Introduction
Heart failure (HF) is characterized by abnormal
cardiac structure or function, which leads to the failure of oxygen
delivery or oxygen delivery with increased filling pressure
(1). Chronic HF (CHF) is considered
a progressive syndrome and represents one of the leading causes of
global disability and death (2).
Changes in dietary habits and the increase in the aging population
contribute to the prevalence of CHF (3). Despite progress in the management of
cardiovascular diseases, the morbidity and mortality of CHF
continue to increase (4). Diagnosis
of CHF mainly depends on clinical manifestations, echocardiography
and several circulating biomarkers, such as B-type natriuretic
peptide (BNP) and N-terminal pro-BNP (NT-proBNP). However, the
increased levels of these established biomarkers are also detected
in patients with primary aldosteronism, renal failure, thyroid
disease, lung disease and cirrhosis, and are influenced by the age
and body mass index of patients (5-7).
Therefore, novel reliable biomarkers with high sensitivity and
specificity are necessary for CHF diagnosis. Vascular endothelial
cells (ECs) are critical in maintaining vascular homeostasis via
their roles in regulation of vascular growth, remodeling and
permeability, cell immune response, cell adhesion and angiogenesis
(8). Thus, impairments in the
function of ECs are major causes of cardiovascular diseases
(9). Dysregulation of ECs, such as
abnormal cell proliferation, was determined to be an important
event involved in the development and progression of CHF (10). Thus, novel therapeutic approaches for
CHF need to be investigated based on the methods to improve EC
function, such as studying biomarkers that are involved in the
regulation of EC function.
Emerging studies have shown that long noncoding RNAs
(lncRNAs) play important functional roles in various human
diseases, including cardiovascular diseases (11). LncRNAs are a group of RNAs >200
nucleotides in length and have regulatory roles in a number of
cellular processes, such as cell proliferation, migration, invasion
and apoptosis (12). Aberrant
expression of lncRNAs has been reported in several cardiovascular
diseases, including CHF, and could be involved in disease
initiation and development (13,14).
Increased expression of lncRNA plasmacytoma variant translocation 1
(PVT1) was found in several heart-related abnormalities, such as
cardiac hypertrophy (15) and atrial
fibrosis (16). In addition, Zheng
et al (17) showed the
promotive effects of PVT1 on the angiogenesis of ECs. However, to
the best of our knowledge, the role of PVT1 in CHF has rarely been
investigated.
MicroRNAs (miRs/miRNAs) are another group of
noncoding RNAs of 18-22 nucleotides in length. Accumulatingevidence
indicates the pivotal roles of miRNAs in the diagnosis, prognosis
and therapy of various diseases (18,19).
miRNAs are involved in disease progression by regulating diverse
cellular processes (20). An
increasing number of studies reported that miRNAs act as direct
targets of lncRNAs, thus mediating the function of lncRNAs
(21). miR-190a-5p was identified as
a target of PVT1 in glioma and mediated the effects of PVT1 on
tumor cell proliferation (22).
Decreased circulating levels of miR-190a-5p werereported in HF
cases (23). However, the precise
role of miR-190-5p in CHF remains to be elucidated.
Considering the important effect of EC function on
the development of CHF, the present study sought to investigate the
regulatory effects of PVT1 and miR-190a-5p on EC proliferation.
Furthermore, clinical research was performed to evaluate the
expressional patterns and diagnostic value of circulating PVT1 and
miR-190a-5p in the ECs of patients with CHF. The results of the
present study might provide a novel insight into the mechanisms
underlying the promotive effect of PVT1 on EC function and as
diagnostic biomarkers for CHF.
Materials and methods
Cell culture and transfection
Human umbilical vein endothelial cells (HUVECs) were
obtained from the American Type Culture Collection. Cells were
cultured in DMEM (Invitrogen; Thermo Fisher Scientific, Inc.)
supplemented with 10% FBS (Gibco; Thermo Fisher Scientific, Inc.)
in a humidified atmosphere of 5% CO2 at 37˚C.
Cell transfection was performed using Lipofectamine™
3000 (Invitrogen; Thermo Fisher Scientific, Inc.) according to the
manufacturer's instructions. miR-190a-5p mimic (50 nM;
5'-UGAUAUGUUUGAUAUAUUAGGU-3') and miR-190a-5p inhibitor (50 nM;
5'-ACCUAAUAUAUCAAACAUAUCA-3') were used to regulate the in
vitro expression of miR-190a-5p, while small interfering RNA
(siRNA)-PVT1 (100 nM; 5'-CCCAACAGGAGGACAGCUUTT-3') was used to
knock down the expression of PVT1. miRNA negative control (miR-NC;
50 nM; 5'-UCACAACCUCCUAGAAAGAGUAGA-3') and siRNA NC (100 nM;
5'-UUCUCCGAACGUGUCACGUTT-3') were used as controls. Untreated cells
served as the mock group. All sequences were synthesized by
Shanghai GenePharma Co., Ltd. Subsequent cell experiments were
performed 48 h after transfection.
Patients and blood collection
A total of 92 CHF patients were recruited from Yidu
Central Hospital of Weifang between January 2015 and February 2017.
The patients were enrolled with the following inclusion criteria:
i) All patients were diagnosed with CHF in accordance with the
guidelines of the American Heart Association (24); ii) patients had good compliance and
could cooperate to complete the present study; and iii) had no
infectious diseases, history of myocardial infarction or other
cardiac diseases. The CHF patients included 58 males and 34 females
with a mean age of 62.5±12.6 years (age range of 35-85 years). In
addition, 60 healthy volunteers who underwent physical examination
were enrolled in the present study during the same time period,
including 38 males and 22 females with a mean age of 62.2±12.1
years (age range of 38-82 years). No significant difference in age
and gender was found between the CHF patients and healthy controls.
Venous blood was collected from all participants and centrifuged at
3,000 x g for 10 min at 4˚C for serum isolation. The experimental
procedures were approved by the Ethics Committee of Yidu Central
Hospital of Weifang. Written informed consent was obtained from
each participant.
