Over the years, there has been an important number
of attempts to classify allergic diseases, especially on
respiratory level. Respiratory allergies may begin at any level of
the respiratory tract (nose, sinuses, lungs) and, over time, they
can generalize throughout, process known as the United Airway
Disease (UAD) (1). One of them was
based on the evaluation of specific mechanisms that represent the
pathophysiological background of the disease: endotypes (2). A disease endotype includes the specific
biological pathway (describing an etiology and/or a firm
pathophysiologic mechanism) that explains the observable properties
of a phenotype (clinical description of a disease without a
connection with underlying pathology). Lötvall et al
(2) were the first to propose the
criteria that characterizes an endotype. In order to be accepted,
an endotype description should take into account at least 5 of
these 7 aspects: Clinical, biomarkers, lung function, genetics,
histopathology, epidemiology and treatment response (3). At the moment, there are two main
endotypes described: Th2 and non-Th2.
Over the last decade, one of the most studied
endotypes was the one based on type 2 immune response.
Historically, allergic asthma and rhinitis were hypothesized to be
produced by this type of mechanism (4,5). Type 2
immune response is based on the contribution of the following
cells: Th2 cells (6), type 2 B cells
(7), interleukin-4 (IL-4) secreting
NK cells (8), IL-4 secreting T-NK
cells (9), mast cells, eosinophils,
basophils, their cytokines (CK) (5):
IL-4, IL-5, IL-9, IL-13, along with those CKs secreted by tissue
cells (5): IL-25(10), IL-31(11), IL-33(12) and TSLP (13). Starting from the characterization of
Th2 endotype, experts identified two endotypes: Th2-high and
Th2-low (14), which were divided
further in several sub-endotypes: IL-5-high, IL-13-high, IgE-high
(5). This kind of endotype may also
be characterized by several other biomarkers: blood or sputum
eosinophilia (15), periostin
(16), considered to be
characteristic for it. Type 2 immune response underlines atopic
asthma and allergic rhinitis (AR) as fundamental for the united
airway concept (5). In addition,
type 2 immune response seems to be important in chronic
rhinosinusitis (CRS) with nasal polyposis (CRSwNP), tissue
eosinophilia and evidence of eosinophil activation, being closely
associated with remodeling features of CRS (5). The whole disease spectrum of atopic
dermatitis (AD) from background inflammation in asymptomatic
patients to chronic disease is also covered by this type of
mechanism (5).
Recent years brought to the allergist's attention a
new endotype in allergic respiratory diseases: non-type 2 immune
response driven endotype. This endotype is related to neutrophilic
inflammation, Th17 activation (17),
neurogenic inflammation and tissue remodeling (4). There are two major mechanisms that are
considered to contribute to definition of this endotype: the
activation of the IL-17-dependent pathway and neutrophil intrinsic
abnormalities (4). It was
demonstrated that IL-17 is linked to remodeling (18), airway hyper-reactivity (AHR)
(19), asthma severity (20) and inflammation (21). Lung airway neutrophilia seems to be
associated with lower lung function, thickening airway walls and
more air trapping (3). For decades,
Th1 immune response was considered the main mechanism responsible
for the pathophysiology of non-atopic asthma (5). This response is characterized by the
domination of Th1 cells and their mediators: interferon-γ (IFN-γ)
(22) and tumor necrosis factor-α
(TNF-α) (23). Initially described
in non-atopic asthma or in severe asthma, it has recently been
connected with allergic rhinitis and asthma as well (24,25).
TNF-α produces a nasal inflammatory response in patients with AR
characterized by plasma exudation and late phase neutrophil
activity 24 h post nasal challenge (24). An increase of IFN-γ levels was
observed due to increased exposure to polycyclic aromatic
hydrocarbons (PAHs), known for their predisposition to atopy
(25). In recent years, mixed
endotypes/sub-endotypes, such as Th1/Th17(4) or Th2/Th17 endotype were proposed
(26).
Besides the classic difference Th1 vs. Th2,
immunology has been described previously in many other types of
lymphocytes. The importance of Th17 in respiratory allergies has
been presented before. Th9 cells have an important role in the
immune responses regulation. They express predominantly IL-9. IL-9
causes the induction of lung eosinophilia, increased serum total
IgE levels, airway hyperreactivity (27), the generation of cytokines from
active mast cells; it also up-regulates high-affinity IgE receptors
on mast cells (28). Th22 cells are
positive for chemokine receptors CCR4, CCR6 and CCR10 and produce
mostly IL-22. IL-22 has been found to be increased in patients with
AR (29) and asthma (30). GM-CSF producing T cells were also
described. Increased levels of GM-CSF were found during the
birch-pollen season in the nasal lavage (31). T follicular helper cells
(TFH) represent a specialized CXCR5-expressing
CD4+ T cell population, regulated by Bcl-6. Peripheral
circulating TFH can be divided into three subsets:
cTFH cells
(BCL6-CXCR3+CCR6-),
cTFH2 cells
(BCL6-CXCR3-CCR6-), and
cTFH17
(BCL6-CXCR3-CCR6+) cells, based on
the differential expression of the chemokine receptors CXCR3 and
CCL6(32). Significant levels were
found in child and adult asthma patients (33). In addition, TFH were
positively correlated with total IgE levels in the blood (34). Unconventional T lymphocytes, such as
invariant natural killer T (iNKT) and mucosal-associated invariant
T cells (MAIT), are considered potential candidates for studying
the mechanisms underlying the pathophysiology of asthma (35). MAIT cells produce low-to-moderate
levels of IL-4 and IL-13(36). A
recent study suggests that MAIT-17 cells may be associated with
asthma symptoms (37).
Another important aspect is represented by the
plasticity of T cells. Previously it was shown that T lymphocytes
can display an important grade of plasticity when they are exposed
to re-polarizing signals (38).
Signaling via Toll-like receptors can drive Th2 cells to an
IFN-γ-secreting phenotype (39). Th9
cells may develop from Th2 cells under the action of TGF-β
(40). Th1/Th2 hybrid cells may
develop from Th2 precursor cells under the influence of interferons
(41). Th1 and Th17 cells may
produce IL-4 under some circumstances (42).
