Introduction
Pregnancy-induced hypertension is a common disease
observed during the gestational period. If not treated in time,
pregnancy-induced hypertension may cause significant harm to the
mother and fetus (1,2). Recent clinical studies have shown that
maternal mortality as a result of pregnancy-induced hypertension to
be 4.2-10 million worldwide as of 2017, accounting for ~9% of all
maternal deaths and 2.2% of perinatal child mortality (3). Thus, this disease poses a serious
threat to maternal and child health, and represents one of the main
causes for the death of pregnant women and neonates (4). The clinical symptoms of
pregnancy-induced hypertension mainly include transient
hypertension and proteinuria in pregnant women, which normally
disappear following delivery (5). At
present, pregnancy-induced hypertension is generally divided into
five types: Gestational hypertension, pre-eclampsia (mild and
severe), eclampsia, chronic hypertension complicated by
pre-eclampsia and chronic hypertension combined with pregnancy
(6). Pregnant women with severe
pregnancy-induced hypertension may suffer from hemolysis,
thrombocytopenia, liver and kidney dysfunction, pulmonary edema and
visual disturbances (5). Risk
factors of the disease include obesity, pre-pregnancy hypertension,
diabetes and old age (7). The causes
for pregnancy-induced hypertension remain to be elucidated;
however, they may be associated with changes in the immune system
such as histocompatibility antigen-associated immunological
abnormalities (8). In addition,
genetic susceptibility may also be involved in the pathogenesis of
this disease (9,10).
Trophoblast cells are one of the components of the
maternal placental architecture, involved in the regulation of
placental microenvironment remodeling, implantation of embryos and
normal fetal development (11).
Previous studies have shown that trophectoderm cells can
differentiate into two types of trophoblast cells at the early
stages of blastocyst implantation, namely cytotrophoblast cells and
syncytiotrophoblast cells (12,13).
Cytotrophoblast cells can fuse with syncytiotrophoblast cells,
which differentiate into extravillous trophoblasts (EVTs). Some
EVTs infiltrate the deeper layers of the endometrium, and are known
as interstitial trophoblast cells (14), whilst others invade the maternal
uterine spiral artery, and are called endovascular trophoblast
cells (15). Trophoblast cells
exhibit similar migratory capacities to tumor cells, which are
closely related in biological function (16). Studies have shown that EVTs migrate
from the placental villi and invade the endometrium and maternal
spiral arteries, where they participate in uterine artery
revascularization and regulate placental blood flow and immune
responses (17). Downregulation of
the invasive ability of trophoblast cells can result in defects in
uterine spiral artery remodeling and placental insufficiency,
leading to pregnancy-induced hypertension, eclampsia and
miscarriage (18). Therefore, it is
of great clinical significance to study changes in trophoblast cell
invasion and the associated mechanism in the pathogenesis of
pregnancy-induced hypertension.
Tight junction proteins are important in the
maintenance of cell-to-cell connections, which serve important
roles in cell polarity and the formation of cellular barriers such
as the intestinal epithelial barrier (19,20). In
particular, claudins (CLDNs) are members of the tight junction
protein family that serve important roles in the formation of tight
junctions. In total, 24 CLDNs have been identified (21). In recent years, studies have
demonstrated that CLDN3 is abnormally expressed in a number of
tumor tissues, including hepatocellular carcinoma and breast cancer
(22,23) and closely associated with the
invasion and metastasis of tumor cells (24,25).
Notably, when tumor cells metastasize, the tight junctions between
cells must be destroyed, and CLDN3 is an important component of
these junctions (26). During
trophoblast cell invasion, the breaking of tight junctions is also
an important prerequisite for detachment (27). However, the function of CLDN3 in this
process remain unclear. Therefore, in the present study, the
regulatory role of CLDN3 in the invasive abilities of trophoblast
cells was investigated, on tissue and cellular levels.