RNA extraction and reverse
transcription-quantitative PCR (RT-qPCR)
Total RNA was isolated from cells and serum using
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.) according to the manufacturer's instructions. Single-stranded
cDNA was synthesized from 2 µg RNA using SuperScript III Reverse
Transcriptase (cat. no 18080044; Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturer's instructions. The
expression levels of PVT1 and miR-190a-5p were examined by RT-qPCR,
which was performed using a SYBR Green PCR Master Mix kit (cat. no
4364344; Applied Biosystems; Thermo Fisher Scientific, Inc.) on a
7500 Real-Time PCR System (Applied Biosystems; Thermo Fisher
Scientific, Inc.). The following thermocycling conditions were used
for the PCR: Initial denaturation at 95˚C for 5 min; 40 cycles of
95˚C for 10 sec, and 60˚C for 1 min; and a final extension at 72˚C
for 10 min. GAPDH and U6 were used as endogenous controls for PVT1
and miR-190a-5p, respectively. The following primer pairs were used
for the PCR: PVT1 forward, 5'-GGGGAATAACGCTGGTGGAA-3' and reverse,
5'-CCCATGGACATCCAAGCTGT-3'; miR-190a-5p forward,
5'-GCCGAGTGATATGTTTGATAT-3' and reverse, 5'-CTCAACTGGTGTCGTGGA-3';
GAPDH forward, 5'-AGCTGAACGGGAAGCTCACT-3' and reverse,
5'-TGCTTAGCCAAATTCGTTG-3'; and U6 forward, 5'-CTCGCTTCGGCAGCACA-3'
and reverse, 5'-AACGCTTCACGAATTTGCGT-3'. Relative expression levels
of mRNA were calculated using the 2-ΔΔCq method
(25).
Luciferase reporter assay
According to TargetScan (version 7.1; www.targetscan.org/vert_71/) analysis, the sequence of
PVT1 contains a target complementary sequence for miR-190a-5p. PVT1
wild-type (WT) or mutant type (MT) fragments were cloned into the
pMIR-REPORT™ luciferase vector (Ambion; Thermo Fisher Scientific,
Inc.) to construct the luciferase reporter vectors. HUVECs were
seeded and co-transfected with reporter vector and miR-190a-5p
mimic or reporter vector and miR-190a-5p inhibitor using
Lipofectamine™ 3000 (Invitrogen; Thermo Fisher Scientific, Inc.)
according to the manufacturer's instructions. Following 48 h of
transfection, luciferase activity was measured using a Dual
Luciferase Reporter Assay system (Promega Corporation). Renilla
luciferase activity was detected for normalization.
Cell proliferation assay
Cell proliferation was analyzed using a Cell
Counting Kit-8 (CCK-8; Dojindo Molecular Technologies, Inc.).
HUVECs were seeded into 96-well plates at a density of
2x103 cells/well and cultured at 37˚C with 5%
CO2. CCK-8 reagent was added to the cells at time points
of 0, 24, 48 and 72 h with a further 2-h incubation. The absorbance
was measured at a wavelength of 450 nm using a microplate reader
(Omega Bio-Tek, Inc.).
Statistical analysis
Data are presented as the mean ± SD and were
analyzed using SPSS 18.0 (SPSS Inc.) and GraphPad Prism 5.0
(GraphPad Software, Inc.). Comparisons between parameters were
performed using Student's t-test or one-way ANOVA followed by
Tukey's post hoc test. Correlation between parameters was assessed
using the Pearson's correlation coefficient. A receiver operating
characteristic (ROC) curve was plotted to evaluate the diagnostic
value of PVT1 and miR-190a-5p in CHF, and logistics regression
analysis was conducted to obtain the ROC analysis results based on
the combination of PVT1 and miR-190a-5p. P<0.05 was considered
to indicate a statistically significant difference.
Results
PVT1 directly inhibits miR-190a-5p
expression in ECs
The present study investigated the effects of PVT1
on miR-190a-5p expression in ECs. The relative expression of PVT1
was downregulated in ECs in vitro following transfection
with siRNA-PVT1 compared with both mock and siRNA NC groups
(P<0.001; Fig. 1A). As shown in
Fig. 1B, knockdown of PVT1 in ECs
resulted in a significant increase in the expression of miR-190a-5p
compared with mock and siRNA NC groups (P<0.001). PVT1 was
demonstrated to contain a complementary sequence for miR-190a-5p
(Fig. 1C), suggesting the potential
for PVT1 to directly regulate miR-190a-5p. Thus, a luciferase
reporter assay was performed to confirm this interaction in ECs. As
shown in Fig. 1D, the luciferase
activity of the WT group was reduced by the overexpression of
miR-190a-5p (P<0.01) but promoted by the reduction of
miR-190a-5p (P<0.05). However, the luciferase activity of the MT
group did not show significant difference among different
experimental conditions. The data indicated that miR-190a-5p served
as a direct target of PVT1 and could be inhibited by PVT1 in
ECs.
