Introduction
Acute myocardial infarction (AMI), commonly known as
heart attack, results in ischemic damage of cardiomyocytes, which,
by activation of a damage-associated molecular pattern, leads to
initiation of the innate immune response (1,2).
Inflammatory cells infiltrate to the injured heart, ensuring the
clearance of harmful cell debris and repair of the damaged area via
formation of a fibrotic scar (3) and
activation of the endogenous stem cell pool (4). While the default immune response to AMI
appears to ensure a quick fix of the heart, the scarring leads to
pathological remodeling of the heart and compromised cardiac
function over time. Therefore, timely balanced transformation from
pro-inflammatory in the acute stage to an anti-inflammatory state
is rather critical to ensure a reparative course after AMI
(5).
Monocytes are cornerstones of local inflammatory
response that are causatively involved in numerous inflammatory
diseases and tissue homeostasis (6).
Monocytes are recruited to the site of necrosis where they
differentiate into distinct subsets of tissue macrophages (M1/M2)
and function synergistically, but at times counteractively, with
other cellular components in a reparative process, orchestrated
mainly by the local milieu and cytokine profile (7,8).
Although circulating monocytes develop from a single, common
precursor, heterogeneity of the monocyte population has been widely
acknowledged based on the differential expression of the
lipopolysaccharide (LPS) receptor (CD14) and the FccII I receptor
(CD16), with cell subsets being the classical
(CD14++/CD16−), non-classical
(CD14+/CD16++) and intermediate
(CD14++/CD16+) populations (9). Each subset has different immune
functions, phagocytic activity, cytokine profile and reparative
capacity (10). Classical monocytes
express high levels of chemokine receptor 2 (CCR2) and are highly
responsive to LPS stimulation; they produce a broad range of
cytokines and chemokines with high myeloperoxidase and phagocytic
activity, and are thus associated with the severity of inflammatory
response and worse clinical outcome in patients with AMI (11). By contrast, non-classical subsets are
distinguished by high levels of complement C1q tumor necrosis
factor (TNF)-related protein-3 (CTRP3), have numerous patrolling
properties and are therefore thought to be involved in the innate
surveillance of tissues (12). They
may be derived from classical monocytes and have a lifespan of
several days in humans. Non-classical monocytes contribute to the
formation of granulation tissue and exert an array of similarities
to tissue macrophages; they are thus considered to be more mature
than classical monocytes and frequently exert anti-inflammatory and
pro-homeostatic effects (13).
The intermediate subpopulation, although it has the
smallest proportion and appears to only display an intermediate
expression of surface markers, it is a clearly distinguishable
subset and has a distinct gene expression profile and functionality
(14). Intermediate monocytes
selectively express C-C chemokine receptor type 5 (CCR5), which,
upon activation, secretes numerous inflammatory cytokines,
including interleukins (ILs) and TNF-α (5). The proportion of the inflammatory
monocytes in peripheral blood was determined to be elevated in
numerous pathogeneses of cardiovascular diseases and diabetes
(15,16), and has been used as a predictor of
prognosis for patients with cardiovascular disease (CVD) (17); however, the mechanistic association
of the intermediate monocyte subset with the severity of CVD
remains elusive (18).
Given the growing body of evidence on the functional
complexity of different monocyte subsets, modulation of the
inflammatory response by targeting the monocyte sub-population has
conceptually emerged as a novel intervention that supports
endogenous tissue homeostasis in patients with AMI (19). Kinetic studies have provided
circumstantial evidence for this monocyte conversion during the
course of an inflammatory response, which occurred in the
peripheral blood and locally at the site of injury (20). This theory is further supported by
the fact that the level of transcription of genes associated with
maturation progressively increases from classical monocytes via
intermediate to non-classical monocytes (13). Phenotypic switch of monocyte subsets
is known to be regulated by numerous factors. For, instance, in
vitro, CD34+ hematopoietic stem cells first
differentiate into CD14++CD16− monocytes,
which, following subsequent culture, express CD16, suggesting that
a phenotypic transition of the monocyte sub-population may occur
under specific conditions (21).
In the present study, the potential of an adipokine
to limit the excessive inflammatory response was explored by
inducing an intermediate switch of monocytes. The CTRP superfamily
is a cluster of adipokines and 15 isotypes of proteins in humans
have been recently identified (22).
CTRP3 is ubiquitously expressed in chondrocytes and adipose tissue
and is able to improve insulin sensitivity as well as promote
glucose and lipid metabolism (23).
CTRP3 also serves as a potent anti-inflammatory adipokine; it may
be beneficial in the prevention of cardiovascular diseases and
provide a promising therapeutic strategy to attenuate vascular
remodeling (24). In this context,
the present study demonstrated that CTRP3 induced an intermediate
switch of monocyte subsets and antagonized pro-inflammatory
cytokine expression in monocytes, providing a ‘proof-of-concept’
evidence of using CTRP3 to limit inflammatory response in patients
following AMI.
Materials and methods
Sample collection
Blood samples (10 ml) were collected from the
patients during primary hospitalization (3–5 h following AMI) and
follow-up (3 and 7 days) at the Department of Cardiology of the
People's Hospital of Danyang (Danyang, China) between November 2018
and February 2019, as well as from young staff members (age range,
26–53 years) within the department who volunteered for this study.
All patients were angiographically proven to have ST-elevated AMI
and all physical parameters are summarized in Table I. A total of 12 patients with AMI and
8 healthy volunteers were included in the present study. Diagnosed
inflammatory diseases within the last three months were the only
exclusion criterion for all volunteers. The study was approved by
the Ethics Committee of People's Hospital of Danyang (Danyang,
China) and was performed in accordance with the Declaration of
Helsinki. All participants or accompanying relatives provided
written consent prior to the obtainment of peripheral blood
samples, and they all agreed to participate in the present
experimental study regarding the isolation of monocytes for medical
research (according to the Declaration of Helsinki).
 | Table I.Clinicopathological parameters of
subjects included in the present study. |
Table I.
Clinicopathological parameters of
subjects included in the present study.
| Parameter | AMI (n=12) | Volunteers
(n=8) | Reference
range | P-value |
|---|
| Age (years) |
67.31±24.1 |
31.35±14.2 | – | 0.007 |
| Sex (M/F) |
7/5 |
5/3 | – | – |
| Diastolic blood
pressure (mmHg) |
91.28±8.1 |
81.31±4.8 | 80–89 | 0.006 |
| Systolic blood
pressure (mmHg) | 147.26±34.6 |
121.31±8.2 | 120–139 | 0.053 |
| WBC count
(cells/µl) | 11,917±2212 | 10,191.31±1812 |
4,000-11,000 | 0.084 |
| PBMC count
(cells/µl) |
767±212 |
512±117 |
500-3,000 | 0.006 |
| Sub-category of
circulating monocytes (% of total monocytes)a |
|
|
|
|
| Classical |
91.31±4.12 |
94.66±3.44 | 86–95 | 0.07 |
| Non-classical |
2.58±0.31 |
2.85±0.38 | 2–4 | 0.098 |
| Intermediate |
6.10±1.21 |
2.49±0.13 |
3–10 | 0.0001 |
Isolation, cultivation and analysis of
monocyte subsets
Peripheral blood mononuclear cells (PBMCs) were
isolated by Ficoll density gradient centrifugation (1,000 × g for
30 min) at 4°C. The mononuclear fraction (indicated by the red
arrow in Fig. 1) was washed with
saline and RPMI-1640 medium (Sigma-Aldrich; Merck KGaA), suspended
in autologous serum (AS) and counted. The samples were sequentially
subjected to either flow cytometric analysis or cultivation as
schematically illustrated in Fig. 1.