Materials and methods
Study subjects and sample
collection
A total of 51 pregnant women with hypertension,
including 25 patients diagnosed with pregnancy-induced
hypertension, 11 patients with mild pre-eclampsia and 15 patients
with severe pre-eclampsia, and 30 normal pregnant women were
included in this study, all of whom were admitted to Laiwu Maternal
and Child Health Hospital (Laiwu, China) for delivery from December
2016 to December 2017. In these patients, the pregnancy
hypertension was defined as: i) BP ≥140/90 mmHg during pregnancy,
which returned to normal within 12 weeks after delivery; ii) urine
protein (-); and iii) cases that may be associated with upper
abdominal discomfort or thrombocytopenia. Mild pre-eclampsia was
defined as: i) BP ≥140/90 mmHg appeared during pregnancy, which
returned to normal within 12 weeks after delivery; ii) urine
protein (-); and iii) cases that may be associated with upper
abdominal discomfort or thrombocytopenia. Severe pre-eclampsia was
defined as maternal convulsions that cannot be explained by other
reasons. None of the pregnant women had previous pregnancies or
suffered from hypertension, diabetes or underlying diseases,
including liver, kidney and autoimmune diseases, prior to the
present pregnancy. All these patients received cesarean section due
to pregnancy-induced hypertension, abnormal fetal position or other
social factors. There were no significant differences in age,
gestational period or neonatal weight between the pregnancy-induced
hypertension and control group (mean age of patients with
pregnancy-induced hypertension, 29.5±1.08 years; mean gestational
period, 38±2.08 weeks; mean neonatal weight, 7.6±2.3 kg. Mean age
of patients with mild-pre-eclampsia, 33.1±1.08 years; gestational
period, 39±3.08 weeks; neonatal weight, 7.1±1.3 kg. Mean age of
patients with severe pre-eclampsia, 31.45±1.08 years; gestational
period, 40±2.60 weeks; neonatal weight, 7.7±1.7 kg). Prior written
informed consent was obtained from every patient and the study was
approved by the ethics review board of Laiwu Maternal and Child
Health Hospital. In total 10 ml peripheral blood sample was
obtained from each subject and the collection of placental tissues
was performed 10 min after the placenta was delivered, with a 2x2x2
cm sample of tissue dissected from the central area (avoiding
infarction and calcification) of the maternal placenta. The tissue
was washed with saline immediately after collection, and then
stored at -80˚C.
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was extracted from the placental tissue
and peripheral blood samples using TRIzol® (Thermo
Fisher Scientific, Inc.) according to the manufacturer's protocol.
RNA concentrations were quantified using the NanoDrop method.
Reverse transcription was performed with 0.5 µg RNA to obtain cDNA
using BeyoRT™ cDNA First Chain Synthesis kit (cat. no. D7166;
Beyotime Institute of Biotechnology). Subsequent qPCR was performed
using BeyoFast™ SYBR Green qPCR Mix (cat. no. 7260; Beyotime
Institute of Biotechnology) in a StepOnePlus™ Real-Time PCR
instrument. The following primer sequences were used: CLDN3
forward, 5'-GCCACCAAGGTCGTCTACTC-3' and reverse,
5'-CCTGCGTCTGTCCCTTAGAC-3' and GAPDH forward,
5'-CGGAGTCAACGGATTTGGTCGTAT-3' and reverse,
5'-AGCCTTCTCCATGGTGGTGAAGAC-3'. The total 20 µl PCR mixture
consisted of 10 µl RT-qPCR-Mix, 0.5 µl each primer, 2 µl cDNA and 7
µl ddH2O. The thermocycling conditions were as follows:
Initial denaturation at 95˚C for 10 min; followed by 40 cycles of
95˚C for 1 min and 60˚C for 1 min. Target gene expression levels
were calculated using the 2-ΔΔCq method (28). GAPDH was used as internal
reference.
Human trophoblast cell culture
Normal human trophoblast HTR8/SVneo cells were
purchased from American Type Culture Collection. Cells were
cultured using RPMI-1640 medium (Gibco; Thermo Fisher Scientific,
Inc.) containing 10% fetal bovine serum (FBS; Gibco; Thermo Fisher
Scientific, Inc.) and supplemented with 100 U/ml penicillin and 100
U/ml streptomycin, in a humidified atmosphere at 37˚C containing 5%
CO2. When cell confluence reached 90%, the cells were
passaged using 0.5% trypsin.
