Introduction
Acute lung injury (ALI) and acute respiratory
distress syndrome (ARDS) are common in critical medicine, affecting
as many as 200,000 Americans per year and with an overall mortality
rate of approximately 40% (1,2). Acute
respiratory failure is the most serious morbidity stemming from
these pathologies. Mechanical ventilation is a commonly used
modality for respiratory support and has become an indispensable
technique for patient rescue and treatment in both emergency and
critical medicine. However, ventilator use has advantages and
disadvantages. On the one hand, ventilators assist patients in
breathing and provide improved oxygenation. On the other hand,
ventilation results in varying degrees of lung tissue damage, with
a growing body of clinical evidence suggesting that mechanical
ventilation duration is positively correlated with patient
mortality (3-5).
Lung damage induced by mechanical ventilation is referred to as
ventilator-induced lung injury (VILI). Although the mechanisms
underlying VILI remain unclear (6,7), VILI
has been demonstrated to contribute to mortality in patients with
ARDS (8,9).
Phosphorylation is an important post-translational
protein modification, and it is estimated that more than one-third
of all proteins in mammalian cells can be phosphorylated.
Phosphorylated proteins play important roles in cellular functions
(10-14),
including signal transduction, differentiation, proliferation,
cycle regulation, metabolism, transcription and translation,
degradation and survival. Phosphoproteomic analyses identify,
catalog, and characterize phosphorylated proteins (15). Because phosphorylated proteins are
involved in numerous biological processes, the current study aimed
to further clarify the mechanism of lung injury through the
detection of phosphorylated proteins. The present study wish to
further elucidate and clarify the mechanisms of VILI in mice using
phosphoproteomics.
Materials and methods
Animal models
The Animal Use and Care Committee of the Second
Military Medical University of China approved all protocols. Adult
male Institute of Cancer Research (ICR) mice (age, 7-9 weeks;
weight 20-28 g) were obtained from the Experimental Animal Center
of the Second Military Medical University. The ICR mice were
randomly divided between control (CON; n=3) and mechanically
ventilated (VEN; n=3) groups. All mice were housed in individual
cages in a temperature-controlled room (23±1˚C) with 0.05%
CO2 and 12-h light/dark cycles and acclimatized for 1
week prior to experimentation. Food and water were available ad
libitum until 8 h before the experiment. Eight hours prior to
experimentation, access to food was removed, but free access to
water was maintained. All mice were anesthetized using ketamine
(100 mg/kg; Fujian Gutian Yuanhang Medical Co., Ltd.) and xylazine
(10 mg/kg; Sigma-Aldrich; Merck KGaA) via intraperitoneal
injection. Endotracheal intubations were then performed. CON group
mice were able to breathe spontaneously and VEN group mice were
connected to an Inspira ventilator (Harvard Apparatus Ltd.) and
ventilated for 4 h at a volume of 30 ml/kg and a rate of 70
breaths/min. At the 4-h time-point, all mice were administered an
intra-peritoneal dose of ketamine (100 mg/kg) and xylazine (10
mg/kg) and cardiac puncture was carried out as a terminal procedure
to withdraw 0.5-1 ml of blood. The right ventricle was also rapidly
perfused with normal saline (at 4˚C) to remove blood from the
lungs. While still under anesthesia, cervical dislocation was
carried out to euthanize the animals. Lung tissue samples were then
harvested and stored at -80˚C prior to analysis.
Hematoxylin and eosin (H&E)
staining
The lower left lobes of mouse lungs were collected
for examination. Lungs were excised, fixed in 4% paraformaldehyde
for 24 h at 4˚C, dehydrated using a series of increasing
concentrations of ethanol, cleared in xylene, and embedded in
paraffin. Paraffin-embedded lung tissues were cut into 5-µm
sections and stained with H&E before observation using light
microscopy, as previously described (13).
Detection of lung wet to dry
ratio
After euthanasia of the mice, the lower right lobes
of lungs were collected and immediately weighed. This initial value
was termed wet weight (W). Lung tissues were then dried at 80˚C for
48 h and weighed again, with this second value termed dry weight
(D). The W/D ratio was then calculated to evaluate the extent of
lung edema.
Western blotting
Western blotting was performed to determine the
expression levels of target proteins. Briefly, lung tissues from
the middle lobes were homogenized and lysed in ice-cold RIPA buffer
(20 mM Tris pH 7.5, 150 mM NaCl, 50 mM NaF, 1% NP-40, 0.1% DOC,
0.1% SDS, 1 mM EDTA, 1 mM PMSF, 1 mg/ml leupeptin). (Beyotime
Institute of Biotechnology). After incubation for 5 min at 95˚C the
supernatant was harvested and protein concentration quantified
using the bicinchoninic acid assay (Bio-Rad Laboratories, Inc.). A
total of 30 µg of protein from each sample was loaded into a 10%
gel (Bio-Rad Laboratories, Inc.) for SDS-PAGE. The proteins were
subsequently transferred onto PVDF membranes, which were blocked in
5% non-fat milk in 0.1% TBST for 2 h at room temperature. All
antibodies were used at a dilution of 1:1,000. Membranes were then
incubated with primary antibodies against Akt (cat. no. 9272S; Cell
Signaling Technology, Inc.), phospho (p)-Akt and GAPDH (cat. nos.
sc-377556 and sc-47724, respectively; both Santa Cruz
Biotechnology, Inc.) at 4˚C overnight, rinsed in TBST three times
(5 min for each rinse), and incubated with a horseradish
peroxidase-conjugated secondary antibody (cat. no. ab205718;
1:2,000; Abcam) for 1 h at room temperature. Membranes were then
washed with TBST a further 3 times. Visualization was performed
using a chemiluminescence kit (SuperSignal™ West Pico substrate;
Pierce; Thermo Fisher Scientific, Inc.), and images of protein
bands captured Protein densitometry was performed using Quantity
One 4.3.0 imaging software (Bio-Rad Laboratories, Inc.).