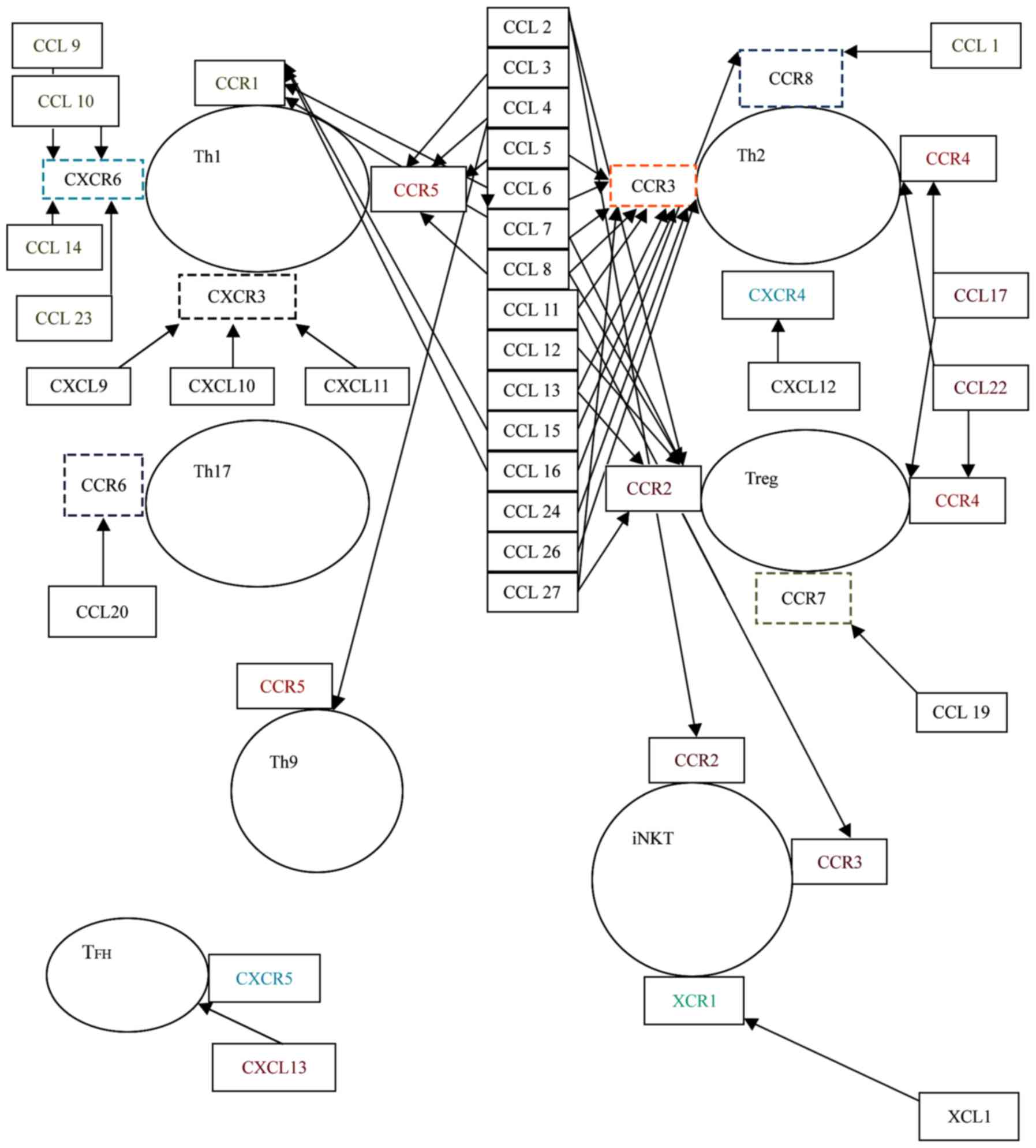
Chemokines (CC) are chemoattractant cytokines that
signal through seven-transmembrane-spanning domain, pertussis
toxin-sensitive, G-protein-coupled receptors (GPCRs). They are
classified into four families, based on the arrangement of the
first two N-terminal cysteine residues within their amino acid
sequence: CXC(α) family, CC(β) family, CX3C(δ) family and C(γ)
family. Chemokines can be divided functionally into inflammatory
and homeostatic. Inflammatory chemokines are produced during an
inflammatory response by activated leukocytes or tissue resident
cells. Examples include CXCL9 [monokine induced by γ-interferon
(MIG)], CXCL10 [interferon-γ-induced protein 10 (IP-10)] and CXCL11
(interferon-inducible T cell α-chemoattractant (ITAC)] that attract
Th1 cells and neutrophil-attracting chemokines: CXCL1
[growth-regulated oncogene (GROα)] and CXCL8 (interleukin-8).
Homeostatic chemokines are produced by healthy tissues and direct
leukocytes to fulfill their normal roles, which are immune
surveillance, hematopoiesis and embryogenesis: CCL19 [EBI1 ligand
chemokine (ELC)], CCL21 [secondary lymphoid-tissue chemokine
(SLC)], CCL25 [thymus-expressed chemokine (TECK)] and CCL27
[cutaneous T cell-attracting chemokine (CTAK)] (43).
Th2 cells are classically associated with the CC (β)
family (best studied until now are CCL1, CCL17, CCL18, CCL22)
(43). They possess three receptors
for CC: CCR3, CCR4, and CCR8. CCL1 (also known as I-309) is
a potent attractant for Th2 lymphocytes (44). CCL1 represents the predominant
CC secreted from IgE-activated mast cells and is found in high
concentrations in asthmatic airways (45). Mast cells release of CCL1 was
proposed to be the key step for early Th2 recruitment through the
CCR8 receptor (46). CCL1 was
found to be significantly elevated in the bronchoalveolar fluid
(BALF) from atopic asthmatic patients as compared with volunteers
(44), and asthmatic vs. controls
(47). The role of CCL1 in the
pathogenesis of asthma was indirectly demonstrated by the
suppression of its serum levels after treating human monocytic
leukemia cell line THP-1 and human monocytes from healthy donors
with a cysteinyl leukotriene receptor antagonist (montelukast)
(48). Murine studies reinforced the
role of CCL1 released by mast cells and basophils and its receptor
(CCR8) in recruitment of IL-4, IL-5 and IL-13-secreting T
lymphocytes into the airways (45,49).
CCL18 [pulmonary and activation-regulated chemokine
(PARC)] is another chemokine, production of which, is induced by
the inflammatory Th2 cytokines. CCL18 exhibits dual functions, with
pro- and anti-inflammatory properties, according to the environment
(baseline or inflammatory) and to the genetic background. CCL18
recruits basophils and Th2 cells activates basophils and induces
histamine release (58). CCL18
levels were found to be elevated in patients with asthma after
segmental allergen challenge (59)
and significantly correlated with sputum eosinophil percentages
(60) in patients with dust mites
allergy (61) and AR (62), results congruent with theoretical
data.
CCL22 [monocyte-derived chemokine (MDC)] induces the
selective migration of Th2 cells (roles in homing and recruitment
of CC chemokine receptor 4-bearing Th2 cells in allergen-induced
inflammation). High levels of CCL22 were found in the serum of
patients with allergic rhinitis with sensitization to birch pollen
(63) and ragweed pollen (56), which suggest a possible role in the
pathogenesis of AR.
The Eotaxin family, which include Eotaxin-1 (CCL11),
Eotaxin-2 (CCL24) and Eotaxin-3 (CCL26), recruits and activates
CCR3-bearing cells, such as Th2 lymphocytes, mast cells and
eosinophils that play an important role in allergic diseases
(64). Eotaxin-1 was also shown to
contribute in producing AHR. Eotaxin-1 presented elevated levels
and good correlations with sputum eosinophilia in children with
stable asthma compared with controls (65), and significant differences between
children with asthma vs. healthy children in BAL fluid (66), which suggests that eotaxin-1 may
regulate eosinophil trafficking into the airways of asthmatic
children in a coordinated manner. High levels of eotaxin-1 were
obtained after nasal allergen challenge in patients with AR
comparing with controls (67) and in
the material from nasal brushing in patients with asthma, allergic
rhinitis and COPD (68). Eotaxin-1
was involved in acute allergic airway response to Aspergillus
fumigatus (69), eosinophilic
inflammation in asthma (70) and
progress to AHR (71). Based on a
review that included 30 studies, Eotaxin-1 was considered as a
potentially useful biomarker for the diagnosis and assessment of
asthma severity and control (72).
Eotaxin-2, found in high levels in patients with nasal polyposis
and perennial allergic rhinitis when comparing to controls,
contribute to eosinophil attraction at the site of inflammation
(73). Eotaxin-3 was expressed by
nasal nerves of patients with AR after allergen challenge (74) and predisposed to AR in a Korean
population (75).
Besides this known relationship between Th2
lymphocytes and presented chemokines, a number of studies pointed
out the association of other CC and Th2 cells in the pathogenesis
of respiratory allergic diseases. For instance, in the last
decades, numerous studies focused on the investigation on CCL2
[monocyte chemoattractant protein-1 (MCP-1)], CCL5
(RANTES-regulated on activation, normal T cell expressed and
secreted), CCL7 [monocyte-chemotactic protein-3 (MCP-3)], CCL13
[monocyte-chemotactic protein-4 (MCP-4)] and CX3CL1 (Fraktaline)
regarding their association with inflammation in asthma and AR.
