20,24,41).
This result represents an interesting finding, since in
conventional cancer chemotherapy, numerous issues hamper successful
treatment. Therefore, the development of more effective and less
toxic therapeutic regimens is vital for the improvement of survival
of patients with cancer. Our previous study revealed that the
treatment of a gastric adenocarcinoma cell line (AGS), a human
cervical adenocarcinoma cell line (HeLa), a melanoma cell line
(M14), a human colon adenocarcinoma cell line (LoVo WT) and its MDR
variant (LoVo DX) cells with AGE was accompanied by characteristic
mitochondrial alterations, intracellular organelles that represent
the bioenergetic key (1). The
mitochondrial probe JC-1 dye exposure on the outer surface of the
mitochondrial membrane, clearly revealed the onset of the MMD
process. In general, the mitochondrial dysfuntions of the
above-mentioned WT cell lines and the resistant phenotype mirrored
the results of the cell survival experiments. In addition, it was
observed that AGE acts in the RLM as a K+/H+
antiporter that directly affects some mitochondrial bioenergetic
functions. In fact, the prolonged incubation of the RLM with AGE
induced depolarization and matrix swelling, typical alterations of
mitochondrial permeability transition phenomenon (1). Further new findings reported in this
study, demonstrated that AGE, during the first 15 min of
incubation, functioned as a typical activator of the
K+/H+ exchanger, as mentioned above. This was
demonstrated by the increase in ΔΨm induced by AGE, that
exhibited a similar effect of the typical
K+/H+ ionophores, nigericin and salinomycin
as previously demonstrated (1). The
decrease in ΔΨm and 58ΔpHm triggered after 15
min of incubation, was accompanied by the oxidation of both
endogenous glutathione and endogenous pyridine nucleotides
(Figs. 2 and 3). The oxidation process can be explained
as reported in our previous study (1). These results demonstrate that the
collapse in the ΔµH+ is indicative of a general
oxidative stress induced by AGE, leading to the phenomenon of
mitochondrial permeability transition and potentially triggering of
apoptotic pathway in the cell. The induction of MPT is strongly
supported by the prevention of the oxidation of both glutathione
and pyridine nucleotides exhibited by CsA, well known typical
inhibitor of MPT. Moreover, these data indicate that AGE is able to
cause oxidative stress in the absence of endogenous
Ca2+, following 15 min of incubation, as suggested in
our previous study (1). The observed
oxidation of glutathione and pyridine nucleotides demonstrate the
involvement of a coupled redox cycling between the above compounds
that is essential for mitochondrial physiology.
To support the anti-proliferative effects of AGE,
experiments were performed on NB cancer cells, the most common
cancer observed in infancy and the most frequent cause of death
from extracranial malignant solid tumors in children (42). NB originates from the sympathoadrenal
lineage of neural crest and accounts for 15% of childhood cancer
mortality. The amplification of the oncogene, N-Myc, is a
well-established poor prognostic marker for NB. The N-Myc
amplification status is strongly associated with a higher tumor
aggression and resistance to treatment. Therefore, besides the
current treatments that include chemotherapy, surgery and
radiation, new therapies for patients with N-Myc-amplified NB need
to be developed (43,44). In this study, two NB cell lines,
SJ-N-KP and MYCN-amplified IMR5, were treated with an AGE
constituent, SAC, with the aim to demonstrate the anticancer
activity of this compound and to elucidate the possible underlying
molecular mechanisms (Figs. 4 and
5, GC-MS chromatograms). During the
aging of garlic (up to 20 months), the hydrolysis of the
γ-glutamyl-S-allylcysteine by γ-glutamyltransferase to form
SAC, the major organosulfur compound of AGE occurs (45). SAC, an antioxidant with
neuroprotective properties, is a water-soluble and non-toxic
compound (46), and is stable since
it remains relatively unaltered in AGE for up to 2 years (45). The antioxidant properties of SAC have
been attributed to its capacity to (reviewed in ref. 47) i)
scavenge ROS/RNS due to the allyl group in its structure (48), thus protecting cells against lipid
peroxidation, protein oxidation and nitration as well as
mitochondrial damage; ii) chelate metal ions implicated in ROS/RNS
formation, such as Fe2+ and Cu2+ (49); iii) increase non-enzymatic
antioxidant defense, such as reduced glutathione (50); iv) inhibit pro-oxidant enzymes, such
as NADPH oxidase and nitric oxide synthase (51); v) inhibit the NF-κB inflammatory
pathway (52,53); and vi) contribute to
anti-inflammatory responses by increasing NO in endothelial cells
(51).
