Introduction
The chemical composition of compounds, including
prototypes and metabolites, is the basis of the effect of
traditional Chinese medicine (1,2). The
majority of Chinese medicines enter into the body via oral
administration and thus, interact with intestinal microbes
(3). A number of Chinese herbal
medicines play a therapeutic role primarily via intestinal flora
metabolism, which has become a popular research topic (2,3).
The intestinal microflora is an extremely complex
microbial system, and is associated with the health of the
individual or animal it exists within (4). Certain chemical components of
traditional Chinese medicine can inhibit or promote the growth of
certain bacteria, altering the composition of the intestinal
microflora (5). Conversely,
intestinal microflora can alter the metabolism of Chinese herbal
medicine directly, by reductive or hydrolytic reactions, or
indirectly, by producing a variety of enzymes, amino acids and
vitamins, such as N-acyl-3-hydroxyglycines and β-glucuronidases,
β-glucosidase and dioxygenase (3,6,7). Therefore, investigating the role of
intestinal microflora in the transformation and metabolism of the
components of Chinese medicines may aid in further understanding
the metabolic pathways of intestinal microflora and the potential
mechanisms of action of Chinese medicines in the body.
Cassia seed is the dried ripe seed of the leguminous
plant Cassia obtusifolia L. or Cassia tora L.
(8). It was first recorded in ‘Shen
Nong's Herbal Classic’ (9). The seed
displays extensive pharmacological actions, for example, modern
pharmacological research has reported that the cassia seed
possesses antibacterial activity (10,11).
Beyond the purgative action of the cassia seed, it has also been
reported to decrease blood pressure and alleviate hyperlipidemia
(12), protect the liver (13), improve immunity (14) and display anti-inflammatory (15) and neuroprotective effects in models
of Parkinson's disease (16,17). The cassia seed is commonly used in
the treatment of hypertension, fatty liver and constipation
(12,13). At present, research into the
pharmacodynamic properties of the cassia seed has primarily focused
on the anthraquinone compounds of the seed, which are related to
its effect on diarrhea (18,19). However, based on the complex
characteristics of Chinese medicine, it is speculated that the role
of the cassia seed in protecting the liver and cardio-cerebral
vessels, as well as improving eyesight may be related to other
unknown components, in particular the components that are
transformed by the intestinal microflora (20,21).
Based on the aforementioned understanding, the present study
cultured human or rat intestinal microflora suspensions with cassia
seeds in vitro, to clarify the mechanism of action of the
cassia seed. The present study focused on the transformation of
active ingredients by the intestinal microflora and examined the
differences in the transformation of active ingredients between
humans and laboratory rats. The water decoction of cassia seeds was
placed into culture medium containing human or rat intestinal
microflora suspension under an anaerobic and sterile environment
in vitro. Subsequently, the metabolites were analyzed using
an ultra-high-performance liquid chromatography (UPLC)-quadrupole
time-of-flight (QTOF) mass spectrometry (MS) system. The present
study hypothesized that if the water decoction of cassia seeds was
transformed by the intestinal microflora, a number of novel
compounds could be detected. Alternatively, if no novel compounds
were identified, the composition of the cassia seed would be as has
previously been described (20).
Materials and methods
Ethics
The present study was approved by the Ethics
Committee of Henan University of Chinese Medicine. Written informed
consent was received from all participants.
Materials and reagents
Raw cassia seeds were collected from the Medicine
Botanical Garden of Henan University of Chinese Medicine in
September 2016. The samples were identified by Professor Suiqing
Chen (College of Pharmacy, Henan University of Chinese Medicine) as
the seeds of the legume C. obtusifolia L. Ellagic acid
(batch no. 1013A022) was obtained from Beijing Solarbio Science
& Technology Co., Ltd. The broth medium (batch no. HB0384-1)
and agar powder (batch no. 01-023) were purchased from Haibo
Biotechnology Co., Ltd. Methanol was purchased from Tianjin Siyou
Fine Chemicals Co., Ltd. Acetonitrile (UPLC/MS grade) and formic
acid (high performance liquid chromatography grade) with a purity
of 99% were purchased from Kareo. Purified water was acquired from
an ESW-1-30 system (Easywell Water System, Inc.). All the other
reagents were obtained from Huayu Biotech Co., Ltd.
Preparation of the cassia seed
decoction
The cassia seeds were broken into pieces and weighed
using an electronic balance. Subsequently, 20.0 g of seeds were
soaked in 200 ml water for 30 min at 25˚C. The suspension was
boiled for at least 30 min and then the filtrate was collected
using a 0.22 µm microporous filter. The filter residue was mixed
with water in the ratio of 1:6 and boiled for 20 min. The resulting
filtrate was collected. The two filtrates were combined and
concentrated to 20 ml at 60˚C under vacuum, using a rotary
evaporator. The concentration of the resulting solution was 1.0
g/ml crude cassia seed, and the solution was stored at 4˚C until
further analysis.
Collection of human fecal samples
Human fecal samples were obtained from three healthy
males (aged 21, 22 and 23 years; body mass 60-70 kg; height 172-178
cm) in July 2018 from Jinshui, Zhengzhou, Henan. Each subject
provided two fecal samples to allow culture experiments to be
performed in duplicate. Samples were stored at 4˚C and were
processed within 1 h of donation. No differences in the microbial
concentrations of the samples were observed between fresh samples
before and after processing. The samples were maintained in anoxic
conditions using the YQX-II anaerobic workstation with 5%
CO2, 5% hydrogen and 90% nitrogen (Shanghai Longyue
Equipment Co., Ltd.).
Collection of rat fecal samples
Rat fecal samples were obtained from three
Sprague-Dawley rats (male; body mass ~250 g; age, 6-7 weeks; Jinan
Pengyue Experimental Animal Breeding Co., Ltd.). Rats were
maintained on a 12 light/dark cycle in a 22±2˚C room with 40%
relative humidity. Food and water were provided ad libitum.
To allow for culture experiments to be performed in duplicate, two
fecal samples were collected from each rat. All procedures
regarding the processing of the rat fecal samples were the same as
for the human fecal samples.
Culture of human or rat intestinal
microflora suspension with cassia seed decoction
The individual human or rat fecal samples were mixed
to an even consistency in a germ-free and anoxic environment.
Subsequently, ~1 g of fecal sample was mixed with 100 ml 0.1 M PBS
to produce an intestinal microflora suspension. Then, 200 µl human
or rat intestinal microflora suspension was placed into solid broth
medium (18 mg/ml broth medium, 16.7 mg/ml agar; sterilization at
121˚C for 20 min) with the water decoction of cassia seeds (0.3,
0.15 or 0.075 g/ml). Additionally, 200 µl human or rat intestinal
microflora suspension was placed into liquid broth medium (18 mg/ml
broth medium; sterilization at 121˚C for 20 min) with the water
decoction of cassia seeds (0.17 g/ml). The human or rat intestinal
microflora suspension was not added to the broth medium for the
control group. Each group had duplicate samples. The culture
protocol was performed in a germ-free laminar flow cabinet. The
culture was incubated in a YQX-II anoxic workstation (Shanghai
Longyue Equipment Co., Ltd.) for 48 h.
Preparation of samples
The culture medium solution was centrifuged at
11,342 x g at 4˚C for 10 min. Subsequently, 1 ml supernatant was
mixed with 1 ml 99.9% of methanol and the mixture was incubated at
4˚C for 10 min. The mixture was then centrifuged at 11,342 x g and
4˚C for 10 min. The resulting solution was filtered through a
0.22-mm membrane filter and then injected into the UPLC-QTOF/MS
system for analysis.
Liquid chromatography
The chromatographic analysis was performed on an
Acquity I-Class UPLC system (Waters Corporation). The separation
was performed using an Acquity BEH C18 column (100x2.1
mm2; particle size, 1.7 µm; Waters Corporation)
maintained at 25˚C with a flow rate of 0.4 ml/min. The injection
volume was 5 µl. The optimal mobile phase consisted of A
(acetonitrile) and B (HCOOH/H2O; 0.1:100). The optimized
UPLC gradient elution conditions were as follows: 0-1 min, 2% A and
98.0% B; 1-10 min, 98.0% A and 2.0% B; and 11-13 min, 2.0% A and
98.0% B. The detection wavelength was 256 nm.