CX3CL1 (Fraktaline) has been found to be increased
in allergic diseases by promoting Th2 cell survival in the inflamed
airways (particularly in asthma). CX3CL1 levels were increased
after segmental allergen challenge in allergic asthmatic patients
(89,90). The CX3CL1/CX3CR1 axis was also
demonstrated to contribute to the development of allergic asthma in
murine studies (91).
Th1 lymphocytes are associated with CXC chemokines,
especially CXCL9, 10 and 11. Th1 lymphocytes possess the receptors
CCR1, CCR5, CXCR3 and CXCR6(43).
Three CC-CXCL9 (MIG), CXCL10 (IP-10) and CXCL11 (ITAC) are
described, which form a mini-cluster along the chromosome 4 at
q21.21. They are inducible by IFN-γ and act on immune cells
expressing CXCR3. Th1 cells express CXCR3 and thus will be
attracted by these chemokines (92).
CXCL9 and CXCL10 were released in large amounts by eosinophils
obtained from the peripheral blood of allergic volunteers when
these were stimulated concomitantly with TNF-α and IFN-γ (93). Also, it was observed that high
concentrations of CXCL9, 10 and 11 were significantly elevated
(94) in BALF after
broncho-provocation with antigen in patients with asthma. Allergen
activation in asthma patients provoked a diminished production of
CXCL9 and CXCL10, which contributed to a skewed Th2 profile
(95). Elevated levels of CXCL9 and
CXCL10 were observed in a pediatric study in patients with an acute
exacerbation in contrast to patients with stable asthma (96). CXCL10 proved to be a useful
inflammatory marker of occupational asthma (OA) exacerbation in
patients with wood dust OA in contrast with healthy controls
(97) and in the exacerbation of
childhood asthma (96). CXCL10
presented significant differences in patients with AR when it was
measured in nasal lavage before and after allergen exposure
(98). The development of
respiratory allergies was related to decreased levels of CXCL10,
particularly in patients with asthma, because this situation
indirectly favors Th2 cytokines production (99-101).
Th17 lymphocytes were found to be associated with
chemokines from the CXC family: CXCL1 (GROα), CXCL2 (GROβ), CXCL3
(GROγ), CXCL5 [epithelial-derived neutrophil-activating peptide 78
(ENA-78)], CXCL6 [granulocyte chemotactic protein 2 (GCP2)] and
CXCL8 (IL-8) (43). These
chemokines, along with CXCL7, belong to the family of ELR+CXC
chemokines (characterized by the highly conserved N-terminal ELR
(glutamic acid-leucine-arginine) triad and agonists for the CXCR2
receptor. Their primary role is to attract and activate neutrophils
(102). Besides these well-known
properties, a small number of studies associated these chemokines
with allergic inflammation. An important body of evidence related
to the contribution of these chemokines regarding allergic
inflammation in respiratory diseases was obtained through
experimental animal murine models. Thus, it was observed that
CXCL1, a chemokine expressed on macrophages, neutrophils and
epithelial cells known for its role in angiogenesis,
arteriogenesis, inflammation, wound healing, and tumorigenesis had
an increased expression in mouse lung epithelial cells in
sensitized animals with Anisakis, which suggest that allergens can
induce airway inflammation by elevating Th2 and Th17 responses
(103). In addition, it was found
that its concentration was increased in the BAL of
cockroach-sensitized mice where the allergen was administered
intranasally during a period of 5 days, which shows that CXCL1
might have roles in the remodeling during asthma (71). The expression of CXCL1 was decreased
in a murine model of AR when the experimental animals were treated
with flagellin-ovalbumin mixture (used as an adjuvant for
immunomodulation) (104) and in a
murine model of allergic severe asthma sensitization with house
dust mites (HDM) when the researchers blocked the activity of
IL17-A and IL17-F, which suggest that CXCL1 might be an important
player in neutrophilic allergic lung inflammation (105).
The contribution of CXCL2 and CXCL3 to mediating
normal and asthmatic airway smooth muscle cell (ASMC) migration
(through the ERK1/2 MAPK pathway) was demonstrated in a study that
used human ASMCs isolated from lung transplant donors, which
suggest a possible role in the pathogenesis of airway remodeling in
asthma (106).
Moreover, it was shown that not only is allergic
sensitization related to elevated levels of IL-8, but it is also
related to pollution. For instance, it was found that diesel
exhaust particles (DEP) induce expression of IL-8 in nasal
fibroblasts (117) and in primary
nasal epithelial cells (NECs) (118), suggesting that air pollution might
induce or aggravate allergic rhinitis through this chemokine
(117).
CCL4 was significantly associated with a mix of
lymphocytes (Th1, Th2, Th9, Th17) in subjects with severe asthma
(119).
Interleukins were used in the last two decades in
order to define and characterize the classical Th2/non-Th2
endotype, as well as the newest Th2/Th17. As shown, CCs appear to
be good candidates for a comprehensive characterization of
endotypes.
Professional editing, linguistic and technical
assistance was provided by Irina Radu, Individual Service Provider,
certified translator in Medicine and Pharmacy (certificate
credentials: Series E no. 0048).
No funding was received.
All data generated or analyzed during this study are
included in this published article.
NOB contributed substantially to the conception and
design of the study, the acquisition, analysis and interpretation
of the data, and was involved in the drafting of the manuscript. MD
and RSC contributed substantially to the acquisition, analysis and
interpretation of the data and were involved in the drafting of the
manuscript. DV, RCC, ASP, RC, CT and CG contributed substantially
to the acquisition of the data and were involved in the critical
revisions of the manuscript for important intellectual content. All
authors agreed to be accountable for all aspects of the work in
ensuring that questions related to the accuracy or integrity of any
part of the work are appropriately investigated and resolved. All
the authors read and approved the final version of the
manuscript.
No applicable.
Not applicable.
The authors declare that they have no competing
interests.
|
1
|
Yii AC, Tay TR, Choo XN, Koh MS, Tee AK
and Wang DY: Precision medicine in united airways disease: A
‘treatable traits’ approach. Allergy. 73:1964–1978. 2018.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Lötvall J, Akdis CA, Bacharier LB, Bjermer
L, Casale TB, Custovic A, Lemanske RF Jr, Wardlaw AJ, Wenzel SE and
Greenberger PA: Asthma endotypes: A new approach to classification
of disease entities within the asthma syndrome. J Allergy Clin
Immunol. 127:355–360. 2011.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Desai M and Oppenheimer J: Elucidating
asthma phenotypes and endotypes: Progress towards personalized
medicine. Ann Allergy Asthma Immunol. 116:394–401. 2016.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Muraro A, Lemanske RF Jr, Hellings PW,
Akdis CA, Bieber T, Casale TB, Jutel M, Ong PY, Poulsen LK,
Schmid-Grendelmeier P, et al: Precision medicine in patients with
allergic diseases: Airway diseases and atopic dermatitis-PRACTALL
document of the European Academy of Allergy and Clinical Immunology
and the American Academy of Allergy, Asthma & Immunology. J
Allergy Clin Immunol. 137:1347–1358. 2016.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Agache I and Akdis CA: Precision medicine
and phenotypes, endotypes, genotypes, regiotypes, and theratypes of
allergic diseases. J Clin Invest. 129:1493–1503. 2019.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Wambre E, Bajzik V, DeLong JH, O'Brien K,
Nguyen QA, Speake C, Gersuk VHH, DeBerg HA, Whalen E, Ni C, et al:
A phenotypically and functionally distinct human TH2 cell
subpopulation is associated with allergic disorders. Sci Transl
Med. 9(eaam9171)2017.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Velez TE, Swartzendruber JA, Hulse KE and
Bryce PJ: Regulation of B cell responses in allergy by histamine
receptors. J Immunol. 196(123.13)2016.
|
|
8
|
Kiniwa T, Enomoto Y, Terazawa N, Omi A,
Miyata N, Ishiwata K and Miyajima A: NK cells activated by
interleukin-4 in cooperation with interleukin-15 exhibit
distinctive characteristics. Proc Natl Acad Sci USA.
