Introduction
Chronic kidney disease, especially end-stage renal
disease, seriously threatens the life and health of humans
(1). At present, blood dialysis and
kidney transplantation are two of the most commonly used strategies
for the treatment of end-stage renal disease. Although blood
dialysis can extend the life of patients, it can seriously
influence their quality of life, and renal transplantation is
considered the best method for treating end-stage renal disease
(2,3). At present, the kidneys used in
transplantation originate from the donation of relatives or others.
However, organ donation by itself cannot meet clinical needs, and
cultivating functional and portable kidneys has substantial
research significance and clinical value for resolving the kidney
shortage problem (4).
A number of previous studies demonstrated that
pluripotent stem cells could differentiate into mature cells and
possess the potential to spontaneously assemble into tissues or
organs (5,6). In 2015, two research teams from
Australia and the USA used different methods to induce human
induced pluripotent stem cells (hiPSCs) into kidney organoids,
comprising collecting tubes, proximal tubule cells, distal tubule
cells, glomerular cells, endothelial cells and other cell types
(7,8). The kidney organoids did not possess
kidney class structures but did have certain renal tubular
absorption characteristics (7,8).
However, inducing 3D renal structuress or organoids
in vitro has two disadvantages; one is the nutrient supply
problem, which limits the number of cells and thus also limits the
number of nephrons that can be generated, and the other is the low
degree of cell differentiation (7,8).
Previous experiments demonstrated that the stem cell
differentiation microenvironment affects the induction of 3D renal
structures or organoids and thus, could promote vascularization of
the graft (9-11).
The renal capsule is one site that is often
transplanted (9). As early as 2004,
Hammerman (12) identified that
renal primordia (metanephroi) transplanted into an animal renal
capsule undergo organogenesis in situ, become vascularized
by blood vessels of host origin and exhibit excretory function.
Also, newly developed nephron/posterior renal primordia have been
suggested to integrate into the host assembly system under the
renal capsule thereby enhancing the renal function of the host
(12). Recently, van den Berg et
al (13) demonstrated that human
pluripotent stem cell-derived kidney organoids, under the kidney
capsule and in the absence of any exogenous vascular endothelial
growth factor, develop host-derived vascularization. These previous
studies demonstrated that the renal capsule is an ideal site for
transplantation.
Unilateral nephrectomy can lead to compensatory
hypertrophy of the other kidney, which is a common clinical
phenomenon (14,15). Taking this phenomenon into
consideration, Dilworth et al (16) demonstrated that a left nephrectomy,
which was performed on a host rat, improved the growth of
transplanted metanephroi. Matsumoto et al (17) observed that grafts were well
differentiated after the transplantation of fetal kidneys into the
omenta or abdominal aortas of rats via unilateral nephrectomy.
Therefore, a unilateral nephrectomy model may be beneficial for
differentiation and vascularization of kidney organoids.
Therefore, the present study aimed to determine
whether the microenvironment of the renal capsule in
immunodeficient mice undergoing unilateral nephrectomy could
promote vascularization and differentiation of kidney organoids.
The present results may be of significance for renal regeneration
research.
Materials and methods
hiPSCs
hiPSCs were gifted by Professor Zhiguo Chen (Cell
Therapy Center, Beijing Institute of Geriatrics, Xuanwu Hospital
Capital Medical University) (18)
and were maintained in mTeSR1 medium (Stemcell Technologies, Inc.;
cat. no. 85850) in 6-well cell culture plates (Costar; Corning,
Inc.; cat. no. 3516) coated with 1% vol/vol human embryonic stem
cell-qualified Matrigel (Corning, Inc.; cat. no. 354277) in a 37˚C
incubator at 5% CO2. hiPSCs were passaged in
Dissociation Solution for human embryonic stem cells/iPSCs
(ReproCELL, Inc.; cat. no. RCHETP002) at a 1:3 ratio every 7 days,
according to the manufacturer's protocol.
Optimizing the conditions of
intermediate mesoderm formation in hiPSCs
hiPSCs were plated on a Matrigel-coated 6-well cell
culture dish at 2,000 cells per cm2 in mTeSR1 medium.
The next day, the cells had reached 20-30% confluence and were
treated with gradient concentrations (0, 6, 8, 10, 12 and 16 µM) of
CHIR99021.e (Sigma-Aldrich; Merck KGaA; cat. no. SML1046) in APEL
basal medium (Stemcell Technologies, Inc.; cat. no. 05270)
supplemented with antibiotic-antimycotic (Thermo Fisher Scientific,
Inc.; cat. no. 15240062) for 4 days.. The cells were
then collected and the mRNA levels of T, TBX6 and LHX1 (molecular
markers of primitive streaking) were detected by RT-qPCR. In the
next step, the cells were treated with gradient concentrations (0,
100, 150, 200, 250 and 300 ng/ml) of FGF9 (R&D Systems, Inc.;
cat. no. 273-F9/CF) for 3 days and 1 µg/ml heparin (Sigma-Aldrich;
Merck KGaA; cat. no. 9041-08-1) was simultaneously added. On the
7th day, qPCR was used to detect the expression of the anterior
intermediate mesoderm marker GATA3, and the posterior intermediate
mesoderm markers HOXD11 and EYA1. The medium was changed every
other day. The above induction processes were all in 37˚C
incubator. The best concentration of inducer was selected by the
expression of molecular marker.
3D kidney organoid formation
On day 7, the cells were collected and dissociated
into single cells using trypsin or TrypLE select (Thermo Fisher
Scientific, Inc.; cat. no. 1905777). Cells (5x105) were
centrifuged at 400 x g for 2 min at room temperature to form a
pellet and then transferred onto a Transwell 0.4-mm-pore polyester
membrane (Corning, Inc.; cat. no. CLS3450). The pellets were
treated with 5 µM CHIR99021 in APEL for 1 h, cultured with FGF9
(200 ng/ml) and heparin (1 µg/ml) for 5 days, and cultured in APEL
basal medium for another 2 weeks; the medium was changed three
times a week. Culture medium was added to the lower chamber of the
Transwell only and was in contact with the membrane but did not
cross the membrane.