Construction and transfection of
lentivirus vector for CLDN3
The lentiviral Lv-GFP-CLDN3 vector was constructed
by Hanbio Biotechnology Co., Ltd., with the titer of
1x108 pfu. HTR8/SVneo cells were first seeded into
24-well plates at a density of 1x105 cells/well and
cultured with RPMI-1640 medium containing 10% FBS. When 70%
confluence was achieved, the HTR8/SVneo cells were transfected with
either empty vector or Lv-GFP-CLDN3 at a multiplicity of
infectionof 20. The medium was changed to fresh RPMI-1640 medium
containing 10% FBS after 6 h, and the cells were cultured at 37˚C
under 5% CO2 for a further 48 h.
Cell counting kit (CCK)-8 assay
CCK-8 assay (Beyotime Institute of Biotechnology)
was performed to measure the viability of the HTR8/SVneo cells.
Briefly, at 48 h after transfection, the cells were seeded into
96-well plates at 2x103 cells/well. A total of 200 µl
RPMI-1640 complete medium containing 10% FBS and 100 U/ml
penicillin and streptomycin was added and the cells were then
incubated at 37˚C under 5% CO2. Following cell adhesion,
at 0, 24, 48 and 72 h, the cells were incubated with 20 µl CCK-8
reaction solution for 30 min. The optical density values at 490 nm
were then obtained for each well using a microplate reader, which
were used to produce a cell viability curve.
Flow cytometry
Flow cytometry was performed to measure HTR8/SVneo
cell proliferation and apoptosis. For cell proliferation, the cells
were collected and washed with PBS. Following fixation with 4%
formaldehyde for 10 min at room temperature, the cells were
centrifuged at 1,000 x g for 5 min at room temperature. The cells
were treated with 0.5% Triton X-100 at room temperature for 5 min
and then incubated with BD Cytofix/Cytoperm™ Plus reagent (cat. no.
555028; BD Biosciences) at room temperature in the dark for 15 min.
Fluorescence was detected and the percentage of Ki-67 calculated
from it using the BD FACSVerse™ flow cytometer (BD Biosciences) and
the FlowJo™ VX10 software (FlowJo LLC).
For apoptosis detection, 48 h after transfection,
1x106 HTR8/SVneo cells were collected and cultured with
a shaker for 2 h at a speed of 200 rpm. After washing twice with
ice-cold PBS, the cells were labeled using the BD Pharmingen™ FITC
Annexin V Apoptosis Detection Kit I (cat. no. 556547; BD
Biosciences), according to the manufacturer's protocol. Flow
cytometry was conducted to detect the fluorescence, from which
apoptosis rate was calculated.
For Ki-67 detection, the cells were collected and
treated with BD Cytofix/Cytoperm™ Plus reagent (cat. no. 555028; BD
Biosciences) according to the manufacturer's protocols. The cells
were then incubated with PE-Cy™7-conjugatedmouse anti-Ki-67
antibody (cat. no. 561283; BD Biosciences) in the dark at room
temperature for 15 min, followed by detection of the fluorescence
with flow cytometry.
Transwell chamber assay
The infiltration ability of the cells was detected
using Transwell chamber assays. Matrigel® solution was
diluted in serum-free RPMI-1640 medium (v:v, 1:3), evenly smeared
onto the upper chamber and incubated at 37˚C for 60 min. The
HTR8/SVneo cells were then seeded onto the upper chamber at a
density of 1x105 cells/well and cultured in 200 µl
serum-free RPMI-1640 medium while 500 µl RPMI-1640 medium
containing 20% FBS was present in the lower chamber. After 24 h,
the cells were fixed with 4% formaldehyde at room temperature for
10 min and subjected to Giemsa staining at room temperature for 2
min. Following rinsing for 2 min, the cells were air-dried.
HTR8/SVneo cells that infiltrated to the lower chamber were
observed and counted under a light microscope, from a total of five
random fields of view under high magnification (x200). For the
measurements of cell migration, identical procedures were performed
as described aforementioned using Transwell chambers that were not
coated with Matrigel.