113:10139–10144. 2016.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Singh AK, Rhost S, Löfbom L and Cardell
SL: Defining a novel subset of CD1d-dependent type II natural
killer T cells using natural killer cell-associated markers. Scand
J Immunol. 90(e12794)2019.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Chen F, Hong H, Sun Y, Hu X, Zhang J, Xu
G, Zhao W, Li H and Shi J: Nasal interleukin 25 as a novel
biomarker for patients with chronic rhinosinusitis with nasal
polyps and airway hypersensitiveness: A pilot study. Ann Allergy
Asthma Immunol. 119:310–316.e2. 2017.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Baumann R, Rabaszowski M, Stenin I,
Gaertner-Akerboom M, Scheckenbach K, Wiltfang J, Schipper J and
Wagenmann M: The release of IL-31 and IL-13 after nasal allergen
challenge and their relation to nasal symptoms. Clin Transl
Allergy. 2(13)2012.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Nechama M, Kwon J, Wei S, Kyi AT, Welner
RS, Ben-Dov IZ, Arredouani MS, Asara JM, Chen CH, Tsai CY, et al:
The IL-33-PIN1-IRAK-M axis is critical for type 2 immunity in
IL-33-induced allergic airway inflammation. Nat Commun.
9(1603)2018.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Meng Q, Liu X, Li P, He L, Xie J, Gao X,
Wu X, Su F and Liang Y: The influence of house dust mite sublingual
immunotherapy on the TSLP-OX40L signaling pathway in patients with
allergic rhinitis. Int Forum Allergy Rhinol. 6:862–870.
2016.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Fahy JV: Type 2 inflammation in asthma -
present in most, absent in many. Nat Rev Immunol. 15:57–65.
2015.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Ciebiada M, Barylski M and Gorska Ciebiada
M: Nasal eosinophilia and serum soluble intercellular adhesion
molecule 1 in patients with allergic rhinitis treated with
montelukast alone or in combination with desloratadine or
levocetirizine. Am J Rhinol Allergy. 27:e58–e62. 2013.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Kralimarkova TZ, Popov TA, Staevska M,
Mincheva R, Lazarova C, Racheva R, Mustakov TB, Filipova V, Koleva
M, Bacheva K, et al: Objective approach for fending off the
sublingual immunotherapy placebo effect in subjects with
pollenosis: Double-blinded, placebo-controlled trial. Ann Allergy
Asthma Immunol. 113:108–113. 2014.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Diamant Z and Hanania NA: Editorial: The
new look on asthma: linking phenotypes, endotypes and biomarkers to
asthma management. Curr Opin Pulm Med. 22:1–2. 2016.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Chakir J, Shannon J, Molet S, Fukakusa M,
Elias J, Laviolette M, Boulet LP and Hamid Q: Airway
remodeling-associated mediators in moderate to severe asthma:
Effect of steroids on TGF-beta, IL-11, IL-17, and type I and type
III collagen expression. J Allergy Clin Immunol. 111:1293–1298.
2003.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Barczyk A, Pierzchala W and Sozañska E:
Interleukin-17 in sputum correlates with airway hyperresponsiveness
to methacholine. Respir Med. 97:726–733. 2003.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Staton TL, Peng K, Owen R, Choy DF,
Cabanski CR, Fong A, Brunstein F, Alatsis KR and Chen H: A phase I,
randomized, observer-blinded, single and multiple ascending-dose
study to investigate the safety, pharmacokinetics, and
immunogenicity of BITS7201A, a bispecific antibody targeting IL-13
and IL-17, in healthy volunteers. BMC Pulm Med.
19(5)2019.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Al-Kufaidy R, Vazquez-Tello A, BaHammam
AS, Al-Muhsen S, Hamid Q and Halwani R: IL-17 enhances the
migration of B cells during asthma by inducing CXCL13 chemokine
production in structural lung cells. J Allergy Clin Immunol.
139:696–699.e5. 2017.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Chambers ES, Nanzer AM, Pfeffer PE,
Richards DF, Timms PM, Martineau AR, Griffiths CJ, Corrigan CJ and
Hawrylowicz CM: Distinct endotypes of steroid-resistant asthma
characterized by IL-17A(high) and IFN-γ(high) immunophenotypes:
Potential benefits of calcitriol. J Allergy Clin Immunol.
136:628–637.e4. 2015.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Berry MA, Hargadon B, Shelley M, Parker D,
Shaw DE, Green RH, Bradding P, Brightling CE, Wardlaw AJ and Pavord
ID: Evidence of a role of tumor necrosis factor alpha in refractory
asthma. N Engl J Med. 354:697–708. 2006.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Widegren H, Erjefält J, Korsgren M,
Andersson M and Greiff L: Effects of intranasal TNFalpha on
granulocyte recruitment and activity in healthy subjects and
patients with allergic rhinitis. Respir Res. 9(15)2008.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Hew KM, Walker AI, Kohli A, Garcia M, Syed
A, McDonald-Hyman C, Noth EM, Mann JK, Pratt B, Balmes J, et al:
Childhood exposure to ambient polycyclic aromatic hydrocarbons is
linked to epigenetic modifications and impaired systemic immunity
in T cells. Clin Exp Allergy. 45:238–248. 2015.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Irvin C, Zafar I, Good J, Rollins D,
Christianson C, Gorska MM, Martin RJ and Alam R: Increased
frequency of dual-positive TH2/TH17 cells in bronchoalveolar lavage
fluid characterizes a population of patients with severe asthma. J
Allergy Clin Immunol. 134:1175–1186.e7. 2014.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Farahani R, Sherkat R, Hakemi MG,
Eskandari N and Yazdani R: Cytokines (interleukin-9, IL-17, IL-22,
IL-25 and IL-33) and asthma. Adv Biomed Res. 3(127)2014.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Kearley J, Erjefalt JS, Andersson C,
Benjamin E, Jones CP, Robichaud A, Pegorier S, Brewah Y, Burwell
TJ, Bjermer L, et al: IL-9 governs allergen-induced mast cell
numbers in the lung and chronic remodeling of the airways. Am J
Respir Crit Care Med. 183:865–875. 2011.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Tang J, Xiao P, Luo X, Bai J, Xia W, Chen
W, Li J, Yu Q, Shi S, Xu Y, et al: Increased IL-22 level in
allergic rhinitis significantly correlates with clinical severity.
Am J Rhinol Allergy. 28:197–201. 2014.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Besnard AG, Sabat R, Dumoutier L, Renauld
JC, Willart M, Lambrecht B, Teixeira MM, Charron S, Fick L, Erard
F, et al: Dual Role of IL-22 in allergic airway inflammation and
its cross-talk with IL-17A. Am J Respir Crit Care Med.
183:1153–1163. 2011.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Linden M, Svensson C, Andersson M, Greiff
L, Andersson E, Denburg JA and Persson CG: Circulating
eosinophil/basophil progenitors and nasal mucosal cytokines in
seasonal allergic rhinitis. Allergy. 54:212–219. 1999.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Gong F, Zheng T and Zhou P: T follicular
helper cell subsets and the associated cytokine IL-21 in the
pathogenesis and therapy of asthma. Front Immunol.