Protein extraction
Phosphoprotein profiling was performed using the
Phospho Explorer Array PEX100 kit (Full Moon BioSystems Inc.)
containing 1,318 antibodies. A magnetic serum albumin depletion kit
(cat. no. LSKMAGD12) was purchased from EMD Millipore. Antibody
arrays and cell lysates were prepared according to manufacturer's
protocol. In brief, cell lysates obtained from the upper lobes of
the lungs of CON and VEN group mice were biotinylated usingthe
aforementioned antibody array assay kit. Antibody microarray slides
were treated with a blocking solution for 30 min at room
temperature, rinsed with Milli-Q grade water for 3-5 min, and dried
with compressed nitrogen. Slides were then incubated with
biotin-labeled cell lysates (~100 µg protein per slide) in coupling
solution at room temperature for 2 h. The array slides were washed
4-5 times with 1x Wash Solution and rinsed extensively with Milli-Q
grade water prior to detection of bound biotinylated protein using
Cy3-conjugated streptavidin. The slides were then scanned using a
GenePix 4000 scanner (Molecular Devices LLC) and the images
analyzed using GenePix Pro 6.0 (Molecular Devices, LLC). The
fluorescence signal of each antibody was obtained from antibody
spot fluorescence intensity. Protein phosphorylation expression
profiling was carried out using a Phospho Explorer protein array. A
ratio computation was used to measure the extent of protein
phosphorylation and was calculated as follows: Phosphorylation
ratio=phosphorylation value/dephosphorylation value.
Bioinformatics analysis
Three independent samples for each experimental
group were prepared under the same conditions and used for antibody
array experiments. Array-measured protein expression was normalized
to the internal positive control β-actin (Sigma-Aldrich; Merck
KGaA) and a saline (Sigma-Aldrich; Merck KGaA) negative control.
Statistical significance was determined by either Student's t-test
or ANOVA using MATLAB 9.1. Omics Bean (http://www.omicsbean.cn/) was used to analyze the data
and the resulting data are presented as the mean ± SEM. In cases
where protein expression array identified significant fold changes,
the roles of the proteins were checked for pathway mapping. For