For subset analysis, the cell fraction was resuspended in MACS
buffer [2% fetal bovine serum (FBS; Gibco; Thermo Fisher
Scientific, Inc.) in PBS with 1 mM EDTA] and stained with CD14 and
CD16 monoclonal antibodies using the methodology described in
detail below. For pharmacological experiments, the isolated cells
were inoculated into a 10-cm petri dish containing Dulbecco's
modified Eagle's medium (DMEM; Sigma-Aldrich; Merck KGaA) culture
medium supplemented with 30% AS, penicillin (100 U/ml),
streptomycin (0.1 mg/ml) and glutamine (2 mM) and incubated at 37°C
with 5% CO2 for treated with CTRP3 for 24 h prior to
extraction of total RNA for reverse transcription-quantitative
(RT-q)PCR.
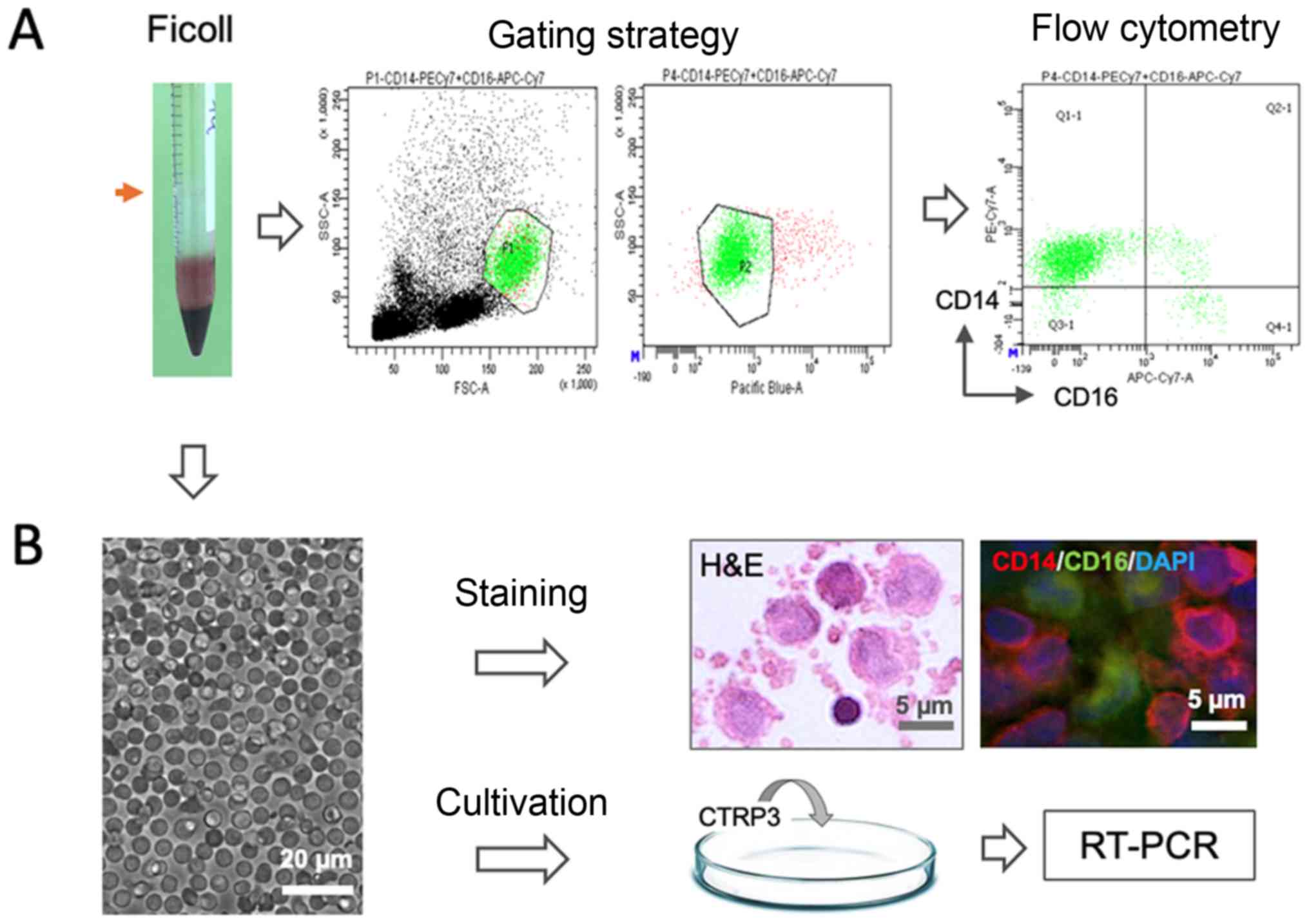 | Figure 1.Schematic illustration of
experimental design. Mononuclear cells (red arrow) were isolated by
Ficoll gradient centrifugation from peripheral blood sample and
were (A) stained with CD14 and CD16 antibodies for flow cytometric
analysis and (B) cultivated for pharmacological testing of CTRP3,
histological H&E staining and immunostaining for CD14 and CD16
(scale bar, 5 or 20 µm). PE, phocoerythrin; APC, allophycocyanine;
Cy, cyanine; FSC, forward scatter; SSC, side scatter; Q, quadrant;
RT-PCR, reverse transcription PCR; CTRP3, complement C1q tumor
necrosis factor-related protein-3. |
Flow cytometric analysis
For immunostaining, cells were diluted at a
concentration of 2×105 in 100 µl MACS buffer (2% FBS in
PBS with 1 mM EDTA) and stained with the following
fluorescence-conjugated antibodies: Phycoerythrin-cyanine
(Cy)7-conjugated anti-human CD14 (clone 63D3; cat. no. 367112;
1:200 dilution; Biolegend) and allophycocyanine-Cy7-conjugated
anti-human CD16 (clone B73.1; cat. no. 360710; 1:200 dilution;
Biolegend) for 10 min at 4°C in the dark. Thereafter, cell
suspensions were washed twice by using MACS buffer and stained with
DAPI (1:250 dilution) for 30 sec. In the gating strategy,
subpopulations of leukocytes (monocytes and lymphocytes) were
identified by distinguishing their locations on the forward and
side scatter scale, and the duplicates and dead cells were excluded
in the channel of pacific blue based on DAPI staining. Monocyte
subsets were discriminated based on the intensity/presence of
expression of CD14 and CD16, namely, classical
(CD14++/CD16−), non-classical
(CD14+/CD16++) and intermediate populations
(CD14++/CD16+). All samples were analyzed
using a flow cytometer (FACSCanto II; BD Biosciences) and the data
were stored and analyzed using Flowjo 7.6 software (Tree Star,
Inc.).