In this study, the effects of cell growth inhibition
induced by SAC on NB cells were investigated. Since the efficacy of
therapeutic agents depends on its long-term effect on cancer cells,
we examined the effects of the treatments on in vitro
tumorigenic capacity by employing MTT assay, which is widely
considered to be a valid method for the evaluation of tumor cell
sensitivity to anticancer drugs. Following treatment of the NB
cells, SJ-N-KP and MYCN-amplified IMR5 cells, with SAC at 20 mM at
37°C for 48 h, cell proliferation was detected by MTT assay. The
cell survival assay revealed that SAC exerted an anti-proliferative
effect on both NB human cancer cells (Fig. 6).
Apoptosis was evaluated by flow cytometry, in the
same experimental condition, using Annexin V-FITC labeling and DNA
staining with propidium iodide. The results revealed that treatment
with SAC increased the percentage of apoptotic cells in a SAC
dose-dependent manner. The percentages of Annexin V-positive cells
were 48.0% in the SJ-N-KP and 50.1% in the IMR5 cells (Fig. 7), indicating that treatment with SAC
was able to induce cell death through apoptosis. Cell cycle
analysis revealed that the percentages of Annexin V-positive cells
were in accordance with those of cells showing a hypodiploid sub-G1
peak (Fig. 8). It is known that the
mitochondria play a pivotal role in the intrinsic apoptotic pathway
and a reduction in Δψm is an early irreversible step (54). By labeling the NB cells with the
probe JC-1 dye, cytofluorimetric analysis revealed that the target
of SAC was the mitochondria. SAC-induced apoptosis was preceded by
the Δψm dissipation in both the IMR5 and SJ-N-KP cells, following
treatment with 30 and 50 mM SAC (Fig.
9). In view of these results, the use of AGE and SAC in cancer
therapy deserves to be taken into consideration. The findings
indicate that both AGE and SAC induce the apoptosis of cancer cells
by altering mitochondrial permeability transition and that the
mitochondria may become a promising target for the treatment of
malignancies.
Acknowledgements
The authors would like to thank the ‘International
Polyamine Foundation-ONLUS’ for the availability to look up the
polyamine documentation. AGE and SAC were a kind gift from Wakunaga
Pharmaceutical Co. Ltd. (Japan).
Funding
This study was funded by the generous support of ‘La
Sapienza’ University of Rome and Italian MIUR (Ministero
dell'Istruzione, dell'Università e della Ricerca), AIRC IG 17575,
IG 20801; Istituto Pasteur-Fondazione Cenci-Bolognetti,
AFM-Telethon grant #21025. EA would like to thank Wakunaga
Pharmaceutical Co. Ltd. (Japan) for the scholarship given to YK for
the support of his PhD research work.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article or are available from the
corresponding author on reasonable request.
Authors' contributions
EA, YK and AT conceived this study and coordinated
the collaboration among the authors. YK, LDV, AM and GC performed
all the experiments on the NB cancer cells. EA, AT, AM and AG
optimized the protocols for the analyses performed by flow
cytometry and GC-MS. All authors wrote the manuscript and all
authors have read and approved the final manuscript.