Immunofluorescence analysis of 3D
organoids
3D kidney organoids were fixed with 4%
paraformaldehyde in PBS for 20 min at room temperature (RT) in a
96-well plate and then washed three times with PBS. The organoids
were then incubated in blocking buffer [0.3% Triton X-100 and 5%
normal donkey serum (cat. no. S9100; Beijing Solarbio Science and
Technology Co., Ltd.) for 1 h at RT and washed three times with
PBS. The organoids were incubated with primary antibodies in
antibody dilution buffer (0.3% Triton X-100 and 1% BSA in PBS)
overnight at 4˚C. The following antibodies and dilutions were used:
Anti-Brachyury (1:500; cat. no. ab209665; Abcam), anti-LHX1 (1:500;
cat. no. sc-515631; Santa Cruz Biotechnology, Inc.), anti-GATA3
(1:1,000; cat. no. ab199428; Abcam), anti-WT1 (1:500; cat. no.
ab89901; Abcam), anti-ECAD (1:500; cat. no. ab1416; Abcam),
anti-PODXL (1:500; cat. no. ab150358; Abcam), anti-NPHS1 (1:300;
cat. no. sc-376522; Santa Cruz Biotechnology, Inc.), anti-CD31
(1:500; cat. no. ab24590; Abcam). The organoids were then washed
with PBS three times for 1 h each. The 4th wash was performed
overnight at 4˚C. For immunostaining with biotinylated Lotus
tetragonolobus lectin (LTL; 1:300; cat. no. B-1325; Vector
Laboratories, Inc.), a Streptavidin/Biotin Blocking kit (cat. no.
SP-2002; Vector Laboratories, Inc.;) was used, according to the
manufacturer's protocol. The organoids were incubated with
secondary antibodies (1:500; Alexa Flour 488 labeled
goat-anti-mouse IgG; cat. no. A0428; Beyotime Institute of
Biotechnology or 1:500; cy3-labeled goat-anti-rabbit IgG; cat. no.
A0516, Beyotime Institute of Biotechnology or 1:500; m-IgGk BP-PE;
cat. no. SC-516141; Santa Cruz Biotechnology, Inc.) in antibody
dilution buffer for 1 h at RT and then washed with PBS three times
for 30 min each. Nuclei were counterstained with DAPI for more than
10 min at room temperature. The organoids were then mounted with
Vectashield (Vector Laboratories, Inc.; cat. no. H-1200) and
examined by confocal microscopy at x200 and x400 magnification
(Zeiss AG; model no. LSM780).
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was purified from cells using the Total
RNA kit Ⅰ (Omega Bio-Tek, Inc.; cat. no. R6834-01). In total, 2 g
RNA was used for RT with the ReverTra Ace qPCR RT kit (Toyobo Life
Science; cat. no. 741200) according to the manufacturer's protocol.
RT-qPCRs were conducted in duplicate using cDNA (1:100), 500 nM
forward and reverse primers and SYBR-Green (Toyobo Life Science;
cat. no. 717000). RT-qPCR was performed using the Roche Real-Time
PCR Detection System (Roche Diagnostics; LightCycler 480II). All
samples were run in two technical replicates and GAPDH was used as
the housekeeping gene. Values were calculated using the
2-∆∆Cq method (19). The
following primers were used: GAPDH, forward,
5'-GGAGCGAGATCCCTCCAAAAT-3' and reverse,
5'-GGCTGTTGTCATACTTCTCATGG-3'; T, forward,
5'-CTGGGTACTCCCAATGGGG-3' and reverse,
5'-GGTTGGAGAATTGTTCCGATGA-3'; T-box transcription factor 6 (TBX6),
forward, 5'-CATCCACGAGAATTGTACCCG-3' and reverse,
5'-AGCAATCCAGTTTAGGGGTGT-3'; LIM homeobox 1 (LHX1), forward,
5'-CCTGGACCGCTTTCTCTTGAA-3' and reverse,
5'-ACCGAAACACCGGAAGAAGTC-3'; GATA binding protein 3 (GATA3),
forward, 5'-GCCCCTCATTAAGCCCAAG-3' and reverse,
5'-TTGTGGTGGTCTGACAGTTCG-3'; homeobox 11 (HOXD11), forward,
5'-TCGACCAGTTCTACGAGGCA-3' and reverse,
5'-AAAAACTCGCGTTCCAGTTCG-3'; EYA transcriptional coactivator and
phosphatase 1 (EYA1), forward, 5'-GTCACAGTCTCAGTCACCTGG-3' and
reverse, 5'-GGGATAAGACGGATAGTCCTGC-3'
Western blotting
Protein sample preparation and western blotting were
performed as previously described (20). Blots were incubated with primary
antibodies against PODXL (1:1,000; cat. no. ab150358; Abcam), RET
(1:500; ab134100; Abcam) and WT1 (1:1,000; cat. no. ab89901;
Abcam), and NPHS1 (1:500; cat. no. sc-376522; Santa Cruz
Biotechnology, Inc.), WNT11 (1:500; cat. no. sc-365033; Santa Cruz
Biotechnology, Inc.) and GAPDH (1:3,000; cat. no. sc-47724; Santa
Cruz Biotechnology, Inc.) overnight at 4˚C, followed by appropriate
peroxidase-conjugated secondary antibodies (1:3,000; anti-mouse
IgG, HRP-linked; cat. no. 7076; Cell Signaling Technology, Inc.;
anti-rabbit IgG, HRP-linked; cat. no. 7074; Cell Signaling
Technology, Inc.). GAPDH served as an internal control.
Visualization of the immunocomplexes was conducted using an
enhanced chemiluminescent HRP substrate (cat. no. 1829501; EMD
Millipore) and followed by exposure to X-ray films.
Animal surgery
Animal care and handling were performed in
accordance with The National Institutes of Health Guide for the
Care and Use of Laboratory Animals. All procedures were approved by
The Ethics Committee of The Second Hospital of Shandong University.