10(2918)2019.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Gong F, Qian C, Zhu H, Zhu J, Pan Y, Dong
Q and Jiang D: Circulating follicular T-helper cell subset
distribution in patients with asthma. Allergy Asthma Proc.
37:154–161. 2016.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Gong F, Zhu HY, Zhu J, Dong QJ, Huang X
and Jiang DJ: Circulating CXCR5+CD4+ T cells
participate in the IgE accumulation in allergic asthma. Immunol
Lett. 197:9–14. 2018.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Lezmi G and Leite-de-Moraes M: Invariant
natural killer T and mucosal-associated invariant T cells in
asthmatic patients. Front Immunol. 9(1766)2018.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Kurioka A, Jahun AS, Hannaway RF, Walker
LJ, Fergusson JR, Sverremark-Ekström E, Corbett AJ, Ussher JE,
Willberg CB and Klenerman P: Shared and distinct phenotypes and
functions of human CD161++ v alpha 7.2+ T
cell subsets. Front Immunol. 8(1031)2017.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Lezmi G, Abou Taam R, Dietrich C,
Chatenoud L, de Blic J and Leite-de-Moraes M: Circulating
IL-17-producing mucosal-associated invariant T cells (MAIT) are
associated with symptoms in children with asthma. Clin Immunol.
188:7–11. 2018.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Krawczyk CM, Shen H and Pearce EJ:
Functional plasticity in memory T helper cell responses. J Immunol.
178:4080–4088. 2007.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Filì L, Ferri S, Guarna F, Sampognaro S,
Manuelli C, Liotta F, Cosmi L, Matucci A, Vultaggio A and
Annunziato F: Redirection of allergen-specific TH2 responses by a
modified adenine through Toll-like receptor 7 interaction and
IL-12/IFN release. J Allergy Clin Immunol. 118:511–517.
2006.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Veldhoen M, Uyttenhove C, van Snick J,
Helmby H, Westendorf A, Buer J, Martin B, Wilhelm C and Stockinger
B: Transforming growth factor-beta ‘reprograms’ the differentiation
of T helper 2 cells and promotes an interleukin 9-producing subset.
Nat Immunol. 9:1341–1346. 2008.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Hegazy AN, Peine M, Helmstetter C, Panse
I, Fröhlich A, Bergthaler A, Flatz L, Pinschewer DD, Radbruch A and
Löhning M: Interferons direct Th2 cell reprogramming to generate a
stable GATA-3(+)T-bet(+) cell subset with combined Th2 and Th1 cell
functions. Immunity. 32:116–128. 2010.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Panzer M, Sitte S, Wirth S, Drexler I,
Sparwasser T and Voehringer D: Rapid in vivo conversion of effector
T cells into Th2 cells during helminth infection. J Immunol.
188:615–623. 2012.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Adkinson NF, Bochner BS, Burks AW, Busse
WW, Holgate ST, Lemanske RF and O'Hehir RE: Chapter 7. Chemokines
in Middleton's Allergy. In: Principles and Practice. Vol. 1. 8th
edition. Elsevier Saunders, Philadelphia. pp98–112. 2014.
|
|
44
|
Montes-Vizuet R, Vega-Miranda A,
Valencia-Maqueda E, Negrete-García MC, Velásquez JR and Teran LM:
CC chemokine ligand 1 is released into the airways of atopic
asthmatics. Eur Respir J. 28:59–67. 2006.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Gonzalo JA, Qiu Y, Lora JM, Al-Garawi A,
Villeval JL, Boyce JA, Martinez-A C, Marquez G, Goya I, Hamid Q, et
al: Coordinated involvement of mast cells and T cells in allergic
mucosal inflammation: Critical role of the CC chemokine ligand
1:CCR8 axis. J Immunol. 179:1740–1750. 2007.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Islam SA and Luster AD: T cell homing to
epithelial barriers in allergic disease. Nat Med. 18:705–715.
2012.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Mutalithas K, Guillen C, Raport C, Kolbeck
R, Soler D, Brightling CE, Pavord ID and Wardlaw AJ: Expression of
CCR8 is increased in asthma. Clin Exp Allergy. 40:1175–1185.
2010.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Lin YC, Huang MY, Lee MS, Hsieh CC, Kuo
HF, Kuo CH and Hung CH: Effects of montelukast on M2-related
cytokine and chemokine in M2 macrophages. J Microbiol Immunol
Infect. 51:18–26. 2018.PubMed/NCBI View Article : Google Scholar
|
|
49
|
Chhiba KD, Hsu CL, Berdnikovs S and Bryce
PJ: Transcriptional heterogeneity of mast cells and basophils upon
activation. J Immunol. 198:4868–4878. 2017.PubMed/NCBI View Article : Google Scholar
|
|
50
|
Takeuchi H, Yamamoto Y, Kitano H and
Enomoto T: Changes in thymus- and activation-regulated chemokine
(TARC) associated with allergen immunotherapy in patients with
perennial allergic rhinitis. J Investig Allergol Clin Immunol.
15:172–176. 2005.PubMed/NCBI
|
|
51
|
Teplyakov A, Obmolova G and Gilliland GL:
Structural insights into chemokine CCL17 recognition by antibody
M116. Biochem Biophys Rep. 13:27–31. 2017.PubMed/NCBI View Article : Google Scholar
|
|
52
|
Staples KJ, Hinks TS, Ward JA, Gunn V,
Smith C and Djukanović R: Phenotypic characterization of lung
macrophages in asthmatic patients: Overexpression of CCL17. J
Allergy Clin Immunol. 130:1404–12.e7. 2012.PubMed/NCBI View Article : Google Scholar
|
|
53
|
Ying S, O'Connor B, Ratoff J, Meng Q,
Mallett K, Cousins D, Robinson D, Zhang G, Zhao J, Lee TH, et al:
Thymic stromal lymphopoietin expression is increased in asthmatic
airways and correlates with expression of Th2-attracting chemokines
and disease severity. J Immunol. 174:8183–8190. 2005.PubMed/NCBI View Article : Google Scholar
|
|
54
|
Chen YL and Chiang BL: Targeting TSLP with
shRNA alleviates airway inflammation and decreases epithelial CCL17
in a murine model of asthma. Mol Ther Nucleic Acids.
5(e316)2016.PubMed/NCBI View Article : Google Scholar
|
|
55
|
Xiao JZ, Kondo S, Takahashi N, Odamaki T,
Iwabuchi N, Miyaji K, Iwatsuki K and Enomoto T: Changes in plasma
TARC levels during Japanese cedar pollen season and relationships
with symptom development. Int Arch Allergy Immunol. 144:123–127.
2007.PubMed/NCBI View Article : Google Scholar
|
|
56
|
Sun J, Wong B, Cundall M, Goncharova S,
Conway M, Dalrymple A, Coyle AJ, Waserman S and Jordana M:
Immunoreactivity profile of peripheral blood mononuclear cells from
patients with ragweed-induced allergic rhinitis. Clin Exp Allergy.
37:901–908. 2007.PubMed/NCBI View Article : Google Scholar
|
|
57
|
Tsybikov NN, Egorova EV, Kuznik BI,
Fefelova EV and Magen E: Biomarker assessment in chronic rhinitis
and chronic rhinosinusitis: Endothelin-1, TARC/CCL17, neopterin,
and α-defensins. Allergy Asthma Proc. 37:35–42. 2016.PubMed/NCBI View Article : Google Scholar
|
|
58
|
Chenivesse C and Tsicopoulos A: CCL18 -
Beyond chemotaxis. Cytokine. 109:52–56. 2018.PubMed/NCBI View Article : Google Scholar
|
|
59
|
Gavala ML, Kelly EA, Esnault S, Kukreja S,
Evans MD, Bertics PJ, Chupp GL and Jarjour NN: Segmental allergen
challenge enhances chitinase activity and levels of CCL18 in mild
atopic asthma. Clin Exp Allergy. 43:187–197. 2013.PubMed/NCBI View Article : Google Scholar
|
|
60
|
Kim HB, Kim CK, Iijima K, Kobayashi T and
Kita H: Protein microarray analysis in patients with asthma:
Elevation of the chemokine PARC/CCL18 in sputum. Chest.
