Introduction
Sperm cryopreservation is not only a routine part of
the daily work performed at sperm banks but also an indispensable
part of assisted reproductive technology. Ultralow temperatures may
lead to impaired sperm motility and the formation of ice crystals
during freezing and thawing, as well as excessive oxidative stress
during this process that may damage the structure and function of
sperm (1,2). According to the World Health
Organization (WHO), ~50% of sperm cells are damaged or killed
during thawing (3). The sperm is
mixed with cryopreservation agents prior to freezing in order to
reduce its mortality (4). The most
commonly used cryopreservation agent is glycerol-egg-yolk-citrate
(GEYC) cryopreservative (5).
However, conventional cryopreservation agents are not sufficiently
able to preserve weak sperm (2). In
recent years, domestic and international studies have reported on
the addition of natural or synthetic substances to cryopreservation
agents to reduce the freezing-associated damage of sperm (2,6-9).
Shahrzad and other researchers (10-12)
indicated that the addition of L-carnitine to cryopreservative may
effectively lower the sperm DNA fragmentation index and apoptosis.
Li et al (13) indicated that
the addition of 200 nmol/l mitochondria-targeted antioxidant
mitoquinone (MitoQ) to cryopreservative may to markedly improve the
quality of cryopreserved sperm, and it was also proposed that MitoQ
could also be used as a cryopreservation additive for semen.
Berkovitz et al (14)
indicated that Zn2+ may also improve the motility of
cryopreserved sperm cells. An increasing number of studies have
also made efforts to identify natural protective agents that are
able to improve the motility of cryopreserved sperm (2,6,7).
Lycium barbarum, also known as ‘Oriental
magic grass’, is commonly used in the Ningxia Hui Autonomous Region
of northwest China, as a traditional Chinese medicine to treat male
infertility, hypertension, high cholesterol and diabetes (15). In recent years, Lycium
barbarum polysaccharides (LBP) extracted from this plant have
been used as antioxidants, scavenging oxygen free radicals and
stabilizing cell membranes (16-20).
A previous study indicated that the addition of LBP during semen
treatment was able to protect sperm structure and function
(21). Therefore, in the present
study, LBP was added into the GEYC cryopreservative to investigate
its effect on the structure and function of mitochondria of weak
sperm, to provide a theoretical basis for further research into
cryopreservation agents that may be beneficial in maintaining the
function of cryopreserved sperm. Sperm motility requires energy
derived from the ATP produced by mitochondria. Therefore, the
structure and function of mitochondria after freezing are crucial
to sperm motility (9). During the
sperm freeze-thawing process, a considerable amount of reactive
oxygen species (ROS), which can damage the mitochondrial membrane,
are produced, resulting in a significant decrease in sperm motility
(1,2). In the present study, the four key
regulators of mitochondrial apoptosis pathways, including Bcl-2,
Bax, cytochrome C (CytC) and caspase-3(22), were assessed as indicators for
mitochondrial apoptosis pathway activation. The effects of LBP on
the freeze-thawing of weak sperm and on the key molecules in these
apoptosis pathways, were studied by measuring the level of ROS. The
aim was to further explore the protective mechanism of improved
cryopreservative and to identify more effective protective agents
that can be used in sperm cryopreservation.
Materials and methods
Sample collection and selection
criteria
Between May and June 2018, 22 patients with
asthenospermia, aged 22-45 (mean age 31.9±8.8) years and with a
semen volume of 2.4±0.6 ml were selected from the reproductive
center of Ningxia Medical University General Hospital. The
selection criteria were in accordance with the WHO Human Semen
Examination and Processing Laboratory Manual (Fifth Edition)
(23) and were as follows: Semen
volume, ≥1.5 ml; total sperm number, ≥39 (106/ml); sperm
forward progressive motility (PR) <32%; sperm [PR +
non-progressive motility (NP)] <40%. Samples with a sperm
concentration range of 6-7x107/ml were used in further
experiments. In order to eliminate dysliquefaction and severe
oligospermia, peroxidase staining was used to eliminate
leukocytospermia (peroxidase-positive leukocytes;
<1.0x106/ml). To perform peroxidase staining 125 mg
benzidine was dissolved in 50 ml 95% methanol. A 150 mg quantity of
rose red B was dissolved in 50 ml of distilled water and the two
solutions were mixed. A 1 ml volume of this mixed solution had 2
drops of H2O2 added and the prepared solution
formed the benzidine dye. A drop of semen was placed on a slide
with a drop of benzidine dye solution and mixed well. The slide was
covered, placed at 37˚C for 20 min and observed under a light
microscope (x1,000). White blood cells were brown after staining.
The number of leukocytes were observed in 200 sperm visual fields.
The present study was approved by the ethics committee of the
General Hospital of Ningxia Medical University (Yinchuan, China)
and written informed consent was obtained from the patients.
Sample preparation
All the subjects were abstinent for 3-5 days. The
semen samples were collected by masturbation into aseptic seminal
cups. Each semen sample was first placed in a 37˚C water bath for
30 min. After the sample was completely liquefied and evenly mixed,
1,500 µl of each semen sample was divided into three groups: Fresh
group (unfrozen sperm), GEYC group (frozen sperm with standard
protectant) and LBP group (frozen group with standard protectant
and added LBP; 500 µl semen in each group). After liquefication,
the semen was diluted with 500 µl Biggers, Whitten and Whittingham
(BWW) sperm culture solution (Beijing Solarbio Science &
Technology Co., Ltd.; cat. no. G2586), as recommended by the WHO
guidelines (23), and centrifuged at
500 x g for 10 min at room temperature. The supernatant was then
discarded, and 1 ml sperm culture solution was added (Fresh: 1 ml
BWW; GEYC: 1 ml BWW, LBP: 1 ml BWW containing 1,000 µg/ml LBP),
followed by incubation at 37˚C for 20 min.
The sperm was then frozen; the cryoprotectant base
fluid was prepared according to the protocol for the GEYC
protectant recommended by the WHO guidelines (23).
Cryopreservation and cryo-survival of
sperm
The manual two-step freezing method was employed
(9). Semen and protectant were added
to the cryopreservation tube at a ratio of 2:1 and the
cryopreservation tube was placed at a height of ~5 cm from the
liquid nitrogen surface for 10 min. Subsequently, it was lowered to
0.1 cm above the liquid level (no contact with the liquid level)
for 5 min and subsequently immersed in liquid nitrogen for
preservation.
Sperm cryo-survival
The cryopreservation tube was taken out of the
liquid nitrogen and placed in a 37˚C water bath for
rewarming for 3 min. The sample was then centrifuged 300 x g for 5
min at room temperature and placed into the BWW culture medium for
10 min at 37˚C. Subsequently, the suspension was gently
reversed 10 times and evenly mixed and then sperm was then
subsequently tested for viability.
Detection of sperm-associated activity
parameters
Computer-aided sperm analysis technology (CASA) was
used to analyse the relevant parameters. Prior to the assessment of
sperm viability, the counting plate [Sefi-Medical Instruments
(9), Ltd.] was placed on the
temperature control plate of the microscope in advance. This was a
single chamber system. A total of six representative fields were
examined under light microscopy (x200), at least 200 spermatozoa
were assessed for each sample. The test was repeated twice. The
color sperm quality detection system (WLJY-9DOO; version 2.0;
Beijing Weili New Century Science & Tech. Deve. Co., Ltd.) was
used to determine the sperm vitality [percentage of sperm forward
movement (PR)] and other relevant parameters [curvilinear velocity
(VCL), straight linear velocity (VSL), sperm average path velocity
(VAP), amplitude of lateral head displacement (ALH) and beat cross
frequency (BCF)] sperm concentration was 2-50x106. Fast
forward motion with a speed ≥25 µm/sec was classified as class A;
if the velocity of forward movement was <25 µm/sec, it was
classified as class B (PR=A+B); if the velocity of non-forward
movement or forward movement was ≤5 µm/S, it was classified as
class C (NP=C); Total immobility was classified as level D
(immobility=D). The spermatozoa were incubated 37˚C for
10 min prior to measuring motility. If the sperm concentration was
>50x106, considering the high frequency of sperm
collision producing errors, the semen was diluted with BWW to
2-50x106 before assessment.
Sperm survival rate experiment
The survival rate was assessed using eosin aniline
black (Beijing Solarbio Science & Technology, Co., Ltd.; cat.
no. G2581), as recommended by the WHO guidelines (23). The sperm concentration was adjusted
to 2-50x106 using normal saline and 25 µl of semen and
an equal volume of eosin aniline black suspension were added to a
1.5 ml centrifuge tube, mixed with micropipette at room temperature
and left to stand for 30 sec. A sperm smear was made with each
sperm suspension, dried and checked immediately after drying. Each
glass slide was examined using a light-field microscope under an
oil immersion objective (magnification, x1,000). Finally, with the
help of a sperm counter (cat. no. XK06-6J; Jiangsu Xinkang Medical
Devices, Co., Ltd.), the stained and non-stained sperm were
counted. Each sample was evaluated twice and 200 sperm from each
sample were evaluated to achieve an acceptably low sampling error.
If the difference was too large, re-sampling and re-evaluation were
performed. Fig. 1 presents the
evaluation criteria used.