this, Gene Ontology software (http://omicslab.genetics.ac.cn/GOEAST/php/customized_microarray.php)
was used to map these proteins which were subsequently checked for
protein-protein interaction (PPI) mapping using Cytoscape software
(version 3.2.0; https://cytoscape.org/) to identify candidate pathways
and inherent hubs formed. GO and Kyoto Encyclopedia of Genes and
Genomes (KEGG) analysis were used to further understand the
signaling pathways involved in VILI.
Statistical analysis
Differences between control (CON; n=3) and
mechanically ventilated (VEN; n=3) groups were analyzed using
unpaired Student's t-test in SPSS 17.0 (SPSS, Inc.). Only proteins
found to differ in expression level by >30% and that returned a
P<0.05 were considered statistically significant. Protein
phosphorylation expression profiling was carried out using a
Phospho Explorer protein array and Agilent GeneSpring GX 9 software
(Agilent Technologies Inc.) and Metacore bioinformatics software
(https://portal.genego.com/) were used to
test the phosphorylation levels of proteins.
Results
Confirmation of the VILI mouse
model
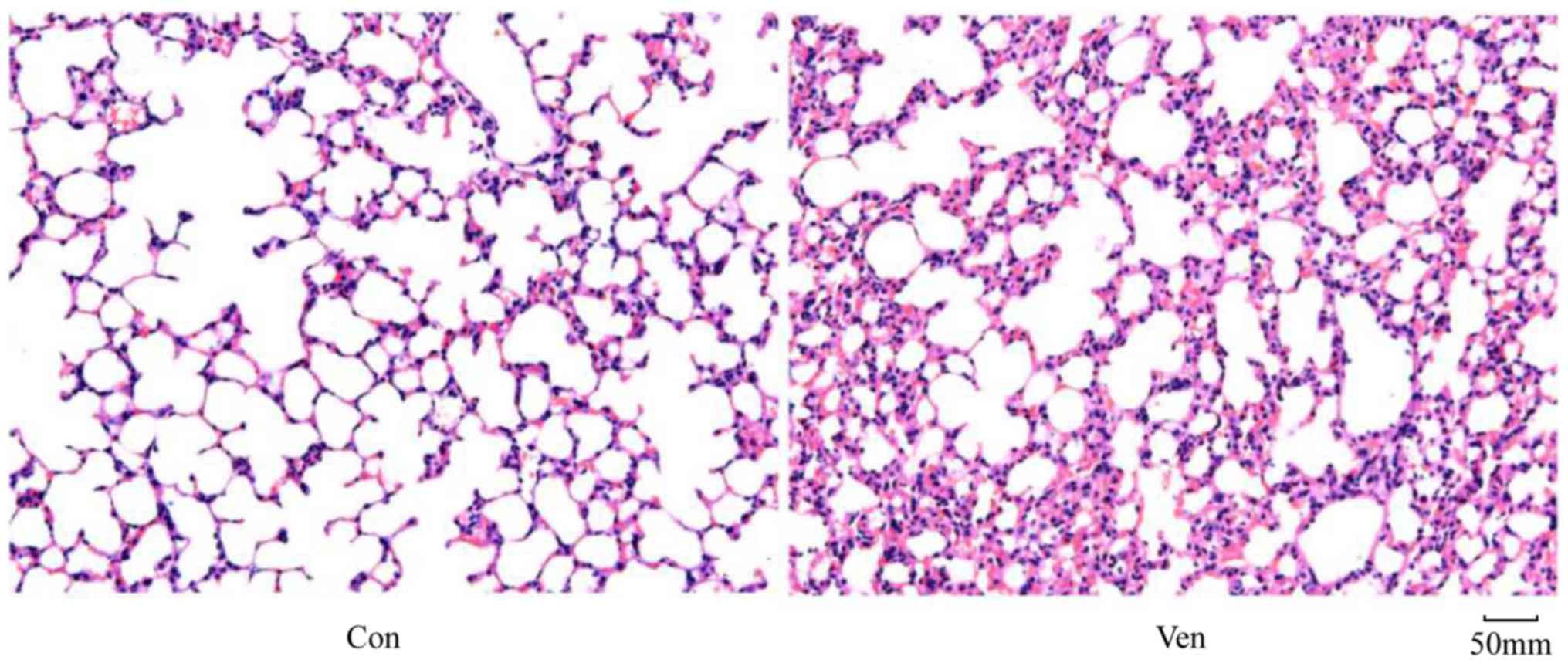
In order to determine whether the application of
mechanical ventilation induced lung injury in mice, lung morphology
was examined in VEN and CON mice post-mortem using H&E
staining. While the CON group demonstrated normal lung structure
and minimal inflammatory cell presence (Fig. 1), the VEN group showed neutrophil
sequestration and infiltration around the pulmonary vessels and the
airway, specifically distributed in the alveolar and interstitial
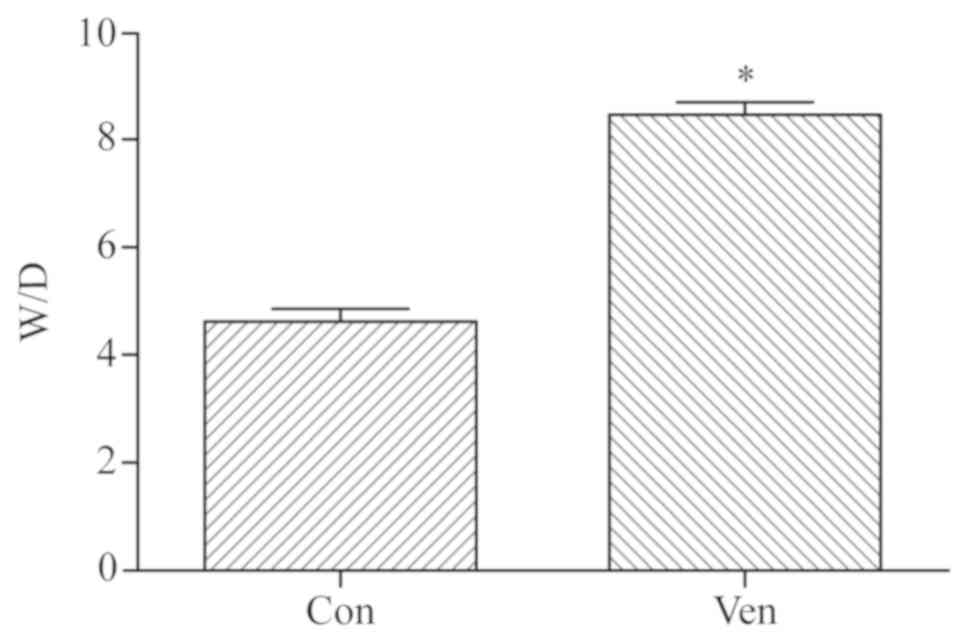
spaces (Fig. 1). Edema extent was
examined using the W/D weight ratio, and the results indicated that
edema extent was significantly elevated in the VEN group compared
with the CON group (Fig. 2;
P<0.05).
Proteomic analysis
Raw protein expression values were normalized and
then subjected to a fold-change cut-off of >|1.5|.
Fold-differences in protein expression with P<0.05 were
considered significant. Notable differences in protein expression
between CON and VEN group mice were observed in 115 proteins, with
37 increased and 78 decreased protein levels (Table SI). Gene changes are presented as a
heatmap (Fig. 3).