135:295–302. 2009.PubMed/NCBI View Article : Google Scholar
|
|
61
|
de Nadaï P, Charbonnier AS, Chenivesse C,
Sénéchal S, Fournier C, Gilet J, Vorng H, Chang Y, Gosset P,
Wallaert B, et al: Involvement of CCL18 in allergic asthma. J
Immunol. 176:6286–6293. 2006.PubMed/NCBI View Article : Google Scholar
|
|
62
|
Peterson S, Poposki JA, Nagarkar DR,
Chustz RT, Peters AT, Suh LA, Carter R, Norton J, Harris KE,
Grammer LC, et al: Increased expression of CC chemokine ligand 18
in patients with chronic rhinosinusitis with nasal polyps. J
Allergy Clin Immunol. 129:119–27.e1, 9. 2012.PubMed/NCBI View Article : Google Scholar
|
|
63
|
Yanai M, Sato K, Aoki N, Takiyama Y,
Oikawa K, Kobayashi H, Kimura S, Harabuchi Y and Tateno M: The role
of CCL22/macrophage-derived chemokine in allergic rhinitis. Clin
Immunol. 125:291–298. 2007.PubMed/NCBI View Article : Google Scholar
|
|
64
|
Holgate ST and Davies DE: Rethinking the
pathogenesis of asthma. Immunity. 31:362–367. 2009.PubMed/NCBI View Article : Google Scholar
|
|
65
|
Kim CK, Kita H, Callaway Z, Kim HB, Choi
J, Fujisawa T, Shin BM and Koh YY: The roles of a Th2 cytokine and
CC chemokine in children with stable asthma: Potential implication
in eosinophil degranulation. Pediatr Allergy Immunol. 21:e697–e704.
2010.PubMed/NCBI View Article : Google Scholar
|
|
66
|
Rojas-Ramos E, Avalos AF, Pérez-Fernandez
L, Cuevas-Schacht F, Valencia-Maqueda E and Terán LM: Role of the
chemokines RANTES, monocyte chemotactic proteins-3 and -4, and
eotaxins-1 and -2 in childhood asthma. Eur Respir J. 22:310–316.
2003.PubMed/NCBI View Article : Google Scholar
|
|
67
|
Semik-Orzech A, Barczyk A, Wiaderkiewicz R
and Pierzchała W: Eotaxin, but not IL-8, is increased in upper and
lower airways of allergic rhinitis subjects after nasal allergen
challenge. Allergy Asthma Proc. 32:230–238. 2011.PubMed/NCBI View Article : Google Scholar
|
|
68
|
Paplińska-Goryca M, Nejman-Gryz P, Górska
K, Białek1-Gosk K, Hermanowicz-Salamon J and Krenke R: Expression
of inflammatory mediators in induced sputum: Comparative study in
asthma and COPD. Adv Exp Med Biol. 1040:101–112. 2018.PubMed/NCBI View Article : Google Scholar
|
|
69
|
Schuh JM, Blease K, Kunkel SL and Hogaboam
CM: Eotaxin/CCL11 is involved in acute, but not chronic, allergic
airway responses to Aspergillus fumigatus. Am J Physiol Lung
Cell Mol Physiol. 283:L198–L204. 2002.PubMed/NCBI View Article : Google Scholar
|
|
70
|
Mori A, Ogawa K, Kajiyama Y, Suko M and
Kaminuma O: Th2-cell-mediated chemokine synthesis is involved in
allergic airway inflammation in mice. Int Arch Allergy Immunol. 140
(Suppl 1):55–58. 2006.PubMed/NCBI View Article : Google Scholar
|
|
71
|
Arizmendi NG, Abel M, Puttagunta L,
Asaduzzaman M, Davidson C, Karimi K, Forsythe P and Vliagoftis H:
Mucosal exposure to cockroach extract induces allergic
sensitization and allergic airway inflammation. Allergy Asthma Clin
Immunol. 7(22)2011.PubMed/NCBI View Article : Google Scholar
|
|
72
|
Wu D, Zhou J, Bi H, Li L, Gao W, Huang M,
Adcock IM, Barnes PJ and Yao X: CCL11 as a potential diagnostic
marker for asthma? J Asthma. 51:847–854. 2014.PubMed/NCBI View Article : Google Scholar
|
|
73
|
Špadijer Mirković C, Perić A, Vukomanović
Đurđević B and Vojvodić D: Effects of fluticasone furoate nasal
spray on parameters of eosinophilic inflammation in patients with
nasal polyposis and perennial allergic rhinitis. Ann Otol Rhinol
Laryngol. 126:573–580. 2017.PubMed/NCBI View Article : Google Scholar
|
|
74
|
Thornton MA, Akasheh N, Walsh MT, Moloney
M, Sheahan PO, Smyth CM, Walsh RM, Morgan RM, Curran DR, Walsh MT,
et al: Eosinophil recruitment to nasal nerves after allergen
challenge in allergic rhinitis. Clin Immunol. 147:50–57.
2013.PubMed/NCBI View Article : Google Scholar
|
|
75
|
Chae SC, Park YR, Oh GJ, Lee JH and Chung
HT: The suggestive association of eotaxin-2 and eotaxin-3 gene
polymorphisms in Korean population with allergic rhinitis.
Immunogenetics. 56:760–764. 2005.PubMed/NCBI View Article : Google Scholar
|
|
76
|
Singh SR, Sutcliffe A, Kaur D, Gupta S,
Desai D, Saunders R and Brightling CE: CCL2 release by airway
smooth muscle is increased in asthma and promotes fibrocyte
migration. Allergy. 69:1189–1197. 2014.PubMed/NCBI View Article : Google Scholar
|
|
77
|
Little FF, Delgado DM, Wexler PJ,
Oppenheim FG, Mitchell P, Feldman JA, Walt DR, Peng RD and Matsui
EC: Salivary inflammatory mediator profiling and correlation to
clinical disease markers in asthma. PLoS One.
9(e84449)2014.PubMed/NCBI View Article : Google Scholar
|
|
78
|
Simons B, Ferrini ME, Carvalho S, Bassett
DJ, Jaffar Z and Roberts K: PGI2 controls pulmonary NK cells that
prevent airway sensitization to house dust mite allergen. J
Immunol. 198:461–471. 2017.PubMed/NCBI View Article : Google Scholar
|
|
79
|
Mellado M, Martín de Ana A, Gómez L,
Martínez C and Rodríguez-Frade JM: Chemokine receptor 2 blockade
prevents asthma in a cynomolgus monkey model. J Pharmacol Exp Ther.
324:769–775. 2008.PubMed/NCBI View Article : Google Scholar
|
|
80
|
Batra J, Das S, Chatterjee R, Chandra S
and Ghosh B: Monocyte chemotactic protein (MCP3) promoter
polymorphism is associated with atopic asthma in the Indian
population. J Allergy Clin Immunol. 128:239–242.e3. 2011.PubMed/NCBI View Article : Google Scholar
|
|
81
|
Zhang YL, Han DH, Kim DY, Lee CH and Rhee
CS: Role of Interleukin-17A on the chemotactic responses to CCL7 in
a murine allergic rhinitis model. PLoS One.