Sperm morphology test
The sperm morphology was observed using a diff Quik
rapid staining kit (cat. no. G1541; Beijing Solarbio Science &
Technology Co., Ltd.) First, the sperm concentration was adjusted
to (2-50x106) using normal saline, and a sperm smear was
then prepared and immersed in fast dye solution I at room
temperature for 10 sec, followed by fast dye solution II at room
temperature for 5 sec. The slide was then rinsed under running
water to remove excess dye solution. A total of 200 sperms were
evaluated after sealing under a light microscope (x1,000). The
numbers of normal and abnormal spermatozoa were recorded, and the
percentage was calculated. This staining method is recommended by
the WHO laboratory manual for human semen examination and
processing (Fifth Edition) (23).
Sperm function tests DNA fragment
index (DFI) test
The DFI of sperm was assessed using a sperm
chromatin structure assay (SCSA) (24). A DFI cell flow cytometer test kit
(cat. no. CP0101-10T; Cellpro; Zhejiang Xingbo Biological
Technology Co., Ltd.) was selected for testing; this kit has been
approved by the State Food and Drug Administration (SFDA) for
clinical semen examination (approval no. 20160051). Solution A
contained: 0.1 mol/l NaCl, 0.4 mmol/l KH2PO4,
2.7 mmol/l C6H6O6, 4.7 mmol/l KCl,
0.2 mmol/l MgSO4•7H2O, 2 mmol/l
CaCl2•2H2O, 0.02 mol/l HEPES-Na, 4 mmol/l
NaHCO3, 0.3 mmol/l
C3H3O3Na, 0.34% (v/v)
C3H5O3Na, 40.5% (v/v) Percol.
Solution B contained 0.1 mol/l NaCl, 0.4 mmol/l
KH2PO4, 2.7 mmol/l
C6H6O6, 4.7 mmol/l KCl, 0.2 mmol/l
MgSO4•7H2O, 2 mmol/l
CaCl2•2H2O, 0.02 mol/l HEPES-Na, 4 mmol/l
NaHCO3, 0.3 mmol/l
C3H3O3Na, 81% (v/v) Percol. 1%
(v/v) Triton X-100 (Beijing Solarbio Science & Technology Co.,
Ltd.; T8200); Solution C contained 6 µg/ml acridine orange, 37
mmol/l citric acid, 126 mmol/l Na2HPO4, 1
mmol/l disodium EDTA, 0.15 mol/l NaCl. The specific operation was
as follows: Sperm 1-2x106/ml was diluted with solution A
to a final volume of 100 µl. Subsequently, 200 µl of solution B was
added, followed by incubation at room temperature for 30 sec
(accurate timing was required). A total of 600 µl solution C was
added and a BD accuri C6 (BD Biosciences) flow cytometer was used
to select the cell groups to be studied on the working interface.
The sperm flow rate was controlled at 100-300/sec and after two
minutes of stable operation, at least 5,000 sperm were collected,
and each sample was assessed at least twice. After data collection,
the flow cytometry (FCS) files were copied into DFI-View software
(DFIView; version 1.10; Cellpro; Zhejiang Xingbo Biological
Technology Co., Ltd.) for analysis.
Sperm mitochondrial membrane potential
(MMP) test
The sperm tail interruption staining kit (cat. no.
CP01014-10T; Cellpro; Zhejiang Xingbo Biological Technology Co.,
Ltd.) was used for the detection of MMP. The kit has been approved
by the SFDA for clinical semen examination (zyxb no. 20160052). The
specific operation was as follows: The semen was diluted to
2x106/ml and 0.5 ml JC-1 working solution was added,
followed by incubation at 37˚C for 20 min and centrifugation at 4˚C
and 300 x g for 5 min. The supernatant was removed, and the
precipitated cells were washed twice with JC-1 buffer solution, and
the cells were analyzed using a BD Accuri C6 (BD Biosciences) with
an excitation wavelength of 488 nm and emission wavelength was 530
nm. The sperm flow rate was controlled at 100-300/sec, and at least
5,000 sperm were collected, and each sample was detected at least
twice. Following data collection, the FCS files were copied into
MMP-test software (MMP-Test; Version 1.0; Cellpro; Zhejiang Xingbo
Biological Technology Co., Ltd.) for analysis. For sperm with a
high MMP, JC-1 spontaneously forms JC-1 aggregates with intense red
fluorescence, which is measured on the FL2 channel. For sperm with
low MMP, JC-1 remains in the monomer form and exhibits green
fluorescence, which is measured on the FL1 channel (25).
Acrosome reaction (AR) of sperm
test
The Sperm Acrosome Staining kit (acrosome test; cat.
no. CP01011-10T; Cellpro; Zhejiang Xingbo Biological Technology
Co., Ltd.) was selected for detection. The kit has been approved by
the SFDA for clinical semen examination (approval no. 20160054).
The specific operation was as follows: The sperm concentration was
adjusted to 4-5x106 live sperm using solution A; two
1.5-ml centrifuge tubes were prepared and 10 µl solution B and 10
µl solution C was added. The sample was then mixed and incubated at
37˚C for 15 min, and 100 µl pre-cooled 70% ethanol was added into
each tube, followed by incubation for 2 min at room temperature.
The mixture was centrifuged at 500 x g for 5 min at room
temperature, the supernatant was discarded and 500 µl staining
working solution (mixture of solution D and solution E) was added,
followed by shaking and mixing. Subsequently, the mixture was
incubated for 15 min at 37˚C. On the BD Accuri C6 equipment the
voltage of the FL1 channel was adjusted so that the major group was
in the specified gates. The sperm flow rate was controlled at
100-300/sec, and at least 5,000 sperm were collected each sample
was analyzed at least twice. (Solution A, NaCl,
KH2PO4,
C6H6O6, KCl, NaHCO3;
Solution B, Ca2+ carrier A23187, DMSO, Solution C, DMSO;
Solution D, NaCl, KH2PO4, KCl,
NaH2PO4; Solution E, Isothiocyanate-coupled
pea agglutinin).
Detection of malondialdehyde
(MDA)
MDA was detected using thiobarbituric acid (TBA)
colorimetry. The MDA test kit of Nanjing Jiancheng Biological
Preparation Co., Ltd. was selected for testing. The operation was
performed according to the manufacturer's protocol and the
absorbance value at 532 nm wavelength. Was read using a
spectrophotometer (1410101 Thermo Scientific™ Multiskan™ FC; Thermo
Fisher Scientific, Inc.).
Detection of ROS
The sperm cell oxidative stress activity superoxide
anion flow cytometry (cat. no. GMS10096; GenMed, Ltd.) test kit was
used for the detection of ROS, and the operation was strictly in
accordance with the manufacturer's protocol. In this assay,
2',7'-dichlorofluorescein diacetate is a completely free dye that
passes through the cell membrane. Once it is oxidized by molecules
including hydrogen peroxide, peroxides and peroxynitrite anions, it
produces fluorescence (26). Using
this assay, the concentration of ROS in spermatozoa were measured.
The specific steps were as follows: Fresh semen (1 ml) was added to
a 1.5-ml centrifuge tube, followed by centrifugation at room
temperature for 10 min at a speed of 300 x g. After discarding the
supernatant, 500 ml of reagent C was added, followed by thorough
mixing. Subsequently, 25 µl of dye A and 25 µl of dye B was added
to the aforementioned sperm cells. Following thorough mixing, the
cells were incubated at room temperature (25˚C) for 40 min under
the exclusion of light. The cells were then centrifuged at room
temperature for 5 min at a speed of 300 x g. The supernatant was
carefully removed and 500 µl of storage solution (reagent C) was
added. Following mixing, dual-channel analysis was immediately
performed using the BD Accuri C6 (Beckton, Dickinson and Company)
with an excitation wavelength at 488 nm. The voltage of the FL1 and
FL2 channels was regulated to make the major group appear in the
specified gate. After the experiment, the data was imported into
FlowJo version 10 (FlowJo LLC) for analysis. The sperm flow rate
was controlled at 100/sec, and at least 10,000 sperms were
collected, and each sample was analyzed at least twice.
Immunofluorescence
The sperm smears were fixed in 4% paraformaldehyde
at room temperature for 20 min and then washed with PBS three
times, for 5 min/wash. The slides were placed in a humidified
chamber and 0.30% Triton-X 100 (Beijing Solarbio Science &
Technology Co., Ltd.; T8200) was used to penetrate the membrane at
room temperature for 10 min. PBS was used to wash the slides
thrice, for 5 min/wash. Subsequently, two drops of 3% peroxidase
inhibitor were added to each slide to block the endogenous
peroxidase at room temperature. The slides were incubated at room
temperature for 10 min and then washed by PBS three times, for 5
min/wash. To block endogenous biotin and avidin, normal goat serum
(Beijing Solarbio Science & Technology Co., Ltd.; cat. no.
SL038) was added to the sample, followed by incubation at room
temperature for 10 min. The samples were then incubated with
primary antibodies in blocking solution overnight at 4˚C. The
primary antibodies used were as follows: Rabbit anti-human Bcl-2
(cat. no., 12789-1-AP; ProteinTech Group, Inc.), Bax (cat. no.