Gene Ontology (GO) and Kyoto
Encyclopedia of Genes and Genomes (KEGG) analysis
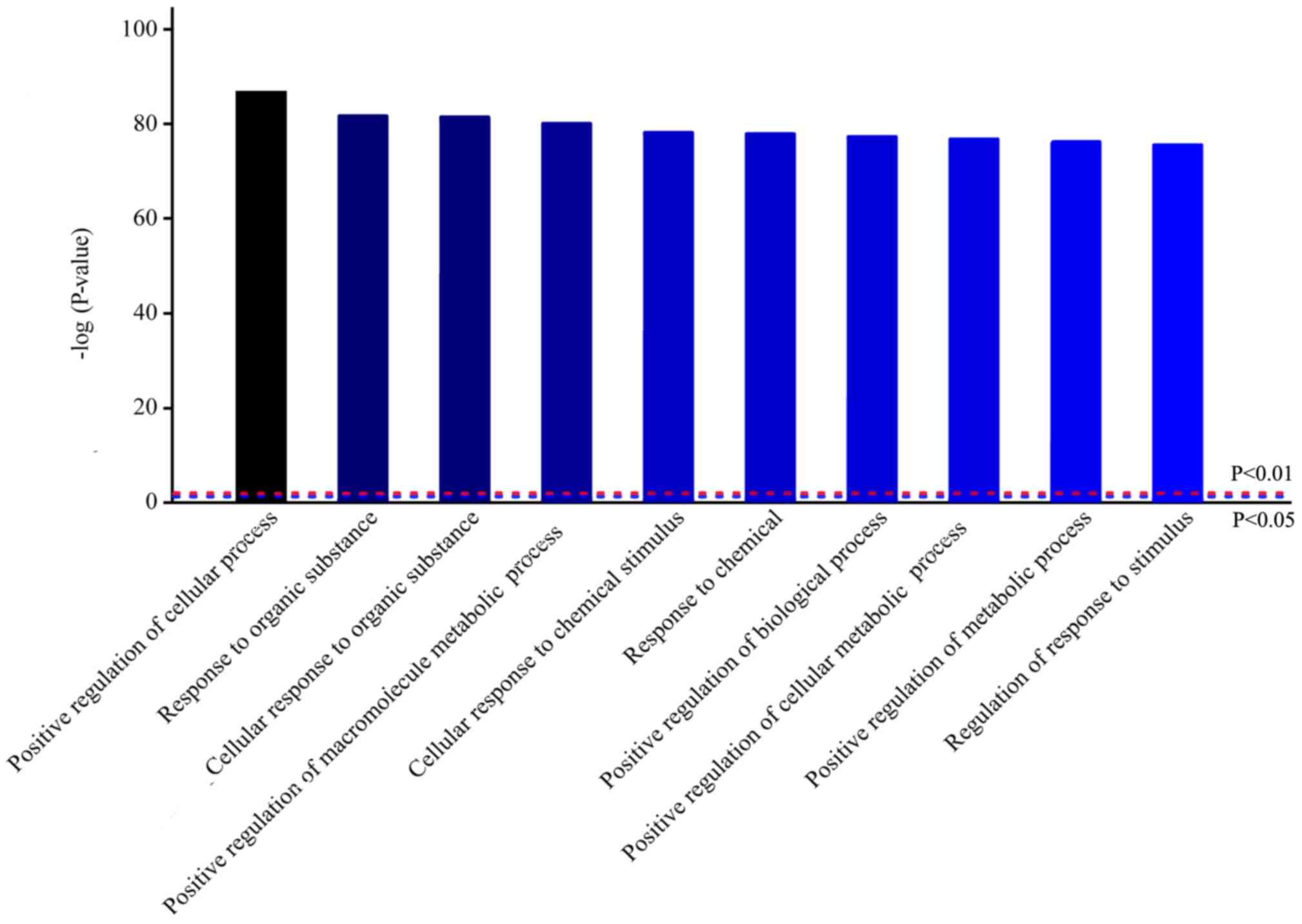
To further understand the signaling pathways
involved in VILI, bioinformatics analysis was performed. Homologous
analysis of differentially expressed proteins based on GO was
performed, which showed that differences in protein expression
levels observed in VEN group mice were primarily in biological
processes (Fig. 4). Biological
process analysis found that most of these proteins were involved in
‘positive regulation of cellular processes’, ‘response to organic
substance’ and ‘positive regulation of metabolic process’.
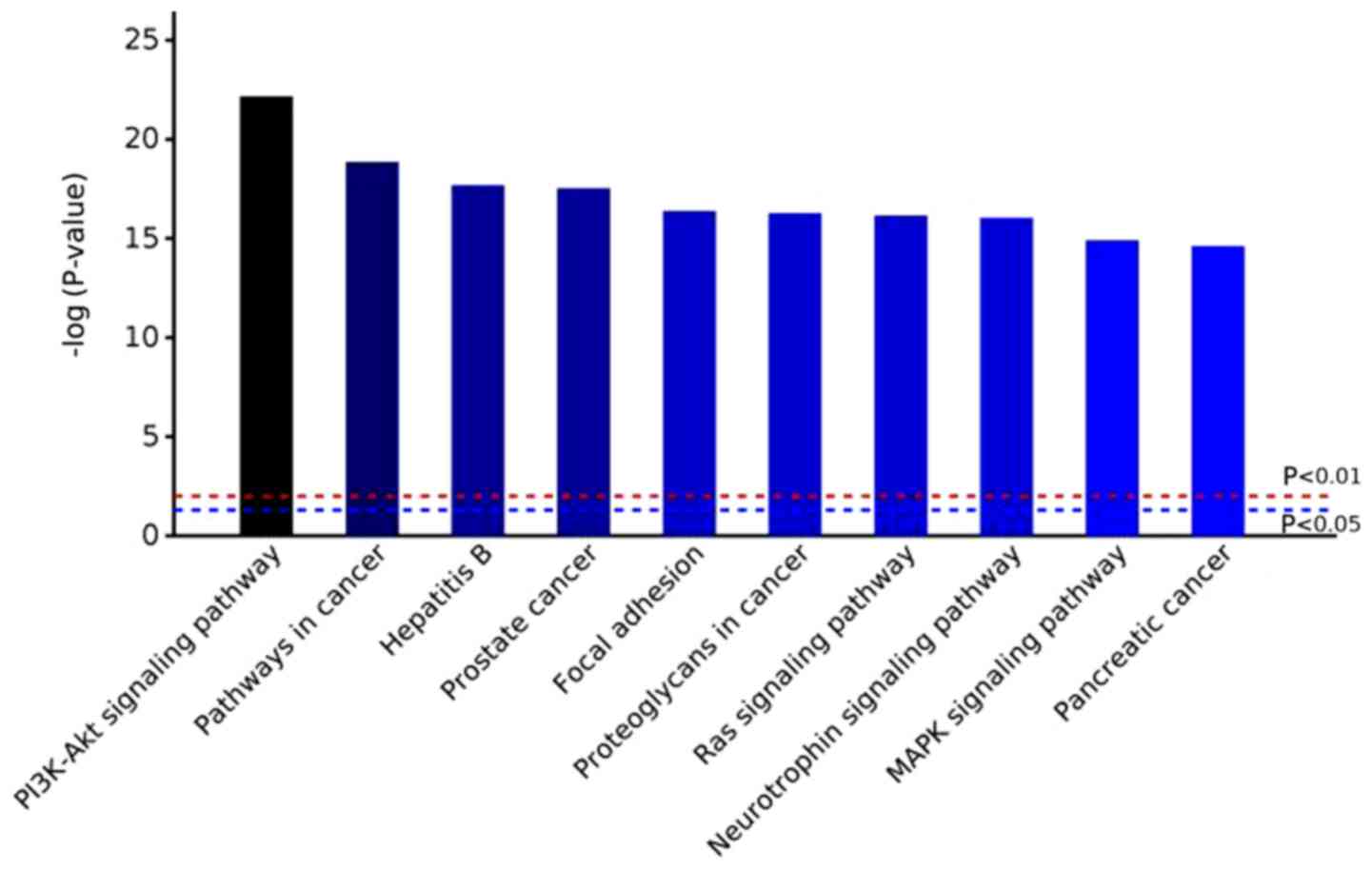
Based on GO analysis results, potential pathways