Ethics approval and consent to
participate
The animal experimental procedures were approved by
the local Ethics Committee for Animal Experimentation (CEASA)
(protocol no. 3619, 15.1.2014) and performed in agreement with the
international guidelines as well as European Communities Council
Directive and National Regulations (CEE Council 86/609 and DL
116/92).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
AGE
|
aged garlic extract
|
|
SAC
|
S-allyl-L-cysteine
|
|
SAMC
|
S-allylmercaptocysteine
|
|
ROS
|
reactive oxygen species
|
|
RNS
|
reactive nitrogen species
|
|
CNS
|
central nervous system
|
|
WT
|
wild-type
|
|
MDR
|
multidrug-resistant
|
|
NB
|
neuroblastoma
|
|
RLM
|
rat liver mitochondria
|
|
MTT
|
thiazolyl blue tetrazolium bromide
|
|
FBS
|
fetal bovine serum
|
|
JC-1
|
5,5′,6,6′-
tetrachloro-1,1′,3,3′-tetraethylimidacarbocyanine iodide
|
|
BSA
|
bovine serum albumin
|
|
Δψm
|
mitochondrial membrane potential
|
|
TPP+
|
tetraphenylphosphonium
|
|
ΔµH+
|
electrochemical gradient
|
|
ΔpHm
|
mitochondrial electrochemical
gradient
|
|
MMD
|
mitochondrial membrane
depolarization
|
|
MPT
|
mitochondrial permeability
transition
|
|
CsA
|
cyclosporin A
|
|
NADH
|
nicotinamide adenine dinucleotide
reduced form
|
|
NADPH
|
nicotinamide adenine dinucleotide
phosphate reduced form
|
|
DMO
|
5,5′-dimethyl-oxazolidine-2,4-dione
|
|
OPA
|
o-phthalaldehyde
|
|
GC-MS
|
gas chromatography-mass
spectrometry
|
|
TBDMS
|
tert-butyldimethylsilyl
|
|
MTBSTFA
|
N-tert-butyldimethylsilyl-N-methyltrifluoro-acetamide
|
|
TIC
|
total ion current
|
|
SIM
|
single ion monitoring
|
References
|
1
|
Ohkubo S, Dalla Via L, Grancara S,
Kanamori Y, García-Argáez AN, Canettieri G, Arcari P, Toninello A
and Agostinelli E: The antioxidant, aged garlic extract, exerts
cytotoxic effects on wild-type and multidrug-resistant human cancer
cells by altering mitochondrial permeability. Int J Oncol.
53:1257–1268. 2018.PubMed/NCBI
|
|
2
|
Fleischauer AT, Poole C and Arab L: Garlic
consumption and cancer prevention: Meta-analyses of colorectal and
stomach cancers. Am J Clin Nutr. 72:1047–1052. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Rahman K and Lowe GM: Garlic and
cardiovascular disease: A critical review. J Nutr. 136 (Suppl
3):736S–740S. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Banerjee SK and Maulik SK: Effect of
garlic on cardiovascular disorders: A review. Nutr J. 1:42002.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Sundaresan S and Subramanian P: Prevention
of N-nitrosodiethylamine-induced hepatocarcinogenesis by
S-allylcysteine. Mol Cell Biochem. 310:209–214. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Thomson M and Ali M: Garlic [Allium
sativum]: A review of its potential use as an anti-cancer
agent. Curr Cancer Drug Targets. 3:67–81. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ray B, Chauhan NB and Lahiri DK: Oxidative
insults to neurons and synapse are prevented by aged garlic extract
and S-allyl-L-cysteine treatment in the neuronal culture and APP-Tg
mouse model. J Neurochem. 117:388–402. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Amagase H, Petesch BL, Matsuura H, Kasuga
S and Itakura Y: Intake of garlic and its bioactive components. J
Nutr. 131:955S–962S. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Cemil B, Gokce EC, Kahveci R, Gokce A,
Aksoy N, Sargon MF, Erdogan B and Kosem B: Aged garlic extract
attenuates neuronal injury in a rat model of spinal cord
ischemia/reperfusion injury. J Med Food. 19:601–606. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Hu X, Cao BN, Hu G, He J, Yang DQ and Wan
YS: Attenuation of cell migration and induction of cell death by
aged garlic extract in rat sarcoma cells. Int J Mol Med. 9:641–643.