50599-2-Ig; ProteinTech Group, Inc.), CytC (cat. no. 12245-1-AP;
ProteinTech Group, Inc.), and caspase-3 (cat. no. 19677-1-AP;
ProteinTech Group, Inc.) antibody (1:200 dilution; 50 µl/tablet. BS
was used instead of the primary antibody in the negative control
group.
The slides were then incubated with secondary
antibody (goat anti-rabbit IgG-H&L (cat. no. ab6721; 1:1,000;
Abcam) for 1 h at room temperature and after washing with PBS,
samples were mounted with DAPI (cat. no. ZLI-9557; OriGene
Technologies, Inc.). Confocal scanning images were captured using a
DM 2000 microscope (x1,000; Leica Microsystems) with a digital
camera (cat. no. DFC450C; Leica Microsystems).
Western blot analysis
Protein was extracted with a whole protein
extraction kit (cat. no. KGP2100; Nanjing KeyGen Biotech Co., Ltd.)
and protein was then quantified using the BCA method (cat. no.
E162-01; Nanjing KeyGen Biotech Co., Ltd.). In each lane, 18-20 µg
protein lysate was separated by 10% SDS-PAGE and proteins were
transferred to PVDF membranes (Merck KGaA) and the membranes were
blocked in 5% non-fat milk (blocking solution) at room temperature
for 1 h. The membrane was then incubated with primary antibodies
overnight at 4˚C. The membranes were washed with Tris-buffered
saline containing Tween-20 three times and then incubated with
horseradish peroxidase-conjugated goat anti-rabbit IgG-H&L
(cat. no. ab6721; 1:5,000; Abcam) secondary antibody at room
temperature for 1 h. The PVDF membranes were then visualized using
an enhanced chemiluminescence (ECL) detection system reagents and
images were captured with the ChemiDocXRS (Bio-Rad Laboratories,
Inc.). After the experiment, the images were assessed using ImageJ
version 1.8.0 (National Institutes of Health). The primary
antibodies were as follows: Caspase-3 (1:10,000; cat. no.
19677-1-AP; ProteinTech Group, Inc.), Bax (1:5,000; cat. no.
50599-2-Ig; ProteinTech Group, Inc.), Bcl-2 (1:10,000; cat. no.,
12789-1-AP; ProteinTech Group, Inc.), CytC (1:1,000; cat. no.
12245-1-AP; ProteinTech Group, Inc.), GAPDH (1:10,000; cat. no.
10494-1-AP; ProteinTech Group, Inc.).
Determination of RNA levels
Total RNA from each group was extracted with a total
RNA preparation kit (cat. no. AP-MN-MS-RNA-520G; Corning, Inc.) and
reverse transcribed (42˚C for 15 min, then 85˚C for 5 sec) into
complementary (c)DNA using transcript all in one first-strand cDNA
synthesis super mix (cat. no. AT341-02; TransGen Biotech Co.,
Ltd.). The SYBR premix ex taq II dye fluorescence quantitative kit
(cat. no. RR820A; Takara Biotechnology Co., Ltd.) was used for PCR
amplification in an ABI 7500 fast quantitative PCR instrument
(Thermo Fisher Scientific, Inc.). After 30 sec of denaturation at
95˚C, 40 cycles of amplification were performed: 5 sec of
denaturation at 95˚C, annealing/elongation at 60˚C for 30 sec. The
primer sequences were as follows: Caspase-3 forward,
5'-TGGAATGTCAGCTCGCAATG-3' and reverse, 5'-CAGGTCCGTTCGTTCCAAAA-3';
Bax forward, 5'-AGATCATGAAGACAGGGGCC-3' and reverse,
5'-ATCCTCTGCAGCTCCATGTT-3'; Bcl-2 forward,
5'-AACTCTTCAGGGATGGGGTG-3' and reverse, 5'-GCTGGGGCCATATAGTTCCA-3';
CytC forward, 5'GCCTCAATGTCCCTGCTTCT-3' and reverse,
5'-TCGTTTCCAGCATGAGTGCT-3'; GAPDH forward,
5'-AGTCTACTGGCGTCTTCACC-3' and reverse,
5'-CCACGATGCCAAAGTTGTCA3'.
Transmission electron microscopy
(TEM)
After liquefaction, the semen samples were washed
twice with 0.85% NaCl and centrifuged at room temperature 1,000 x g
for 10 min. The supernatant was then discarded and then 2.5%
glutaraldehyde was added slowly, and samples were fixed in a 4˚C
refrigerator for 2-4 h. Subsequently, the samples were fixed with
osmium acid (1%), washed with distilled water, embedded for 2 h at
room temperature in Epon812 epoxy resin, cut into semi thin section
(1 nm), stained at room temperature for 1 h by toluidine blue and
positioned under a light microscope. The sperm cell area was
subjected to semi thin section (1 nm) and a JEM1200-EX TEM (JEOL,
Ltd.) was used to observe the changes of mitochondria in the sperm
tail. An experienced electron microscopist blinded to the origin of
the samples tested and recorded the information. The specific
evaluation methods were as follows: Under an electron microscope,
at least 100 sperm from each group of samples was evaluated,
focusing on evaluation of the microstructure of mitochondria in the
tail of the sperm, observing whether the arrangement of
mitochondria was orderly, whether there was oedema and the severity
of oedema in mitochondria and whether there was damage to the
structure of the mitochondrial membrane.
Statistical analysis
Values are expressed as the mean ± standard error of
the mean unless otherwise noted in the figure legends. Statistical
analysis was performed using a SPSS 24.0 statistical package (IBM
Corp.), differences among multiple groups were analyzed by Welch's
one-way ANOVA, following by Tukey's HSD test. P<0.05 was
considered to indicate a statistically significant difference.
Results
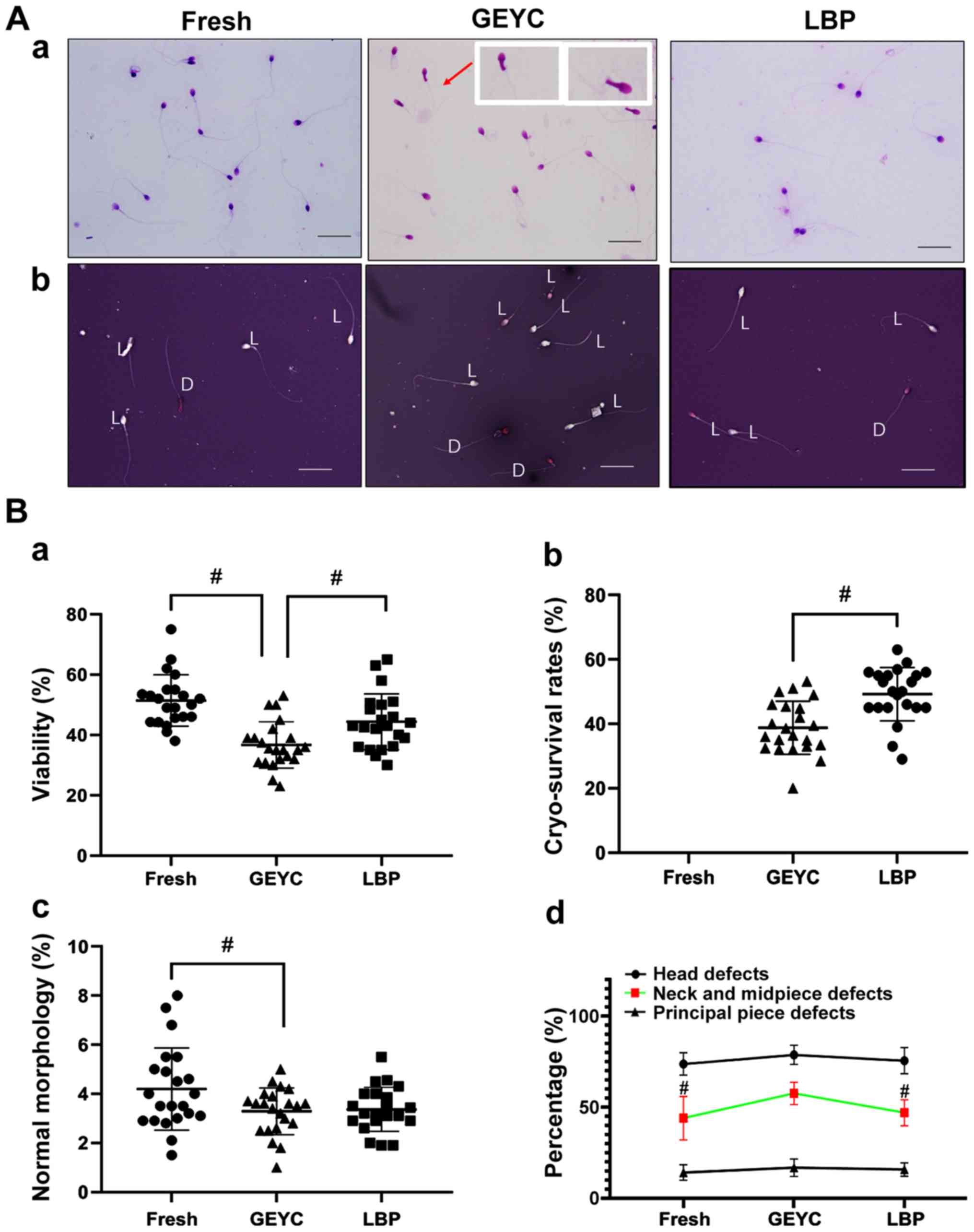
The experimental images of sperm morphology and the
sperm survival rate are presented in Fig. 1A, from which it can be seen that
sperm neck abnormality (Fig. 1A-a)
and sperm mortality (Fig. 1A-b) are
indicated to increase during the freezing process. The results also
indicated that the sperm viability decreased after sperm
cryopreservation. The viability of the LBP group was higher
compared with the GEYC group (P<0.05), however, the viability of
the GEYC group was lower compared with the Fresh group (P<0.05),
as presented in Fig. 1B-a. The sperm
cryo-survival rate in the LBP group was also higher compared with
the GEYC group (P<0.05), as indicated in Fig. 1B-b. The normal morphology rates of
GEYC group was lower compared with the Fresh group (P<0.05), and
there was no obvious difference between the LBP group and the GEYC
group (Fig. 1B-c), but the abnormal
morphology rate at the middle part of sperms in the LBP group was
significantly lower compared with the GEYC group (P<0.05), the
abnormal morphology rate at the middle part of sperms in the GEYC
group was significantly higher compared with the Fresh group
(P<0.05) as presented in Fig.