involved in VILI and control group were investigated using KEGG
pathway analysis (Fig. 5). In VEN
group mice, the PI3K-AKT signaling pathway was the most markedly
altered. Additionally, ‘pathways in cancer’, ‘focal adhesion’,
‘MAPK signaling pathway’ and ‘Ras signaling pathway’ were also
significantly enriched (Fig. 5;
Table SII; and
Figs. S1-S5).
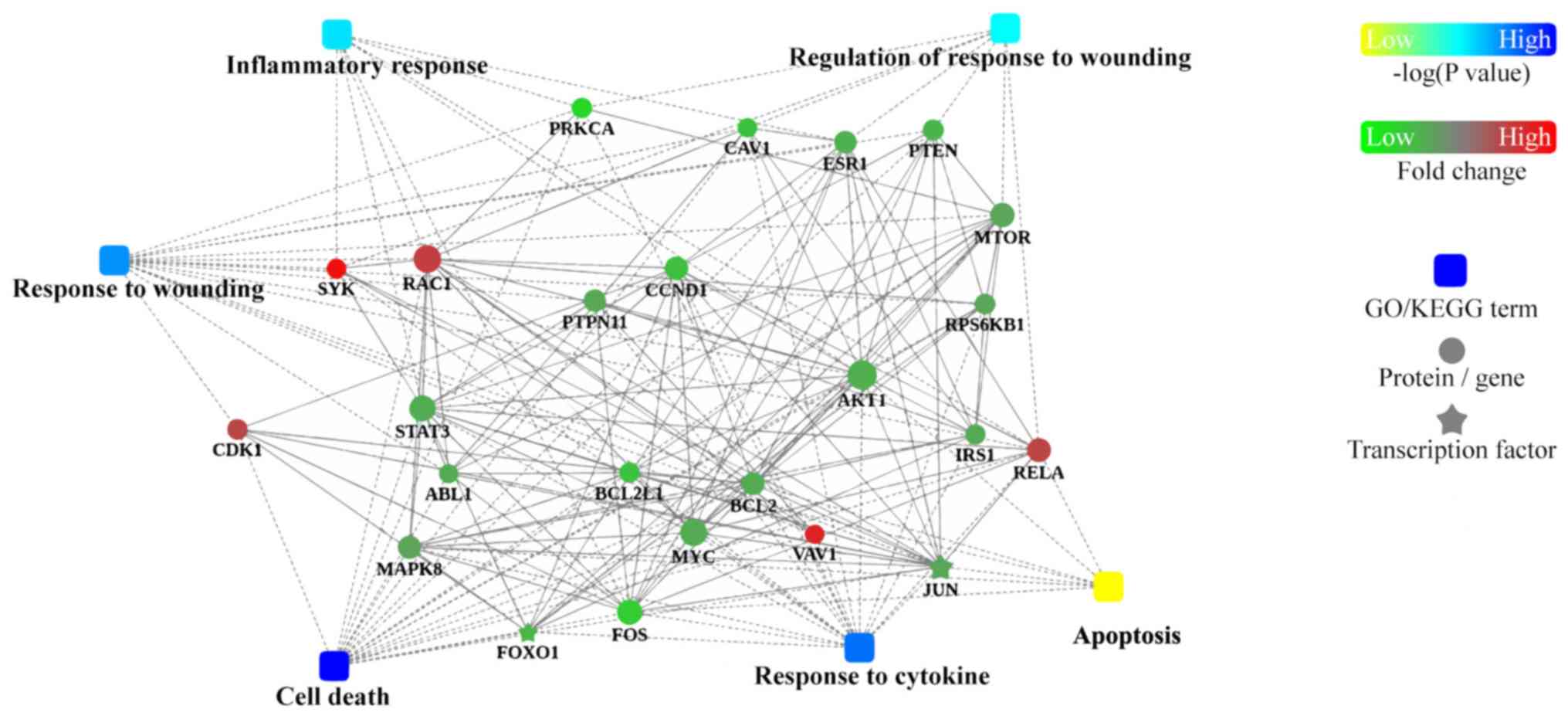
PPI analysis
To further understand how the identified proteins of
interest interact with each other, bioinformatics analysis using
Cytoscape was performed to visualize a PPI network (Fig. 6). This analysis revealed that hub
proteins were enriched in ‘inflammatory response’ and ‘regulation
of response to wounding’ in VILI mice.