A total of 12 male immunodeficient mice (BAlB/c Nude; age, 5-6
weeks; weight, 22-25 g; Beijing Vital River Laboratory Animal
Technology Company) were initially anesthetized with 1%
pentobarbital sodium (50 mg/kg) via intraperitoneal injection. The
right kidneys of the mice were removed and ligated with a surgical
line. Mice were divided into a control group and an experimental
group, with 6 mice in each group. In the experimental group, kidney
organoids were transplanted into the kidney capsule, which were
then sutured. In the control group, kidney organoids were
transplanted into the subcutaneous armpit of mice. No mice died
during the experiment. A total of 2 weeks after transplantation,
all mice were euthanized with 1% pentobarbital sodium (150 mg/kg)
via intraperitoneal injection. Criteria for judging death in mice
included continuous no spontaneous breathing for 2-3 min and no
blinking reflex.
The mice were maintained at 22-25˚C, at a relative
humidity of 50-60% and on a 12 h light/dark cycle. The health of
the mice was observed every day and it was ensured that the mice
had adequate access to food and water. The cages and mattresses
were changed once per week and the mice were kept in a warm
environment after anesthesia.
Hematoxylin and eosin staining
The graft was fixed in 4% paraformaldehyde for 48 h
at room temperature and embedded in paraffin, which was then cut
into sections with a thickness of 4 µm. An H&E staining kit
(cat. no. G1120; Beijing Solarbio Science & Technology Co.,
Ltd.) was used according to the manufacturer's instructions. Images
were captured using a light microscope at x100 magnification.
Statistical analysis
Statistical analysis was performed using GraphPad
Prism software (version 5.0; GraphPad Software, Inc.). A paired
Student's t-test was used for analysis of statistical significance
between the control and treated groups. The comparative data are
presented as the mean ± SD of at least three independent
experiments. P<0.05 was considered to indicate a statistically
significant difference.
Results
Optimizing the conditions of
intermediate mesoderm formatoin in hiPSCs
The methods of inducing kidney organoid formation
from hiPSCs have been widely discussed (7,8).
However, the dosages of factors required for induction of kidney
organoid formation in different hiPSC strains may be different;
therefore, the induction procedure was optimized according to the
method by Takasato et al (7).
First, hiPSCs were treated with gradient concentrations (0, 6, 8,
10, 12 and 16 µM) of CHIR99021 for 4 days in a 37˚C incubator at 5%