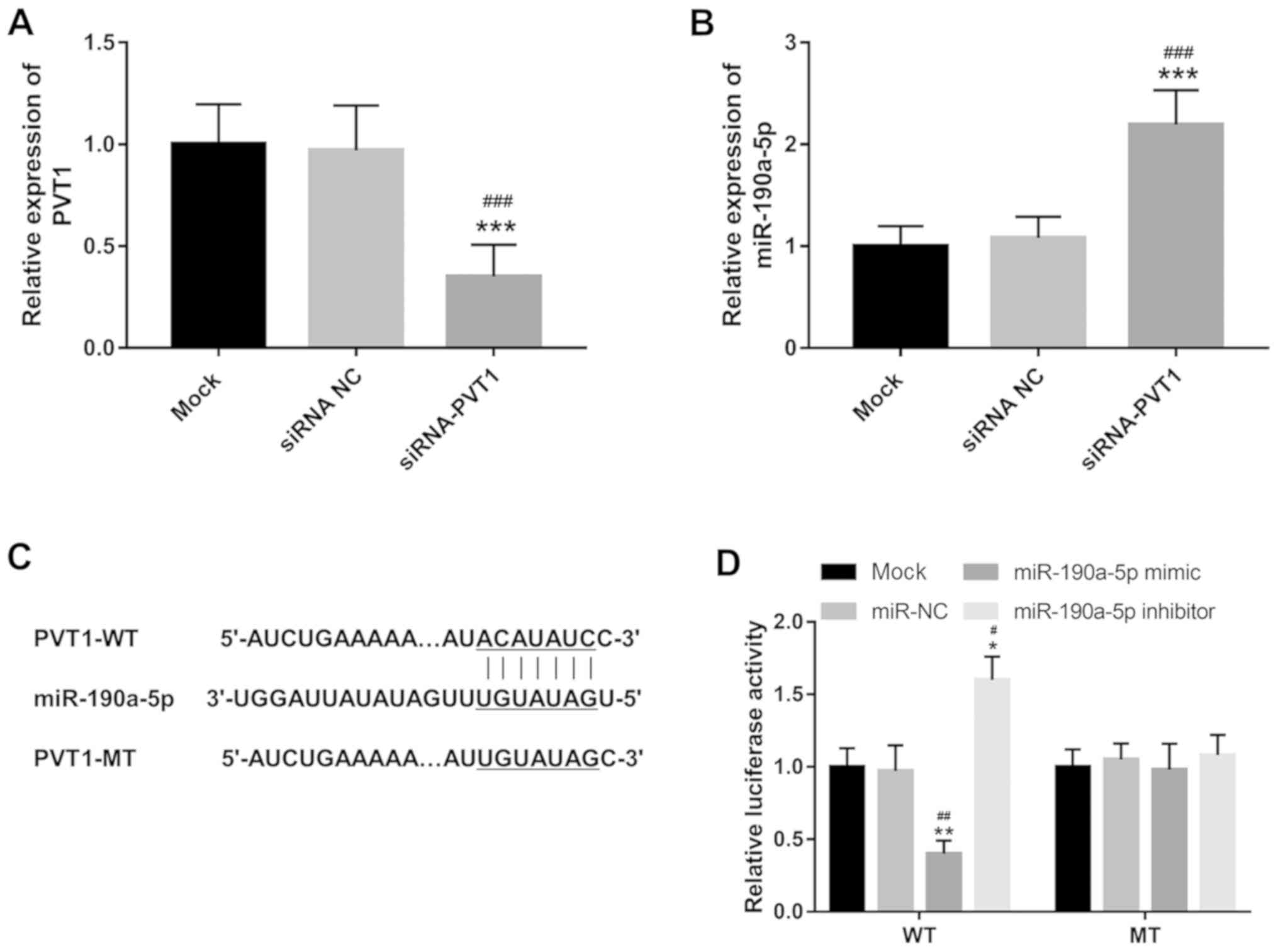 | Figure 1PVT1 directly regulates the expression
of miR-190a-5p in HUVECs. (A) Expression of PVT1 was suppressed by
siRNA-PVT1 in HUVECs. ***P<0.001, vs. the mock group;
###P<0.001 vs. siRNA NC group. (B) Expression of
miR-190a-5p was promoted by the knockdown of PVT1 in HUVECs.
***P<0.001, vs. the mock group;
###P<0.001 vs. siRNA NC group. (C) PVT1 contains a
complementary sequence to miR-190a-5p. (D) Luciferase reporter
assay results confirmed the direct interaction between PVT1 and
miR-190a-5p. *P<0.05 and **P<0.01, vs.
the mock group; #P<0.05 and ##P<0.01
vs. the miR-NC group. HUVEC, human umbilical vein endothelial
cells; PVT1, plasmacytoma variant translocation 1; siRNA-PVT1,
small interfering RNA targeting PVT1, NC, negative control; WT,
wild-type; MT, mutant-type; miR, microRNA. |
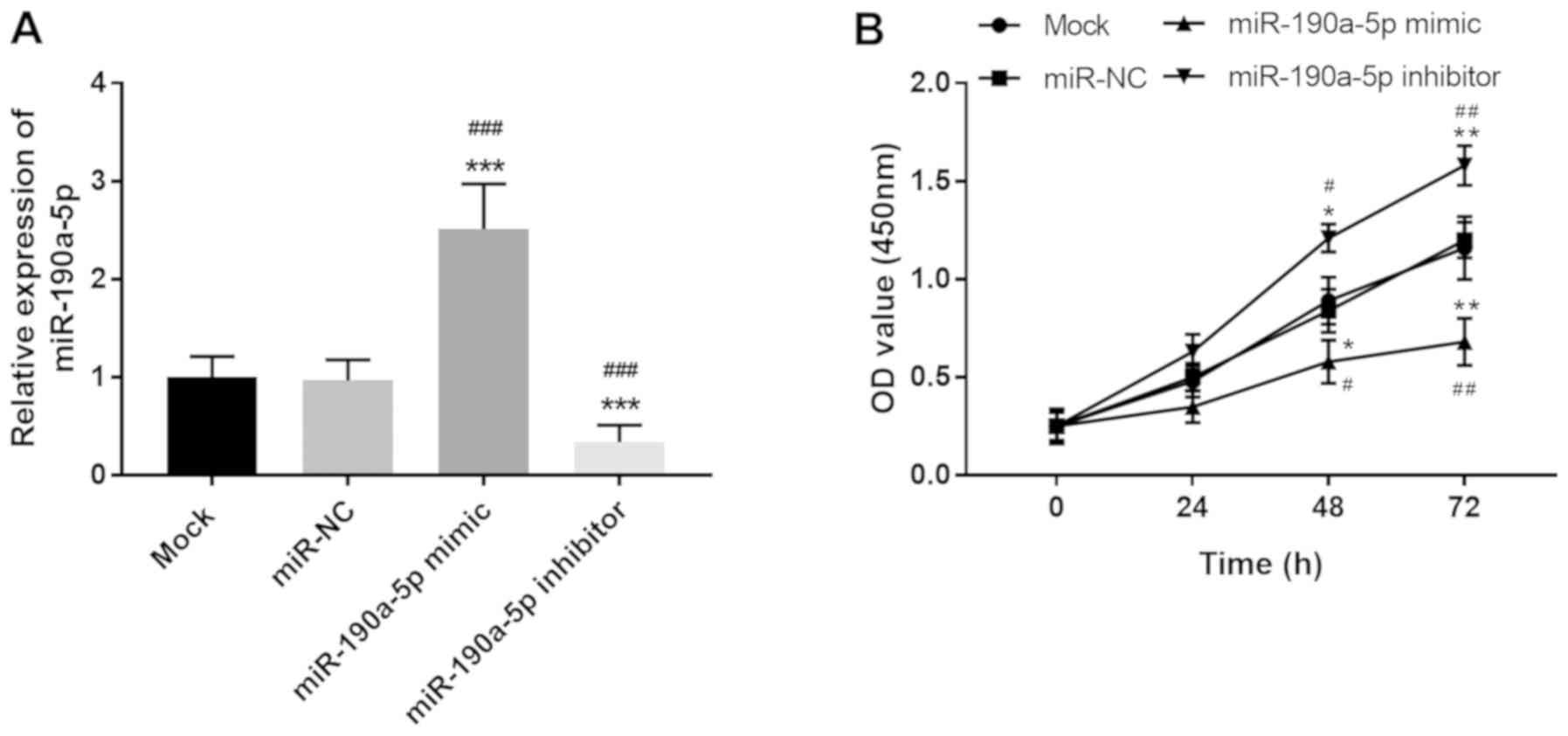
miR-190a-5p suppresses cell
proliferation of HUVECs
Considering the critical regulatory role of PVT1 in
the angiogenesis of ECs (17), the
present study further focused on the effects of miR-190a-5p on the
proliferation of ECs. miR-190a-5p expression in HUVECs was
regulated by cell transfection. miR-190a-5p mimic transfection led
to the overexpression of miR-190a-5p, while miR-190a-5p inhibitor
resulted in the knockdown of miR-190a-5p expression compared with