Validation
In order to confirm the proteomics, GO, and KEGG
analyses, western blotting was performed to examine AKT
phosphorylation levels in VEN and CON mice. AKT phosphorylation was
significantly reduced in the VEN group compared with the CON group
(Fig. 7; CON group p-AKT/AKT ratio,
0.758±0.055; VEN group, 0.387±0.037; P<0.05).
Discussion
Numerous studies have shown that protein
phosphorylation and dephosphorylation play important roles in a
range of physiological and biochemical processes, including cell
cycle regulation, signal transduction, neuronal growth and memory
processes (10,11). Therefore, exploring the molecular
mechanisms underlying protein phosphorylation is critical in
examining disease mechanisms and identifying potential therapeutic
targets. Since approximately 30% of proteins in eukaryotic cells
can be phosphorylated, each protein kinase selectively modifies a
specific substrate protein to ensure signal transduction within a
complex cellular environment (8,10-17).
The results of the present study suggest that phosphorylated
proteins are involved in VILI, in particular, phosphorylation of
proteins involved in PI3K-AKT and MAPK signaling pathways.
VILI is essentially caused by an uncontrolled
inflammatory response. Abnormal mechanical tension in the lung, due
to artificial ventilation, initiates inflammatory signal
transduction in inflammatory cells such as neutrophils and
macrophages (18-21),
resulting in cell activation and the promotion of phagocytic
function, as well as the synthesis and release of inflammatory
mediators such as interleukin (IL)-16, IL-10, and tumor necrosis
factor-α (1). PI3K activation has
been found to regulate neutrophil activation observed in ALI
(18), and both pharmacological and
induced pluripotent stem cell-mediated inhibition of the PI3K
pathway have been documented to ameliorate inflammatory responses
and inflammatory cell activation associated with VILI (19,20).
PI3K-AKT-mediated reactive oxygen species (ROS) production has also
been linked to VILI (20). Moreover,
Wnt/β-catenin signaling, which can be induced by PI3K pathway
activation, has been shown to be activated by mechanical
ventilation and has been found to have a role in the promotion of
inflammation and fibrosis in otherwise healthy rat lungs (21).
The MAPK pathway is likewise involved in a myriad of
cellular and systemic processes and mechanisms. In the context of
VILI, mechanical stress and stretching of the alveolar epithelium
activates epidermal growth factor receptor (EGFR)-mediated MAPK
signaling, resulting in elevated pulmonary permeability (22,23).
Similarly, inhibition of EGFR-mediated MAPK signaling has been
shown to reduce lung inflammation (23). Mechanical stress has been shown to
induce the release of ROS through MAPK-dependent pathway activation
of xanthine oxidoreductase (24).
Finally, MAPK pathway signaling has been demonstrated to facilitate
microRNA-127-induced inflammation and pulmonary permeability
(22-25).
In the present study, bioinformatics-based protein phosphorylation
analysis suggested that the ‘MAPK signaling pathway’ was activated
in VILI. Focal adhesion formation and behavior has been
demonstrated to be pivotal to pulmonary epithelial function, and
has been linked with modulation of ALI injury severity (15). Focal adhesions (FAs), large
macromolecular complexes mediating interactions between cells and
the extracellular matrix (ECM), have been linked to the development
of pulmonary fibrosis and injury, as FAs play a part in mediating
epithelial function, and epithelial dysfunction is a hallmark of
pulmonary injury (16). The
inhibition of focal adhesion kinase (FAK), a protein that is
activated by integrin-ECM interactions and fibroblast-secreted
transforming growth factor-β (TGF-β), has been shown to exacerbate
bleomycin-induced lung injury in mice (16). Moreover, FA formation, FAK
phosphorylation, and FA arrangement and rearrangement have been
shown to mediate preservation of pulmonary vascular barrier
function (17). In the present
study, the phosphorylation of FA-associated proteins was found to
be significantly influenced by VILI. In particular,
3-phosphoinositide-dependent protein kinase 1 (PDK1)
phosphorylation was elevated in the VEN group. PDK1 is a downstream
kinase of PI3K, and has been linked with cell adhesion and the
organization of structural fibers and tubules (26,27).
PDK1 has also been suggested to play a role in fibroblast-mediated
pulmonary fibrosis, acting as a secondary messenger to activate Akt
upon TGFβ-mediated PI3K activation (27). However, PDK1 activation has also been
demonstrated to be protective, when in conjunction with
serum/glucocorticoid-induced protein kinase 1, in an in
vitro cellular model (28).
Given the complexity of the PI3K-PDK1-AKT pathway and the diverse
range of mechanisms that it modulates, more research is required to
clarify how this pathway modulates FA formation during health and
injury.
The results of the present study suggested that the
expression level of RAC1 was upregulated in VEN mice as shown in
Table SII. Rac is a protein that
mediates epithelial barrier integrity through interactions with
microtubules (29). Given that
epithelial dysfunction is a principle element of VILI, changes in
Rac levels were anticipated as Rac upregulation and activation in
lung injury models is typically linked to protective mechanisms
(30). As such, the elevated
phosphorylation observed in VEN group mice may be a compensatory
mechanism in response to physical damage. In-depth pathway analysis
and functional investigation is required to fully characterize the
role of Rac in VILI.