MS
MS detections were performed on a Xevo-G2-XS QTOF
tandem mass spectrometer (Waters Corporation) with negative and
positive electrospray ionization (ESI) modes. The sensitivity of
the system ensured the identification of as high of a number of
putative compounds as possible. QTOF-MS was performed for the mass
ranges of 100-1,200 m/z, and the experiments were run with 200 msec
accumulation time. Positive and negative ionization modes were
tested and the negative ionisation mode was selected for improved
sensitivity. The conditions used for the ESI source were as
follows: Ion source injection voltage, 4 kV; capillary voltage, 3.0
kV; sampling cone, 40 V; source temperature, 120˚C; and desolvation
temperature, 500˚C. Nitrogen was used as a cone and desolvation gas
with a flow rate of 50 and 800 l/h, respectively. The nebulizer
pressure of the nitrogen gas was 100 psi. Argon was used as the
collision gas with a coll ision energy of 10-30 V, a scan time of
0.5 sec and an interval scan time of 0.02 sec. Acquiring data in
this manner allowed for information regarding the precursor and
fragment ions to be collected. Mass tolerance was set at <5 ppm
to reduce the number of options used to determine the elemental
compositions of both the precursor and the product ions.
Data processing and analysis
strategy
For data processing, Masslynx software (version 4.1;
Waters Corporation) was used for qualitative analyses. Extracted
ion chromatograms and the MS Library made by identification for
unknown components of the elucidation tool in the Masslynx software
were used to identify the target compounds. A formula database of
target compounds, including names, molecular formulas, accurate
molecular weights and chemical structures, was established for the
target compounds. This database was prepared using previously
reported information (22,23). Subsequently, the names of the target
compounds were imported into the extracted ion chromatograms in the
UNIFI Scientific Information System (Water Corporation) to finalize
the screening of the target compounds. After screening, the
compounds that matched the names, molecular formulas, accurate
molecular weights and chemical structures of the target compounds
in the formula database were extracted. Then, the target compound
information was compared with that of standard compounds (Beijing
Solarbio Science & Technology Co., Ltd.) whose spectra were
obtained by matching the MS/MS fragments. Hence, the common
compounds existing in the water decoction of cassia seeds were
identified. The structures of the metabolites were presumed
primarily based on accurate mass and mass fragmentation using the
UNIFI Scientific Information System. Finally, the fragment ions
were used to further confirm the chemical structures by making
comparisons with previously reported data (24-31).
Results
Discovery and identification of
ellagic acid in cassia seeds
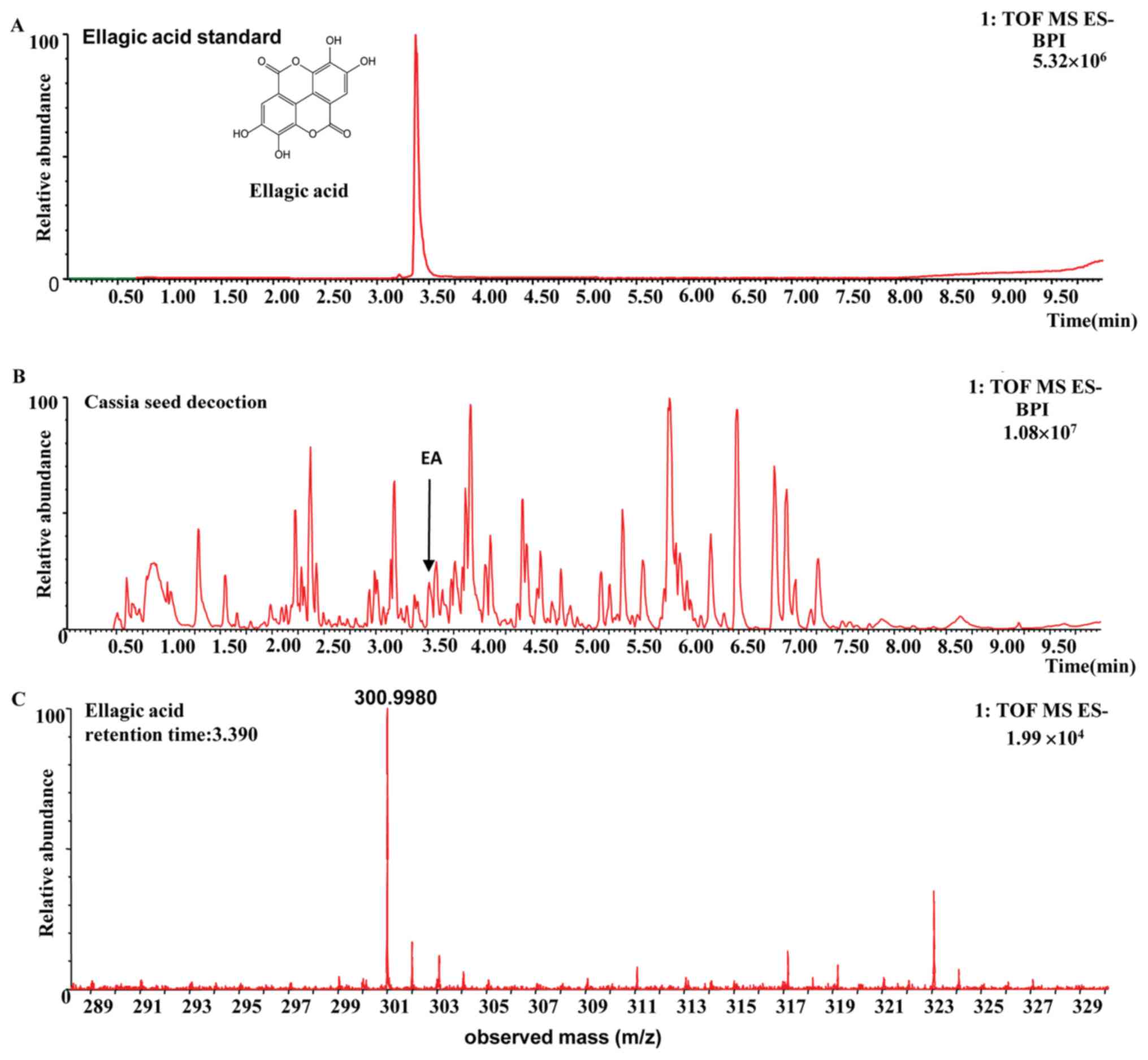
The compound was identified as ellagic acid on the
basis of its retention time and the m/z of characteristic ions,
compared with those of the standard compounds and data obtained
from the literature (22,23). The fragmentation pathway of ellagic
acid was recorded using a collision energy of 10-30 V and the
collision-induced dissociation MS/MS spectrum for ellagic acid is
shown in Fig. 1. In the MS/MS
spectrum, the [M-H]- ion at m/z 300.9980 was selected as
a precursor ion to provide fragmentation information. The precursor
ions produced were an [M-H-OH]- ion at m/z 283.9951
(C14H4O7) and an
[M-H-CO2]- ion at m/z 257.0080
(C13H5O7). The
[M-H-CO2]- ion yielded ions at m/z 229.01328
(C12H5O5), 201.01843
(C11H5O4), 173.02396
(C10H5O3) and 145.02948
(C9H5O2), consecutively by the
sequential loss of CO, and were identified as the characteristic
ions of ellagic acid (22,23). The
[M-H-CO2-CO]- ion at m/z 229.01328 underwent
direct elimination of CO2 and yielded the product
[M-H-2CO2-CO]- ion at m/z 185.02388
(C11H5O3). The structures,
retention times and MS data of compounds are summarized in Fig. 1 and Table
I.
 | Table IMass spectrometry data and proposed
fragmentation pathways of ellagic acid. |
Table I
Mass spectrometry data and proposed
fragmentation pathways of ellagic acid.