both mock and miR-NC groups (all P<0.001; Fig. 2A). Based on the CCK-8 assay results,
cell proliferation of HUVECs was markedly suppressed by the
upregulation of miR-190a-5p but enhanced by the downregulation of
miR-190a-5p at both 48 h (P<0.05) and 72 h (P<0.01; Fig. 2B). The results demonstrated that
miR-190a-5p exerted an opposite effect on EC proliferation compared
with PVT1, specifically inhibiting the proliferation of HUVECs.
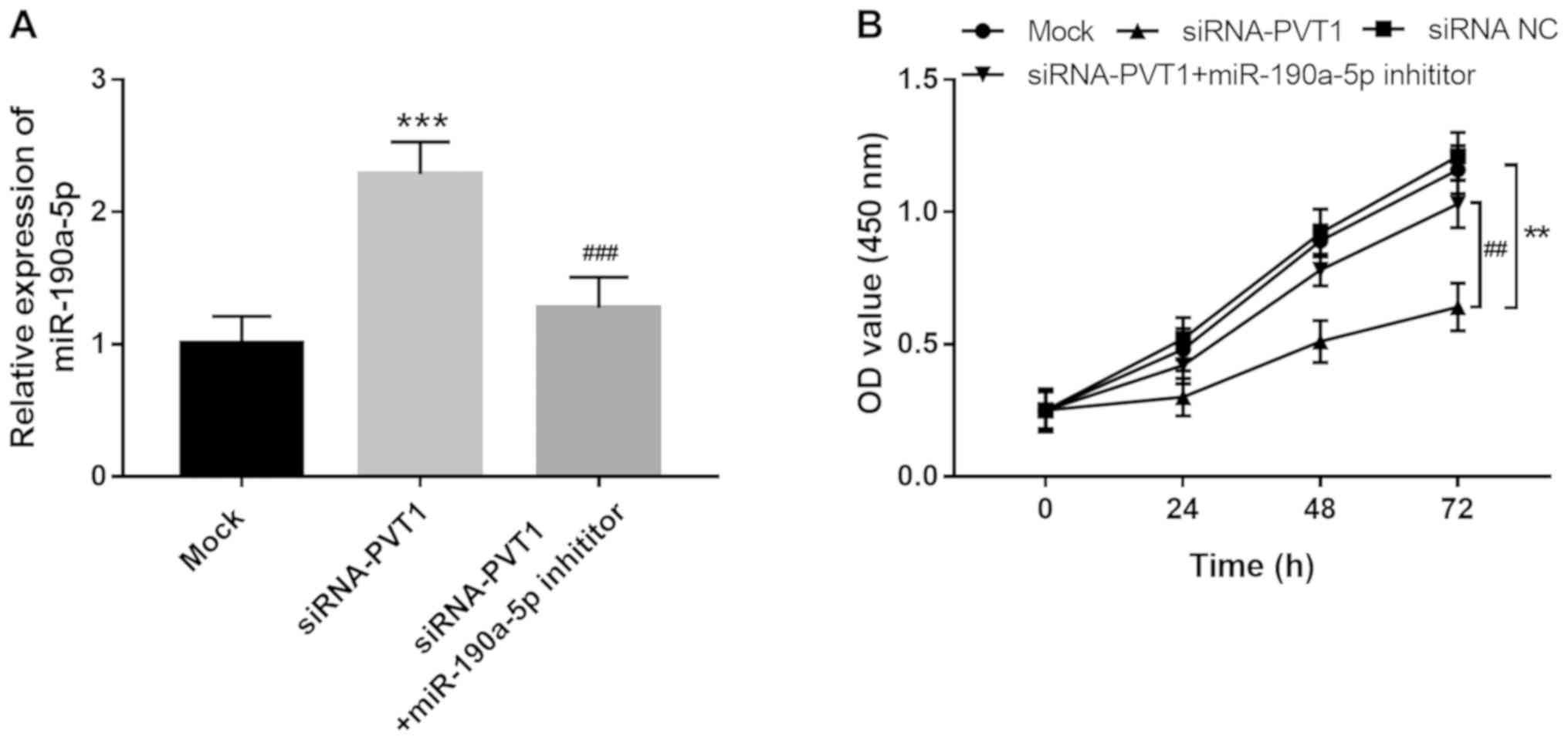
PVT1 facilitates EC proliferation via
regulation of miR-190a-5p
PVT1 was previously determined to be a promoter of
EC angiogenesis (17). Considering
the direct interaction between PVT1 and miR-190a-5p in ECs, the
present study further investigated the role of miR-190a-5p in the
regulatory effects of PVT1 on EC proliferation. As shown in
Fig. 3A, increased expression of
miR-190a-5p induced by the knockdown of PVT1 was reversed by
miR-190a-5p inhibitor transfection (P<0.001). Cell proliferation
assay results showed that the proliferation of HUVECs was
suppressed by the downregulation of PVT1 (P<0.01), but this
inhibitory effect was rescued by inhibiting miR-190a-5p (P<0.01;
Fig. 3B), which suggested that the
promotive effect of PVT1 on EC proliferation might be mediated by
downregulating miR-190a-5p.