1B-d. After cryo-survival, the sperms' progressive motility,
VCL, VSL and VAP, BCF in the LBP group were higher compared with
those in the GEYC group (P<0.05), the sperm progressive
motility, VCL, VSL, VAP, BCF and ALH in the GEYC group were lower
compared with those in the Fresh group (P<0.05) as presented in
Table I.
 | Table IAnalysis and detection of sperm
motility related indexes by CASA. |
Table I
Analysis and detection of sperm
motility related indexes by CASA.
| Group | PR (%) | VCL (µm/sec) | VSL (µm/sec) | VAP (µm/sec) | ALH (µm) | BCF
(times/sec) |
|---|
| Fresh |
22.0±5.2a |
38.5±3.5a |
19.7±3.8a |
22.7±2.2a |
4.4±1.1a |
13.3±1.5a |
| GEYC | 12.3±3.1 | 26.5±3.1 | 11.2±3.2 | 13.0±2.6 | 2.1±0.4 | 9.3±1.4 |
| LBP |
19.3±4.7a |
32.9±2.8a,b |
16.5±3.4a |
20.9±3.4a |
2.4±0.5b |
10.2±1.1a |
Sperm function
To assess the DFI, a minimum of 5,000 cells from
each sample were acquired and analyzed using fluorescence-assisted
cell sorting interfaced with a data analysis software. Sperm DFI
was assessed using a SCSA. The green fluorescence was that of
acridine orange dye binding to double-stranded DNA, while the red
fluorescence was that of acridine orange dye binding to
single-stranded DNA. As presented in Fig. 2A, following sperm cryopreservation,
the DFI rose and the DFI of the LBP group (27.3±5.1%) was lower
compared with the GEYC group (38.2±5.6%) but significantly higher
compared with the Fresh group (17.5±3.1%).
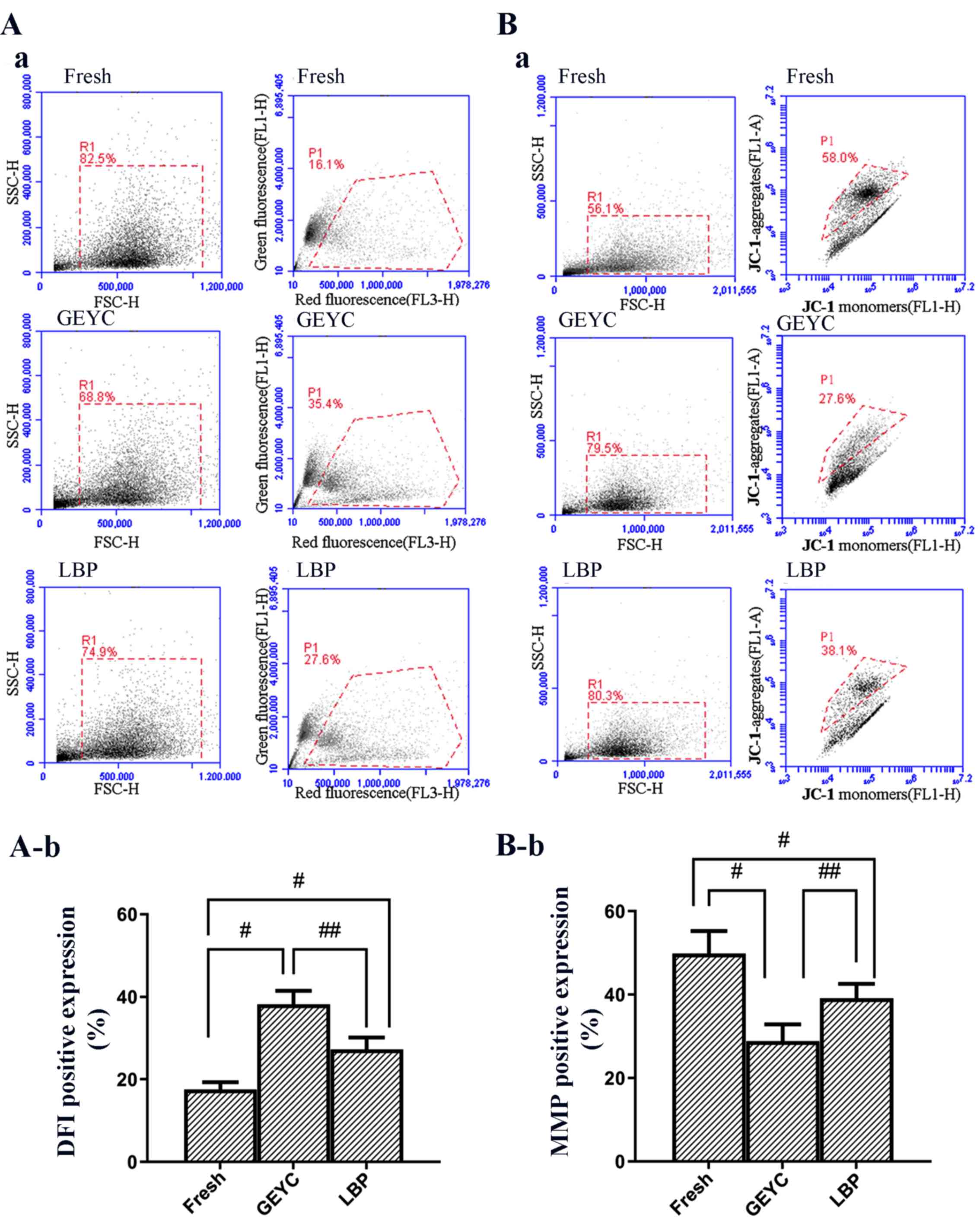 | Figure 2(A-a) FSC/SSC dot plot (52) (R1) is indicated in the left-hand
panel and the cytogram (red vs. green fluorescence) in the
right-hand panel, SSC-H reflects the fluorescence concentration per
unit area of cells. (A-b) Quantitative analysis of the
fluorescence-assisted cell sorting data (DFI). (B-a) The
scattergate and point diagram of representative sperm samples
stained by JC-1. In the right-hand panel, P1 contains
high-mitochondrial membrane potential sperm subsets. (B-b)
Quantitative analysis of the fluorescence-assisted cell sorting
data (MMP). (n=22). #P<0.05 vs. Fresh group;
##P<0.05 vs. GEYC group. GEYC, glycerin-yolk-citrate
cryopreservative; LBP, Lycium barbarum polysaccharides;
FSC-H, forward scatter-height; SSC, side scatter; MMP,
mitochondrial membrane potential; AR, acrosome reaction; DFI, DNA
fragmentation index; R1, region 1; FL, fluorescence. |
JC-1 dye molecules accumulate in the mitochondrial
matrix and produce red fluorescence; however, if the MMP is low,
the JC-1 dye is not able to accumulate in the mitochondrial matrix
and green fluorescence is then produced. As presented in Fig. 2B, the MMP of cryopreserved
spermatozoa was increased. The MMP of the LBP group (39.2±6.1%) was
higher compared with the GEYC group (28.8±7.0%) but significantly
lower compared with the Fresh group (49.8±9.5%).