| tR
(min) | Ion (m/z) | Diff (ppm) | Chemical
formula | Fragmentation
pathways | Diff (ppm) | Identification |
|---|
| 3.39 | 300.9980 | -3.29 |
C14H5O8 | 283.99510
[M-H-OH]- | -4.052 | Ellagic acid |
| | | | | 257.00800
[M-H-CO2]- | -4.518 | |
| | | | | 229.01328
[M-H-CO2-CO]- | -4.220 | |
| | | | | 201.01843
[M-H-CO2-2CO]- | -4.478 | |
| | | | | 185.02388
[M-H-2CO2-CO]- | -2.904 | |
| | | | | 173.02396
[M-H-CO2-3CO]- | -2.643 | |
| | | | | 157.02930
[M-H-2CO2-2CO]- | -1.291 | |
| | | | | 145.02948
[M-H-CO2-4CO]- | -0.157 | |
General metabolites in the culture of
human or rat intestinal microflora suspension with the water
decoction of cassia seeds
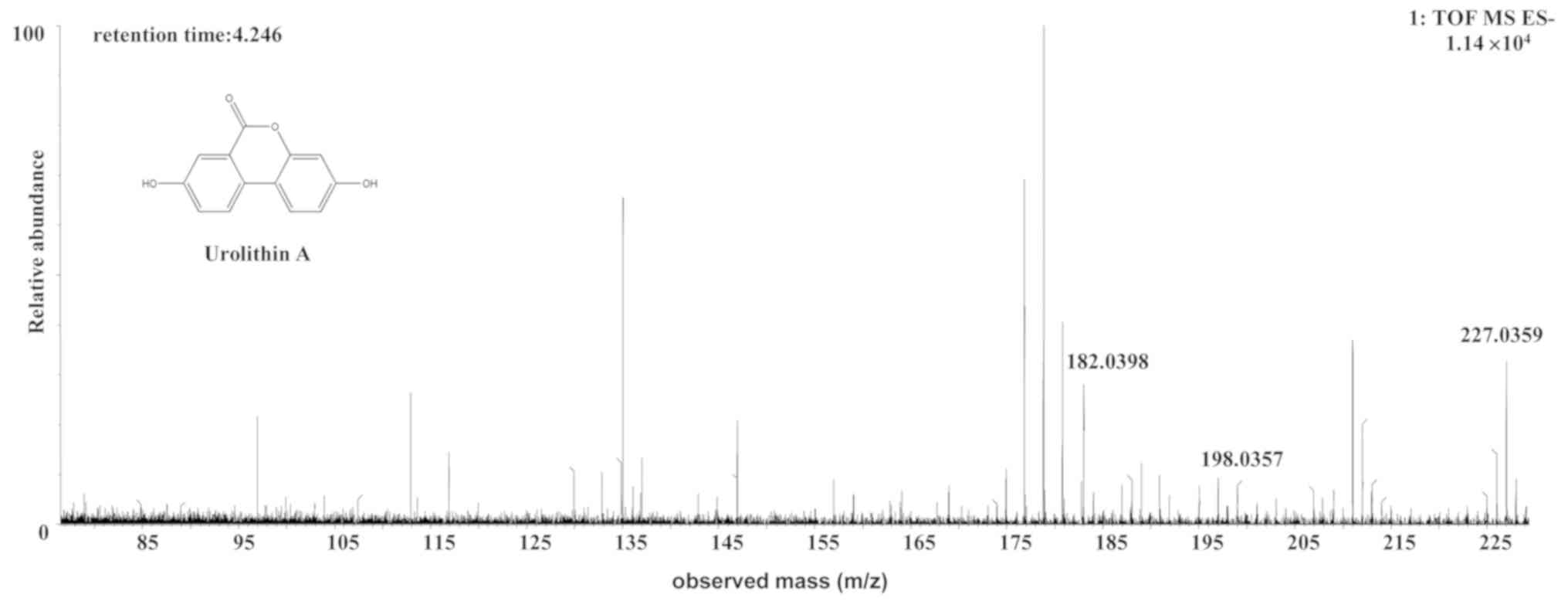
The [M-H]- ion at m/z 227.0329, with a
retention time of 4.25 min, gave rise to a fragment ion at m/z
198.0357 by the loss of an hydrogen ion and a carbonyl ion. The
fragment ion then gave rise to an [M-CO-O-H]- ion at m/z
182.0398 by the loss of an oxygen ion. It was speculated that the
[M-CO-O-H]- ion was an aromatic acid ester compound. The
characteristics of the [M-CO-O-H]- ion were the same as
those of the urolithin (uro)-A
(C13H8O4) metabolite of ellagic
acid (24). Therefore, the
[M-H]- ion was identified as uro-A. The structures,
retention times and MS data of the compounds are summarized in
Fig. 2 and Table II.