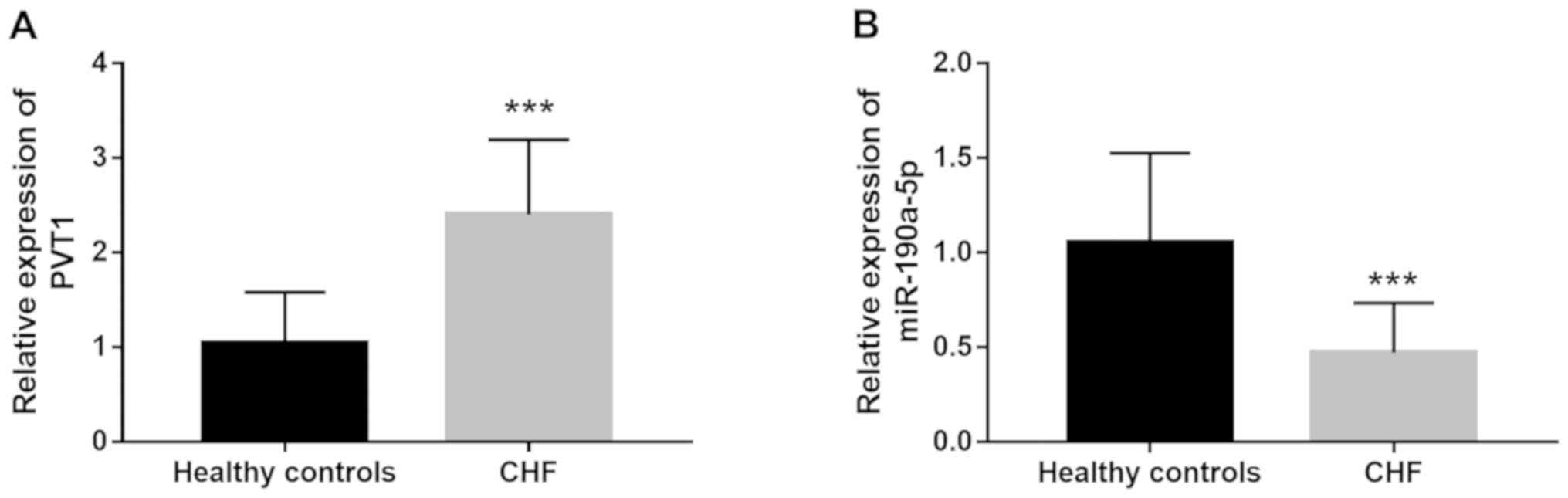
Expression of PVT1 and miR-190a-5p in
CHF patients
Circulating lncRNAs and miRNAs were previously
identified as a group of non-invasive clinical biomarkers for
disease diagnosis (18). In the
present study, serum expression of PVT1 and miR-190a-5p was
measured in patients with CHF. As shown in Fig. 4A, circulating PVT1 expression was
significantly higher in CHF cases compared with healthy individuals
(P<0.001). By contrast, the serum expression of miR-190a-5p
significantly decreased in CHF patients compared with healthy
controls (P<0.001; Fig. 4B).
Dysregulation of circulating PVT1 and miR-190a-5p levels might be a
potential biomarker for CHF diagnosis.
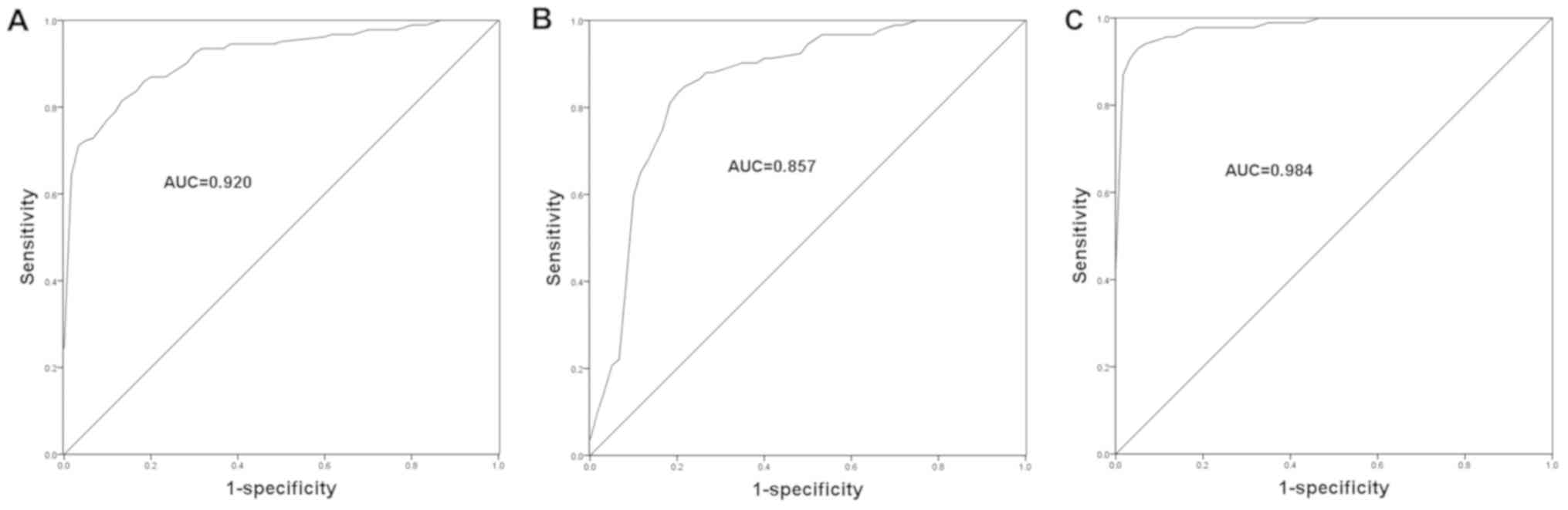
Clinical significance of PVT1 and
miR-190a-5p in the diagnosis of CHF
Considering the dysregulation of PVT1 and
miR-190a-5p in the serum specimens of CHF patients, the current
study aimed to further assess the diagnostic potential of these
molecules in CHF. The ROC curves for CHF patients were plotted
based on the expression levels of PVT1 and miR-190a-5p, and the
area under the curve (AUC) was calculated to reflect diagnostic
accuracy. As shown in Fig. 5, the
ROC curves showed that the AUCs were 0.920 and 0.857 for PVT1
(Fig. 5A) and miR-190a-5p (Fig. 5B), respectively. The best cut-off
values for CHF diagnosis were 2.015 and 0.730 for PVT1 and
miR-190a-5p, respectively. These data indicated the relatively high
diagnostic accuracy of PVT1 and miR-190a-5p. Furthermore, the study
assessed the diagnostic value of the combination of PVT1 and
miR-190a-5p in CHF. As shown in Fig.