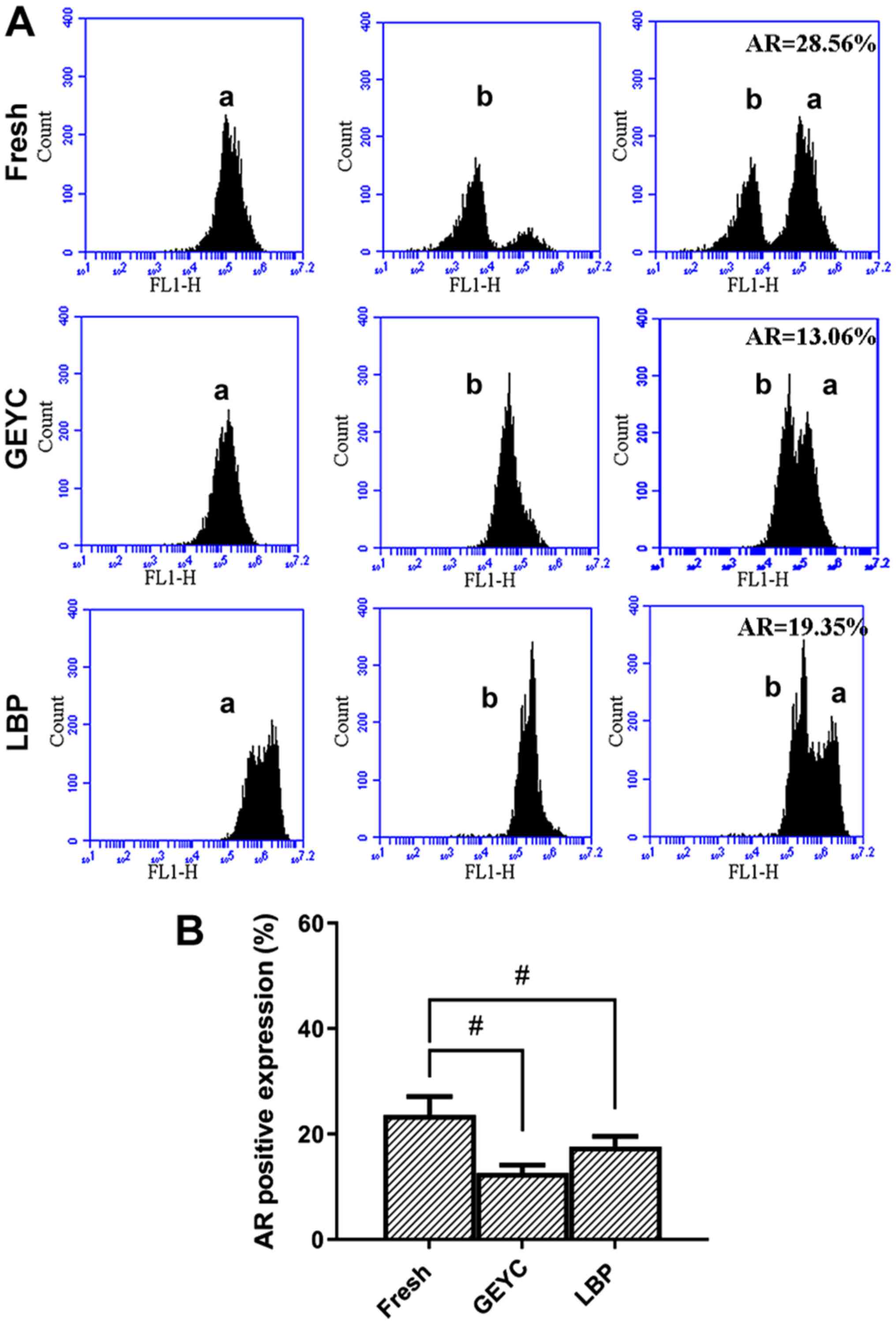
After the acrosome reaction was induced, the
relative fluorescence intensity was weaker compared with the
control, indicating that the fluorescence peak shifted to the left.
The AR of cryopreserved sperm was reduced and there was no
significant difference between the LBP group (17.7±3.3%) and the
GEYC group (14.7±2.5%); The AR of the GEYC group was lower than
that of the Fresh group (24.6±4.3%; Fig.
3A and B).
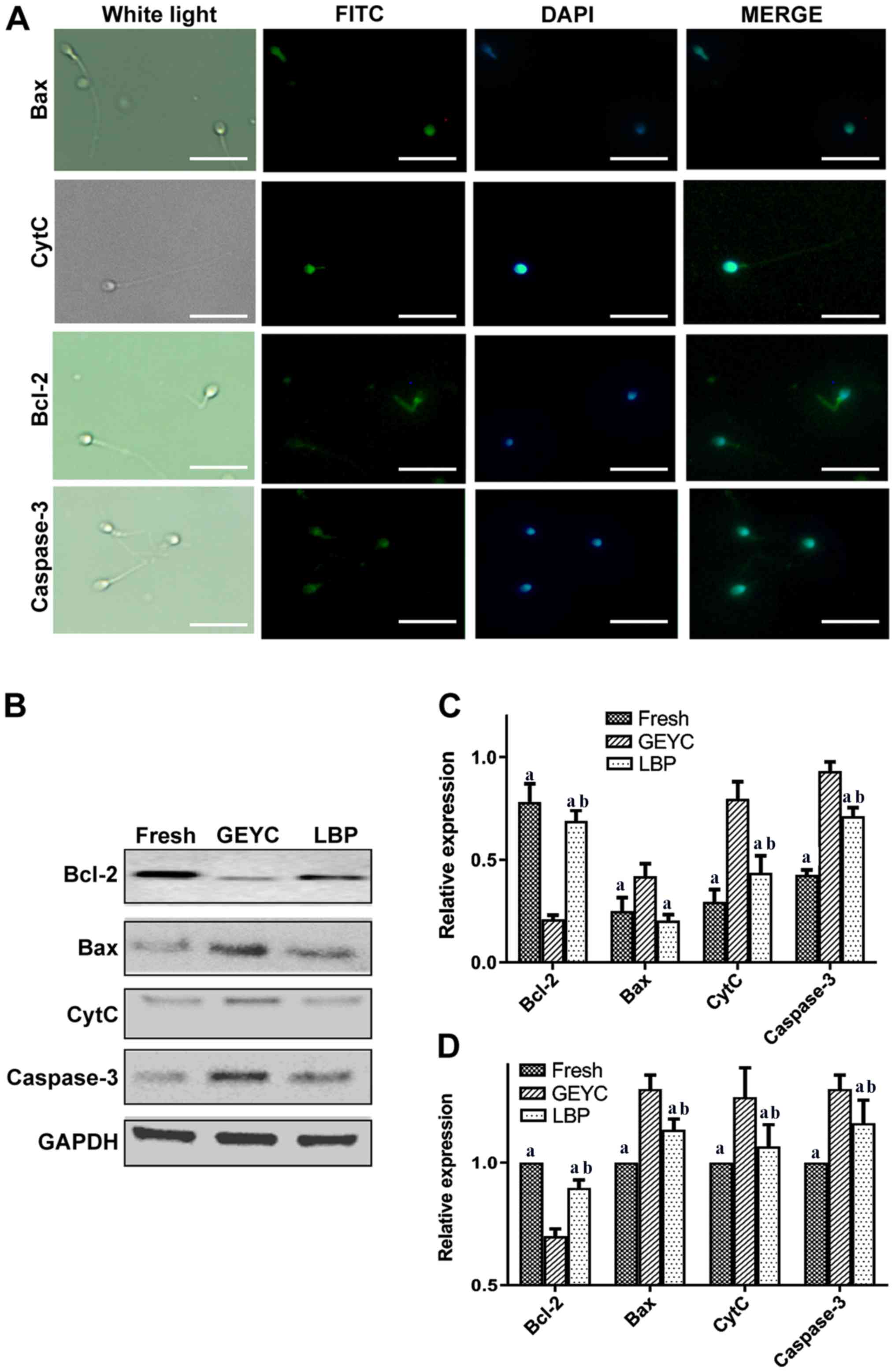
Apoptosis-associated proteins
The results of the immunofluorescence analysis
indicated that the expression of Bcl-2, Bax, CytC and caspase-3 was
concentrated in the head and middle part of the sperms (Fig. 4A). Western blot analysis suggested
that the LBP group had a higher content of Bcl-2 protein compared
with the GEYC group, while the expression levels of Bax, CytC and
caspase-3 protein were lower compared with the GEYC group. Bcl-2,
CytC, Caspase-3 in the LBP group and Fresh group differed
significantly, while the differences in Bax were not significant,
as presented in Fig. 4C. The results
of the reverse transcription PCR analysis indicated that, compared
with the GEYC group, Bax, CytC and caspase-3 were significantly
decreased in the LBP group. Bcl-2 in the LBP group was higher
compared with the GEYC group, Bcl-2, Bax, CytC, Caspase-3 in the
LBP group and Fresh group differed significantly, as presented in
Fig. 4D.