Assessment of phagocytic and migratory
activity
To compare the biological nature of monocytes from
two cohorts of subjects (healthy, n=8 vs. AMI on day 3, n=12), two
complementary approaches were used to assess the phagocytic
activity. The cells isolated by Ficoll gradient centrifugation were
seeded into 6-well plates [1×105 in 2 ml DMEM
supplemented with 10% FBS, penicillin (100 U/ml), streptomycin (0.1
mg/ml) and glutamine (2 mM)] and incubated at 37°C for 30 min. In
the first part, FITC-labeled Zymosan (Thermo Fisher Scientific,
Inc.) was used as a reference tracer to mimic infectious
conditions. Zymosan was added into the culture medium at the final
concentration of 200 µg/ml and plates were again placed at 37°C
under gentle shaking (60 rpm) for an additional 1 h. Thereafter,
cells were collected and washed 3 times with PBS and stained again
with antibodies against CD14 and CD16, as described above. For flow
cytometric analysis of particle uptake, the mean fluorescence
intensity (MFI) in the FITC channel (wavelength, 495 nm) was
measured as an indicator for the amount of ingested Zymosan
particles. In the second part, 19F-perfluorocarbon (PFC)
nanoparticles were used as a sterile inflammatory tracer as
previously described (25). In
brief, 100 µl (10% in PBS) was added to each well of a 6-well plate
and incubation was maintained at 37°C under gentle shaking (60 rpm)
for 60 min, following which cells were collected. After several
washing steps, the cell pellet was fixed with 4% paraformaldehyde
and the 19F content was measured by using an NMR
spectrometer (9.4 Tesla; Bruker) at frequencies of 400.13 MHz for
1H and 376.46 MHz for 19F detection. For
superimposing the images of the same nuclei, the ‘hot iron’ color
lookup table (ParaVision; Bruker) was applied to 19F
images and the intensity of 19F was considered as a
phagocytic tracer in the comparison of the activity in different
subpopulations from individual subjects.
Migratory analysis was performed using a commercial
migration assay in a 12-well plate with a pore size of 3 µm (Thin
Cert™; Austria). In brief, 500,000 monocytes were transferred to
the upper layer of a Thin Cert™ set in 10 ml culture medium and the
lower compartment contained 2 ml DMEM with 10% FBS supplemented
with monocyte chemoattractant protein 1 (MCP-1) at a concentration
of 20 ng/ml to facilitate the migratory process of monocytes. The
migratory unit was placed in an incubator at 37°C for 3 h and
subsequently, the entire medium of the lower compartment was
collected. Migrated cells were quantitatively counted via flow
cytometry (FACSCanto II; BD Biosciences) using CountBright Absolute
counting beads (Thermo Fisher Scientific, Inc.).
Pharmacological interventions
In the present series of experiments, to minimize
potential skewing of epitopes and spontaneous differentiation into
a macrophage phenotype of cultured monocytes, FBS was replaced by
AS of individual subjects in the culture protocol. In doing so,
monocytes from patients 3 days after AMI (3D-AMI) were isolated and
cultivated under similar conditions as above, except that 10% FBS
was changed to 30% AS, and under these conditions, the proportions
of the three phenotypic subsets of monocytes, as well as the
expression profile of cytokines (IL-6 and TNF-α), were unaffected
after 24 h in culture. To test the induction effects of CTRP3
(Aviscera Bioscience) on phenotypic switch of monocytes that
display anti-inflammatory properties, recombinant CTRP3 was added
into the culture medium at a final concentration of 1 µg/ml and the
subsets of monocytes were analyzed by flow cytometry, as specified
above. To explore potential downstream cascades mediating this
biological effect, the cells were first challenged with LPS (100
ng/ml; Sigma-Aldrich; Merck KGaA) and a p38 mitogen-activated
protein kinase (MAPK)/ERK inhibitor, SB203580 (1 µM;
MedChemExpress), was optionally added prior to treatment with
CTRP3. All chemicals were dissolved in DMSO and added into the
medium at a ratio of 1:1,000. The wells supplemented with 0.1% of
DMSO were applied as DMSO control. In all interventions, cells were
cultivated at 37°C with 5% CO2 for a period of 24 h
prior to the extraction of total RNA was for RT-qPCR.
RT-qPCR
RT-qPCR was performed to determine the mRNA
expression levels of IL-6 and TNF-α following the pharmacological
treatments. Total RNA of monocytes was isolated using the RNeasy
Micro Kit (Qiagen) and complementary DNA was synthesized using the
QuantiTect Reverse Transcription Kit (Qiagen) according to the
manufacturer's protocol and as previously described (8). The thermocycling conditions were as
follows: Initial denaturation for 2 min at 50°C, and 40 cycles of
10 min at 95°C and 1.5 min at 60°C. Amplification efficiencies were
calculated from the linear part of the quantification curve. Gene
expression was normalized to GAPDH and calculated using the
2−∆∆Cq method (8).
Pre-designed Taqman commercial primers (IL-6: Hs00174131_m1; TNF-α,
Hs02621508_s1; GAPDH: Hs02786624_g1) were purchased from Thermo
Fisher Scientific, Inc. and their efficiency was confirmed in all
amplification plots. Quantification of gene expression was
performed using a StepOnePlus Real-time PCR system (Thermo Fisher
Scientific, Inc) following the manufacturer's protocol and all PCR
assays were performed in duplicate.
Statistical analysis
Values are expressed as the mean ± standard
deviation. Differences in sub-populations of monocytes over time,
migratory activity and the phagocytic activity of FITC-Zymosan
within different monocyte subsets were analyzed using two-way
repeated-measures analysis of variance (ANOVA). The expression
levels of IL-6 and TNF-α were analyzed using one-way ANOVA. Tukey's
multiple-comparisons test was used as a post hoc analysis to reveal
significant differences between the groups. An unpaired Student's
t-test was performed to analyze the phagocytic activity of
19F-nanoparticles in the comparison of healthy vs.
3D-AMI monocytes. Statistical analysis was performed using GraphPad
Prism© (version 7; GraphPad Software Inc.). P<0.05
was considered to indicate a statistically significant
difference.
Results
Subjects
In total, 12 AMI patients (age, 67.31±24.1 years;
male/female, 7/5) and 8 volunteers (age, 31.35±14.2 years;
male/female, 5/3) were included in the present study. Physical data
of all subjects included in the present study are summarized in
Table I.
Dynamic changes of circulating
monocyte subsets after AMI
Peripheral whole blood samples were obtained
directly after the angiographic exam (usually 3–5 h after
hospitalization), as well as 3 and 7 days after the onset of AMI
and subjected to flow cytometric analysis. A total of three
distinct monocyte subpopulations were defined according to CD14 and
CD16 expression in the cytometric plots (Fig. 2A), with the classical subset
accounting for the majority of cells in healthy volunteers and
patients with AMI (Table I).