5C and Table I, a higher AUC of
0.984 was observed when the two indicators were combined, with
improved sensitivity and specificity. This suggested that the
combined detection of PVT1 and miR-190a-5p might be an efficient
novel strategy for the diagnosis of CHF.
 | Table IDiagnostic value of PVT1 and
miR-190a-5p in chronic heart failure. |
Table I
Diagnostic value of PVT1 and
miR-190a-5p in chronic heart failure.
| Indicator | AUC | Sensitivity, % | Specificity, % |
|---|
| PVT1 | 0.920 | 71.7 | 96.7 |
| miR-190a-5p | 0.857 | 83.7 | 80.0 |
| PVT1 and
miR-190a-5p | 0.984 | 92.4 | 96.7 |
Discussion
The present study focused on the treatment of CHF by
investigating the roles of PVT1 and miR-190a-5p in the regulation
of vascular EC proliferation and disease diagnosis. Knockdown of
PVT1 in ECs led to increased miR-190a-5p expression and PVT1
suppressed cell proliferation by targeting miR-190a-5p. The present
study demonstrated that miR-190a-5p antagonized the effect of PVT1
on ECs proliferation, which provided a novel insight into the
mechanisms underlying the role of PVT1 in the regulation of EC
angiogenesis. The clinical study showed that the expression of PVT1
was upregulated, while the expression of miR-190a-5p was
downregulated in serum samples collected from CHF patients compared
with healthy controls. Furthermore, the ROC curves indicated the
high diagnostic accuracy of PVT1 and miR-190a-5p, particularly when
the two indicators were combined, in patients with CHF. Thus, the
combined detection of PVT1 and miR-190a-5p might be a novel
efficient diagnostic approach for CHF.
Vascular ECs are basic components in the innermost
layer of blood vessels and are considered a pivotal vascular
barrier (26). The dysfunction of
ECs contributes to the occurrence of various cardiovascular
diseases, such as thrombus formation, atherosclerosis, hypertension
and CHF (27-29).
Accumulating evidence indicated that numerous molecules are
involved in the progression of cardiovascular diseases through the
regulation of abnormal cell proliferation of ECs. For example,
Zheng et al (30)
demonstrated that overexpression of miR-155 could suppress the
proliferation of ECs, leading to increased vascular endothelial
permeability and contributing to the progression of
atherosclerosis. Another study by Schober et al (31) also showed the regulatory role of
miR-126-5p in the proliferation of ECs, thereby improving
atherosclerosis. In CHF, miR-214 expression was increased in
patients with this disease and served as a potential therapeutic
target by regulating EC proliferation (32). The aforementioned studies indicated
that the treatment of cardiovascular diseases, including CHF,
should focus on the improvement of impaired EC function.
The present study found suppressed EC proliferation
induced by the reduction in PVT1 expression. Zheng et al
(17) demonstrated the promotive
effect of PVT1 on the proliferation of ECs, consistent with the
present results. PVT1 regulates the expression of miR-190a-5p
during the tumorigenesis of glioma, miR-190a-5p is identified as a
target of PVT1 in the regulation of glioma cell proliferation
(22). However, whether miR-190a-5p
is also a target of PVT1 in ECs remains to be elucidated. In the
present study, ECs with knocked down PVT1 were constructed, and the
results showed that the expression of miR-190a-5p was enhanced. The
luciferase activity data further confirmed the direct interaction
between PVT1 and miR-190a-5p in ECs. miR-190a-5p was reported to
suppress cell proliferation in glioma (22). Similarly, overexpression of
miR-190a-5p inhibited the proliferation of ECs, indicating the role
of miR-190a-5p in the regulation of EC biological function.
Inhibition of EC proliferation induced by PVT1 knockdown was
rescued by downregulation of miR-190a-5p. The results implied that
PVT1 contributes to the cell proliferation of ECs through targeting
miR-190a-5p. Thus, the PVT1/miR-190a-5p axis may be a novel
therapeutic target for the treatment of cardiovascular
diseases.
A study by Wong et al (23) reporteddecreased circulating
miR-190a-5p in HF patients. However, thestudy did not investigate
the precise clinical role of miR-190a-5p in HF. Considering the
role of the PVT1/miR-190a-5p axis in EC proliferation, the
expressional patterns and clinical significance of the two RNAs
were further assessed. Increased expression of PVT1 and decreased
expression of miR-190a-5p were found in the serum specimens of CHF
patients, and the aberrant expression of the two molecules had
relatively high diagnostic accuracy. Currently, the diagnosis of
CHF is limited by the cost of the ultrasound examination and
lowspecificity of established circulating biomarkers, such as BNP
and NT-proBNP (6,7,33). In
the current study, the combination of PVT1 and miR-190a-5p
presented a diagnostic potential for CHF with considerable
sensitivity and specificity, providing a novel potential approach
for the diagnosis of CHF. Aberrant expression levels of lncRNAs and
miRNAs are considered valuable diagnostic biomarkers in various
human diseases, including CHF (34,35).