While the present study was limited by the small
sample size and the fact that only AKT phosphorylation was assessed
in a validation study, the results of this study suggest several
candidate cellular mechanisms that may be involved in VILI. Future
studies will focus on determining how other elements of the
PI3K-AKT pathway, as well as other potential candidate pathways,
may be involved in VILI. Determining the precise roles of specific
proteins, as well as establishing proof of principle for these
causal relationships, will be a primary focus for future studies to
establish potential targets for future therapeutics against
VILI.
Supplementary Material
‘PI3K-Akt signaling pathway’ is the
top Kyoto Encyclopedia of Genes and Genomes enriched pathway in VEN
vs. CON group. Red represents the proteins with upregulated
phosphorylation levels in the VEN group. Blue represents the
proteins with downregulated phosphorylation levels in the VEN
group. Green represents unaltered protein phosphorylation levels in
the VEN group. CON, control; VEN, mechanically ventilated.
‘Pathways in cancer’ is the second
most enriched Kyoto Encyclopedia of Genes and Genomes pathway in
VEN vs. CON group. Red represents the proteins with upregulated
phosphorylation levels in the VEN group. Blue represents the
proteins with downregulated phosphorylation levels in the VEN
group. Green represents unaltered protein phosphorylation levels in
the VEN group. CON, control; VEN, mechanically ventilated.
‘Hepatitis B’ is the third most
enriched Kyoto Encyclopedia of Genes and Genomes pathway in VEN vs.
CON group. Red represents the proteins with upregulated
phosphorylation levels in the VEN group. Blue represents the
proteins with downregulated phosphorylation levels in the VEN
group. Green represents unaltered protein phosphorylation levels in
the VEN group. CON, control; VEN, mechanically ventilated.
‘Prostate cancer’ is the foruth most
enriched Kyoto Encyclopedia of Genes and Genomes pathway in VEN vs.
CON group. Red represents the proteins with upregulated
phosphorylation levels in the VEN group. Blue represents the
proteins with downregulated phosphorylation levels in the VEN
group. Green represents unaltered protein phosphorylation levels in
the VEN group. CON, control; VEN, mechanically ventilated.
‘Focal adhesion’ is the fifth most
enriched Kyoto Encyclopedia of Genes and Genomes pathway in VEN vs.
CON group. Red represents the proteins with upregulated
phosphorylation levels in the VEN group. Blue represents the
proteins with downregulated phosphorylation levels in the VEN
group. Green represents unaltered protein phosphorylation levels in
the VEN group. CON, control; VEN, mechanically ventilated.
Differentially phosphorylated protein
UniProt IDs and fold changes between CON and VEN groups.
Pathway summary of proteins
differentially expressed in the VEN group compared with the CON
group.
Acknowledgments
The authors would like to thank Mr Ye Changlin from
the Shanghai Institute of Physical Education for his technical
support.
Funding
This study was funded by the Shanghai Municipal
Commission of Health and Family Planning Funding for Key Developing
Disciplines (grant no. 2015ZB0101) and the Multi-Center Clinical
Study of Pre-operative Pain Facilitation and Post-operative
Cognitive Dysfunction, Joint Research Project of Pudong New Area
Health and Family Planning Commission (grant no. PW2015D-3).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
RR and WY made substantial contributions to
conception and design; ZR, HD and JD were involved in acquisition
of data, analysis and interpretation of data; RR and JD were
involved in drafting the manuscript; WY were involved in revising
it critically for important intellectual content; all authors gave
final approval of the version to be published.
Ethics approval and consent to
participate
All protocols were approved by the Animal Use and
Care Committee of the Second Military Medical University of China.
The animal production license number was SYXK (Shanghai)
2012-0003.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Rezoagli E, Fumagalli R and Bellani G:
Definition and epidemiology of acute respiratory distress syndrome.
Ann Transl Med. 5(282)2017.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Liu KD and Matthay MA: Advances in
critical care for the nephrologist: Acute lung injury/ARDS. Clin J
Am Soc Nephrol. 3:578–586. 2008.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Kuipers MT, van der Poll T, Schultz MJ and
Wieland CW: Bench-to-bedside review: Damage-associated molecular
patterns in the onset of ventilator-induced lung injury. Crit Care.
15(235)2011.PubMed/NCBI View
Article : Google Scholar
|
|
4
|
Rocco PR, Dos Santos C and Pelosi P:
Pathophysiology of ventilator-associated lung injury. Curr Opin
Anaesthesiol. 25:123–130. 2012.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Frank JA and Matthay MA: Science review:
Mechanisms of ventilator-induced injury. Crit Care. 7:233–241.
2003.PubMed/NCBI View
Article : Google Scholar
|
|
6
|
Silva PL, Negrini D and Rocco PR:
Mechanisms of ventilator-induced lung injury in healthy lungs. Best
Pract Res Clin Anaesthesiol. 29:301–313. 2015.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Han GJ, Li JQ, Pan CG, Sun JX, Shi ZX, Xu
JY and Li MQ: Experimental study of airway pressure release
ventilation in the treatment of acute respiratory distress
syndrome. Exp Ther Med. 14:1941–1946. 2017.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Manning G, Whyte DB, Martinez R, Hunter T
and Sudarsanam S: The protein kinase complement of the human
genome. Science. 298:1912–1934. 2002.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Gattinoni L, Tonetti T and Quintel M:
Regional physiology of ARDS. Crit Care. 21 (Suppl
3)(S312)2017.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Alonso A, Sasin J, Bottini N, Friedberg I,
Friedberg I, Osterman A, Godzik A, Hunter T, Dixon J and Mustelin
T: Protein tyrosine phosphatases in the human genome. Cell.