2002.PubMed/NCBI
|
|
11
|
Fallah-Rostami F, Tabari MA, Esfandiari B,
Aghajanzadeh H and Behzadi MY: Immunomodulatory activity of aged
garlic extract against implanted fibrosarcoma tumor in mice. N Am J
Med Sci. 5:207–212. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Li G, Qiao C, Lin R, Pinto J, Osborne M
and Tiwari R: Antiproliferative effects of garlic constituents in
cultured human breast-cancer cells. Oncol Rep. 2:787–791.
1995.PubMed/NCBI
|
|
13
|
Shirin H, Pinto JT, Kawabata Y, Soh JW,
Delohery T, Moss SF, Murty V, Rivlin RS, Holt PR and Weinstein IB:
Antiproliferative effects of S-allylmercaptocysteine on
colon cancer cells when tested alone or in combination with
sulindac sulfide. Cancer Res. 61:725–731. 2001.PubMed/NCBI
|
|
14
|
Hosono T, Fukao T, Ogihara J, Ito Y, Shiba
H, Seki T and Ariga T: Diallyl trisulfide suppresses the
proliferation and induces apoptosis of human colon cancer cells
through oxidative modification of beta-tubulin. J Biol Chem.
280:41487–41493. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Howard EW, Ling MT, Chua CW, Cheung HW,
Wang X and Wong YC: Garlic-derived S-allylmercaptocysteine is a
novel in vivo antimetastatic agent for androgen-independent
prostate cancer. Clin Cancer Res. 13:1847–1856. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Sriram N, Kalayarasan S, Ashokkumar P,
Sureshkumar A and Sudhandiran G: Diallyl sulfide induces apoptosis
in Colo 320 DM human colon cancer cells: Involvement of caspase-3,
NF-kappaB, and ERK-2. Mol Cell Biochem. 311:157–165. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lai KC, Kuo CL, Ho HC, Yang JS, Ma CY, Lu
HF, Huang HY, Chueh FS, Yu CC and Chung JG: Diallyl sulfide,
diallyl disulfide and diallyl trisulfide affect drug resistant gene
expression in colo 205 human colon cancer cells in vitro and in
vivo. Phytomedicine. 19:625–630. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yan JY, Tian FM, Hu WN, Zhang JH, Cai HF
and Li N: Apoptosis of human gastric cancer cells line SGC 7901
induced by garlicderived compound S-allylmercaptocysteine (SAMC).
Eur Rev Med Pharmacol Sci. 17:745–751. 2013.PubMed/NCBI
|
|
19
|
Zhang H, Wang K, Lin G and Zhao Z:
Antitumor mechanisms of S-allyl mercaptocysteine for breast cancer
therapy. BMC Complement Altern Med. 14:2702014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ng KT, Guo DY, Cheng Q, Geng W, Ling CC,
Li CX, Liu XB, Ma YY, Lo CM, Poon RT, et al: A garlic derivative,
S-allylcysteine (SAC), suppresses proliferation and metastasis of
hepatocellular carcinoma. PLoS One. 7:e316552012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Liu Z, Li M, Chen K, Yang J, Chen R, Wang
T, Liu J, Yang W and Ye Z: S-allylcysteine induces cell cycle
arrest and apoptosis in androgen-independent human prostate cancer
cells. Mol Med Rep. 5:439–443. 2012.PubMed/NCBI
|
|
22
|
Chung LY: The antioxidant properties of
garlic compounds: Allyl cysteine, alliin, allicin, and allyl
disulfide. J Med Food. 9:205–213. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
García E, Santana-Martínez R, Silva-Islas
CA, Colín-González AL, Galván-Arzate S, Heras Y, Maldonado PD,
Sotelo J and Santamaría A: S-allyl cysteine protects against
MPTP-induced striatal and nigral oxidative neurotoxicity in mice:
Participation of Nrf2. Free Radic Res. 48:159–167. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Xu YS, Feng JG, Zhang D, Zhang B, Luo M,
Su D and Lin NM: S-allylcysteine, a garlic derivative, suppresses
proliferation and induces apoptosis in human ovarian cancer cells
in vitro. Acta Pharmacol Sin. 35:267–274. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Compston A and Coles A: Multiple
sclerosis. Lancet. 372:1502–1517. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lassmann H: Pathophysiology of
inflammation and tissue injury in multiple sclerosis: What are the
targets for therapy. J Neurol Sci. 306:167–169. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zeinali H, Baluchnejadmojarad T, Fallah S,
Sedighi M, Moradi N and Roghani M: S-allyl cysteine improves
clinical and neuropathological features of experimental autoimmune
encephalomyelitis in C57BL/6 mice. Biomed Pharmacother. 97:557–563.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kyo E, Uda N, Kasuga S and Itakura Y:
Immunomodulatory effects of aged garlic extract. J Nutr.