Increased expression of lncRNAcolorectal neoplasia differentially
expressed-h was identified as a diagnostic and prognostic indicator
in colorectal carcinoma (34).
Decreased plasma lncRNAHOX transcript antisense intergenic RNA
serves as an efficient biomarker for the diagnosis of acute
myocardial infarction (35). The two
circulating lncRNAs NRON and myosin heavy chain associated RNA
transcripts actas two candidate diagnostic markers, with the
ability to distinguish HF patients from healthy controls (36). In a previous study, circulating
upregulated miR-195-3p was described as a potential biomarker for
the diagnosis of HF (7). To further
improve the diagnosis of CHF, the present study provided evidence
for PVT1 and miR-190a-5p as two candidate diagnostic biomarkers,
and the combination of PVT1 and miR-190a-5p as markers suggested a
more effective clinical significance for CHF diagnosis.
Although the present study demonstrated the
regulatory effect of the PVT1/miR-190a-5p axis on EC proliferation,
the biological function of the PVT1/miR-190a-5p axis in ECs
warrants further investigation. In addition, the present study
found aberrant expression of PVT1 and miR-190a-5p in CHF patients.
Thus, the PVT1/miR-190a-5p axis might be involved in CHF
development via regulation of EC function. However, this hypothesis
needs to be verified in future studies, such as investigation into
the role of the PVT1/miR-190a-5p axis in EC apoptosis and
angiogenesis.
Taken together, the present study provided evidence
for the inhibitory effect of miR-190a-5p on the proliferation of
ECs and showed that PVT1 promoted EC proliferation by directly
suppressing miR-190a-5p, implying that the PVT1/miR-190a-5p axis
may serve as a therapeutic target in diseases with EC dysfunction.
Additionally, increased circulating PVT1 levels and decreased
miR-190a-5p expression in CHF patients may serve as two candidate
diagnostic biomarkers, and the combined detection of PVT1 and
miR-190a-5p may be a novel effective approach to diagnose CHF
cases.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the currnt
study are available from the corresponding author on reasonable
request.
Authors' contributions
BS made substantial contributions to conception and
design, acquisition of data, analysis and interpretation of data,
and drafting of the manuscript. MM and JW contributed to
acquisition of data and statistical analysis. SW designed the
study, analysed the data and revised the manuscript. All authors
read and approved the final manuscript.
Ethics approval and consent to
participate
The experimental procedures were approved the Ethics
Committee of Yidu Central Hospital of Weifang. Written informed
consent was obtained from each participant.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Tanai E and Frantz S: Pathophysiology of
heart failure. Compr Physiol. 6:187–214. 2015.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Hoffman TM: Chronic heart failure. Pediatr
Crit Care Med. 17 (8 Suppl 1):S119–S123. 2016.
|
|
3
|
Ford I, Robertson M, Komajda M, Böhm M,
Borer JS, Tavazzi L and Swedberg K: SHIFT Investigators: Top ten
risk factors for morbidity and mortality in patients with chronic
systolic heart failure and elevated heart rate: The SHIFT Risk
Model. Int J Cardiol. 184:163–169. 2015.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Maggioni AP: Epidemiology of heart failure
in Europe. Heart Fail Clin. 11:625–635. 2015.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Heil B and Tang WH: Biomarkers: Their
potential in the diagnosis and treatment of heart failure. Cleve
Clin J Med. 82 (12 Suppl 2):S28–S35. 2015.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Schaub JA, Coca SG, Moledina DG, Gentry M,
Testani JM and Parikh CR: Amino-Terminal Pro-B-type natriuretic
peptide for diagnosis and prognosis in patients with renal
dysfunction: A systematic review and meta-analysis. JACC Heart
Fail. 3:977–989. 2015.PubMed/NCBI View Article : Google Scholar
|
|
7
|
He X, Ji J, Wang T, Wang MB and Chen XL:
Upregulation of Circulating miR-195-3p in Heart Failure.
Cardiology. 138:107–114. 2017.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Eelen G, de Zeeuw P, Treps L, Harjes U,
Wong BW and Carmeliet P: Endothelial cell metabolism. Physiol Rev.
98:3–58. 2018.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Sieve I, Munster-Kuhnel AK and
Hilfiker-Kleiner D: Regulation and function of endothelial
glycocalyx layer in vascular diseases. Vascul Pharmacol. 100:26–33.
2018.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Sager HB, Hulsmans M, Lavine KJ, Moreira
MB, Heidt T, Courties G, Sun Y, Iwamoto Y, Tricot B, Khan OF, et
al: Proliferation and recruitment contribute to myocardial
macrophage expansion in chronic heart failure. Circ Res.
119:853–864. 2016.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Uchida S and Dimmeler S: Long noncoding
RNAs in cardiovascular diseases. Circ Res. 116:737–750.
2015.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Kong Q and Qiu M: Long noncoding RNA
SNHG15 promotes human breast cancer proliferation, migration and
invasion by sponging miR-211-3p. Biochem Biophys Res Commun.
495:1594–1600. 2018.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Dangwal S, Schimmel K, Foinquinos A, Xiao
K and Thum T: Noncoding RNAs in heart failure. Handb Exp Pharmacol.
243:423–445. 2017.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Kumarswamy R, Bauters C, Volkmann I, Maury
F, Fetisch J, Holzmann A, Lemesle G, de Groote P, Pinet F and Thum
T: Circulating long noncoding RNA, LIPCAR, predicts survival in
patients with heart failure. Circ Res. 114:1569–1575.
2014.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Yu YH, Hu ZY, Li MH, Li B, Wang ZM and
Chen SL: Cardiac hypertrophy is positively regulated by long
non-coding RNA PVT1. Int J Clin Exp Pathol. 8:2582–2589.