| Table IIMetabolites in the co-culture of the
human or rat intestinal microflora suspension and the cassia seed
decoction. |
Table II
Metabolites in the co-culture of the
human or rat intestinal microflora suspension and the cassia seed
decoction.
| Metabolites | Source | Chemical
formula | Retention time
(min) | Parent ion
(m/z) | Error (mda) | Molecular ion | MS/MS m/z
(error/ppm) |
|---|
| Uro-A | Human or rat |
C13H8O4 | 4.25 | 227.0329 | -0.8 (human)/0.3
(rat) |
[C13H7O4]- |
MS2[227.0329]: 198.0357,
182.0398 |
| Uro-B | Human or rat |
C13H8O3 | 5.92 | 211.0381 | -0.3 (human)/0.3
(rat) |
[C13H7O3]- |
MS2[211.0381]: 167.0509
(3.962), 139.0531(-15.991) |
| Uro-M6 | Human or rat |
C13H8O6 | 3.64 | 259.0249 | 0.1 (human)/0.5
(rat) |
[C13H7O6]- |
MS2[259.0249]: 213.0155
(-17.9898), 187.0428(14.690), 159.0453 (0.925) |
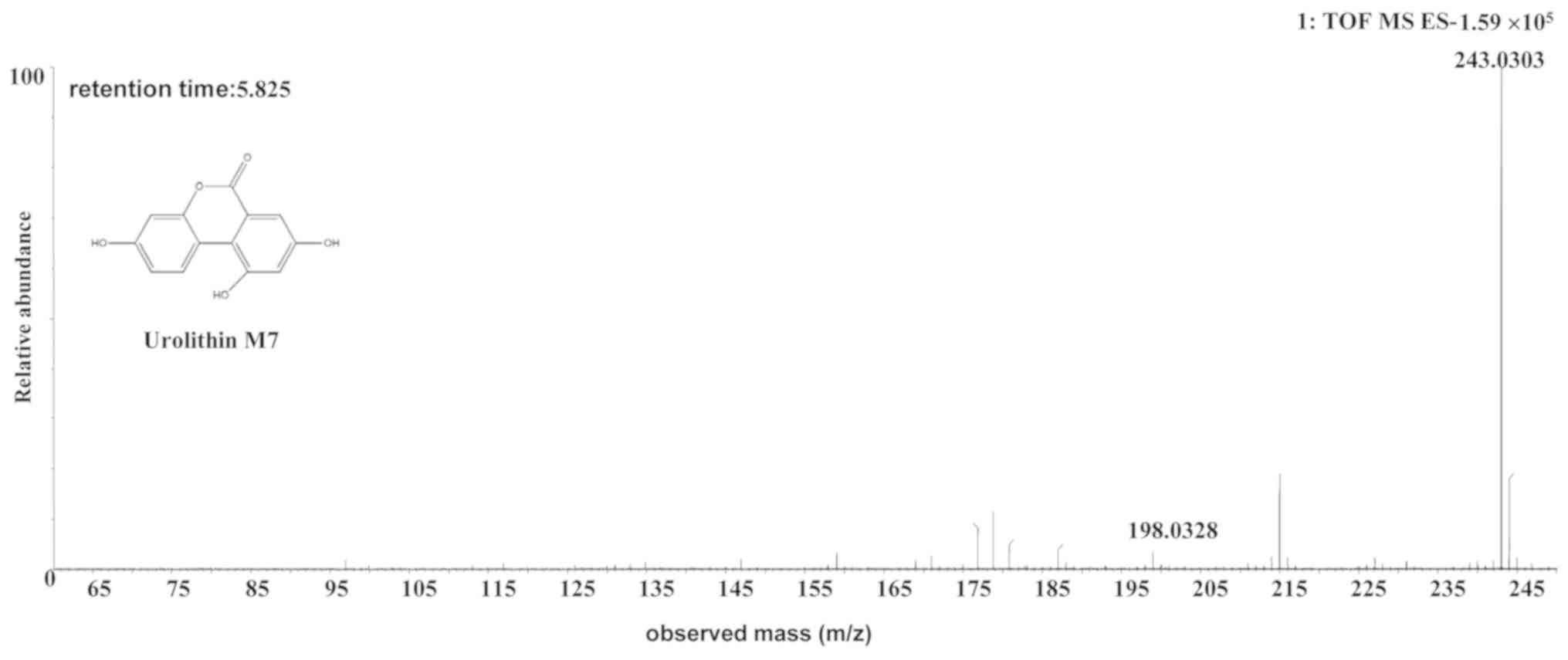
| Uro-M7 | Human or rat |
C13H8O5 | 5.82 | 243.0303 | -0.7 (human)/-0.5
(rat) |
[C13H7O5]- |
MS2[243.0303]: 198.0328 |
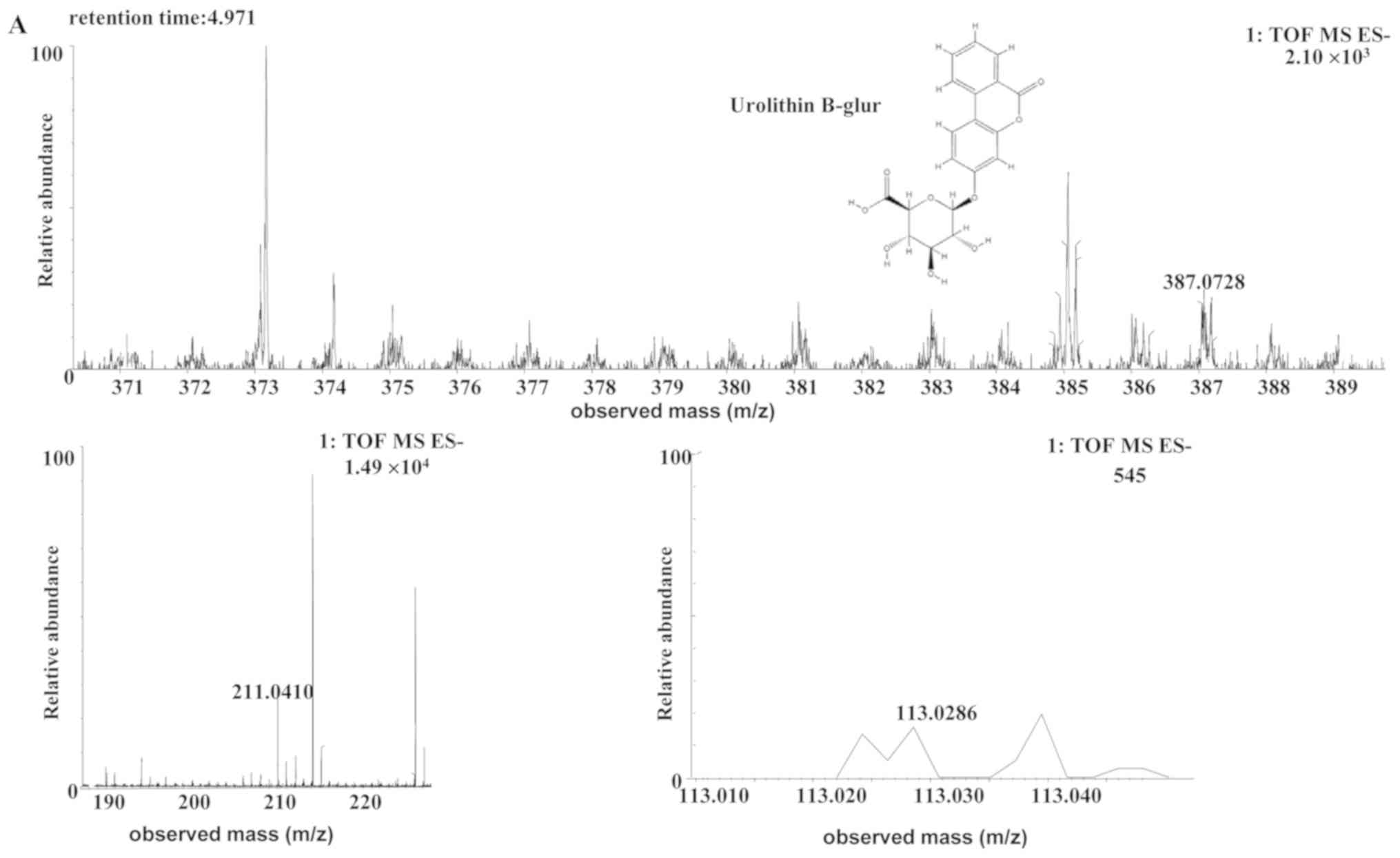
| Uro-B glur | Human |
C19H16O9 | 4.97 | 387.0728 | 0.4 (human) |
[C19H15O9]- |
MS2[387.0728]: 211.0410
(4.419), 113.0286 |
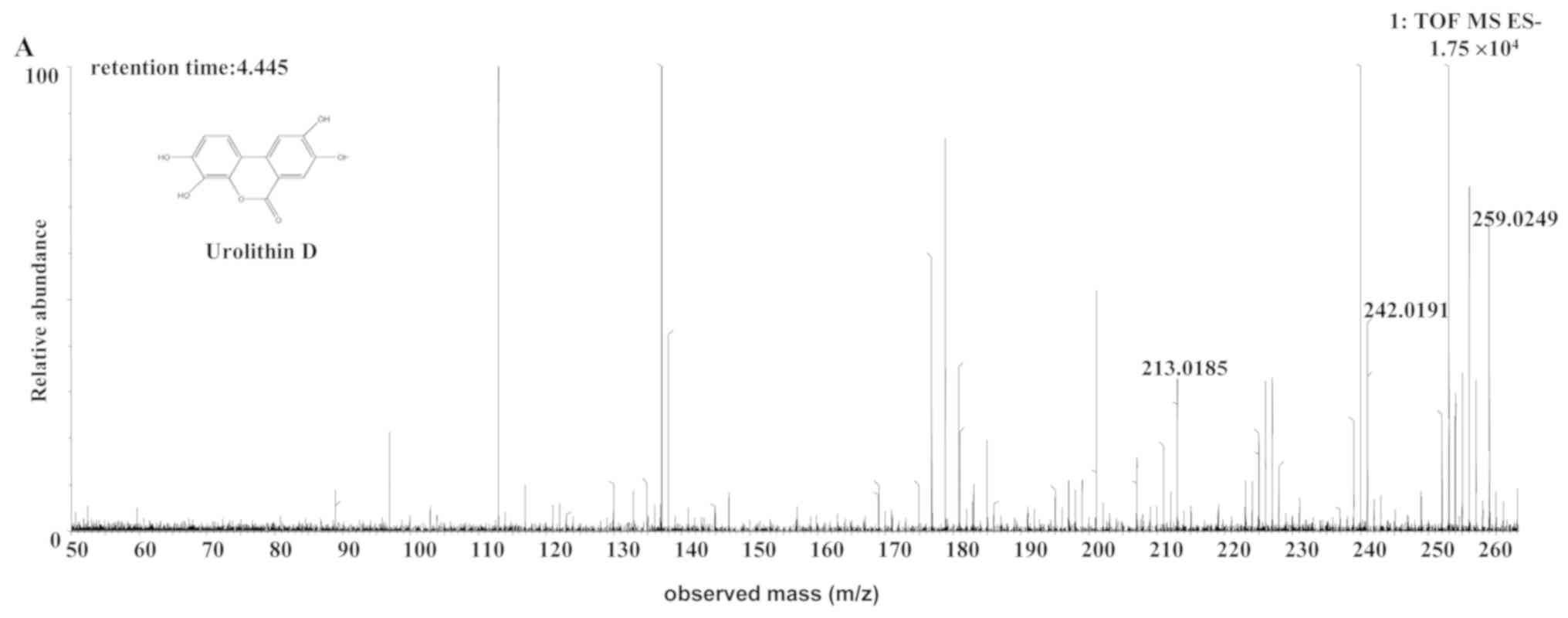
| Uro-D | Rat |
C13H8O6 | 4.45 | 259.0249 | 1.1 (rat) |
[C13H7O6]- |
MS2[259.0249]: 242.0191,
213.0185 (-3.906) |