131:1075S–1079S. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Morihara N, Sumioka I, Moriguchi T, Uda N
and Kyo E: Aged garlic extract enhances production of nitric oxide.
Life Sci. 71:509–517. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Imai J, Ide N, Nagae S, Moriguchi T,
Matsuura H and Itakura Y: Antioxidant and radical scavenging
effects of aged garlic extract and its constituents. Planta Med.
60:417–420. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Timeus F, Crescenzio N, Doria A, Foglia L,
Pagliano S, Ricotti E, Fagioli F, Tovo PA and Cordero di
Montezemolo L: In vitro anti-neuroblastoma activity of
saquinavir and its association with imatinib. Oncol Rep.
27:734–740. 2012.PubMed/NCBI
|
|
32
|
van Engeland M, Nieland LJ, Ramaekers FC,
Schutte B and Reutelingsperger CP: Annexin V-affinity assay: A
review on an apoptosis detection system based on phosphatidylserine
exposure. Cytometry. 31:1–9. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Nicoletti I, Migliorati G, Pagliacci MC,
Grignani F and Riccardi C: A rapid and simple method for measuring
thymocyte apoptosis by propidium iodide staining and flow
cytometry. J Immunol Methods. 139:271–279. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Jiménez-Martín E, Ruiz J, Pérez-Palacios
T, Silva A and Antequera T: Gas chromatography-mass spectrometry
method for the determination of free amino acids as their
dimethyl-tert-butylsilyl (TBDMS) derivatives in animal source food.
J Agric Food Chem. 60:2456–2463. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Frezza C, Cipolat S and Scorrano L:
Organelle isolation: Functional mitochondria from mouse liver,
muscle and cultured fibroblasts. Nat Protoc. 2:287–295. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Gornall AG, Bardawill CJ and David MM:
Determination of serum proteins by means of the biuret reaction. J
Biol Chem. 177:751–766. 1949.PubMed/NCBI
|
|
37
|
Kamo N, Muratsugu M, Hongoh R and Kobatake
Y: Membrane potential of mitochondria measured with an electrode
sensitive to tetraphenyl phosphonium and relationship between
proton electrochemical potential and phosphorylation potential in
steady state. J Membr Biol. 49:105–121. 1979. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Rottenberg H: Non-equilibrium
thermodynamics of energy conversion in bioenergetics. Biochim
Biophys Acta. 549:225–253. 1979. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Tietze F: Enzymic method for quantitative
determination of nanogram amounts of total and oxidized
glutathione: Applications to mammalian blood and other tissues.
Anal Biochem. 27:502–522. 1969. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Kanda Y: Investigation of the freely
available easy-to-use software ‘EZR’ for medical statistics. Bone
Marrow Transplant. 48:452–458. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Ho JN, Kang M, Lee S, Oh JJ, Hong SK, Lee
SE and Byun SS: Anticancer effect of S-allyl-L-cysteine via
induction of apoptosis in human bladder cancer cells. Oncol Lett.