CO2, before they were observed under a light microscope
at x100 magnification (Fig. 1A). The
cells were then collected and the mRNA levels of T, TBX6 and LHX1
(molecular markers of primitive streaking) were detected by RT-qPCR
(Fig. 1B).
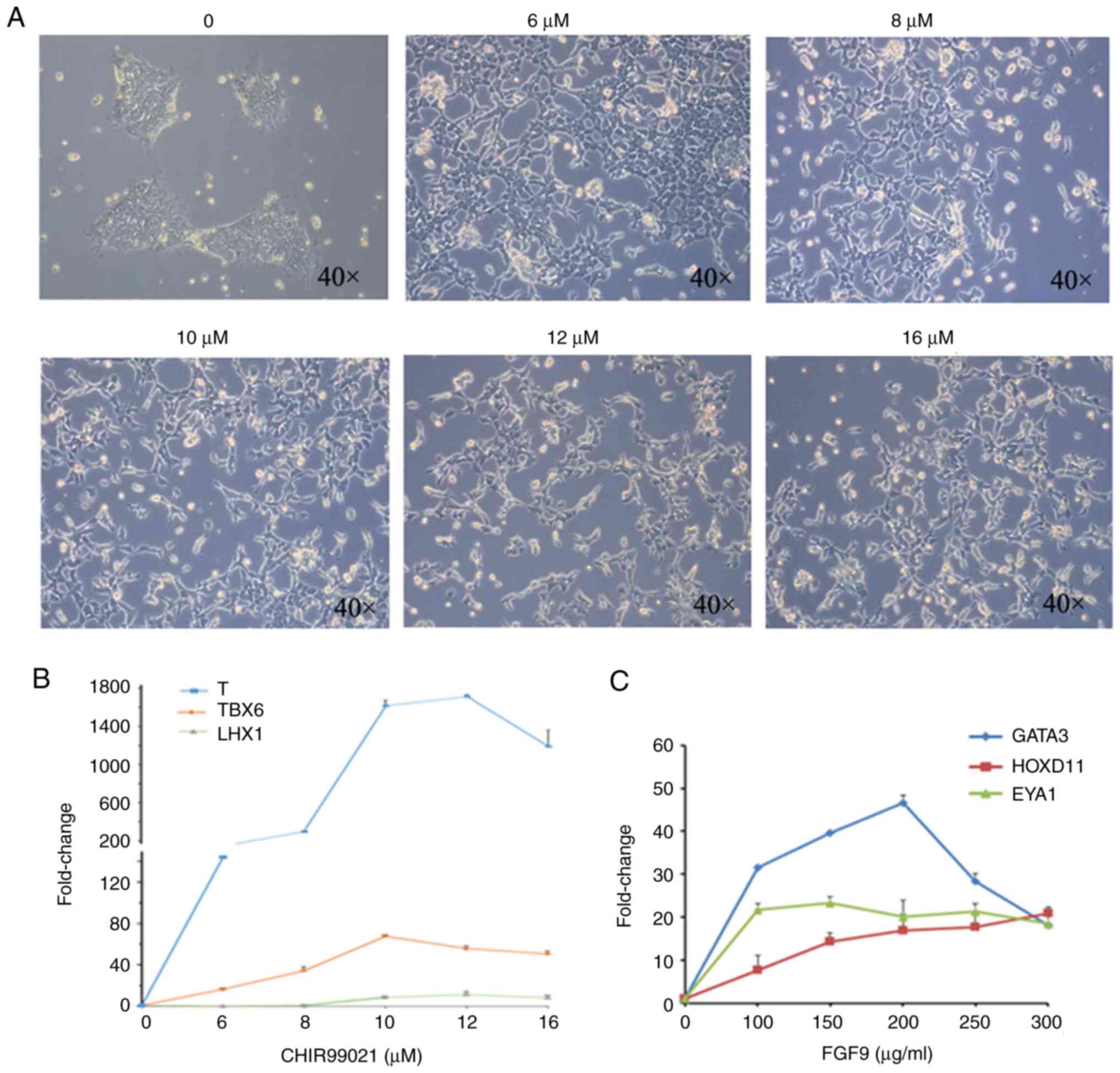 | Figure 1Condition optimization of intermediate
mesoderm induced by human induced pluripotent stem cells. (A)
Bright field observations at day 4 of induction by gradient
concentration of CHIR99021. (B) RT-qPCR detection of the primitive
streak markers T, TBX6 and LHX1 at day 4 of induction. (C) RT-qPCR
detection of anterior intermediate mesoderm marker GATA3, and
posterior intermediate mesoderm markers HOXD11 and EYA1, at day 4
of induction. The data are representative from a minimum of three
independent experiments. RT-qPCR, reverse
transcription-quantitative PCR; TBX6, T-box transcription factor 6;
LHX1, LIM homeobox 1; GATA3, GATA binding protein 3; HOXD11,
homeobox 11; EYA1, EYA transcriptional coactivator and phosphatase
1; FGF9, fibroblast growth factor 9. |
While 10 µM CHIR99021 was determined to be the best
concentration for induction based on the expression of TBX6, 12 µM
was deemed the best induction concentration based on the expression
of T and LHX1. The present study used 12 µM as the working
concentration of CHIR99021. In the second step, the cells were
treated with gradient concentrations of FGF9 (0, 100, 150, 200, 250
and 300 ng/ml) for 3 days (1 µg/ml heparin was simultaneously
added). On the 7th day, qPCR was used to detect the expression of
the anterior intermediate mesoderm marker GATA3, and the posterior
intermediate mesoderm markers HOXD11 and EYA1 (Fig. 1C). Among these markers, the
expression of HOXD11 increased with increasing FGF9 concentrations,
while the expression of EYA1 was not affected by changes in the
FGF9 concentration. Based on the expression of GATA3, 200 µg/ml
FGF9 was determined to be the best concentration for induction.
During the induction, the cellular morphology was
markedly altered. After the initial cell clones were treated with
CHIR99021, they were dispersed and exhibited a long shuttle shape,
but the cells gradually exhibited a fuller shape after CHIR99021
was replaced with FGF9 (Fig. 2A).
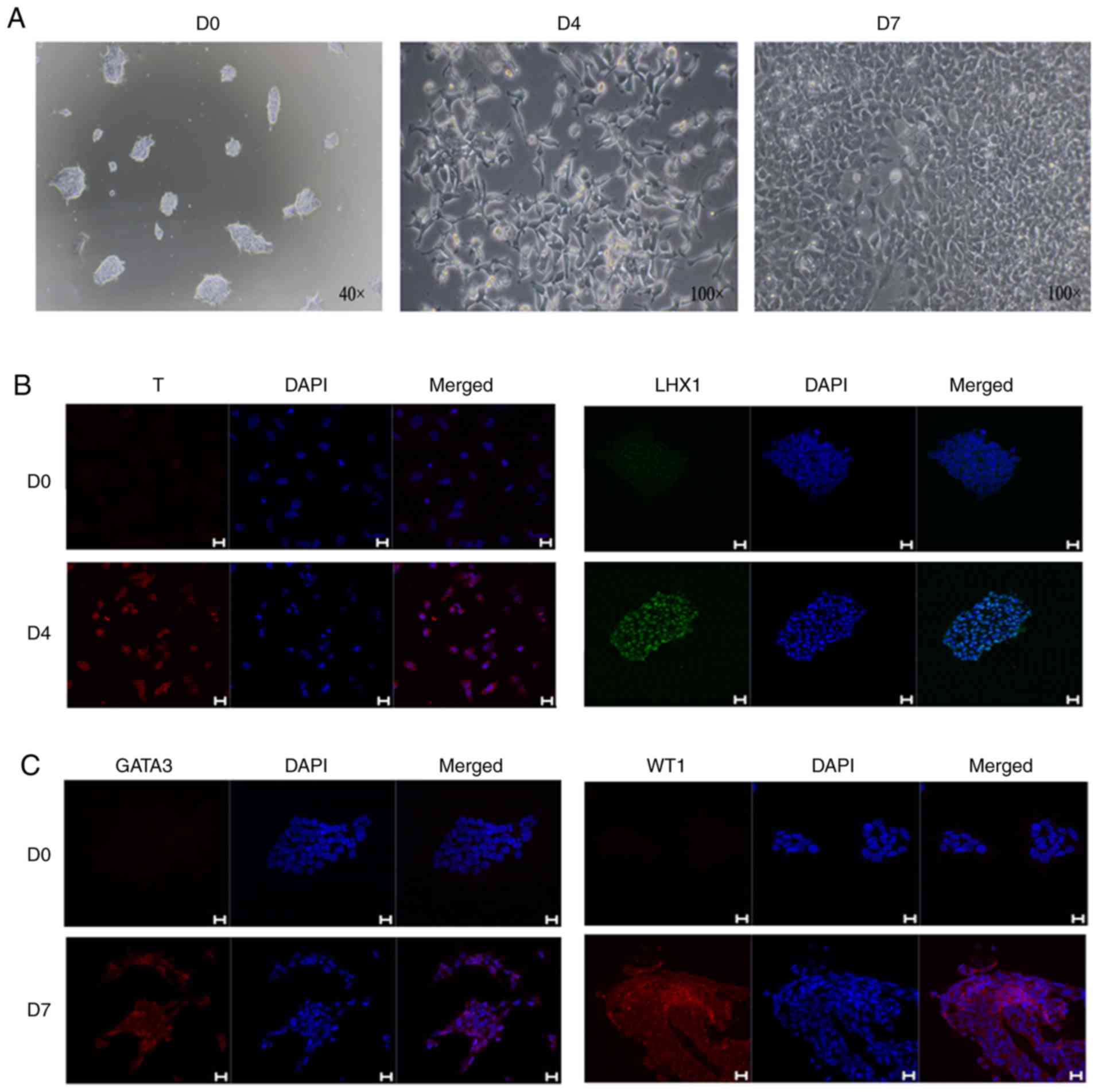
hiPSCs were analyzed and treated with CHIR9901 for 4 days, and
immunofluorescence analysis showed that the cells were positive for
both T and LHX1 (Fig. 2B). On the
7th day of induction, mesoderm markers were detected, and
immunofluorescence analysis demonstrated that the cells were
positive for the mesoderm markers GATA3 and WT1 (Fig. 2C).