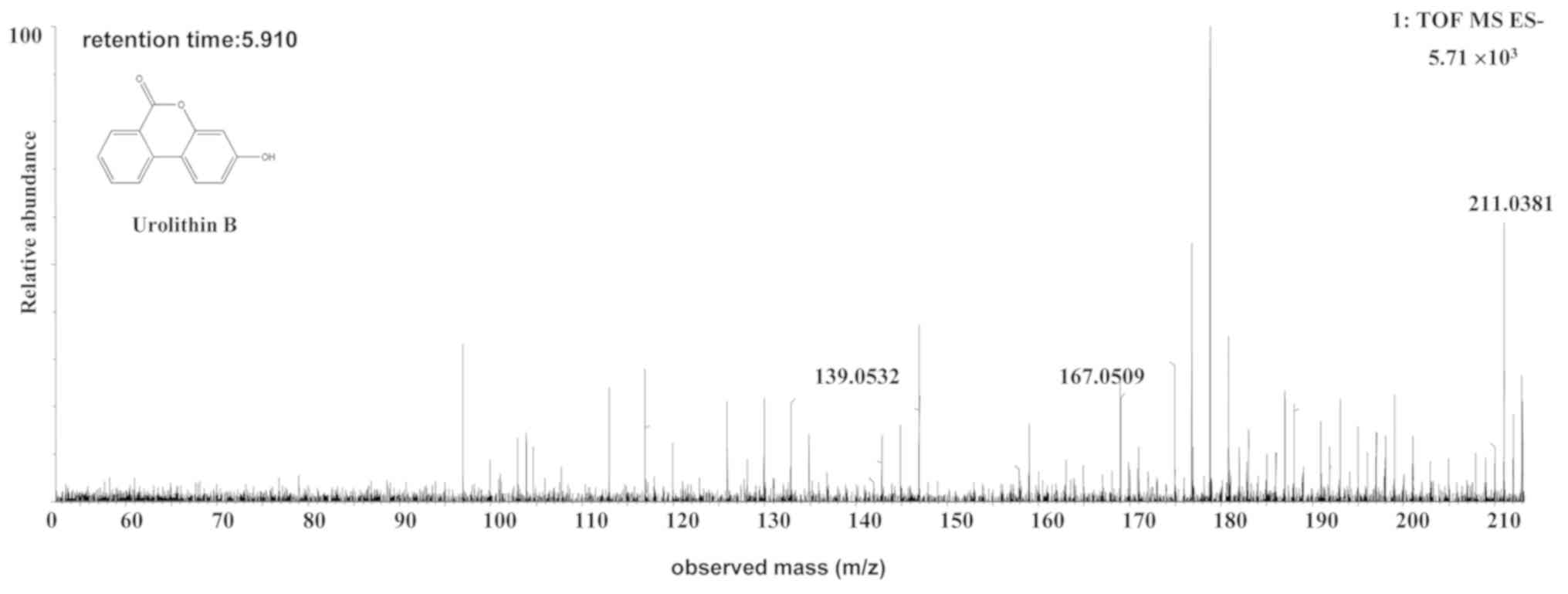
The [M-H]- ion at m/z 211.0381, with a
retention time of 5.92 min, gave rise to a fragment ion at m/z
167.0509, by the loss of an hydrogen ion and a carbonyl ion. The
fragment ion then gave rise to an [M-H-COO-CO]- fragment
ion at m/z 139.0531 by the loss of a carbonyl ion. The
characteristics of this ion were the same as those of the uro-B
(C13H8O3) metabolite of ellagic
acid (25). Therefore, the product
ion was identified as uro-B. The structures, retention times and MS
data of the compounds are summarized in Fig. 3 and Table
II.
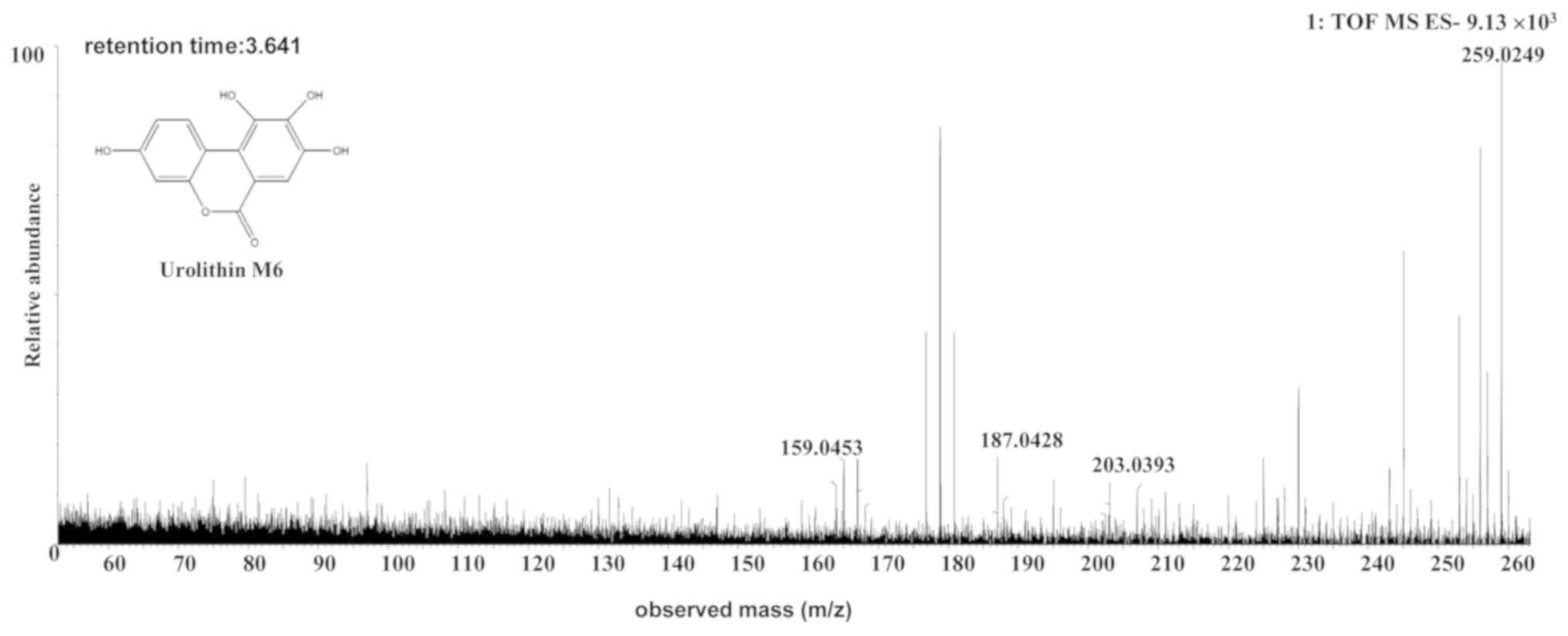
The [M-H]- ion at m/z 259.0249, with a
retention time of 3.64 min, gave rise to a fragment ion at m/z
213.0155, 187.0428 and 159.0453, by MS/MS analyses. The
characteristics of the [M-H]- ion were the same as those
of the uro-M6 (C13H8O6) metabolite
of ellagic acid (26,27). Therefore, the product ion was
identified as uro-M6. The structures, retention times and MS data
of the compounds are summarized in Fig.
4 and Table II.
The [M-H]- ion at m/z 243.0303, with a
retention time of 5.82 min, gave rise to a fragment ion at m/z
198.0328 [M-H-COOH]-, by MS/MS analyses. The
characteristics of this [M-H]- ion were the same as
those of the uro-M7 (C13H8O5)
metabolite of ellagic acid (28,29).