Although the relative percentage of the subsets in patients with
AMI at the early stage was similar to that in the healthy controls,
the absolute cell counts were already significantly increased,
suggesting that recruitment of monocytes occurred as early as a few
hours after the onset of MI. Thereafter, the total monocytes and
classical subset further increased, peaking on day 3 after the
onset of MI, and declined to a level close to that in the healthy
controls on day 7 (Fig. 2B). The
non-classical and intermediate subset increased more vigorously
than the classical population and remained significantly increased
at the end of the observation period of 7 days (Fig. 2B). Taken together, the time-course
study indicates dynamical recruitment of monocyte subsets that is
required to orchestrate the reparative process of the damaged heart
tissue.
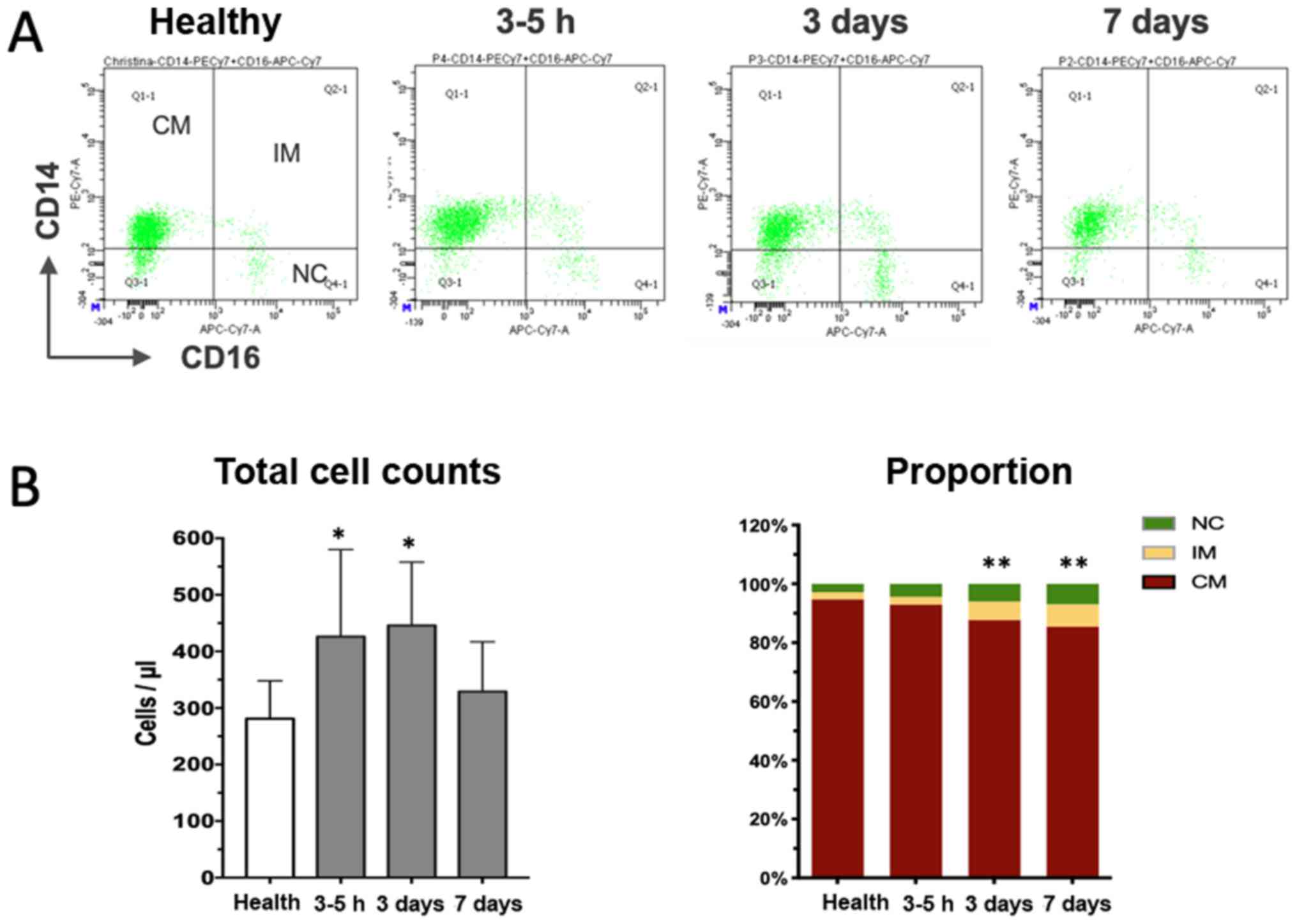 | Figure 2.Dynamical changes of monocyte subsets
after myocardial infarction. (A) Representative illustration of
three monocyte subsets according to CD14 and CD16 staining at
different time-points. (B) Quantitative analysis indicated that the
total cell counts initially increased (3–5 h) and then declined
almost to the level of healthy controls (left panel in B). The
right panel in B displays the proportional changes of three subsets
of monocytes over time, revealing the progressive increase of the
percentage of the IM subpopulation. *P<0.05, total cell counts
vs. healthy; **P<0.01, proportion of IM vs. healthy. CM,
classical monocytes; NC, non-classical subsets; IM, intermediate
subpopulation; PE, phycoerythrin; APC, allophycocyanine; Cy,
cyanine; FSC, forward scatter; SSC, side scatter; Q, quadrant. |
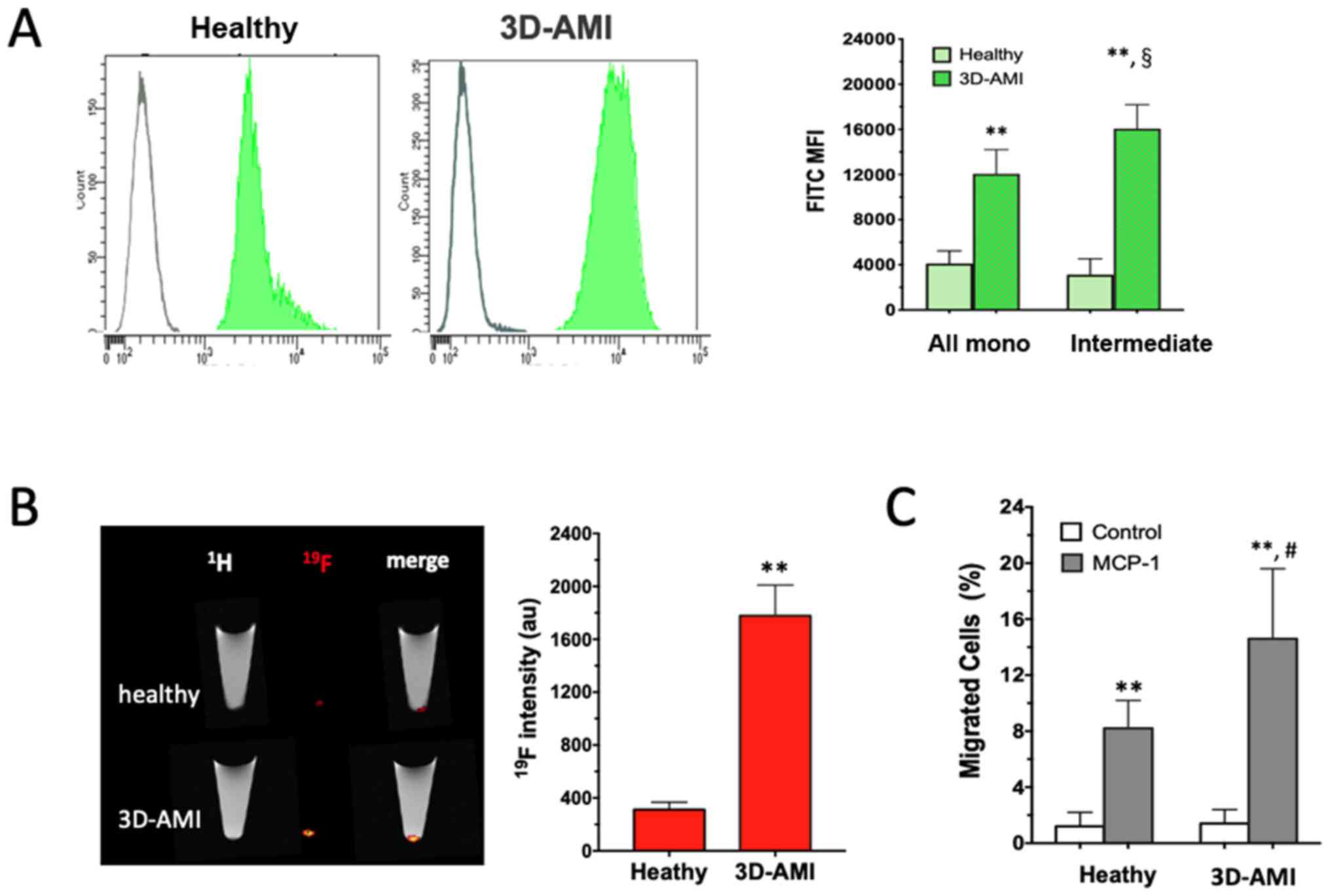
Enhanced phagocytic and migratory
activity of monocytes in patients with AMI
The functional enhancement of the circulating
monocytes in patients with AMI was further analyzed. In this
attempt, two complementary approaches were used. When FITC-Zymosan
was added to the culture medium of the isolated monocytes, they
exhibited strong phagocytic activity, as shown by the right-shifted
MFI in Fig. 3A. In comparison to the
healthy control, the monocytes derived from patients in the 3D-AMI
group exhibited more vigorous phagocytic activity in response to
Zymosan in all monocyte subsets, which was even more pronounced in
the intermediate sub-population (P<0.01). In addition,
19F-containing nanoparticles were used as an alternative
phagocytic tracer to compare monocytes from either healthy
volunteers or patients with AMI. After 60 min of co-cultivation,