3D culture of renal organoids in
vitro
The 3D culture of renal organoids in vitro
was examined (Fig. 3). The
morphology of kidney organoids was viewed under x40 light
microscope (Fig. 3A). H&E
staining analyses showed that hiPSCs were induced to form a tubular
structure in vitro (Fig. 3B).
The expression levels of renal-related molecular markers in the
cultured cells were detected by immunofluorescence. The tubule
marker LTL was identified to be strongly positive in the tubule
structure, while E-cadherin (ECAD) was weakly positive, suggesting
that the culture appeared to contain an immature renal tubule
structure (Fig. 3C). Some of the
tubules were also found to be GATA3+ ECAD+,
which indicated the existence of a collecting tube structure in the
culture (Fig. 3D). In addition, the
specific markers for ureteric buds, such as RET and WNT11, were
detected by western blotting. The expression of WNT11 was highest
on the 12th day of induction and then decreased. The expression
trend of RET was similar to that of WNT11 (Fig. 3G). These results suggested that the
production of ureteric progenitor cells during induction peaks on
the 12th day and then decreases. The culture was positive for
PODXL, a marker of the renal podocyte, an important cell in the
glomerular structure (5) (Fig. 3E). To confirm this result, the
molecular markers of podocytes (NPHS1, PODXL and WT1) were detected
by western blotting (Fig. 3F). WT1
began to be expressed on the 4th day of induction and was still
highly expressed on the D18 day of the kidney organoids formation
period The other two key podocyte markers were both highly
expressed on D18 day of the kidney organoid formation period. These
results suggested that renal podocyte-like cells may exist in the
kidney organoids.
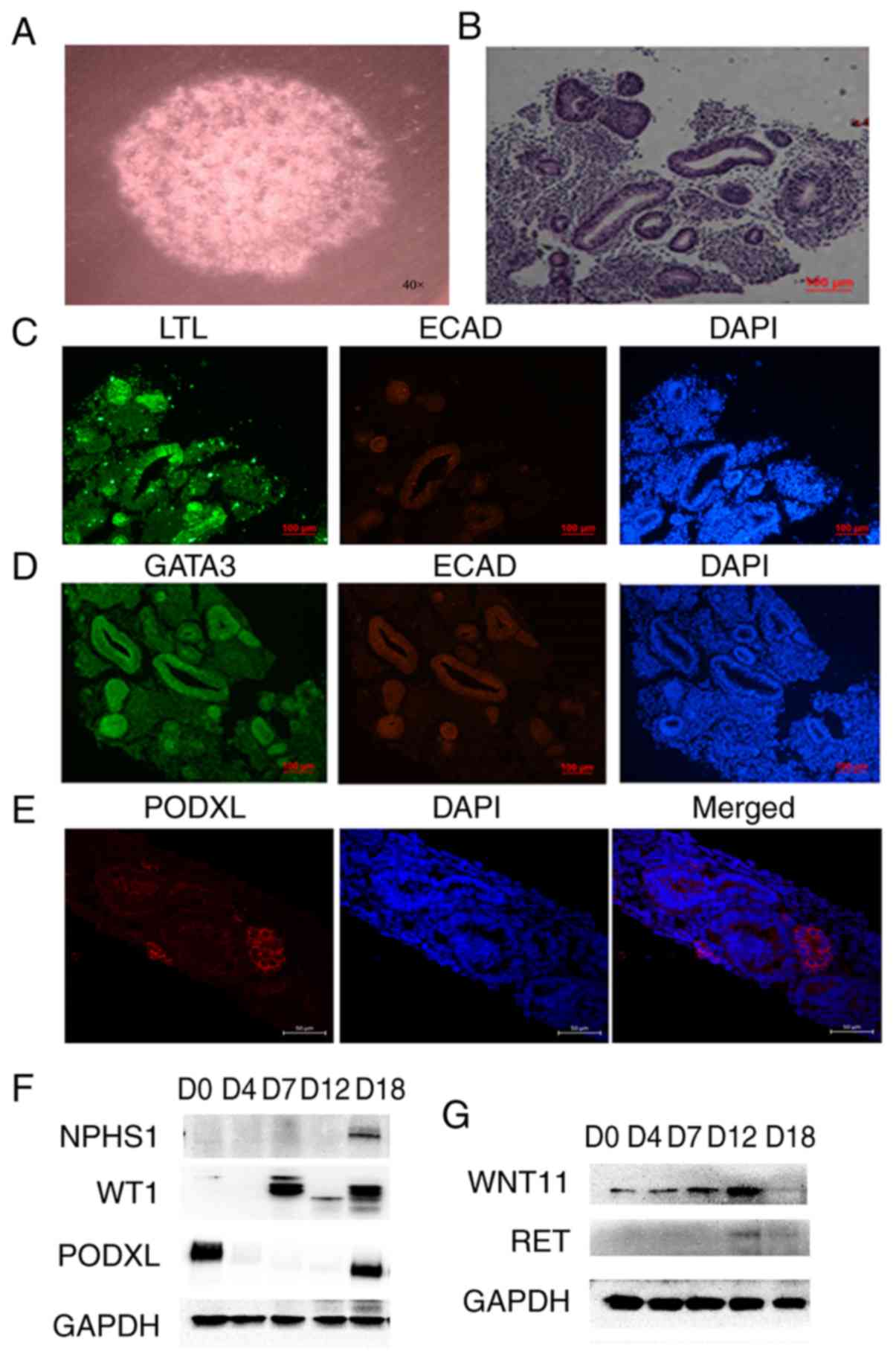 | Figure 33D culture of kidney organoids. (A)
Bright field observations of the kidney organoids. (B) Hematoxylin
and eosin staining results of the kidney organoids. (C) Expression
detection of renal tubular markers LTL and ECAD in kidney organoids
by immunofluorescence. (D) Expression detection of collection tube
markers GATA3 and ECAD in kidney organoids by immunofluorescence.