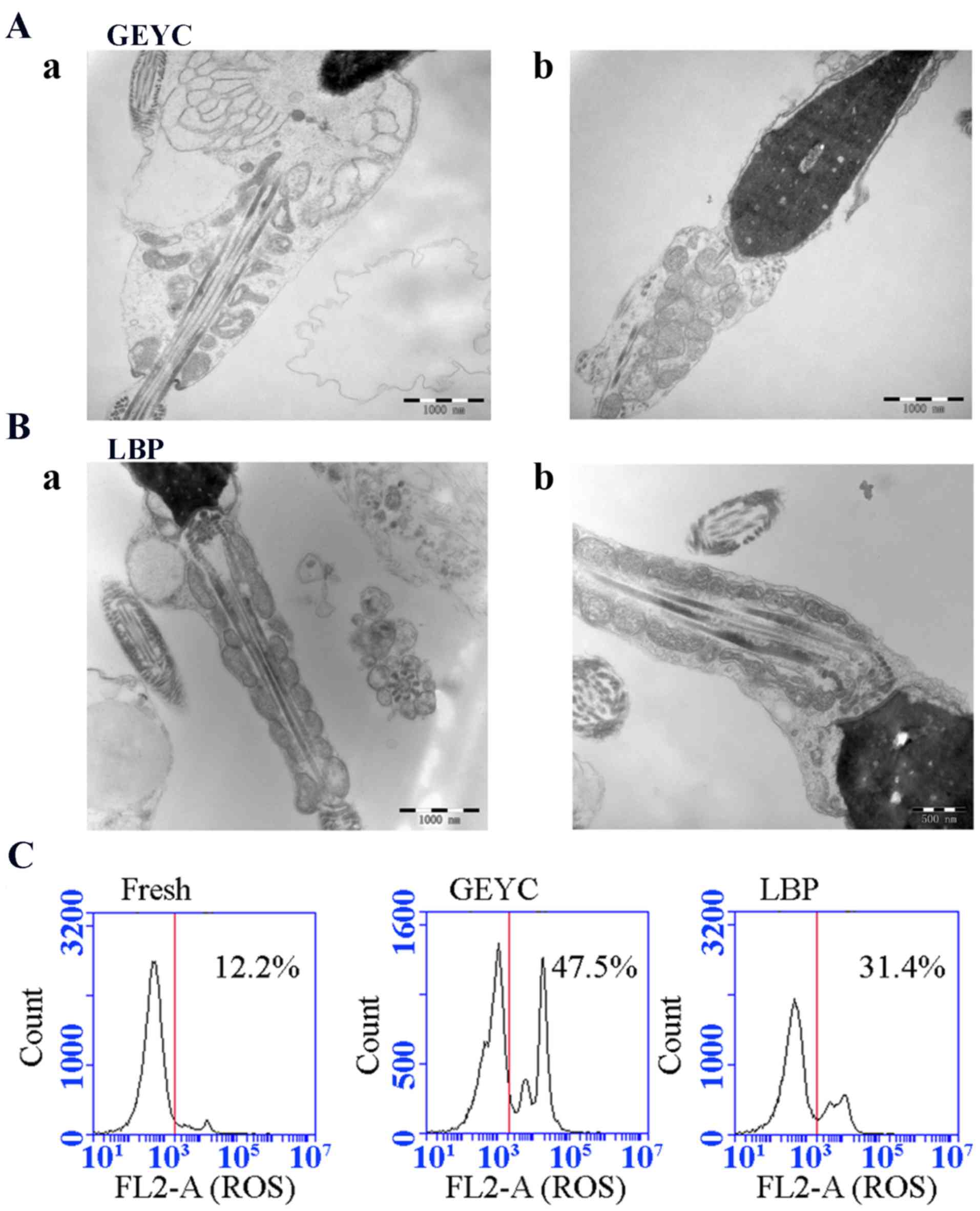
TEM and ROS detection results
TEM was used to detect the microstructure of sperm.
It was indicated that the mitochondrial structure in the middle
part of the sperm tail was severely damaged after freezing, the
mitochondrial cristae structure was disorganized and normal
mitochondrial cristae disappeared. By contrast, there was a
significant improvement in the LBP group. Specifically, the
mitochondria at the sperm neck were orderly arranged and abnormal
mitochondrial structures were significantly less frequent, as
presented in Fig. 5A-B.
The TBA colorimetric method was used for MDA
detection. The results suggested that the LBP group (18.0±3.6
nmol/ml) had a significantly lower sperm MDA content than the GEYC
group (23.9±4.1 nmol/ml), which was higher than the fresh group
(16.3±3.4 nmol/ml), but there was no significant difference between
the LBP group and the Fresh group. ROS detection based on flow
cytometry indicated that the rate of ROS-positive cells in the LBP
group (38.1±6.7%) was significantly lower compared with the GEYC
group (58.3±10.8%), and this rate in the LBP group increased as
compared with the Fresh group (17.4±5.2%; P<0.05; Table II; Fig.
5C).
| Table IIMDA and ROS in sperm. |
Table II
MDA and ROS in sperm.
| Group | MDA (nmol/ml) | ROS (%) |
|---|
| Fresh |
16.3±3.4a |
17.4±5.2a |
| GEYC | 23.9±4.1 | 58.3±10.8 |
| LBP |
18.0±3.6a |
38.1±6.7a,b |
Discussion
With the development of assisted reproductive
technology, the cryopreservation of germ cells is improving. The
selection of protective agents is progressing, and the
cryopreservation scheme is undergoing constant optimization and
innovation. The formation of ice crystals during freezing-thawing,
the production of massive ROS and the toxic effect of glycerol in
GEYC cryopreservative on cells all seriously affect the quality and
mobility of sperm after freezing (27-29).
Specifically, the ROS generated during the freeze-thawing process
may damage the mitochondrial membrane (30,31) and
reduce the MMP of sperm mitochondria (32), thereby leading to a sharp decline in
sperm motility. Sperm plasma membranes are vulnerable to oxidative
stress, as they are rich in polyunsaturated fatty acids (33,34).
Furthermore, oxidative damage may lead to sperm plasma membrane
injury, respiratory depression, intracellular enzyme loss, axoneme
protein damage and mitochondrial membrane injury (1,9). In
addition, the structure of sperm is damaged after freezing and
thawing, the integrity of the sperm plasma membrane is broken, the
sperm tail is injured, the mitochondrial structure and function are
impaired, ATP malfunctions occur, the sperm motility parameters
decrease significantly and sperm function is damaged. Due to this,
sperm are weakened and their survival impaired after
cryo-preservation, having a negative impact on its fertilization
ability (35-40).
The damage to sperm that is caused by excessive ROS
produced by freezing and thawing is reduced by the addition of
antioxidants, and thus, sperm motility may be improved. For
instance, Zheng et al (41)
reported that the addition of a certain concentration of VitE to
the sperm cryopreservative may reduce sperm damage that is caused
by the freeze-thawing process and partly improve the motility,
survival rate and DNA integrity of sperm. However, most previous
studies are concentrated on sperm viability and morphology after
freezing, rather than the impact on sperm microstructure (7-9,41).
There are few studies eluding to the role of antioxidants in sperm
freezing and thawing and indicating which pathways and molecules
are affected during this process (10-13,35).
LBP, which is extracted from a traditional Chinese
medicinal plant named Lycium barbarum, exhibits biological
effects including antioxidative effects, scavenging of oxygen free
radicals and the stabilization of the cell membrane (15). In a previous study, LBP was added at
different concentrations to conventional GEYC cryopreservative and
it was determined that LBP at 1,000 µg/ml had the most significant
effect in improving sperm motility after freezing, and for this
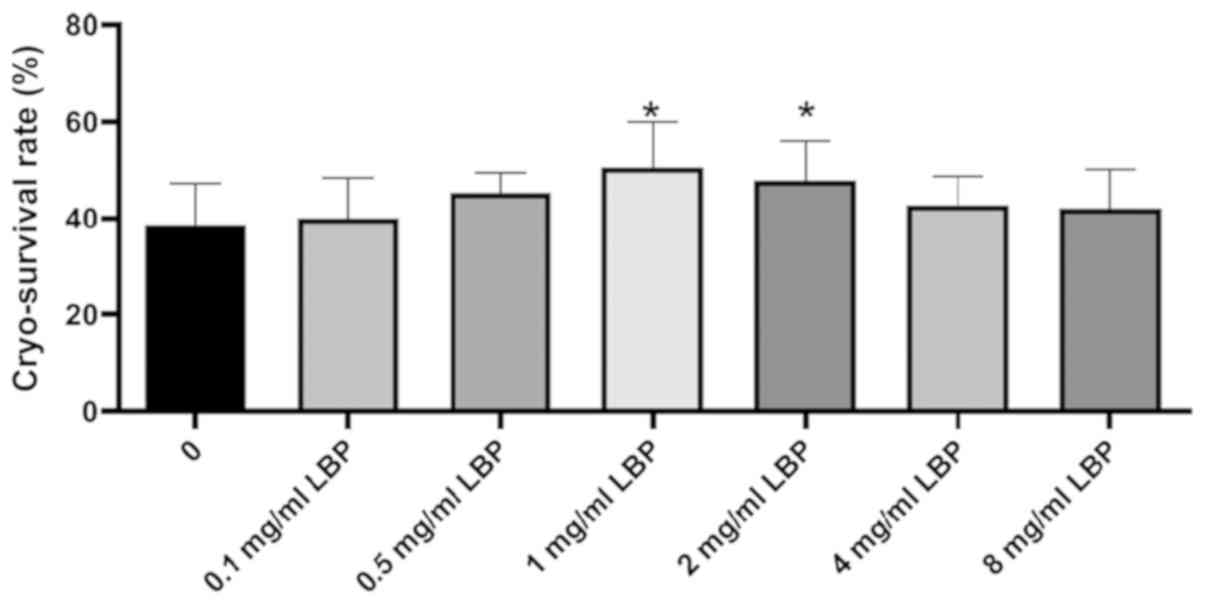
reason, the concentration of the added LBP in the present study was
1,000 µg/ml (Fig. 6). The results
indicated although there was no significant difference between the
LBP group and the GEYC group in terms of the normal sperm
morphology rate, the deformity rate of the sperm tail at the middle
part in the LBP group was significantly declined as compared with
that the GEYC group. TEM observation indicated that the
mitochondrial area in the middle part of the sperm tail was
obviously swollen, the normal mitochondrial cristae disappeared,
and the mitochondrial structure was disorganized. In the LBP group,
the ultrastructures of mitochondria and abnormal spermatozoa in the
neck were significantly improved.