Confocal laser scanning microscopy
(CLSM)
The cytoskeleton was imaged using CLSM. After
transfection, when 80% confluence was reached, HTR8/SVneo cells
were fixed with 4% formaldehyde at room temperature for 10 min.
After washing with PBS, the cells were permeabilized using 0.5%
Triton X-100 at room temperature for 5 min and incubated with 200
µl 100 nmol/l Rhodamine Phalloidin at room temperature in the dark
for 30 min. The cytoskeleton was then observed using a confocal
microscope (no. of fields taken, 5; magnification, x400; model SP8;
Leica Microsystems GmbH).
Western blot analysis
At 48 h after transfection, the cells were lysed
using RIPA buffer (Beyotime Institute of Biotechnology)
supplemented with PMSF (100 mM) on ice. The protein concentration
was determined using Bicinchoninic Acid assay (Beyotime Institute
of Biotechnology). Then, 20 µg sample was separated by 10%
SDS-PAGE, and then transferred onto PVDF membranes. After blocking
with 50 g/l fat-free milk diluted in 0.1% TBST at room temperature
for 1 h, the membranes were incubated with rabbit anti-human
anti-CLDN3 (1:1,000 dilution; cat. no. 83609), rabbit anti-human
anti-matrix metalloprotease (MMP)-2 (1:1,000 dilution; cat no.
40994), rabbit anti-human anti-MMP-9 (1:1,000 dilution; cat no.
13667), rabbit anti-human anti-ERK1/2 (1:1,000 dilution; cat no.
5013), mouse anti-human anti-p-ERK1/2 (1:1,000 dilution; cat. no.
9106) or rabbit anti-human anti-GAPDH (1:5,000 dilution; cat no.
5174) primary antibodies (all from Cell Signaling Technology, Inc.)
at 4˚C overnight. After washing with TBST, the membranes were
incubated with horseradish peroxidase-conjugated goat anti-rabbit
(cat. no. A0208) or goat anti-mouse (cat. no. A0216) secondary