Scale bars, 100 µm. (E) Expression detection of kidney podocytes
marker PODXL in kidney organoids by immunofluorescence. Scale bars,
50 µm. (F) Expression detection of the molecular markers of
podocyte (NPHS1, PODXL and WT1) by western blotting. (G) Expression
detection of the specific markers for ureteric buds RET and WNT11
by western blotting. The data are representative from a minimum of
three independent experiments. LTL, Lotus tetragonolobus lectin;
ECAD, e-cadherin; GATA3, GATA binding protein 3; NPHS1, nephrin;
PODXL, podocalyxin; WT1, Wilms tumor protein; RET, proto-oncogene
tyrosine-protein kinase receptor Ret; WNT11, protein Wnt-11; D,
day. |
Renal organoids survive and grow under
the kidney capsule
As nutrient supply is a problem during in
vitro culture, organoid transplantation was carried out in the
present study. The present results demonstrated that the renal
capsule provided a differentiation microenvironment that was
sufficient for metanephros, while unilateral nephrectomy promoted
compensation of the other side of the kidney by increasing the
blood supply. On the 12th day of the experiment, the cultured
organoids were transplanted into the renal capsules of
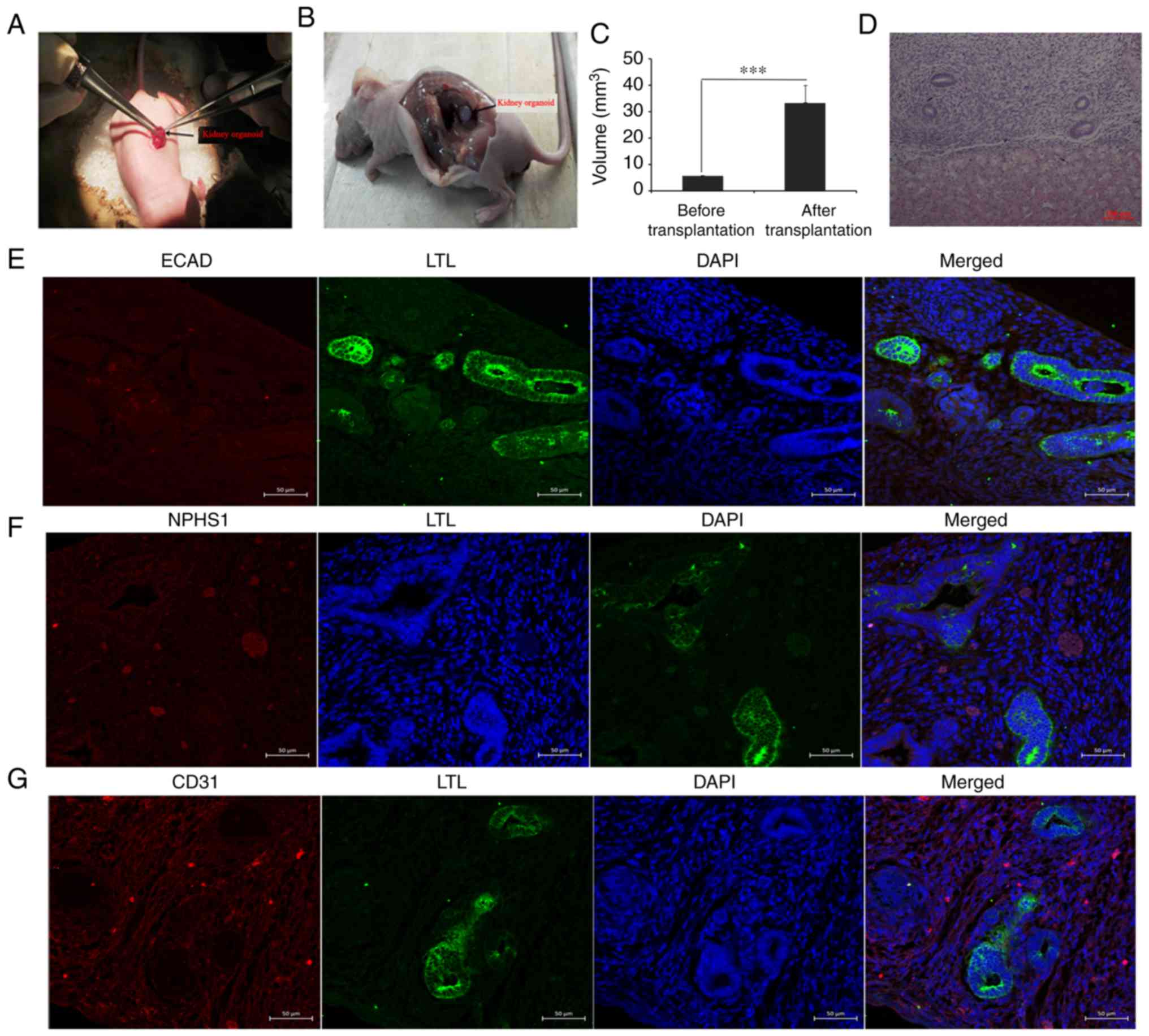
immunodeficient mice via unilateral nephrectomy (Fig. 4A). After 2 weeks of culture in
vivo, the grafts were removed and related molecular markers
were detected (Fig. 4B-G). Kidney
organoids transplanted in the subcutaneous armpit of mice could not
be detected in the control group (data not shown). Statistical
analysis identified that the graft volumes (kidney organoids)
increased significantly after transplantation, which suggested that
the host supplied nutrients for the graft (Fig. 4C). HE staining results showed tubular
structures similar to renal tubules (Fig. 4D). A renal tubular marker (LTL) and
glomerular marker (NPHS1) were detected using immunofluorescence.