Therefore, the product ion was identified as uro-M7. The
structures, retention times and MS data of the compounds are
summarized in Fig. 5 and Table II.
Metabolites in the culture of human
intestinal microflora suspension with the water decoction of cassia
seeds
The [M-H]- ion at m/z 387.0728, with a
retention time of 4.97 min, gave rise to a fragment ion at m/z
211.0410 and 113.0286, by MS/MS analyses. The characteristics of
this [M-H]- ion were the same as those of the metabolite
uro-B-glucuronide (glur; C19H16O9)
of ellagic acid (30). Therefore,
the product ion was identified as uro-B-glur. The structures,
retention times and MS data of the compounds are summarized in
Fig. 6 and Table II.
Metabolites in the culture of rat
intestinal microflora suspension with the water decoction of cassia
seeds
The [M-H]- ion at m/z 259.0249, with a
retention time of 4.45 min, gave rise to an [M-OH-H]-
fragment ion at m/z 242.0191, by the loss of an hydroxide ion. The
fragment ion then gave rise to an [M-OH-CHO-H]- ion at
m/z 213.0185, by the loss of a CHO. The characteristics of this
[M-H]- ion were the same as those of the uro-D
(C13H8O6) metabolite of ellagic
acid (31). Therefore, the product
ion was identified as uro-D. The structures, retention times and MS
data of the compounds are summarized in Fig. 7 and Table
II.
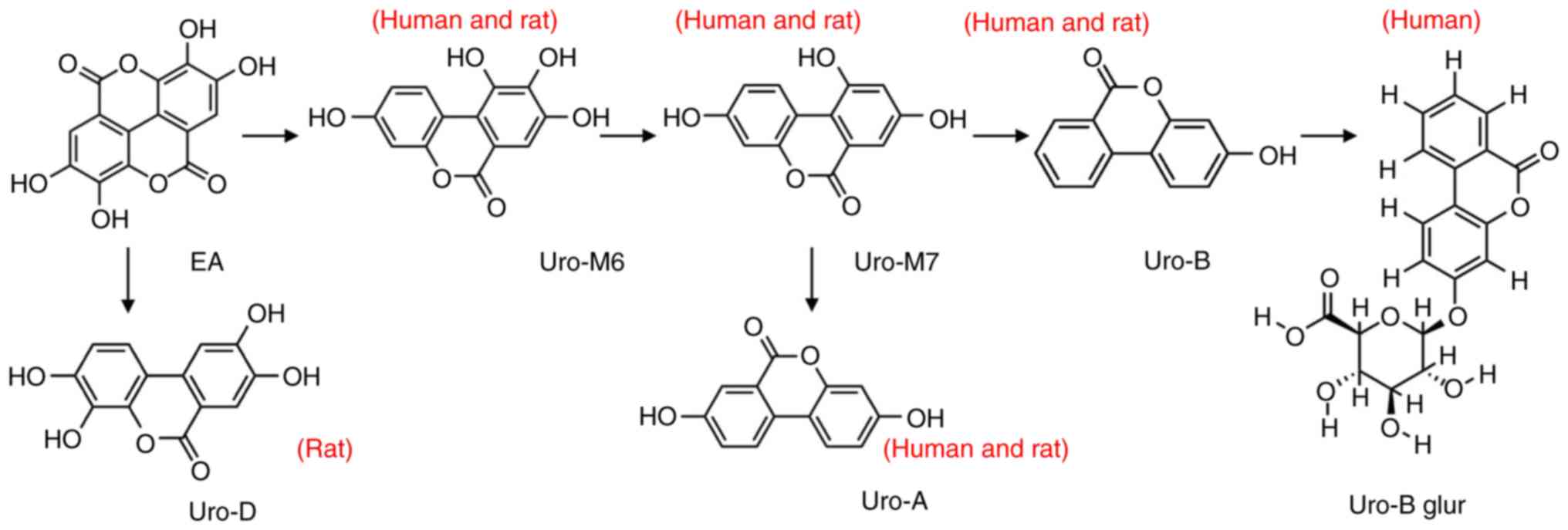
Metabolic pathways of ellagic
acid
Urolithin was generated from ellagic acid via three
metabolic pathways (Fig. 8). The
metabolites were uro-A, uro-B and uro-D. Moreover, the metabolic
pathways were different for human or rat intestinal microflora.
uro-B was further transformed into uro-B-glur by human intestinal
microflora and uro-D was generated only by the rat intestinal
microflora.
Discussion
In the present study, the interaction between the
intestinal micoflora and the cassia seed decoction was
investigated. UPLC-MS/MS technology is a combination of
high-resolution UPLC and highly sensitive and selective MS. The use
of this technology does not require the measured samples to be
subjected to purification or derivatization with complicated
separation and enrichment processes prior to analysis (32). A large amount of reliable qualitative
and quantitative information can be quickly obtained following a
simple pre-treatment with UPLC-MS/MS technology (32). The present study described a novel,
rapid and effective UPLC-MS/MS method for the discovery and
identification of ellagic acid in the cassia seed decoction.
Ellagic acid is a derivative of gallic acid, which is a tannin with
antioxidant, anticancer and anti-inflammatory effects (33). Ellagic acid also regulates the
intestinal microflora and performs other functions, including its
anti-atherogenic and neuroprotective effects (33). Despite displaying a wide range of
biological activities, the permeability of ellagic acid is too low
for direct absorption into the body. Synthesis and transformation
of ellagic acid are still widely used today. In general, ellagic
acid is obtained in two main forms. One form can be produced by the
oxidative polymerization of gallic acid or gallic acid ester under
the action of peroxidase. The other form is prepared by hydrolyzing
ellagitannin, which requires a high temperature and is performed
under acidic conditions (33).
Extraction methods using hot water reflux in this experiment do not
provide the correct conditions for the conversion of other
components into ellagic acid (34).
Therefore, the present study identified ellagic acid as one of the
natural components and parent compounds in the cassia seed.
Previous studies have reported that ellagic acid is
metabolized to the more easily absorbed urolithin by the intestinal
microflora (28,30). Compared with traditional methods such
as HPLC or UPLC, UPLC-MS/MS can identify a number of metabolites in
a short time, reduce the use of animals and facilitate the
metabolite identification process (23). The environmental factors influencing
the metabolic process are easy to control in the UPLC-MS/MS system
(28,32,35).
Moreover, certain active metabolites in the biological samples were
likely to be generated at relatively low levels and UPLC-MS/MS is
able to detect low levels of metabolites. A number of studies have
used UPLC-MS/MS to detect novel compounds in the co-culture of
human or rat intestinal microflora and the cassia seed decoction
(20,35). The cassia seed contains a number of
chemical components, including anthraquinones and tannins (12). Although the cassia seed contains a
large amount of anthraquinones, anthraquinones are also present in
a number of other traditional Chinese medicines, including rhubarb
(Rheum palmatum L.) (12,35). The
biotransformation of anthraquinone metabolites under the action of
the intestinal microflora has been previously investigated using
tUPLC-QTOF/MS (35).
The present study identified ellagic acid as a novel
compound in the cassia seed decoction. To further investigate the
presence of ellagic acid in the cassia seed decoction, the
biotransformation metabolites of ellagic acid were detected using a
UPLC-Q-TOF/MS system. The six types of urolithin, which are the
metabolites of ellagic acid under the action of the intestinal
microflora, were detected and identified. To the best of our
knowledge, the present study was the first to identify ellagic acid
as a component of the cassia seed. Furthermore, the present study
suggested that the urolithin metabolites of ellagic acid may be the
active component of cassia seeds.
The therapeutic effect of urolithin has been widely
recognized (36-40).
Urolithin can enhance the anti-inflammatory effects of neutrophils,
alleviate the symptoms of inflammation (36) and protect against prostate cancer
(37). Uro-A and uro-B increase cell
viability and protect cells, reduce malondialdehyde content and
enhance superoxide dismutase activity (38). Uro-A regulates the expression of
intercellular adhesion factor-1 via the ERK/peroxisome proliferator
activated receptor-γ signaling pathway, which exerts
anti-inflammatory effects (39).