The energy required for sperm motility is derived
from the ATP produced by mitochondria and the structure and
functional integrity of mitochondria after freezing is important
for sperm motility and quality (42). The MMP in the frozen group with LBP
was significantly higher compared with the GEYC group (P<0.05)
and the DFI of sperm declined after LBP was added, which may be
attributed to the biological effects of LBP to act as an
antioxidant, scavenge oxygen free radicals and stabilize the cell
membrane. In the present study, the AR detection result in the LBP
group was higher compared with the GEYC group but the difference
was not statistically significant, suggesting that LBP could
promote sperm acrosome reaction, but LBP at this concentration did
not have a significant promoting effect on every sample. This may
be related to the difference of each sperm sample, such as the
difference in the redox ability. If the sample size was expanded,
the current study may have gained more satisfactory results.
Furthermore, the addition of LBP reduced the contents of MDA and
ROS, which demonstrated that LBP exhibits an antioxidant capacity.
A large number of previous studies indicated that LBP may scavenge
oxygen free radicals, increase SOD levels in cells and withstand
oxidation (43-45).
In the present study, after LBP was added to the cryopreservative,
the sperm forward motility and movement parameters (VCL, VSL and
VAP) increased significantly, and the MMP and DFI were also
markedly improved, indicating that the antioxidant capacity of LBP
reduced the damage to sperm DNA and mitochondrial membrane caused
by ROS, thereby protecting mitochondria and DNA.
The mitochondrial signaling pathway is a common
apoptosis pathway that may be activated by a number of different
factors to trigger the interaction between Bcl-2 and Bax, causing
the latter to oligomerize and be inserted into the mitochondrial
membrane, leading to changes in the permeability of the
mitochondrial membrane and release of CytC. Subsequently, the
caspase cascade is initiated and the downstream caspase-3 is
activated, resulting in cell apoptosis (46). Hezavehei et al (47) indicated that freezing resulted in an
increase of caspase-3 and apoptosis of human sperm. Therefore, four
key genes, namely Bcl-2, Bax, CytC and caspase-3, were used as
indicators to observe the effect of LBP on sperm freezing and
thawing in the present study. The protein imprinting results
indicated that in the LBP group, Bcl-2 increased compared with the
GEYC group, while Bax, CytC and caspase-3 decreased, indicating
that LBP affected the key molecules in this pathway and inhibited
cell apoptosis. The detection results for RNA levels were
consistent with those for the protein levels. The biological
functions of LBP to exert antioxidant effects and scavenge oxygen
free radicals attenuate the effect of excessive ROS on the sperm
mitochondrial pathway in the freeze-thawing process, thereby
protecting the sperms' mitochondria and function. Similar to the
results of the present study, Dai et al (48) reported that the addition of LBP when
freezing goat spermatogonial stem cells led to the upregulation of
the expression of anti-apoptotic Bcl-2 and downregulation of Bax
protein, which significantly reduced apoptosis of the cells. Ma
et al (49) and Huang et
al (50) indicated that LBP my
reduce CytC and caspase-3 levels, thus significantly reducing
apoptosis of the studied cells. In addition, Li et al
(51) added LBP when freezing goat
semen and revealed that the integrity rate of the sperm plasma
membrane and the mitochondrial activity were significantly
increased.
In the sperm treatment process of the present study,
sperm was separated from semen using centrifugation after semen
liquefaction and then cultured in BWW medium with LBP for 20 min
prior to freezing, so that LBP was able to better interact with
sperm and obtain a better freezing effect. In this way, the damage
of non-sperm components to the structure and function of sperms in
the semen, as well as any interference with the experimental
results, were avoided.
Lycium barbarum is a traditional medicinal
plant in China, and research into its major functional component,
LBP, is still ongoing. Further study should be performed on the
protective mechanism of LBP in sperm cryopreservation. With the
continuous advance of various omics methods, it is possible to
explore whether LBP has an interactive molecular target and further
study its function, to develop a safer natural sperm
cryopreservative and ultimately to improve the preservation of
sperm.
Acknowledgements
Not applicable.
Funding
The current study was supported by the Natural
Science Foundation of Ningxia (grant no. NZ17140), the Key R and D
Plan Project of Ningxia Hui Autonomous Region (grant no.
2020BFH02002), Guangzhou Science and Technology Plan Project (grant
no. 201707010394) and the open project of National Health Committee
Key Laboratory of Male Reproduction and Genetics (grant no.
KF201709).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
HW and YT designed the study, BY and XZ performed
sperm function tests, TEM and sperm protein and RNA analyses; SJ
and YZ performed sperm motility tests and morphological tests. JT
and JW performed the statistical analysis of the data in this
study. HW and YT wrote and edited the paper. All authors read and
approved the final manuscript.
Ethics approval and consent to
participate
The present study was approved by the ethics
committee of the General Hospital of Ningxia Medical University
(approval no. 2019-402).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Shabani Nashtaei M, Nekoonam S, Naji M,
Bakhshalizadeh S and Amidi F: Cryoprotective effect of resveratrol
on DNA damage and crucial human sperm messenger RNAs, possibly
through 5' AMP-activated protein kinase activation. Cell Tissue
Bank. 19:87–95. 2018.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Li SJ, Su WD, Qiu LJ, et al: Protective
effect of resveratrol on human sperm freezing injury. Chin J
Androl. 24:499–503. 2018.
|
|
3
|
Nie ZY, Wu HF, Zhang N, et al: The effect
of male age on sperm oxidative stress and nuclear DNA damage. Chin
J Fam Planning. 20(30)2012.(In Chinese).
|
|
4
|
Ma CJ, Zhuang JM, Deng SM, et al: The
effect of docosahexaenoic acid added to glycerin yolk citrate
cryoprotectors on the kinematical parameters of human sperm before
and after cryopreservation. Chin Contemporary Med. 25:4–8. 2018.(In
Chinese).
|
|
5
|
Chen ZW, Fan LQ, Wen RQ, Lu WH, et al:
Assisted reproductive male technology. 1st edition. Beijing,
People's Health Press, 2016.
|
|
6
|
Li Y: Study on the cryopreservation effect
of Epimedium Polysaccharide on goat semen. Northwest Univ
Agriculture Forestry Sci Technol, 2019.
|
|
7
|
Ren F, Feng T, Dai G, Wang Y, Zhu H and Hu
J: Lycopene and alpha-lipoic acid improve semen antioxidant enzymes
activity and cashmere goat sperm function after cryopreservation.
Cryobiology. 84:27–3. 2018.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Losano JDA, Angrimani DSR, Rui BR, Bicudo
LC, Dalmazzo A, Silva BCS, Viana CHC, Mendes CM, Assumpção MEOA,
Barnabe VH and Nichi M: The addition of docosahexaenoic acid (DHA)
and antioxidants (glutathione peroxidase and superoxide dismutase)
in extenders to epididymal sperm cryopreservation in bulls. Zygote.
26:199–206. 2018.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Tian J, Zhang SH, Ma K, et al: The role of
Rho/rock signal pathway in sperm anti freezing injury. Chin J
Androl. 4:322–328. 2019.(In Chinese).
|
|
10
|
Shahrzad E, Zahiri S, Ghasemi F and
Jahromi HK: A study of effects of L-carnitine on morphology and
apoptosis in cryopreserved Sperm. Adv Environmental Biol.
7:2126–2134. 2013.
|
|
11
|
Yujie Z, Jing Y, Tailang Y, et al: The
effect of acetyl-L-carnitine on oxidative damage of human sperm
during cryopreservation. J Wuhan Univ (Med Ed). 35:426–431.
2014.
|
|
12
|
Yujie Z, et al: The protective effect of
acetyl-L-carnitine on acrosome integrity and ultrastructure of
human sperm before and after cryopreservation. J Med Mol Biol.
14:268–273. 2017.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Li L, et al: Protective effect of
mitoquinone, a mitochondrial targeted antioxidant, on oxidative
stress injury of human sperm during freezing thawing. Chin J
Androl. 22:205–211. 2016.
|
|
14
|
Berkovitz A, Fitoussi D, Zhakov D and
Breitbart H: Cryopreservation of human sperm in the presence of
Zn2+ increases the motility rate. J Obstetrics
Gynecological Investigations. 1:6–12. 2018.
|
|
15
|
Wu W, Xue SJ, Zhu LQ, et al: Study on the
protective effect of Lycium barbarum polysaccharide on the
injury of vascular endothelial cells induced by hydrogen peroxide
through antioxidation and anti apoptosis. Shi Zhen Guo Yi Guo Yao.
30:1047–1049. 2019.
|
|
16
|
Rui Z, Kang KA, Mei JP, Kim KC, Kim AD,
Chae S, Park JS, Youn UJ and Hyun JW: Cytoprotective effect of the
fruits of Lycium Chinense Miller against oxidative stress-induced
hepatotoxicity. J Ethnopharmacol. 130:299–306. 2010.
|
|
17
|
Yufang C, Yanrong W, Bin K, et al: Effect
of Lycium barbarum polysaccharide on cryopreservation of
fetal ovarian tissue. China Tissue Engineering Res. 16:7475–7479.