12(e0169353)2017.PubMed/NCBI View Article : Google Scholar
|
|
82
|
Saad-El-Din Bessa S, Abo El-Magd GH and
Mabrouk MM: Serum chemokines RANTES and monocyte chemoattractant
protein-1 in Egyptian patients with atopic asthma: Relationship to
disease severity. Arch Med Res. 43:36–41. 2012.PubMed/NCBI View Article : Google Scholar
|
|
83
|
Keskin O, Keskin M, Kucukosmanoglu E,
Ozkars MY, Gogebakan B, Kul S, Bayram H and Coskun Y: Exhaled
RANTES and interleukin 4 levels after exercise challenge in
children with asthma. Ann Allergy Asthma Immunol. 109:303–308.
2012.PubMed/NCBI View Article : Google Scholar
|
|
84
|
Marcella R, Croce A, Moretti A, Barbacane
RC, Di Giocchino M and Conti P: Transcription and translation of
the chemokines RANTES and MCP-1 in nasal polyps and mucosa in
allergic and non-allergic rhinopathies. Immunol Lett. 90:71–75.
2003.PubMed/NCBI View Article : Google Scholar
|
|
85
|
Xie ZK, Zhao H, Huang J and Xie ZF: The
regulated upon activation normal T-cell expressed and secreted
(RANTES) -28C/G and -403G/A polymorphisms and asthma risk: A
meta-analysis. Mol Diagn Ther. 18:523–531. 2014.PubMed/NCBI View Article : Google Scholar
|
|
86
|
Kalayci O, Sonna LA, Woodruff PG, Camargo
CA Jr, Luster AD and Lilly CM: Monocyte chemotactic protein-4
(MCP-4; CCL-13): A biomarker of asthma. J Asthma. 41:27–33.
2004.PubMed/NCBI View Article : Google Scholar
|
|
87
|
Odaka M, Matsukura S, Kuga H, Kokubu F,
Kasama T, Kurokawa M, Kawaguchi M, Ieki K, Suzuki S, Watanabe S, et
al: Differential regulation of chemokine expression by Th1 and Th2
cytokines and mechanisms of eotaxin/CCL-11 expression in human
airway smooth muscle cells. Int Arch Allergy Immunol. 143 (Suppl
1):84–88. 2007.PubMed/NCBI View Article : Google Scholar
|
|
88
|
Baumann R, Rabaszowski M, Stenin I,
Tilgner L, Scheckenbach K, Wiltfang J, Schipper J, Chaker A and
Wagenmann M: Comparison of the nasal release of IL-4, IL-10, IL-17,
CCL13/MCP-4, and CCL26/eotaxin-3 in allergic rhinitis during season
and after allergen challenge. Am J Rhinol Allergy. 27:266–272.
2013.PubMed/NCBI View Article : Google Scholar
|
|
89
|
Rimaniol AC, Till SJ, Garcia G, Capel F,
Godot V, Balabanian K, Durand-Gasselin I, Varga EM, Simonneau G,
Emilie D, et al: The CX3C chemokine fractalkine in allergic asthma
and rhinitis. J Allergy Clin Immunol. 112:1139–1146.
2003.PubMed/NCBI View Article : Google Scholar
|
|
90
|
El-Shazly A, Berger P, Girodet PO, Ousova
O, Fayon M, Vernejoux JM, Marthan R and Tunon-de-Lara JM:
Fraktalkine produced by airway smooth muscle cells contributes to
mast cell recruitment in asthma. J Immunol. 176:1860–1868.
2006.PubMed/NCBI View Article : Google Scholar
|
|
91
|
Mionnet C, Buatois V, Kanda A, Milcent V,
Fleury S, Lair D, Langelot M, Lacoeuille Y, Hessel E, Coffman R, et
al: CX3CR1 is required for airway inflammation by promoting T
helper cell survival and maintenance in inflamed lung. Nat Med.
16:1305–1312. 2010.PubMed/NCBI View Article : Google Scholar
|
|
92
|
Kaplan AP: Chemokines, chemokine receptors
and allergy. Int Arch Allergy Immunol. 124:423–431. 2001.PubMed/NCBI View Article : Google Scholar
|
|
93
|
Liu LY, Bates ME, Jarjour NN, Busse WW,
Bertics PJ and Kelly EA: Generation of Th1 and Th2 chemokines by
human eosinophils: Evidence for a critical role of TNF-α. J
Immunol. 179:4840–4848. 2007.PubMed/NCBI View Article : Google Scholar
|
|
94
|
Liu L, Jarjour NN, Busse WW and Kelly EA:
Enhanced generation of helper T type 1 and 2 chemokines in
allergen-induced asthma. Am J Respir Crit Care Med. 169:1118–1124.
2004.PubMed/NCBI View Article : Google Scholar
|
|
95
|
Lun SW, Wong CK, Ko FW, Ip WK, Hui DS and
Lam CW: Aberrant expression of CC and CXC chemokines and their
receptors in patients with asthma. J Clin Immunol. 26:145–152.
2006.PubMed/NCBI View Article : Google Scholar
|
|
96
|
Lai ST, Hung CH, Hua YM, Hsu SH, Jong YJ
and Suen JL: T-helper 1-related chemokines in the exacerbation of
childhood asthma. Pediatr Int. 50:99–102. 2008.PubMed/NCBI View Article : Google Scholar
|
|
97
|
Ragusa F and Fallahi P: IP-10 in
occupational asthma: Review of the literature and case-control
study. Clin Ter. 168:e151–e157. 2017.PubMed/NCBI View Article : Google Scholar
|
|
98
|
Tworek D, Kuna P, Młynarski W, Górski P,
Pietras T and Antczak A: MIG (CXCL9), IP-10 (CXCL10) and I-TAC
(CXCL11) concentrations after nasal allergen challenge in patients
with allergic rhinitis. Arch Med Sci. 9:849–853. 2013.PubMed/NCBI View Article : Google Scholar
|
|
99
|
Romagnani P, Maggi L, Mazzinghi B, Cosmi
L, Lasagni L, Liotta F, Lazzeri E, Angeli R, Rotondi M and Filì L:
CXCR3-mediated opposite effects of CXCL10 and CXCL4 on TH1 or TH2
cytokine production. J Allergy Clin Immunol. 116:1372–1379.
2005.PubMed/NCBI View Article : Google Scholar
|
|
100
|
Ko FW, Lun SW, Wong CK, Szeto CC, Lam CW,
Leung TF and Hui DS: Decreased T-bet expression and changes in
chemokine levels in adults with asthma. Clin Exp Immunol.
147:526–532. 2007.PubMed/NCBI View Article : Google Scholar
|
|
101
|
Lun SW, Wong CK, Ko FW, Ip WK, Hui DS and
Lam CW: Aberrant expression of CC and CXC chemokines and their
receptors in patients with asthma. J Clin Immunol. 26:145–152.
2006.PubMed/NCBI View Article : Google Scholar
|
|
102
|
Sawant KV, Poluri KM, Dutta AK, Sepuru KM,
Troshkina A, Garofalo RP and Rajarathnam K: Chemokine CXCL1
mediated neutrophil recruitment: Role of glycosaminoglycan
interactions. Sci Rep. 6(33123)2016.PubMed/NCBI View Article : Google Scholar
|
|
103
|
Cho MK, Park MK, Kang SA, Caballero ML,
Perez-Pinar T, Rodriguez-Perez R, Ock MS, Cha HJ, Hong YC and Yu
HS: Allergenicity of two Anisakis simplex allergens evaluated in
vivo using an experimental mouse model. Exp Parasitol. 146:71–77.