the 19F content in the cell pellet, which represents the
ingested nanoparticles, was analyzed by 19F-MRI
detection, which consistently indicated a ~5-fold increase in
19F intensity in the AMI-derived monocytes (P<0.01;
Fig. 3B). Therefore, the circulating
monocytes in patients with AMI, likely triggered by the
inflammatory milieu after AMI, reached a state with enhanced
phagocytic activity.
Next, cell migration was examined as an additional
function of monocytes by using a commercial Transwell assay (Thim
Cert™). When primed with chemotactic MCP-1 in the lower chamber,
monocytes mobilizing from the upper layer into the lower chamber
were observed in the healthy as well as the 3D-AMI group. Of note,
the monocytes from the 3D-AMI patients migrated more vigorously,
with the proportion of monocytes moving across the membrane
reaching 15±5.0% in comparison to only 8±2.0% in the healthy
control group (P<0.01; Fig. 3C).
Taken together, circulating monocytes in patients with AMI not only
increased in quantity but were also induced to acquire an increased
function, as evidenced by their elevated phagocytic and migratory
activity.
CTRP3 induced a phenotypic switch of
monocytes to acquire anti-inflammatory properties
Given that an ‘uncontrolled’ excessive inflammatory
response may cause secondary cardiac damage in patients with AMI
and that dynamic changes of monocyte subsets may provide a novel
therapeutic target in the treatment of AMI, the anti-inflammatory
properties of a newly discovered adipokine, CTRP3, on monocyte
functionality were then tested in vitro. When monocytes from
patients in the 3D-AMI group were isolated and cultivated with 10%
FBS, a significant spontaneous differentiation into macrophages was
noticed over 24 h (24 h dark grey in Fig. 4A), but the phenomenon was
substantially minimized and no phenotypic changes were observed
(DMSO control in Fig. 4B) when FBS
was replaced by AS, confirming the stability of in vitro
monocyte cultivation for pharmacological interventions (Fig. 4A). When CTRP3 was added to the
medium, the proportion of the classical
(CD14++CD16−), non-classical
(CD14+CD16++) and intermediate
(CD14++CD16+) subset was analyzed by either
immunostaining or flow cytometry. The results indicated that all
three subsets did not change in the first 2 h, but that the
percentage of the intermediate population was elevated to 6.8±1.1%
after 24 h of culture in comparison to 4.1±1.2% in the DMSO control
(P<0.05; Fig. 4B), suggesting
that CTRP3 is able to induce a phenotypic switch among monocyte
subpopulations.