Uro-A also induces mitophagy, prolongs the lifespan of
Caenorhabditis elegans and increases muscle function in
rodents (40,41). In the present study, it was
speculated that uro-A was the active component that induced the
antioxidant effect of cassia seeds (12,42).
Furthermore, the present study identified a
difference between the presence of urolithins in human and rat
intestinal microflora. Uro-B-glur was only identified in human
intestinal microflora and uro-D only in rat intestinal microflora.
This finding indicated that the differences between metabolic
products in human and rat intestinal microflora might be related to
the abundance and diversity of the intestinal microflora. A
significant difference between human and rat intestinal microflora
has been previously reported. For example, the proportion of
Bacteroides and Variovorax was the highest in human
and rat intestinal microflora, respectively (43,44).
Furthermore, ellagic acid was metabolized to urolithin and its
derivatives under the action of lactonase (45) and high concentrations of G.
urolithinfaciens DSM 27213T (≥107 cfu/g
feces) (46). Moreover, the
degradation of ellagic acid by the intestinal microflora leads to
the formation of different urolithin metabolites in fecal cultures
in vitro (20). The
intestinal microflora is a micro-ecological system that is
important in the interaction between the animal, medicine and
disease. However, in the present study it was difficult to
understand the complexity of this interaction because the
individual differences between the three human or animal samples
were large. This is a potential limitation of the present study.
Furthermore, additional drug metabolism systems, including the
liver, are present in whole organisms. Therefore, cassia seed
treatment in vivo could reveal different results compared
with in vitro treatment. The effect of the intestinal
microbiota on drug metabolism has gained importance in clinical
research because it can influence the hepatic drug metabolism of
the host (21,47). In the present study, although ellagic
acid was not a key or main ingredient of the crude cassia seed, it
could be identified in the chemical profiles of the cassia seed.
Therefore, further investigation into ellagic acid and the
components of the cassia seed is required.
In summary, the cassia seed has a complex chemical
composition and a wide range of pharmacological activities
(12). The seed is a natural
botanical drug, which in recent years has been widely used in
research (12). In the present
study, ellagic acid was identified in the cassia seed decoction and
subsequently, ellagic acid and its metabolite urolithin were
studied. The present study suggested that urolithin and the
intermediate metabolites were produced by ellagic acid in the
cassia seed decoction, under the action of the intestinal
microflora by hydrolysis and glucuronidation. The mechanism of
action of the intestinal microflora might be related to the
abundance and diversity of the human or rat intestinal microflora.
The metabolism of ellagic acid involved the biotransformation of
urolithin into a number of final products, including uro-B-glur.
However, with regards to uro-B-glur, the related mass spectrometry
signal was buried in the baseline noise, potentially due to the low
content of the product in the sample. The present study provided
rationale for further pharmacological and clinical investigation
into the mechanisms of action of the cassia seed and to clarify the
pathways and mechanisms of action of phytochemicals containing
ellagic acid.
Acknowledgements
The authors would like to thank Dr Wei-Xia Li and Dr
Ying-Jie Cao of the Pharmaceutical Department of the First
Affiliated Hospital of the Henan University of Chinese Medicine for
their technical guidance.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant nos. 81503363 and
81473435) and the Science and Technology Innovation Talents Support
Project of the Henan University of Chinese Medicine (grant no.
2015XCXRC02).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
SHW and HBL conceived and designed the study. NL and
YJQ performed the LC-MS analysis and collected the data. GLL
prepared the samples. SHW wrote the manuscript. All authors read
and approved the final manuscript.
Ethics approval and consent to
participate
The present study was approved by the Ethics
committee of Henan University of Chinese Medicine (approval no.
DWLL2018030060). Written informed consent was obtained from all
participants.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Yang C, Yin X, Dong X Zhang X, You L, Wang
W, Wang J, Chen Q and Ni J: Determination of the phytochemical
composition of Jingning fang and the in vivo pharmacokinetics of
its metabolites in rat plasma by UPLC-MS/MS. J Chromatogr B Analyt
Technol Biomed Life Sci. 1067:71–88. 2017.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Wu S, Xu W, Wang FR and Yang XW: Study of
the biotransformation of tongmai formula by human intestinal flora
and its intestinal permeability across the caco-2 cell monolayer.
Molecules. 20:18704–18716. 2015.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Feng W, Ao H, Peng C and Yan D: Gut
microbiota, a new frontier to understand traditional Chinese
medicines. Pharmacol Res. 142:176–191. 2019.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Goodman AL and Gordon JI: Our unindicted
coconspirators: Human metabolism from a microbial perspective. Cell
Metab. 12:111–116. 2010.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Wang RF, Yuan M, Yang XB, Xu W and Yang
XW: Intestinal bacterial transformation-a nonnegligible part of
Chinese medicine research. J Asian Nat Prod Res. 15:532–549.
2013.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Koeth RA, Wang Z, Levison BS, Buffa JA,
Org E, Sheehy BT, Britt EB, Fu X, Wu Y, Li L, et al: Intestinal
microbiota metabolism of L-carnitine, a nutrient in red meat,
promotes atherosclerosis. Nat Med. 19:576–585. 2013.PubMed/NCBI View
Article : Google Scholar
|
|
7
|
Cohen LJ, Esterhazy D, Kim SH, Lemetre C,
Aguilar RR, Gordon EA, Pickard AJ, Cross JR, Emiliano AB, Han SM,
et al: Commensal bacteria make GPCR ligands that mimic human
signalling molecules. Nature. 549:48–53. 2017.PubMed/NCBI View Article : Google Scholar
|
|
8
|
State Pharmacopoeia Commission of the PRC:
Pharmacopoeia of the People's Republic of China. Vol. Ⅰ. Beijing.
People's Medical Publishing House, 2015.
|
|
9
|
Xin T, Zhang Y, Pu X, Gao R, Xu Z and Song
J: Trends in herbgenomics. Sci China Life Sci. 62:288–308.
2019.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Huang YL, Chow CJ and Tsai YH:
Composition, characteristics, and in-vitro physiological effects of
the water-soluble polysaccharides from Cassia seed. Food Chem.
134:1967–1972. 2012.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Sahu J, Koley KM and Sahu BD: Attribution
of antibacterial and antioxidant activity of Cassia tora extract
toward its growth promoting effect in broiler birds. Vet World.
10:221–226. 2017.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Dong X, Fu J, Yin X, Yang C, Zhang X, Wang
W, Du X, Wang Q and Ni J: Cassiae semen: A review of its
phytochemistry and pharmacology (Review). Mol Med Rep.
16:2331–2346. 2017.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Xie Q, Guo FF and Zhou W: Protective
effects of cassia seed ethanol extract against carbon
tetrachloride-induced liver injury in mice. Acta Biochim Pol.
59:265–270. 2012.PubMed/NCBI
|
|
14
|
Kim M, Lim SJ, Lee HJ and Nho CW: Cassia
tora seed extract and its active compound aurantio-obtusin inhibit
allergic responses in IgE-mediated mast cells and anaphylactic
models. J Agric Food Chem. 63:9037–9046. 2015.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Yi JH, Park HJ, Lee S, Jung JW, Kim BC,
Lee YC, Ryu JH and Kim DH: Cassia obtusifolia seed ameliorates
amyloid β-induced synaptic dysfunction through anti-inflammatory
and Akt/GSK-3β pathways. J Ethnopharmacol. 178:50–57.
2016.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Ju MS, Kim HG, Choi JG, Ryu JH, Hur J, Kim
YJ and Oh MS: Cassiae semen, a seed of Cassia obtusifolia, has
neuroprotective effects in Parkinson's disease models. Food Chem
Toxicol. 48:2037–2044. 2010.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Drever BD, Anderson WG, Riedel G, Kim DH,
Ryu JH, Choi DY and Platt B: The seed extract of Cassia obtusifolia
offers neuroprotection to mouse hippocampal cultures. J Pharmacol
Sci. 107:380–392. 2008.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Shi BJ, Zhang WD, Jiang HF, Zhu YY, Chen
L, Zha XM, Lu YY and Zhang WM: A new anthraquinone from seed of
Cassia obtusifolia. Nat Prod Res. 30:35–41. 2016.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Xu YL, Tang LY, Zhou XD, Zhou GH and Wang
ZJ: Five new anthraquinones from the seed of Cassia obtusifolia.