antibody (both 1:4,000 dilution; Beyotime Institute of
Biotechnology) at room temperature for 1 h. Protein bands were
visualized using the ECL method (Pierce; Thermo Fisher Scientific,
Inc.) and analyzed using the Quantity one V4.6.7 software (Bio-Rad
Laboratories, Inc.).
Statistical analysis
Data are presented as the mean ± SD. SPSS 20.0
software (IBM Corp.) was used for statistical analysis. Pairwise
comparisons were performed using the Student's t-test, whereas
one-way ANOVA followed by Dunnett's test was used for multiple
group comparisons. P<0.05 was considered to indicate a
statistically significant difference.
Results
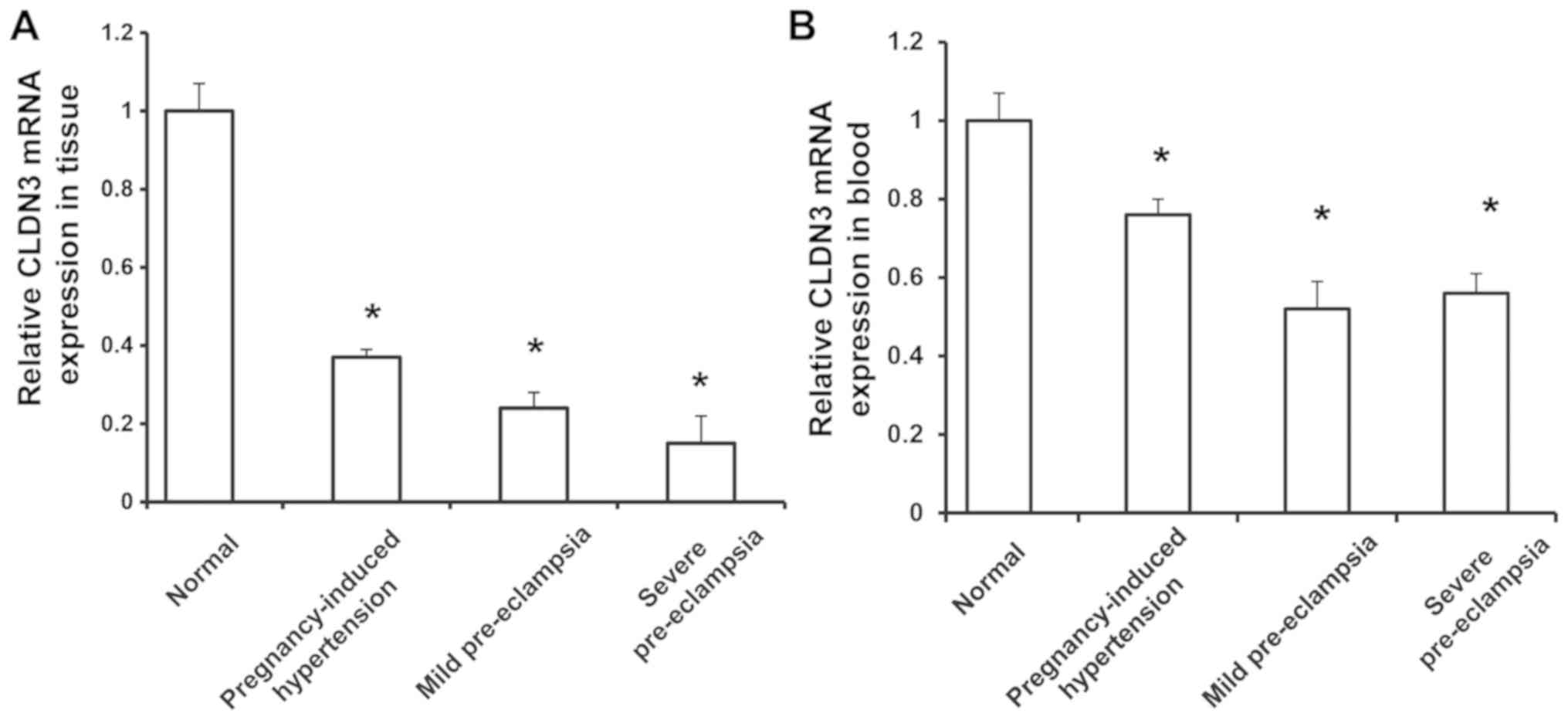
Expression of CLDN3 in placental
tissues from patients with pregnancy-induced hypertension
To investigate the expression of CLDN3 in the
placenta of the pregnant patients with and without hypertension,
RT-qPCR was performed. Compared with the healthy control group, the
expression levels of CLDN3 mRNA in the placental tissue were
significantly reduced in the pregnant patients with hypertension
(P<0.01; Fig. 1A). In addition,
relative CLDN3 mRNA expression levels in the placental tissues from
the severe pre-eclampsia, mild pre-eclampsia and pregnancy-induced
hypertension groups were significantly reduced compared with the
healthy control group (P<0.05; Fig.
1A). In the peripheral blood, compared with the control group,
the relative CLDN3 mRNA expression levels in the pregnancy-induced
hypertension, mild pre-eclampsia and severe pre-eclampsia groups
were significantly lower (P<0.05; Fig. 1B). These results suggest that at the
onset of pregnancy-induced hypertension, CLDN3 expression was
reduced in placental tissues, and may be associated with disease
pathogenesis.