The present results demonstrated that some of the grafts were
positive for LTL, while the expression of ECAD in LTL-positive
tubules was negative, indicating that the tubular structure was
immature (Fig. 4E). There were
sporadic NPHS1-positive grafts, suggesting the presence of kidney
podocytes (Fig. 4F). The molecular
marker CD31 was partially positive in vascular epithelial cells,
indicating the formation of blood cells (Fig 4G).
Discussion
Organoid-related research has attracted increasing
attention, and numerous methods for the in vitro induction
of kidney organoids have been reported. Takasato et al
(7) induced anterior and posterior
intermediate mesoderms by using a Wnt agonist and FGF9 as the main
inducers, and these components were further differentiated into
metanephric mesenchymal cells and ureteral progenitor cells,
eventually forming kidney organoids. Differentiation of iPSCs is a
complex and long-term process. Volpato et al (21) demonstrated that interlaboratory
reproducibility of iPSC differentiation is poor and cell type
heterogeneity is a major source of interlaboratory variation.
Phipson et al (22) analyzed
the sources of transcriptional variation in a specific kidney
organoid protocol, and demonstrated interexperimental and
inter-clonal variation in kidney organoid differentiation within a
same laboratory. The interexperimental and inter-clonal variation
in kidney organoid differentiation is unavoidable at present.
Therefore, at least three biological repeats were required to yield
similar results in the present study.
Based on the method established by Taksato et
al (7), the conditions for
induction at the 2D stage were optimized, demonstrating that 12 µM
CHIR99021 led to the most optimal induction when the hiPSCs used in
the present study changed to the primitive streak. On the 7th day,
RT-qPCR assays indicated that 200 ng/ml FGF9 led to the best
induction, which was consistent with the results of the previous
study (7).
The cells were placed on a Transwell membrane and 3D
culturing was performed. After 5 days, 200 ng/ml FGF9 was replaced
with a simple differentiation medium, APEL, for 2 weeks.
Immunofluorescence analysis confirmed that hiPSC-induced kidney
organoids in vitro comprised renal tubules, collecting ducts
and renal podocytes. However, numerous problems are still
associated with inducing kidney organoids in vitro, as renal
tubular differentiation is not mature, no mature glomerular
structure was observable, and the nutrient supply limitation had
not been solved. Another experiment (Fig. S1) and other previous studies
(16,17) demonstrated that unilateral nephrectomy promoted
the growth of another kidney graft. Kidney organoids induced in
vitro were transplanted into the renal capsules of
immunodeficient mice and then one of their kidneys was removed to
create an environment conducive to kidney organoid growth. At 2
weeks after transplantation, the kidneys were removed, stained with
HE and subjected to immunofluorescence analysis. The present
results demonstrated that the kidney organoid volumes were
increased significantly compared with those before transplantation,
suggesting that the kidney organoids obtained nutrients from the
host, and CD31 positivity indicated the presence of vascular
endothelial cells (23). However, no
positive ECAD expression was observed, which is related to the
differentiation and maturation of renal tubules (7). No marked difference in the expression
of molecular markers of podocytes was observed between the in
vivo and in vitro-induced kidney organoids.
van den Berg et al (13) demonstrated generation of vascularized
human kidney tissues derived from hiPSCs upon transplantation of
human kidney organoids to renal capsule in mice. In addition, their
research demonstrated that renal subcapsular transplantation of
PSC-derived kidney organoids induced significant glomerular and
tubular maturation in vivo (13). In comparison with the previous study
by van den Berg et al (13),
the present study has a number of differences, including the animal
surgery, in vitro induction time, in vivo culture
time and identification methods. The present study demonstrated
that the internal environment can provide nutrients to the graft
(kidney organoids), but short-term culture does not significantly
promote the differentiation or maturation of renal organ-related
cells. Additionally, it was observed that the tubules in the kidney
organoids induced or cultured in vitro were very sparse
compared with those in the normal kidneys, and a large area of low
or osteogenic differentiation was observed, suggesting that the
efficiency of kidney organoid induction was still substantially
improved. To improve the induction efficiency of kidney organoids,
future studies should further optimize the induction conditions,
and consider screening renal interstitial cells and ureteral
progenitor cells via flow separation, and then culturing these
cells. Considering the degree of differentiation observed, the time
of kidney organoid culture in vitro or in vivo should
be extended. After kidney organoid transplantation, VEGF and FGF9
may be added to promote the angiogenesis and differentiation of
renal-related cells.
In summary, the present study demonstrated that the
microenvironment of the kidney capsule after unilateral nephrectomy
is conducive to the proliferation of kidney organoids. However,
there are many limitations in the present study that should be
addressed.
Supplementary Material
Figure S1. Effect of unilateral
nephrectomy on engraftment of kidney organoids below the kidney
capsules. Kidney organoids (cultured in 96‑well, round bottom,
ultra‑low attachment plates in the 3‑D culture stage; the induction
method for the first 7 days was the same as the original one)
derived from human pluripotent stem cells were transplanted into
the kidney capsule of mice. Experimental group: Unilateral
nephrectomy performed on mice. Control group: Unilateral
nephrectomy was not performed on mice. There were six mice in each
group. (A) A total of 2 weeks later after transplantation, the
kidney organoids were removed and imaged. (B) Statistical results
showed that the volume of kidney organoids in the experimental
group was 23.89±6.39 mm3 and the volume of kidney
organoids in the control group was 8.94±2.25 mm3. There
was a significant difference between the two groups. Data are
presented as the mean ± SD. P‑values were calculated using an
independent t‑test. **P<0.01.
Acknowledgements
The authors would like to thank Dr Yong Guan and Dr
Yanxia Guo (Engineering Laboratory of Shandong Province for
Structure and Functional Reconstruction of Urinary Organs) for
providing helpful discussions and technical support concerning the
present study.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant nos. 81700592 and
81670625) and the Shandong Provincial Natural Science Foundation
(grant no. ZR2017PH058).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
SZ and FK conducted the study. SZ and GC designed
the experiments. DZ, XD, KL and XZ performed experiments, and
collected and analyzed the data. All authors read and approved the
final manuscript.