Arch Pharm Res. 38:1054–1058. 2015.PubMed/NCBI View Article : Google Scholar
|
|
20
|
García-Villalba R, Beltrán D, Espín JC,
Selma MV and Tomás-Barberán FA: Time course production of
urolithins from ellagic acid by human gut microbiota. J Agric Food
Chem. 61:8797–8806. 2013.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Koppel N, Maini Rekdal V and Balskus EP:
Chemical transformation of xenobiotics by the human gut microbiota.
Science. 356:2017.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Mullen W, Yokota T, Lean ME and Crozier A:
Analysis of ellagitannins and conjugates of ellagic acid and
quercetin in raspberry fruits by LC-MSn. Phytochemistry.
64:617–624. 2003.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Lee JH, Johnson JV and Talcott ST:
Identification of ellagic acid conjugates and other polyphenolics
in muscadine grapes by HPLC-ESI-MS. J Agric Food Chem.
53:6003–6010. 2005.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Cerdá B, Periago P, Espín JC and
Tomás-Barberán FA: Identification of urolithin a as a metabolite
produced by human colon microflora from ellagic acid and related
compounds. J Agric Food Chem. 53:5571–5576. 2005.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Lucas R, Alcantara D and Morales JC: A
concise synthesis of glucuronide metabolites of urolithin-B,
resveratrol, and hydroxytyrosol. Carbohydr Res. 344:1340–1346.
2009.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Rupiani S, Guidotti L, Manerba M, Di Ianni
L, Giacomini E, Falchi F, Di Stefano G, Roberti M and Recanatini M:
Synthesis of natural urolithin M6, a galloflavin mimetic, as a
potential inhibitor of lactate dehydrogenase A. Org Biomol Chem.
14:10981–10987. 2016.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Nuñez-Sánchez MA, García-Villalba R,
Monedero-Saiz T, García-Talavera NV, Gómez-Sánchez MB,
Sánchez-Álvarez C, García-Albert AM, Rodríguez-Gil FJ, Ruiz-Marín
M, Pastor-Quirante FA, et al: Targeted metabolic profiling of
pomegranate polyphenols and urolithins in plasma, urine and colon
tissues from colorectal cancer patients. Mol Nutr Food Res.
58:1199–1211. 2014.PubMed/NCBI View Article : Google Scholar
|
|
28
|
González-Barrio R, Truchado P, Ito H,
Espin JC and Tomás-Barberán FA: UV and MS identification of
Urolithins and Nasutins, the bioavailable metabolites of
ellagitannins and ellagic acid in different mammals. J Agric Food
Chem. 59:1152–1162. 2011.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Seeram NP, Henning SM, Zhang Y, Suchard M,
Li Z and Heber D: Pomegranate juice ellagitannin metabolites are
present in human plasma and some persist in urine for up to 48
hours. J Nutr. 136:2481–2485. 2006.PubMed/NCBI View Article : Google Scholar
|
|
30
|
García-Villalba R, Espín JC and
Tomás-Barberán FA: Chromatographic and spectroscopic
characterization of urolithins for their determination in
biological samples after the intake of foods containing
ellagitannins and ellagic acid. J Chromatogr A. 1428:162–175.
2016.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Giorgio C, Mena P, Del Rio D, Brighenti F,
Barocelli E, Hassan-Mohamed I, Callegari D, Lodola A and Tognolini
M: The ellagitannin colonic metabolite urolithin D selectively
inhibits EphA2 phosphorylation in prostate cancer cells. Mol Nutr
Food Res. 59:2155–2167. 2015.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Fan M, Qin K, Ding F, Huang Y, Wang X and
Cai B: Identification and differentiation of major components in
three different ‘Sheng-ma’ crude drug species by UPLC/Q-TOF-MS.
Acta Pharm Sin B. 7:185–192. 2017.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Vattem DA and Shetty K: Biological
functionality of ellagic acid: A review. J Food Biochem.
29:234–266. 2005.
|
|
34
|
Jadhav PD and Laddha KS: Synthesis of new
ellagic acid derivatives. Indian J Chem B. 45:1551–1553. 2006.
|
|
35
|
Huang ZH, Xu Y, Wang Q and Gao XY:
Metabolism and mutual biotransformations of anthraquinones and
anthrones in rhubarb by human intestinal flora using UPLC-Q-TOF/MS.
J Chromatogr B Analyt Technol Biomed Life Sci. 1104:59–66.
2019.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Piwowarski JP, Granica S, Zwierzyńska M,
Stefańska J, Schopohl P, Melzig MF and Kiss AK: Role of human gut
microbiota metabolism in the anti-inflammatory effect of
traditionally used ellagitannin-rich plant materials. J
Ethnopharmacol. 155:801–809. 2014.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Stolarczyk M, Piwowarski JP, Granica S,
Stefanska J, Naruszewicz M and Kiss AK: Extracts from
Epilobium sp. herbs, their components and gut microbiota
metabolites of Epilobium ellagitannins, urolithins, inhibit
hormone-dependent prostate cancer cells-(LNCaP) proliferation and
PSA secretion. Phytother Res. 27:1842–1848. 2013.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Haddad EH, Gaban-Chong N, Oda K and Sabaté
J: Effect of a walnut meal on postprandial oxidative stress and
antioxidants in healthy individuals. Nutr J. 13(4)2014.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Han QA, Yan C, Wang L, Li G, Xu Y and Xia
X: Urolithin A attenuates ox-LDL-induced endothelial dysfunction
partly by modulating microRNA-27 and ERK/PPAR-γ pathway. Mol Nutr
Food Res. 60:1933–1943. 2016.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Ryu D, Mouchiroud L, Andreux PA, Katsyuba
E, Moullan N, Nicolet-Dit-Félix AA, Williams EG, Jha P, Lo Sasso G,
Huzard D, et al: Urolithin A induces mitophagy and prolongs
lifespan in C. elegans and increases muscle function in rodents.
Nat Med. 22:879–893. 2016.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Chen P, Chen F, Lei J, Li Q and Zhou B:
Activation of the miR-34a-mediated SIRT1/mTOR signaling pathway by
urolithin A attenuates D-galactose-induced brain aging in mice.
Neurotherapeutics. 16:1269–1282. 2019.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Kumar RS, Narasingappa RB, Joshi CG,
Girish TK, Prasada Rao UJ and Danagoudar A: Evaluation of Cassia
tora Linn. against oxidative stress-induced DNA and cell membrane
damage. J Pharm Bioallied Sci. 9:33–43. 2017.PubMed/NCBI View Article : Google Scholar
|
|
43
|
David LA, Maurice CF, Carmody RN,
Gootenberg DB, Button JE, Wolfe BE, Ling AV, Devlin AS, Varma Y,
Fischbach MA, et al: Diet rapidly and reproducibly alters the human
gut microbiome. Nature. 505:559–563. 2014.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Ezenwa VO, Gerardo NM, Inouye DW, Medina M
and Xavier JB: Microbiology. Animal behavior and the microbiome.
Science. 338:198–199. 2012.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Espín JC, Larrosa M, García-Conesa MT and
Tomás-Barberán F: Biological significance of urolithins, the gut
microbial ellagic Acid-derived metabolites: The evidence so far.
Evid Based Complement Alternat Med. 2013(270418)2013.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Selma MV, Beltrán D, García-Villalba R,
Espín JC and Tomás-Barberán FA: Description of urolithin production
capacity from ellagic acid of two human intestinal Gordonibacter
species. Food Funct. 5:1779–1784. 2014.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Clarke G, Sandhu KV, Griffin BT, Dinan TG,
Cryan JF and Hyland NP: Gut reactions: Breaking down
xenobiotic-microbiome interactions. Pharmacol Rev. 71:198–224.
2019.PubMed/NCBI View Article : Google Scholar
|