2015.PubMed/NCBI
|
|
16
|
Cao F, Li Z, Ding WM, Yan L and Zhao QY:
LncRNA PVT1 regulates atrial fibrosis via
miR-128-3p-SP1-TGF-β1-Smad axis in atrial fibrillation. Mol Med.
25(7)2019.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Zheng J, Hu L, Cheng J, Xu J, Zhong Z,
Yang Y and Yuan Z: lncRNA PVT1 promotes the angiogenesis of
vascular endothelial cell by targeting miR26b to activate
CTGF/ANGPT2. Int J Mol Med. 42:489–496. 2018.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Qiu Z, Li H, Wang J and Sun C: MiR-146a
and miR-146b in the diagnosis and prognosis of papillary thyroid
carcinoma. Oncol Rep. 38:2735–2740. 2017.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Zhao Y, Ponnusamy M, Zhang L, Zhang Y, Liu
C, Yu W, Wang K and Li P: The role of miR-214 in cardiovascular
diseases. Eur J Pharmacol. 816:138–145. 2017.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Liu B, Li J and Cairns MJ: Identifying
miRNAs, targets and functions. Brief Bioinform. 15:1–19.
2014.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Zhao L, Han T, Li Y, Sun J, Zhang S, Liu
Y, Shan B, Zheng D and Shi J: The lncRNA SNHG5/miR-32 axis
regulates gastric cancer cell proliferation and migration by
targeting KLF4. FASEB J. 31:893–903. 2017.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Xue W, Chen J, Liu X, Gong W, Zheng J, Guo
X, Liu Y, Liu L, Ma J, Wang P, et al: PVT1 regulates the malignant
behaviors of human glioma cells by targeting miR-190a-5p and
miR-488-3p. Biochim Biophys Acta Mol Basis Dis. 1864:1783–1794.
2018.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Wong LL, Armugam A, Sepramaniam S,
Karolina DS, Lim KY, Lim JY, Chong JP, Ng JY, Chen YT, Chan MM, et
al: Circulating microRNAs in heart failure with reduced and
preserved left ventricular ejection fraction. Eur J Heart Fail.
17:393–404. 2015.PubMed/NCBI View
Article : Google Scholar
|
|
24
|
WRITING COMMITTEE MEMBERS, Yancy CW,
Jessup M, Bozkurt B, Butler J, Casey DE Jr, Drazner MH, Fonarow GC,
Geraci SA, Horwich T, et al: 2013 ACCF/AHA guideline for the
management of heart failure: A report of the American College of
Cardiology Foundation/American Heart Association Task Force on
practice guidelines. Circulation 128: e240-e327, 2013.
|
|
25
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Aman J, Weijers EM, van Nieuw Amerongen
GP, Malik AB and van Hinsbergh VW: Using cultured endothelial cells
to study endothelial barrier dysfunction: Challenges and
opportunities. Am J Physiol Lung Cell Mol Physiol. 311:L453–L466.
2016.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Jufri NF, Mohamedali A, Avolio A and Baker
MS: Mechanical stretch: Physiological and pathological implications
for human vascular endothelial cells. Vascular Cell.
7(8)2015.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Koyama T, Sakurai T, Kamiyoshi A,
Ichikawa-Shindo Y, Kawate H and Shindo T: Adrenomedullin-RAMP2
system in vascular endothelial cells. J Atheroscler Thromb.
22:647–653. 2015.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Franssen C, Chen S, Unger A, Korkmaz HI,
De Keulenaer GW, Tschöpe C, Leite-Moreira AF, Musters R, Niessen
HW, Linke WA, et al: Myocardial microvascular inflammatory
endothelial activation in heart failure with preserved ejection
fraction. JACC Heart Fail. 4:312–324. 2016.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Zheng B, Yin WN, Suzuki T, Zhang XH, Zhang
Y, Song LL, Jin LS, Zhan H, Zhang H, Li JS and Wen JK:
Exosome-mediated miR-155 transfer from smooth muscle cells to
endothelial cells induces endothelial injury and promotes
atherosclerosis. Mol Ther. 25:1279–1294. 2017.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Schober A, Nazari-Jahantigh M, Wei Y,
Bidzhekov K, Gremse F, Grommes J, Megens RT, Heyll K, Noels H,
Hristov M, et al: MicroRNA-126-5p promotes endothelial
proliferation and limits atherosclerosis by suppressing Dlk1. Nat
Med. 20:368–376. 2014.PubMed/NCBI View
Article : Google Scholar
|
|
32
|
Duan Q, Yang L, Gong W, Chaugai S, Wang F,
Chen C, Wang P, Zou MH and Wang DW: MicroRNA-214 Is upregulated in
heart failure patients and suppresses xbp1-mediated endothelial
cells angiogenesis. J Cell Physiol. 230:1964–1973. 2015.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Hill SA, Booth RA, Santaguida PL,
Don-Wauchope A, Brown JA, Oremus M, Ali U, Bustamam A, Sohel N,
McKelvie R, et al: Use of BNP and NT-proBNP for the diagnosis of
heart failure in the emergency department: A systematic review of
the evidence. Heart Fail Rev. 19:421–438. 2014.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Liu T, Zhang X, Gao S, Jing F, Yang Y, Du
L, Zheng G, Li P, Li C and Wang C: Exosomal long noncoding RNA
CRNDE-h as a novel serum-based biomarker for diagnosis and
prognosis of colorectal cancer. Oncotarget. 7:85551–85563.
2016.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Gao L, Liu Y, Guo S, Yao R, Wu L, Xiao L,
Wang Z, Liu Y and Zhang Y: Circulating long noncoding RNA HOTAIR is
an essential mediator of acute myocardial infarction. Cell Physiol
Biochem. 44:1497–1508. 2017.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Xuan L, Sun L, Zhang Y, Huang Y, Hou Y, Li
Q, Guo Y, Feng B, Cui L, Wang X, et al: Circulating long non-coding
RNAs NRON and MHRT as novel predictive biomarkers of heart failure.
J Cell Mol Med. 21:1803–1814. 2017.PubMed/NCBI View Article : Google Scholar
|