Ethics approval and consent to
participate
Animal care and handling were performed in
accordance with The National Institutes of Health Guide for the
Care and Use of Laboratory Animals. All procedures were approved by
The Ethics Committee of The Second Hospital of Shandong
University.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Ng JK and Li PK: Chronic kidney disease
epidemic: How do we deal with it? Nephrology (Carlton). 23:116–120.
2018.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Tsai HB, Chao CT, Chang RE and Hung KY:
COGENT Study Group: Conservative management and health-related
quality of life in end-stage renal disease: A systematic review.
Clin Invest Med. 40:E127–E134. 2017.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Thuret R, Timsit MO and Kleinclauss F:
Chronic kidney disease and kidney transplantation. Prog Urol
(French). 26:882–908. 2016.PubMed/NCBI View Article : Google Scholar
|
|
4
|
van Gelder MK, Mihaila SM, Jansen J,
Wester M, Verhaar MC, Joles JA, Stamatialis D, Masereeuw R and
Gerritsen KG: From portable dialysis to a bioengineered kidney.
Expert Rev Med Devices. 15:323–336. 2018.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Kitamura S, Sakurai H and Makino H: Single
adult kidney stem/progenitor cells reconstitute 3-dimensional
nephron structures in vitro. Stem Cells. 33:774–784.
2015.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Taguchi A, Kaku Y, Ohmori T, Sharmin S,
Ogawa M, Sasaki H and Nishinakamura R: Redefining the in vivo
origin of metanephric nephron progenitors enables generation of
complex kidney structures from pluripotent stem cells. Cell Stem
Cell. 14:53–67. 2014.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Takasato M, Er PX, Chiu HS, Maier B,
Baillie GJ, Ferguson C, Parton RG, Wolvetang EJ, Roost MS, Chuva de
Sousa Lopes SM and Little MH: Kidney organoids from human iPS Cells
contain multiple lineages and model human nephrogenesis. Nature.
526:564–568. 2015.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Morizane R, Lam AQ, Freedman BS, Kishi S,
Valerius MT and Bonventre JV: Nephron organoids derived from human
pluripotent stem cells model kidney development and injury. Nat
Biotechnol. 33:1193–1200. 2015.PubMed/NCBI View
Article : Google Scholar
|
|
9
|
Cunha GR and Baskin L: Use of sub-renal
capsule transplantation in developmental biology. Differentiation.
9:4–9. 2016.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Schmidt C: Pancreatic islets find a new
transplant home in the omentum. Nat Biotechnol.
35(8)2017.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Gould DJ, Mehrara BJ, Neligan P, Cheng MH
and Patel KM: Lymph node transplantation for the treatment of
lymphedema. J Surg Oncol. 118:736–742. 2018.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Hammerman MR: Renal organogenesis from
transplanted metanephric primordia. J Am Soc Nephrol. 15:1126–1132.
2004.PubMed/NCBI View Article : Google Scholar
|
|
13
|
van den Berg CW, Ritsma L, Avramut MC,
Wiersma LE, van den Berg BM, Leuning DG, Lievers E, Koning M,
Vanslambrouck JM, Koster AJ, et al: Renal subcapsular
transplantation of psc-derived kidney organoids induces
neo-vasculogenesis and significant glomerular and tubular
maturation in vivo. Stem Cell Rep. 10:751–765. 2018.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Urie BK, Tillson DM, Smith CM, Brawner WR,
Almond GT, Beard DM, Lenz SD and Lothrop CD Jr: Evaluation of
clinical status, renal function, and hematopoietic variables after
unilateral nephrectomy in canine kidney donors. J Am Vet Med Assoc.
230:1653–1656. 2007.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Kendi Celebi Z, Peker A, Kutlay S, Kocak
S, Tuzuner A, Erturk S, Keven K and Sengul S: Effect of unilateral
nephrectomy on urinary angiotensinogen levels in living kidney
donors: 1 year follow-up study. J Renin Angiotensin Aldosterone
Syst. 18(1470320317734082)2017.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Dilworth MR, Clancy MJ, Marshall D,
Bravery CA, Brenchley PE and Ashton N: Development and functional
capacity of transplanted rat metanephroi. Nephrol Dial Transplant.
23:871–879. 2008.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Matsumoto K, Yokoo T, Yokote S, Utsunomiya
Y, Ohashi T and Hosoya T: Functional development of a transplanted
embryonic kidney: Effect of transplantation site. J Nephrol.
25:50–55. 2012.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Wang S, Wang B, Pan N, Fu L, Wang C, Song
G, An J, Liu Z, Zhu W, Guan Y, et al: Differentiation of human
induced pluripotent stem cells to mature functional Purkinje
neurons. Sci Rep. 5(9232)2015.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2-Delta Delta C (T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Yuan H, Gong A and Young CY: Involvement
of transcription factor Sp1 in quercetin -mediated inhibitory
effect on the androgen receptor in human prostate cancer cells.
Carcinogenesis. 26:793–801. 2005.
|
|
21
|
Volpato V, Smith J, Sandor C, Ried JS,
Baud A, Handel A, Newey SE, Wessely F, Attar M, Whiteley E, et al:
Reproducibility of molecular phenotypes after long-term
differentiation to human iPSC-derived neurons: A multi-site omics
study. Stem Cell Reports. 11:897–911. 2018.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Phipson B, Er PX, Combes AN, Forbes TA,
Howden SE, Zappia L, Yen HJ, Lawlor KT, Hale LJ, Sun J, et al:
Evaluation of variability in human kidney organoids. Nat Methods.
16:79–87. 2019.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Lertkiatmongkol P, Liao D, Mei H, Hu Y and
Newman PJ: Endothelial functions of platelet/endothelial cell
adhesion molecule-1 (CD31). Curr Opin Hematol. 23:253–259.
2016.PubMed/NCBI View Article : Google Scholar
|