2012.
|
|
18
|
Lianghong M, Zhijun Q, Shengnan Y, et al:
Effect of Lycium barbarum polysaccharide on proliferation of
spermatogonial stem cells in vitro. China Tissue Engineering Res.
15:4277–4281. 2011.
|
|
19
|
Huan Z, Na T, Lihua W, Jianan P, Yi D and
Menghua C: Lycium barbarum polysaccharides alleviate kidney
injury and oxidative stress in unilateral ureteral obstruction
rats. Chin J Nephrol. 34:377–384. 2018.
|
|
20
|
Vittoria MV, Pasciu V, Gadau SD, Baralla
E, Serra E, Palomba D and Demontis MP: Possible antioxidant effect
of Lycium barbarum polysaccharides on hepatic
cadmium-induced oxidative stress in rats. Environmental Sci
Pollution Res. 24:2946–2955. 2017.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Qingqing H, Bei Y, Tao Y, Fang M and Juan
W: The protective effect of Lycium barbarum polysaccharide
on the washing process of spermatozoa after freezing. Ningxia Med
J. 41:1073–1075. 2019.
|
|
22
|
Zhang WD, Zhang Z, Jia LT, Zhang LL, Fu T,
Li YS, Wang P, Sun L, Shi Y and Zhang HZ: Oxygen free radicals and
mitochondrial signaling in oligospermia and asthenospermia. Mol Med
Rep. 10:1875–1880. 2014.PubMed/NCBI View Article : Google Scholar
|
|
23
|
World Health Organization, Department of
Reproductive Health and Research: WHO laboratory manual for the
Examination and processing of human semen, Fifth edition.
Switzerland, WHO Press, 2010.
|
|
24
|
Lu JC, Jing J, Chen L, Ge YF, Feng RX,
Liang YJ and Yao B: Analysis of human sperm DNA fragmentation index
(DFI) related factors: A report of 1010 subfertile men in China.
Reproductive Biol Endocrinol. 16(23)2018.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Guowei Z, Zhi W, Xi L, Zou P, Yang H, Chen
Q, Zhon N, Sun L, Gao J, Zhon Z, et al: Mitochondrial biomarkers
reflect semen quality: Results from the MARCHS study in Chongqing,
China. PLoS One. 11(e0168823)2016.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Zhao ZH, Zhang TT and Wang H: Effect of
breviscapine on apoptosis of cortical neurons induced by
hypoxia/reoxygenation. Practical Med Clin. 1:9–13. 2020.
|
|
27
|
Ma CJ, Jiang F, Zhuang JM, et al: The
effect of glycerol maohuang sodium citrate cryoprotectors on human
sperm motility parameters without cryopreservation. Chin J Androl.
24:37–39. 2010.
|
|
28
|
Zhao K, Chen W, Xu AM, et al: Preliminary
study on the protective effect of isocitrate on sperm freezing and
thawing. J Nanjing Med Univ Nat Sci Ed. 37:165–168. 2017.
|
|
29
|
Cheng S, Yan JQ, Zhu JQ, et al: DNA damage
of Pseudosciaena crocea sperm cryopreserved with glycerol as
antifreeze. China J Animal Husbandry. 49:34–36. 2013.
|
|
30
|
Dong QY, Shi H and Li JY: Research on
cryopreservation injury and protection of human sperm. Chin J Fam
Planning. 27:1259–1263. 2019.
|
|
31
|
Fu LL, Zhang LY, et al: The effect of
levocarnitine on the motility parameters and mitochondrial function
of frozen sperm. Chin J Androl. 24:1059–1063. 2018.
|
|
32
|
Gürler H, Molama E, Heppelmann M, Calisici
O, Leiding C, Kastelic JP and Bollwein H: Effects of
cryopreservation on sperm viability, synthesis of reactive oxygen
species, and DNA damage of bovine sperm. Theriogenology.
86:562–571. 2016.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Baumber J, Ball BA, Linfor JJ and Meyers
SA: Reactive oxygen species and cryopreservation promote DNA
fragmentation in equine spermatozoa. J Androl. 24:621–628.
2003.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Johnson AE, Freeman EW, Wildt DE and
Songsasen N: Spermatozoa from the maned wolf (Chrysocyon
brachyurus) display typical canid hyper-sensitivity to osmotic and
freezing-induced injury, but respond favorably to dimethyl
sulfoxide. Cryobiology. 68:361–370. 2014.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Zribi N, Chakroun NF, Ben Abdallah F,
Elleuch H, Sellami A, Gargouri J, Rebai T, Fakhfakh F and Keskes
LA: Effect of freezing-thawing process and quercetin on human sperm
survival and DNA integrity. Cryobiology. 65:326–331.
2012.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Chen SJ, Allam JP, Duan YG and Haidl G:
Influence of reactive oxygen species on human sperm functions and
fertilizing capacity including therapeutical approaches. Arch
Gynecol Obstet. 288:191–199. 2013.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Ribas-Maynou J, García-Peiró A,
Fernández-Encinas A, Abad C, Amengual MJ, Prada E, Navarro J and
Benet J: Comprehensive analysis of sperm DNA fragmentation by five
different assays: TUNEL assay, SCSA, SCD test and alkaline and
neutral Comet assay. Andrology. 1:715–722. 2013.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Huang CC, Lin DP, Tsao HM, Cheng TC, Liu
CH and Lee MS: Sperm DNA fragmentation negatively correlates with
velocity and fertilization rates but might not affect pregnancy
rates. Fertil Steril. 84:130–140. 2005.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Mahfouz R, Sharma R, Thiyagarajan A, Kale
V, Gupta S, Sabanegh E and Agarwal A: Semen characteristics and
sperm DNA fragmentation in infertile men with low and high levels
of seminal reactive oxygen species. Fertil Steril. 94:2141–2146.
2010.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Tran M, Uriondo H, Nodar F and Sedó CA:
Cryopreservation promotes sperm DNA damage through oxidative stress
(38N). Obstetr Gynecol. 131 (Suppl)(162S)2018.
|
|
41
|
Zheng JJ, Chen X, Zhang LY, et al:
Protective effects of vitamin E and B12 on viability and DNA of
frozen sperm. J Reproductive Med. 27:264–268. 2016.
|
|
42
|
O'Connell M, Mcclure N and Lewis SEM: The
effect of cryopreservation on sperm morphology, motility and
mitochondrial function. Hum Reprod. 17:704–709. 2002.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Liu L, Lao W, Ji QS, Yang ZH, Yu GC and
Zhong JX: Lycium barbarum polysaccharides protected human
retinal pigment epithelial cells against oxidative stress-induced
apoptosis. Int J Ophthalmol. 8:11–16. 2015.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Zhang WJ, Yang H, Zhu L, Luo Y, Nie L and
Li G: Role of EGFR/ErbB2 and PI3K/AKT/e-NOS in Lycium
barbarum polysaccharides ameliorating endothelial dysfunction
induced by oxidative stress. Am J Chin Med. 47:1523–1539.
2019.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Li XM, Ma YL and Liu XJ: Effect of the
Lycium barbarum polysaccharides on age-related oxidative
stress in aged mice. J Ethnopharmacol. 111:504–511. 2007.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Taheri Moghadam M, Asadi Fard Y, Saki G
and Nikbakht R: Effect of vitamin D on apoptotic marker, reactive
oxygen species and human sperm parameters during the process of
cryopreservation. Iran J Basic Med Sci. 22:1036–1043.
2019.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Hezavehei M, Mohseni Kouchesfahani H,
Shahverdi AH, Sharafi M, Hosseini Salekdeh GH and Eftekhari-Yazdi
P: Induction of Sublethal oxidative stress on human sperm before
cryopreservation: A time-dependent response in post-thawed sperm
parameters. Cell J. 20:537–543. 2019.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Dai GC: Effects of Lycium barbarum
polysaccharide on apoptosis inhibition and cryopreservation of Goat
Spermatogonial stem cells. Northwest Agricultural Forestry Univ,
2019.
|
|
49
|
Ma XF: Effects of Lycium barbarum
polysaccharide on apop-tosis, gene expression and delayed rectifier
potassium current of retinal ganglion cells induced by high
glucose. J Hainan Med Coll. 23:2017.
|
|
50
|
Huang H, P H, W Y, et al: Inhibitory
effect of Lycium barbarum polysaccharide on apoptosis of
bone marrow monocytes induced by ionizing radiation. Environmental
Occupational Med. 35:67–71. 2018.
|
|
51
|
Li FZ, Wang YH, Wen F, et al: Effects of
Laminaria polysaccharide and Lycium barbarum polysaccharide on
cryopreservation of goat semen. Environmental Occupational Med.
39:48–52. 2018.
|
|
52
|
Muratori M, Tarozzi N, Carpentiero F,
Danti S, Perrone FM, Cambi M, Casini A, Azzari C, Boni L, Maggi M,
et al: Sperm selection with density gradient centrifugation and
swim up: Effect on DNA fragmentation in viable spermatozoa. Sci
Rep. 9(7492)2019.PubMed/NCBI View Article : Google Scholar
|