15:623–629. 2018.PubMed/NCBI
|
|
42
|
Hoehner JC, Gestblom C, Hedborg F,
Sandstedt B, Olsen L and Påhlman S: A developmental model of
neuroblastoma: Differentiating stromapoor tumors' progress along an
extra-adrenal chromaffin lineage. Lab Invest. 75:659–675.
1996.PubMed/NCBI
|
|
43
|
Huang M and Weiss WA: Neuroblastoma and
MYCN. Cold Spring Harb Perspect Med. 3:a0144152013. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Fonseka P, Liem M, Ozcitti C, Adda CG, Ang
CS and Mathivanan S: Exosomes from N-Myc amplified neuroblastoma
cells induce migration and confer chemoresistance to non-N-Myc
amplified cells: Implications of intratumour heterogeneity. J
Extracell Vesicles. 8:15976142019. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Lawson LD: Garlic: A review of its
medicinal effects and indicated active compounds. Phytomedicines of
Europe: Chemistry and biological activity. Lawson LD and Bauer R;
ACS symposium series, : 691. American Chemical Society; Washington,
DC, USA: pp. 176–209. 1998, View Article : Google Scholar
|
|
46
|
Kodera Y, Suzuki A, Imada O, Kasuga S,
Sumioka I, Kanezawa A, Taru N, Fujikawa M, Nagae S, Masamoto K, et
al: Physical, chemical, and biological properties of
S-allylcysteine, an amino acid derived from garlic. J Agric Food
Chem. 50:622–632. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Colín-González AL, Santana RA, Silva-Islas
CA, Chánez-Cárdenas ME, Santamaría A and Maldonado PD: The
antioxidant mechanisms underlying the aged garlic extract- and
S-allylcysteine-induced protection. Oxid Med Cell Longev.
2012:9071622012. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Maldonado PD, Alvarez-Idaboy JR,
Aguilar-González A, Lira-Rocha A, Jung-Cook H, Medina-Campos OM,
Pedraza-Chaverri J and Galano A: Role of allyl group in the
hydroxyl and peroxylradical scavenging activity of S-allylcysteine.
J Phys Chem B. 115:13408–13417. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Dairam A, Fogel R, Daya S and Limson JL:
Antioxidant and iron-binding properties of curcumin, capsaicin, and
S-allylcysteine reduce oxidative stress in rat brain homogenate. J
Agric Food Chem. 56:3350–3356. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Pinto JT, Qiao C, Xing J, Rivlin RS,
Protomastro ML, Weisler ML, Tao Y, Thaler H and Heston WD: Effects
of garlic thioallyl derivatives on growth, glutathione
concentration, and polyamine formation of human prostate carcinoma
cells in culture. Am J Clin Nutr. 66:398–405. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Kim KM, Chun SB, Koo MS, Choi WJ, Kim TW,
Kwon YG, Chung HT, Billiar TR and Kim YM: Differential regulation
of NO availability from macrophages and endothelial cells by the
garlic component S-allyl cysteine. Free Radic Biol Med. 30:747–756.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Kim SR, Jung YR, An HJ, Kim DH, Jang EJ,
Choi YJ, Moon KM, Park MH, Park CH, Chung KW, et al: Anti-wrinkle
and anti-inflammatory effects of active garlic components and the
inhibition of MMPs via NF-κB signaling. PLoS One. 8:e738772013.
View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Mong MC and Yin MC: Nuclear factor
κB-dependent anti-inflammatory effects of S-allyl cysteine and
S-propyl cysteine in kidney of diabetic mice. J Agric Food Chem.
60:3158–3165. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Grancara S, Zonta F, Ohkubo S, Brunati AM,
Agostinelli E and Toninello A: Pathophysiological implications of
mitochondrial oxidative stress mediated by mitochondriotropic
agents and polyamines: The role of tyrosine phosphorylation. Amino
Acids. 47:869–883. 2015. View Article : Google Scholar : PubMed/NCBI
|