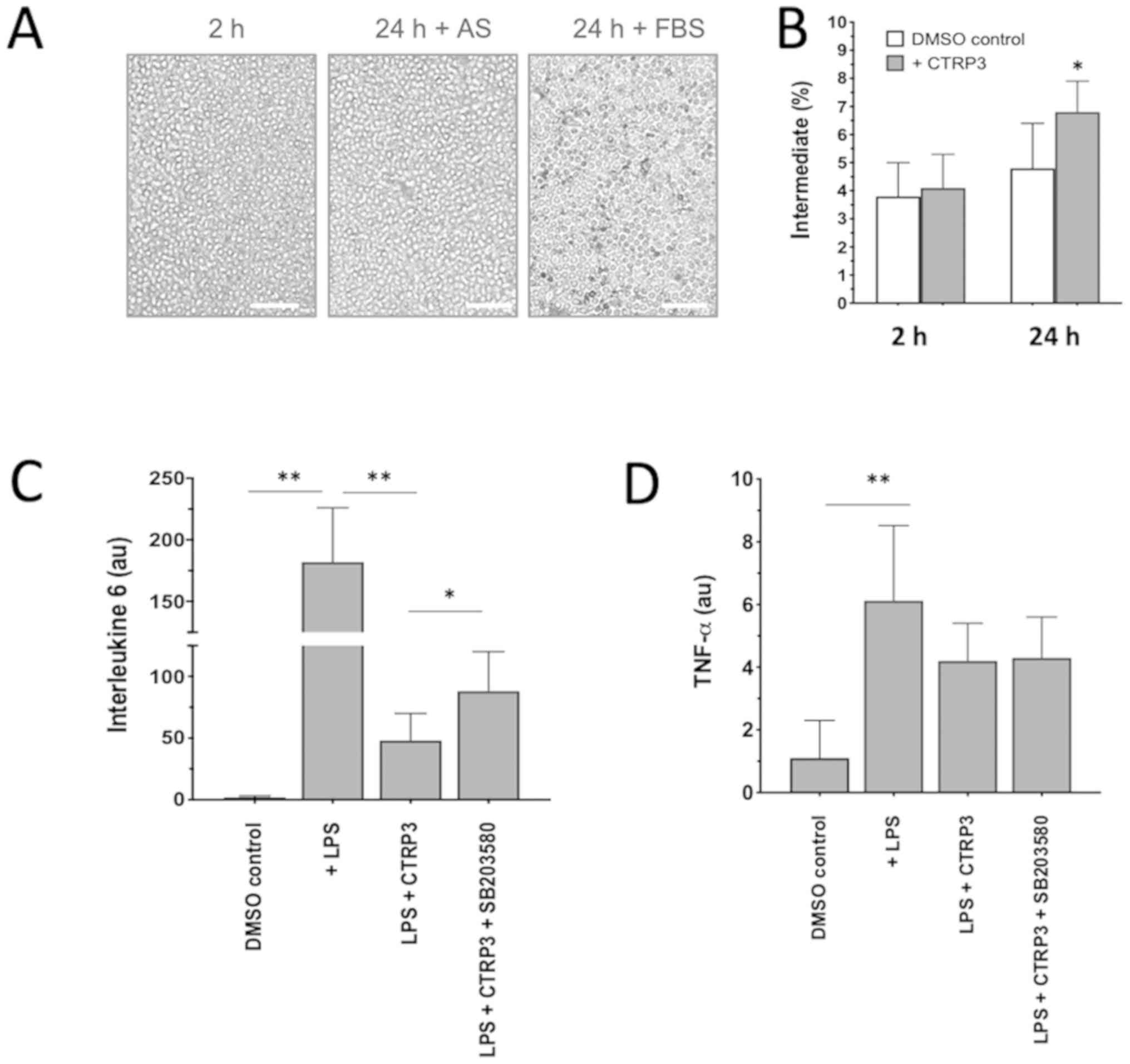 | Figure 4.Anti-inflammatory effect of CTRP3 on
phenotypic switch and cytokines. (A) When isolated monocytes were
cultivated ex vivo with 30% AS for 24 h, they were
morphologically similar to cells after 2 h cultivation with 10%
FBS. Incubation with 10% FBS for 24 h appeared to have induced
spontaneous macrophage differentiation, as characterized by the
attached, elongated morphology. Scale bars, 50 µm. (B) Addition of
CTRP3 induced a significant enrichment of the intermediate subset
of cultivated monocytes. (C and D) LPS-induced stress stimulated
expression of (C) interleukin 6 and (D) TNF-α, which was blunted in
the presence of CTRP3, highlighting the anti-inflammatory effects
of CTRP3 on monocytes. Of note, the inhibitory effect on IL-6 but
not TNF-α was partially counteracted when SB203580, a p38
mitogen-activated protein kinase/ERK inhibitor, was added.
*P<0.05 and **P<0.01. CTRP3, complement C1q TNF-related
protein-3; AS, autologous serum; TNF, tumor necrosis factor; FBS,
fetal bovine serum; LPS, lipopolysaccharides. |
To demonstrate the effects of CTRP3 on the secretory
profiles of monocytes, two representative cytokines, IL-6 and TNF-α
were examined as major pro-inflammatory cytokines derived from
activated monocytes (26). When
monocytes were stressed by LPS, transcription of IL-6 increased by
>100-fold in comparison with that in the DMSO control
(P<0.01; Fig. 4C), while TNF-α
was only moderately induced (P<0.05; Fig. 4D), indicating that individual
cytokines may react diversely in response to LPS stimulation. Of
note, the LPS-induced IL-6 activation was largely antagonized in
the presence of CTRP3 and partially restored when the activity of
the p38 MAPK/ERK cascade was blocked by the addition of SB203580 to
the culture medium (Fig. 4C).
SB203580 is a pyridinyl imidazole inhibitor widely used to
investigate the roles of p38 MAPK. The present results highlight
the inhibitory activity of CTRP3 on the response to LPS challenge
in monocytes populations upon activation of the p38 MAPK/ERK
cascade. On the other hand, CTRP3 exhibited a less pronounced
effect on LPS-induced TNF-α activation in the present experimental
setting (Fig. 4D).
Discussion
The present study revealed a sequential mobilization
and phenotypical interchange of monocyte subpopulations from the
onset of AMI to the sub-acute phase (7 days) with enhanced
phagocytic and migratory activities, reflecting the salvaging
requirement and severity of the systemic inflammatory response to
tissue damage. The dynamic heterogeneity of circulating monocytes
and their differential cell fates following tissue infiltration
pinpoints monocytes as a novel therapeutic target, which, by
pharmacological manipulation, may augment the cardiac reparative
process in patients following AMI. In this attempt, it was further
demonstrated that CTRP3, a member of the CTRP family, induced an
intermediate switch to the CD14++CD16+
monocyte subpopulation and antagonized the LPS-induced expression
of pro-inflammatory cytokines, suggesting that CTRP3 may represent
an alternative candidate in preventing damage arising from
excessive inflammatory responses by targeting circulating
monocytes.
Cardiac ischemic insult evokes a complex, temporally
and spatially well-coordinated inflammatory response at the damaged
parts of the tissue where immune cells are sequentially recruited
and functionally orchestrated (27).
Among all infiltrating immune cells, monocytes as a part of the
innate immune system are mobilized and appear concomitantly at
different stages of the inflammatory response to pursue distinct
functions in specific ways (5). In
mice after myocardial infarction, Ly-6Chi monocytes,
corresponding to CD14++CD16− monocytes in
humans, accumulate via CCR2, predominate at the site of injury
during the first 3 days and scavenge necrotic debris by a
combination of expression of inflammatory mediators, proteolysis
and phagocytosis, while Ly-6Clo monocytes, corresponding
to CD14+CD16++ monocytes in humans,
infiltrate preferentially via a CX3C chemokine receptor 1-mediated
pathway to initiate a reparative process (5,27). This
paradigm was demonstrated in the present study, as a few hours
after infarction, the total monocyte counts (mainly classical
subsets) in AMI patients expanded and were elevated as early as 3–5
h and retained for up to 3 days before starting to decline, whilst
a persistent increase in intermediate and non-classical subsets at
3 and 7 days was observed, suggesting that the mobilization and
recruitment of monocytes is a sequentially fine-tuned process upon
the salvaging requirement for the maintenance of tissue
homeostasis.
Of note, the peak level of the cell counts of
classical monocytes (CD14++CD16−) has been
proved to be linked to the impairment of myocardial salvage in the
acute phase after AMI and adverse left ventricular remodeling,
suggesting excessive mobilization of circulating monocytes is
likely to be harmful to the initiation of the reparative course
(11,28). The undue response is frequently
associated with the enhanced innate immunity as evidenced by the
phagocytic and migratory activities of the activated monocytes,
mainly occurring in the intermediate subset (29). In the present study, two types of
particle sizes and tracers were employed: FITC-Zymosan has a
relatively big size (1 µm in average dimension) and represents
infectious particles (produced from yeast particles known to be
phagocytosed by various types of cell, including
monocytes/macrophages) (30) and
19F-containing PFC is a small-particle size (100 nm)
nanoemulsion that has been used as an experimental tracer of
monocytes homing to the site of inflammation (25). The two independent approaches
suggested that monocytes from patients with AMI exhibited an
enhanced phagocytic activity, indicating that the monocytes were
systemically tuned to a highly active state in response to
infarction. Of note, the patient-derived monocytes more efficiently
ingested the small-size particles (100 nm) than the big-size
particles (1 µm), although the biological relevance of this
phenomenon requires to be further investigated.