2014.PubMed/NCBI View Article : Google Scholar
|
|
104
|
Kim EH, Kim JH, Samivel R, Bae JS, Chung
YJ, Chung PS, Lee SE and Mo JH: Intralymphatic treatment of
flagellin-ovalbumin mixture reduced allergic inflammation in murine
model of allergic rhinitis. Allergy. 71:629–639. 2016.PubMed/NCBI View Article : Google Scholar
|
|
105
|
Chenuet P, Fauconnier L, Madouri F,
Marchiol T, Rouxel N, Ledru A, Mauny P, Lory R, Uttenhove C, van
Snick J, et al: Neutralization of either IL-17A or IL-17F is
sufficient to inhibit house dust mite induced allergic asthma in
mice. Clin Sci (Lond). 131:2533–2548. 2017.PubMed/NCBI View Article : Google Scholar
|
|
106
|
Al-Alwan LA, Chang Y, Mogas A, Halayko AJ,
Baglole CJ, Martin JG, Rousseau S, Eidelman DH and Hamid Q:
Differential roles of CXCL2 and CXCL3 and their receptors in
regulating normal and asthmatic airway smooth muscle cell
migration. J Immunol. 191:2731–2741. 2013.PubMed/NCBI View Article : Google Scholar
|
|
107
|
Dixit N and Simon SI: Chemokines,
selectins and intracellular calcium flux: Temporal and spatial cues
for leukocyte arrest. Front Immunol. 3(188)2012.PubMed/NCBI View Article : Google Scholar
|
|
108
|
Baos S, Calzada D, Cremades L, Sastre J,
Quiralte J, Florido F, Lahoz C and Cárdaba B: Biomarkers associated
with disease severity in allergic and nonallergic asthma. Mol
Immunol. 82:34–45. 2017.PubMed/NCBI View Article : Google Scholar
|
|
109
|
Charrad R, Kaabachi W, Rafrafi A, Berraies
A, Hamzaoui K and Hamzaoui A: IL-8 gene variants and expression in
childhood asthma. Lung. 195:749–757. 2017.PubMed/NCBI View Article : Google Scholar
|
|
110
|
Lee MF, Song PP, Hwang GY, Lin SJ and Chen
YH: Sensitization to Per a 2 of the American cockroach correlates
with more clinical severity among airway allergic patients in
Taiwan. Ann Allergy Asthma Immunol. 108:243–248. 2012.PubMed/NCBI View Article : Google Scholar
|
|
111
|
Yang T, Li Y, Lyu Z, Huang K, Corrigan CJ,
Ying S, Wang W and Wang C: Characteristics of proinflammatory
cytokines and chemokines in airways of asthmatics: Relationships
with disease severity and infiltration of inflammatory cells. Chin
Med J (Engl). 130:2033–2040. 2017.PubMed/NCBI View Article : Google Scholar
|
|
112
|
Wang X, Zhang N, Bo M, Holtappels G, Zheng
M, Lou H, Wang H, Zhang L and Bachert C: Diversity of TH cytokine
profiles in patients with chronic rhinosinusitis: A multicenter
study in Europe, Asia, and Oceania. J Allergy Clin Immunol.
138:1344–1353. 2016.PubMed/NCBI View Article : Google Scholar
|
|
113
|
Tomassen P, Vandeplas G, Van Zele T,
Cardell LO, Arebro J, Olze H, Förster-Ruhrmann U, Kowalski ML,
Olszewska-Ziąber A, Holtappels G, et al: Inflammatory endotypes of
chronic rhinosinusitis based on cluster analysis of biomarkers. J
Allergy Clin Immunol. 137:1449–1456.e4. 2016.PubMed/NCBI View Article : Google Scholar
|
|
114
|
Cui XY, Chen X, Yu CJ, Yang J, Lin ZP, Yin
M and Cheng L: Increased expression of toll-like receptors 2 and 4
and related cytokines in persistent allergic rhinitis. Otolaryngol
Head Neck Surg. 152:233–238. 2015.PubMed/NCBI View Article : Google Scholar
|
|
115
|
Pelikan Z: Cytokines in tears during the
secondary keratoconjunctival responses induced by allergic reaction
in the nasal mucosa. Ophthalmic Res. 52:32–42. 2014.PubMed/NCBI View Article : Google Scholar
|
|
116
|
Pelikan Z: Cytokine profiles in tears
accompanying the secondary conjunctival responses induced by nasal
allergy. Curr Eye Res. 39:120–132. 2014.PubMed/NCBI View Article : Google Scholar
|
|
117
|
Kim JA, Cho JH, Park IH, Shin JM, Lee SA
and Lee HM: Diesel exhaust particles upregulate interleukins IL-6
and IL-8 in nasal fibroblasts. PLoS One.
11(e0157058)2016.PubMed/NCBI View Article : Google Scholar
|
|
118
|
Ozturk AB, Bayraktar R, Gogebakan B,
Mumbuc S and Bayram H: Comparison of inflammatory cytokine release
from nasal epithelial cells of non-atopic non-rhinitic, allergic
rhinitic and polyp subjects and effects of diesel exhaust particles
in vitro. Allergol Immunopathol (Madr). 45:473–481. 2017.PubMed/NCBI View Article : Google Scholar
|
|
119
|
Hastie AT, Steele C, Dunaway CW, Moore WC,
Rector BM, Ampleford E, Li H, Denlinger LC, Jarjour N, Meyers DA,
et al: NHLBI Severe Asthma Research Program (SARP): Complex
association patterns for inflammatory mediators in induced sputum
from subjects with asthma. Clin Exp Allergy. 48:787–797.
2018.PubMed/NCBI View Article : Google Scholar
|
|
120
|
Miyajima S, Shigehara K, Kamekura R,
Takaki H, Yabe H, Ikegami I, Asai Y, Nishikiori H, Chiba H, Uno E,
et al: Activated circulating T follicular helper cells and skewing
of T follicular helper 2 cells are down-regulated by treatment
including an inhaled corticosteroid in patients with allergic
asthma. Allergol Int. 69:66–77. 2020.PubMed/NCBI View Article : Google Scholar
|
|
121
|
Fujita K, Kobayashi M, Brutkiewicz RR,
Hanafusa T, Herndon DN and Suzuki F: Role for IL-4 nonproducing NKT
cells in CC-chemokine ligand 2-induced Th2 cell generation. Immunol
Cell Biol. 84:44–50. 2006.PubMed/NCBI View Article : Google Scholar
|
|
122
|
Bilenki L, Yang J, Fan Y, Wang S and Yang
X: Natural killer T cells contribute to airway inflammation induced
eosinophilic by ragweed through enhanced IL-4 and eotaxin
production. Eur J Immunol. 34:345–354. 2004.PubMed/NCBI View Article : Google Scholar
|
|
123
|
Hong GU, Kim NG, Kim TJ and Ro JY: CD1d
expressed in mast cell surface enhances IgE production in B cells
by up-regulating CD40L expression and mediator release in allergic
asthma in mice. Cell Signal. 26:1105–1117. 2014.PubMed/NCBI View Article : Google Scholar
|
|
124
|
Karisola P, Lehto M, Kinaret P, Ahonen N,
Haapakoski R, Anthoni M, Taniguchi M, Wolff H, Puustinen A and
Alenius H: Invariant natural killer T cells play a role in
chemotaxis, complement activation and mucus production in a mouse
model of airway hyperreactivity and inflammation. PLoS One.
10(e0129446)2015.PubMed/NCBI View Article : Google Scholar
|
|
125
|
Woo YD, Koh J, Kang HR, Kim HY and Chung
DH: The invariant natural killer T cell-mediated chemokine X-C
motif chemokine ligand 1-X-C motif chemokine receptor 1 axis
promotes allergic airway hyperresponsiveness by recruiting
CD103+ dendritic cells. J Allergy Clin Immunol.
142:1781–1792.e12. 2018.PubMed/NCBI View Article : Google Scholar
|