117:699–711. 2004.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Hunter T: Signaling-2000 and beyond. Cell.
100:113–127. 2000.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Charbonneau H and Tonks NK: 1002 protein
phosphatases? Annu Rev Cell Biol. 8:463–493. 1992.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Yanjun Liu, Xuejun Lv, Wei Zhao, Ming Dong
Hu, Yuying Li, Guanchong Wang, Jiancheng Xu and Guisheng Qian:
Sirt1 protects against lipopolysaccharide-induced injury in mouse
type II alveolar epithelial cells by deacetylating RelA/p65 subunit
of nuclear factor-κB. Journal of the Third Military Medical
University. 39:1415–1421. 2017.
|
|
14
|
Cohen P: The origins of protein
phosphorylation. Nat Cell Biol. 4:E127–E130. 2002.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Suwanmajo T and Krishnan J: Mixed
mechanisms of multi-site phosphorylation. J R Soc Interface.
12(20141405)2015.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Wheaton AK, Agarwal M, Jia S and Kim KK:
Lung epithelial cell focal adhesion kinase signaling inhibits lung
injury and fibrosis. Am J Physiol Lung Cell Mol Physiol.
312:L722–L730. 2017.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Wang L, Bittman R, Garcia JG and Dudek SM:
Junctional complex and focal adhesion rearrangement mediates
pulmonary endothelial barrier enhancement by FTY720 S-phosphonate.
Microvasc Res. 99:102–109. 2015.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Li LF, Liu YY, Yang CT, Chien Y, Twu NF,
Wang ML, Wang CY, Huang CC, Kao KC, Hsu HS, et al: Improvement of
ventilator-induced lung injury by IPS cell-derived conditioned
medium via inhibition of PI3K/Akt pathway and IP-10-dependent
paracrine regulation. Biomaterials. 34:78–91. 2013.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Uhlig U, Fehrenbach H, Lachmann RA,
Goldmann T, Lachmann B, Vollmer E and Uhlig S: Phosphoinositide
3-OH kinase inhibition prevents ventilation-induced lung cell
activation. Am J Respir Crit Care Med. 169:201–208. 2004.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Spassov SG, Donus R, Ihle PM,
Engelstaedter H, Hoetzel A and Faller S: Hydrogen sulfide prevents
formation of reactive oxygen species through PI3K/Akt signaling and
limits ventilator-induced lung injury. Oxid Med Cell Longev.
2017(3715037)2017.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Villar J, Cabrera NE, Valladares F, Casula
M, Flores C, Blanch L, Quilez ME, Santana-Rodríguez N, Kacmarek RM
and Slutsky AS: Activation of the Wnt/β-catenin signaling pathway
by mechanical ventilation is associated with ventilator-induced
pulmonary fibrosis in healthy lungs. PLoS One.
6(e23914)2011.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Otulakowski G, Engelberts D, Gusarova GA,
Bhattacharya J, Post M and Kavanagh BP: Hypercapnia attenuates
ventilator-induced lung injury via a disintegrin and
metalloprotease-17. J Physiol. 592:4507–4521. 2014.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Bierman A, Yerrapureddy A, Reddy NM,
Hassoun PM and Reddy SP: Epidermal growth factor receptor (EGFR)
regulates mechanical ventilation-induced lung injury in mice.
Transl Res. 152:265–272. 2008.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Abdulnour RE, Peng X, Finigan JH, Han EJ,
Hasan EJ, Birukov KG, Reddy SP, Watkins JE III, Kayyali US, Garcia
JG, et al: Mechanical stress activates xanthine oxidoreductase
through MAP kinase-dependent pathways. Am J Physiol Lung Cell Mol
Physiol. 291:L345–L353. 2006.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Li Q, Ge YL, Li M, Fang XZ, Yuan YP, Liang
L and Huang SQ: miR-127 contributes to ventilator-induced lung
injury. Mol Med Rep. 16:4119–4126. 2017.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Medina-Tato DA, Ward SG and Watson ML:
Phosphoinositide 3-kinase signalling in lung disease: Leucocytes
and beyond. Immunology. 121:448–461. 2007.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Chang W, Wei K, Ho L, Berry GJ, Jacobs SS,
Chang CH and Rosen GD: A critical role for the mTORC2 pathway in
lung fibrosis. PLoS One. 9(e106155)2014.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Caohuy H, Yang Q, Eudy Y, Ha TA, Xu AE,
Glover M, Frizzell RA, Jozwik C and Pollard HB: Activation of
3-phosphoinositide-dependent kinase 1 (PDK1) and serum- and
glucocorticoid-induced protein kinase 1 (SGK1) by short-chain
sphingolipid C4-ceramide rescues the trafficking defect of
ΔF508-cystic fibrosis transmembrane conductance regulator
(ΔF508-CFTR). J Biol Chem. 289:35953–35968. 2014.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Karki P and Birukova AA:
Microtubules-associated Rac regulation of endothelial barrier: A
role of Asef in acute lung injury. J Investig Med. 65:1089–1092.
2017.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Birukova AA, Wu T, Tian Y, Meliton A,
Sarich N, Tian X, Leff A and Birukov KG: Iloprost improves
endothelial barrier function in lipopolysaccharide-induced lung
injury. Eur Respir J. 41:165–176. 2013.PubMed/NCBI View Article : Google Scholar
|