While phagocytic activity and production of a myriad
of cytokines are necessary to ensure the clearance of cell debris
in the damaged tissue, a timely switch of immune response and
reduction of excessive inflammation are important to maintain
tissue homeostasis (6). The nature
of fluctuations observed in the transient increase of cell counts
of monocyte subsets and persistent elevation of non-classical and
intermediate cells points at a time-dependent change of monocyte
counts in patients after AMI, which opens a possibility of
manipulating CD14++CD16− monocytes as a novel
therapeutic target for salvaging ischemic damage (19,11). In
this attempt, the potential of CTRP3 as an immune modulator to
induce phenotypic switch of monocyte subsets and to balance the
immune response was tested. First, a culture system with individual
AS was established, in which monocytes maintained their phenotypic
stability for as long as 24 h in in vitro culture. It was
also demonstrated that CTRP3 treatment caused a skew of cultured
monocytes towards the intermediate subset
(CD14++CD16+), which mimicked the scenario in
patients, where classical CD14++CD16−
monocytes convert to CD14+CD16++
non-classical monocytes through a CD14++CD16+
monocyte intermediate (14),
although it remains elusive how CTRP3 brought about the phenotypic
changes. Intermediate monocytes are particularly viewed as
pro-atherogenic, as they selectively express CCR5, which has been
associated with atherosclerosis in experimental and large
epidemiological studies (31,32), but
mainly with angiogenic (33) and
reparative properties and reduced cytokine production (13). This paradigm was observed in the
present study, which demonstrated that CTRP3 antagonized the
LPS-induced upregulation of IL-6 expression but had a less
pronounced influence on TNF-α expression. The limited ability of
CTRP3 to inhibit the LPS-induced upregulation of TNF-α expression
is likely associated with the minor response of monocytes to
LPS-induced stress in the present experimental setting in
comparison to that in a previous study (34), or CTRP3 is preferentially effective
in the inhibition of IL-6 production. Furthermore, the inhibitory
effect is likely exerted via blunting the activity of the p38
MAPK/ERK cascade, particularly in elderly patients (26).
Of note, the present study has certain limitations:
i) The physical data and associated parameters (age, sex and
hemodynamics) in the present study were not exactly matched between
AMI patients and volunteer controls, which may yield pre-exiting
bias on subject selection. Nevertheless, the comparison of monocyte
subsets remain valid as the blood cell counts of all subjects were
comparable. ii) The present study mainly focused the intermediate
subset of monocytes as an interventional target. In the biology of
blood monocytes, the intermediate subpopulation, although its
proportion was small, is the most critical and informative to
demonstrate the functional transition from pro-inflammatory to
anti-inflammatory status in patients with AMI. The other two
subsets (classical and non-classical) may be equally important and
should be addressed in a future study. iii) In the ex vivo
experiments, the cultivation time was restricted to 24 h, which may
have somewhat limited the phenotypic changes, particularly after
CTRP3 treatment. However, when the culture time was extended up to
36 or 48 h, most of the cultivated monocytes transformed into a
macrophage phenotype on the dish (see supplementary Fig. S1), indicating phenotypic transition
of monocytes and spontaneous differentiation of monocytes into
macrophages even under culture conditions with AS. This inevitable
phenomenon does not permit any prolongation of culture time in the
present setting and requires technical circumvention in future
studies. iv) There is limited evidence of p38 MAPK/ERK activation
after CTRP3 intervention. As the major focus of the present study
was to reveal the dynamic fluctuation of monocyte subsets and only
provided ‘proof-of-concept’ evidence by using CTRP3 to modulate the
immune response after AMI, further experiments to gain insight at
the molecular level are currently being performed by our team and
the results will be reported in a separate study in the near
future.
In conclusion, the present study indicated a
dynamical fluctuation of monocyte counts and the proportions of
three subsets, which underscores the notion that the clinical use
of monocytes as a predictive marker of cardiac risk should be
performed with caution due to the time-dependent variation of
circulating monocytes in patients with AMI. The increased monocyte
counts and their elevated phagocytic and migratory activities
pinpoint an enhanced immune response to tissue damage. Furthermore,
CTRP3, a member of the CTRP family, may represent an alternative
candidate to modulate the ‘uncontrolled’ inflammatory response and
to augment the cardiac reparative process in patients following
AMI.
Supplementary Material
Supporting Data
Acknowledgements
The authors thank Ms Jiangfang Zhang, Department of
Cardiology, People's Hospital of Danyang, The Affiliated Hospital
of Nantong University (Danyang, China) for her technical support in
the flow cytometric analysis and Prof. Dr. Linlin Qiu from the
Department of Cardiology, People's Hospital of Danyang, The
Affiliated Hospital of Nantong University (Danyang, China) for his
constructive discussions that conceptually formed the major body of
this study.
Funding
This study was supported by the Department of
Zhenjiang S&T (grant no. SHW2015020) and the Social Development
Foundation of Zhenjiang, China (grant no. FZ2017010).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
HZ and WW conceived the study, designed the
experiments and interpreted the data. YD participated in performing
the experiments of monocyte isolation and characterization by flow
cytometry. YZ, XD and JZ performed ex vivo experiments and
RT-qPCR analysis of cytokine expression. WO, JG and YZ acquired all
blood samples from patients and performed statistical analysis of
all experimental data. XL provided conceptual advice, interpreted
the results and wrote the manuscript. All authors have read and
approved the final version of the manuscript.
Ethics approval and consent to
participate
The study was approved by the Ethics Committee of
the People's Hospital of Danyang (Danyang, China) and was performed
in accordance with the Declaration of Helsinki. All participants or
companying relatives provided written informed consent for the use
of their monocytes for scientific research in accordance with the
Declaration of Helsinki directly after AMI.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
AMI
|
acute myocardial infarction
|
|
CTRP3
|
complement C1q tumor necrosis
factor-related protein-3
|
|
AS
|
autologous serum
|
|
LPS
|
lipopolysaccharides
|
References
|
1
|
Godwin JW, Pinto AR and Rosenthal NA:
Chasing the recipe for a pro-regenerative immune system. Semin Cell
Dev Biol. 61:71–79. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Karin M and Clevers H: Reparative
inflammation takes charge of tissue regeneration. Nature.
529:307–315. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Chen B and Frangogiannis NG: Immune cells
in repair of the infarcted myocardium. Microcirculation. 24:2017.
View Article : Google Scholar
|
|
4
|
Tang J, Wang X, Tan K, Zhu H, Zhang Y,
Ouyang W, Liu X and Ding Z: Injury-induced fetal reprogramming
imparts multipotency and reparative properties to pericardial
adipose stem cells. Stem Cell Res Ther. 9:2182018. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Arfvidsson J, Ahlin F, Vargas KG, Thaler
B, Wojta J and Huber K: Monocyte subsets in myocardial infarction:
A review. Int J Cardiol. 231:47–53. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Nahrendorf M, Pittet MJ and Swirski FK:
Monocytes: Protagonists of infarct inflammation and repair after
myocardial infarction. Circulation. 121:2437–2445. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Barberà–Cremades M, Baroja-Mazo A and
Pelegrín P: Purinergic signaling during macrophage differentiation
results in M2 alternative activated macrophages. J Leukoc Biol.
99:289–299. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Tan K, Zhu H, Zhang J, Ouyang W, Tang J,
Zhang Y, Qiu L, Liu X, Ding Z and Deng X: CD73 expression on
mesenchymal stem cells dictates the reparative properties via its
anti-inflammatory activity. Stem Cells Int. 2019:87176942019.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Gordon S and Taylor PR: Monocyte and
macrophage heterogeneity. Nat Rev Immunol. 5:953–964. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ziegler-Heitbrock L: Blood monocytes and
their subsets: Established features and open questions. Front
Immunol. 6:4232015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Berg KE, Ljungcrantz I, Andersson L,
Bryngelsson C, Hedblad B, Fredrikson GN, Nilsson J and Björkbacka
H: Elevated CD14++CD16- monocytes predict cardiovascular events.
Circ Cardiovasc Genet. 5:122–131. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Buscher K, Marcovecchio P, Hedrick CC and
Ley K: Patrolling mechanics of non-classical monocytes in vascular
inflammation. Front Cardiovasc Med. 4:802017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ancuta P, Liu KY, Misra V, Wacleche VS,
Gosselin A, Zhou X and Gabuzda D: Transcriptional profiling reveals
developmental relationship and distinct biological functions of
CD16+ and CD16- monocyte subsets. BMC Genomics. 10:4032009.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zawada AM, Rogacev KS, Rotter B, Winter P,
Marell RR, Fliser D and Heine GH: SuperSAGE evidence for
CD14++CD16+ monocytes as a third monocyte subset. Blood.
118:e50–e61. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Heine GH, Ulrich C, Seibert E, Seiler S,
Marell J, Reichart B, Krause M, Schlitt A, Köhler H and Girndt M:
CD14(++)CD16+ monocytes but not total monocyte numbers predict
cardiovascular events in dialysis patients. Kidney Int. 73:622–629.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Neuser J, Galuppo P, Fraccarollo D, Willig
J, Kempf T, Berliner D, Bauersachs J and Widder JD: Intermediate
CD14++CD16+ monocytes decline after transcatheter aortic valve
replacement and correlate with functional capacity and left
ventricular systolic function. PLoS One. 12:e01836702017.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wildgruber M, Aschenbrenner T, Wendorff H,
Czubba M, Glinzer A, Haller B, Schiemann M, Zimmermann A, Berger H,
Eckstein HH, et al: The ‘Intermediate
CD14++CD16+ monocyte subset increases in
severe peripheral artery disease in humans. Sci Rep. 6:394832016.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Stansfield BK and Ingram DA: Clinical
significance of monocyte heterogeneity. Clin Transl Med. 4:52015.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Tsujioka H, Imanishi T, Ikejima H, Kuroi
A, Takarada S, Tanimoto T, Kitabata H, Okochi K, Arita Y, Ishibashi
K, et al: Impact of heterogeneity of human peripheral blood
monocyte subsets on myocardial salvage in patients with primary
acute myocardial infarction. J Am Coll Cardiol. 54:130–138. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhu L, Yin Y, Zhou R, Lin J, Li J and Ye
J: Changes of monocyte subsets in patients with acute coronary
syndrome and correlation with myocardial injury markers. Int J Clin
Exp Pathol. 8:7266–7271. 2015.PubMed/NCBI
|
|
21
|
Cappellari R, D'Anna M, Bonora BM, Rigato
M, Cignarella A, Avogaro A and Fadini GP: Shift of monocyte subsets
along their continuum predicts cardiovascular outcomes.
Atherosclerosis. 266:95–102. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Li Y, Wright GL and Peterson JM:
C1q/TNF-related protein 3 (CTRP3) function and regulation. Compr
Physiol. 7:863–878. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Peterson JM, Wei Z and Wong GW:
C1q/TNF-related protein-3 (CTRP3), a novel adipokine that regulates
hepatic glucose output. J Biol Chem. 285:39691–39701. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wu D, Lei H, Wang JY, Zhang CL, Feng H, Fu
FY, Li L and Wu LL: CTRP3 attenuates post-infarct cardiac fibrosis
by targeting Smad3 activation and inhibiting myofibroblast
differentiation. J Mol Med (Berl). 93:1311–1325. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Flögel U, Ding Z, Hardung H, Jander S,
Reichmann G, Jacoby C, Schubert R and Schrader J: In vivo
monitoring of inflammation after cardiac and cerebral ischemia by
fluorine magnetic resonance imaging. Circulation. 118:140–148.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Nyugen J, Agrawal S, Gollapudi S and Gupta
S: Impaired functions of peripheral blood monocyte subpopulations
in aged humans. J Clin Immunol. 30:806–813. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Epelman S, Liu PP and Mann DL: Role of
innate and adaptive immune mechanisms in cardiac injury and repair.
Nat Rev Immunol. 15:117–129. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
van der Laan AM, ter Horst EN, Delewi R,
Begieneman MP, Krijnen PA, Hirsch A, Lavaei M, Nahrendorf M,
Horrevoets AJ, Niessen HW and Piek JJ: Monocyte subset accumulation
in the human heart following acute myocardial infarction and the
role of the spleen as monocyte reservoir. Eur Heart J. 35:376–385.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Shantsila E and Lip GY: Monocyte diversity
in myocardial infarction. J Am Coll Cardiol. 54:139–142. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Nienhaus F, Colley D, Jahn A, Pfeiler S,
Flocke V, Temme S, Kelm M, Gerdes N, Flögel U and Bönner F:
Phagocytosis of a PFOB-nanoemulsion for 19F magnetic
resonance imaging: First results in monocytes of patients with
stable coronary artery disease and ST-elevation myocardial
infarction. Molecules. 24(pii): E20582019. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Fingerle G, Pforte A, Passlick B,
Blumenstein M, Ströbel M and Ziegler-Heitbrock HW: The novel subset
of CD14+/CD16+ blood monocytes is expanded in sepsis patients.
Blood. 82:3170–3176. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Rogacev KS, Cremers B, Zawada AM, Seiler
S, Binder N, Ege P, Grosse-Dunker G, Heisel I, Hornof F, Jeken J,
et al: CD14++CD16+ monocytes independently predict cardiovascular
events. J Am Coll Cardiol. 60:1512–1520. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Jaipersad AS, Lip GY, Silverman S and
Shantsila E: The role of monocytes in angiogenesis and
atherosclerosis. J Am Coll Cardiol. 63:1–11. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Weigert J, Neumeier M, Schäffler A, Fleck
M, Schölmerich J, Schütz C and Buechler C: The adiponectin paralog
CORS-26 has anti-inflammatory properties and is produced by human
monocytic cells. FEBS Lett. 579:5565–5570. 2005. View Article : Google Scholar : PubMed/NCBI
|