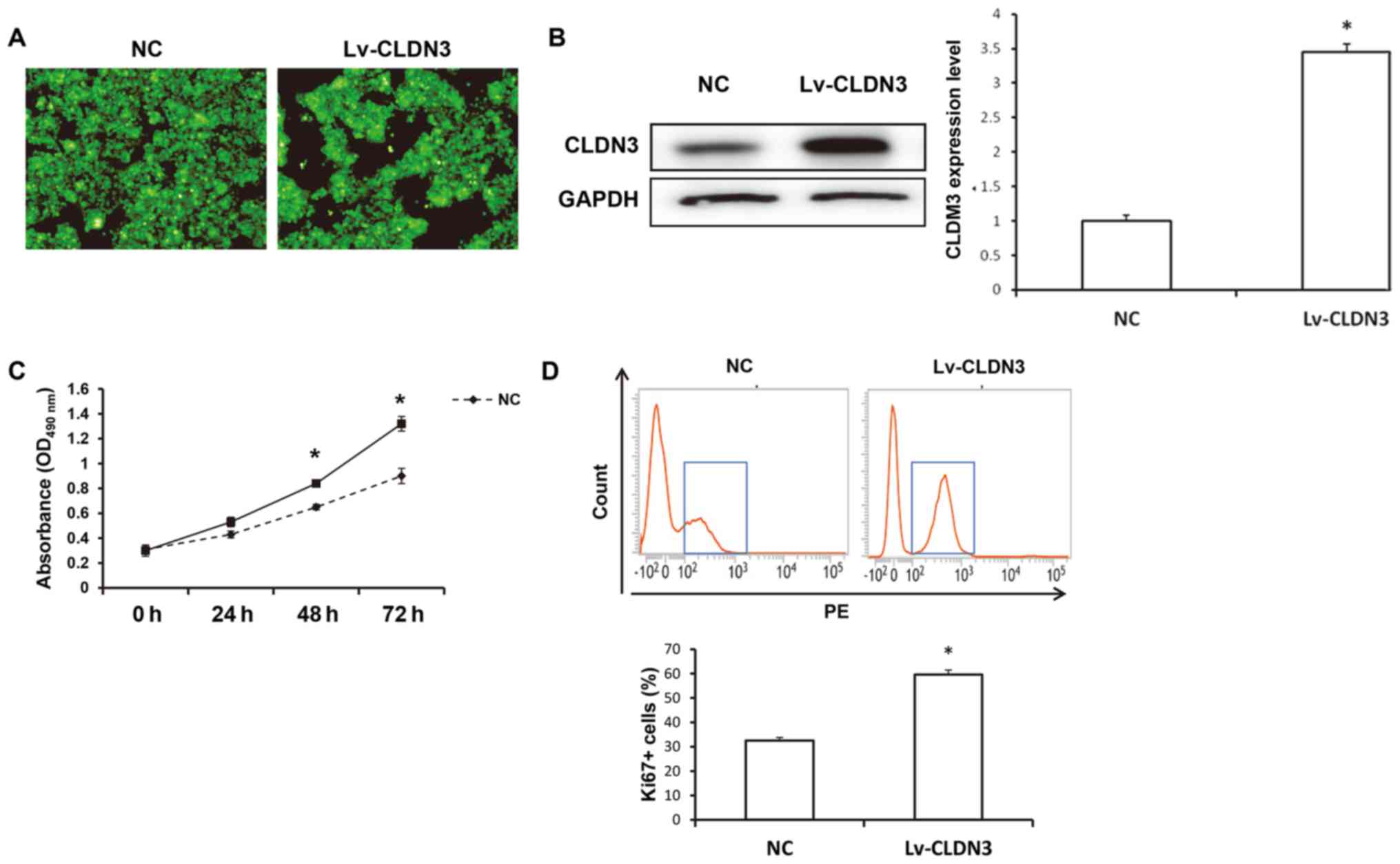
CLDN3 overexpression enhances of
HTR8/SVneo cell viability
The effects of CLDN3 on the viability of HTR8/SVneo
cells were next investigated. At 48 h after transfection with
Lv-GFP-CLDN3, GFP fluorescence was distributed evenly in the
cytoplasm of the HTR8/SVneo cells (Fig.
2A). Western blot analysis confirmed that CLDN3 expression was
elevated in HTR8/SVneo cells after transfection with Lv-GFP-CLDN3
compared with those in the empty vector-transfected control
(P<0.05; Fig. 2B). The viability
of the transfected cells was measured using a CCK-8 assay. Compared
with the control group, CLDN3 overexpression significantly
increased HTR8/SVneo cell viability at 24, 48 and 72 h after
transfection (P<0.05; Fig. 2C).
Consistent results were observed using flow cytometry, which
indicated that the overexpression of CLDN3 significantly
upregulated Ki-67 expression in HTR8/SVneo cells (P<0.05;
Fig. 2D). These results indicate
that CLDN3 overexpression significantly promoted HTR8/SVneo cell
proliferation.
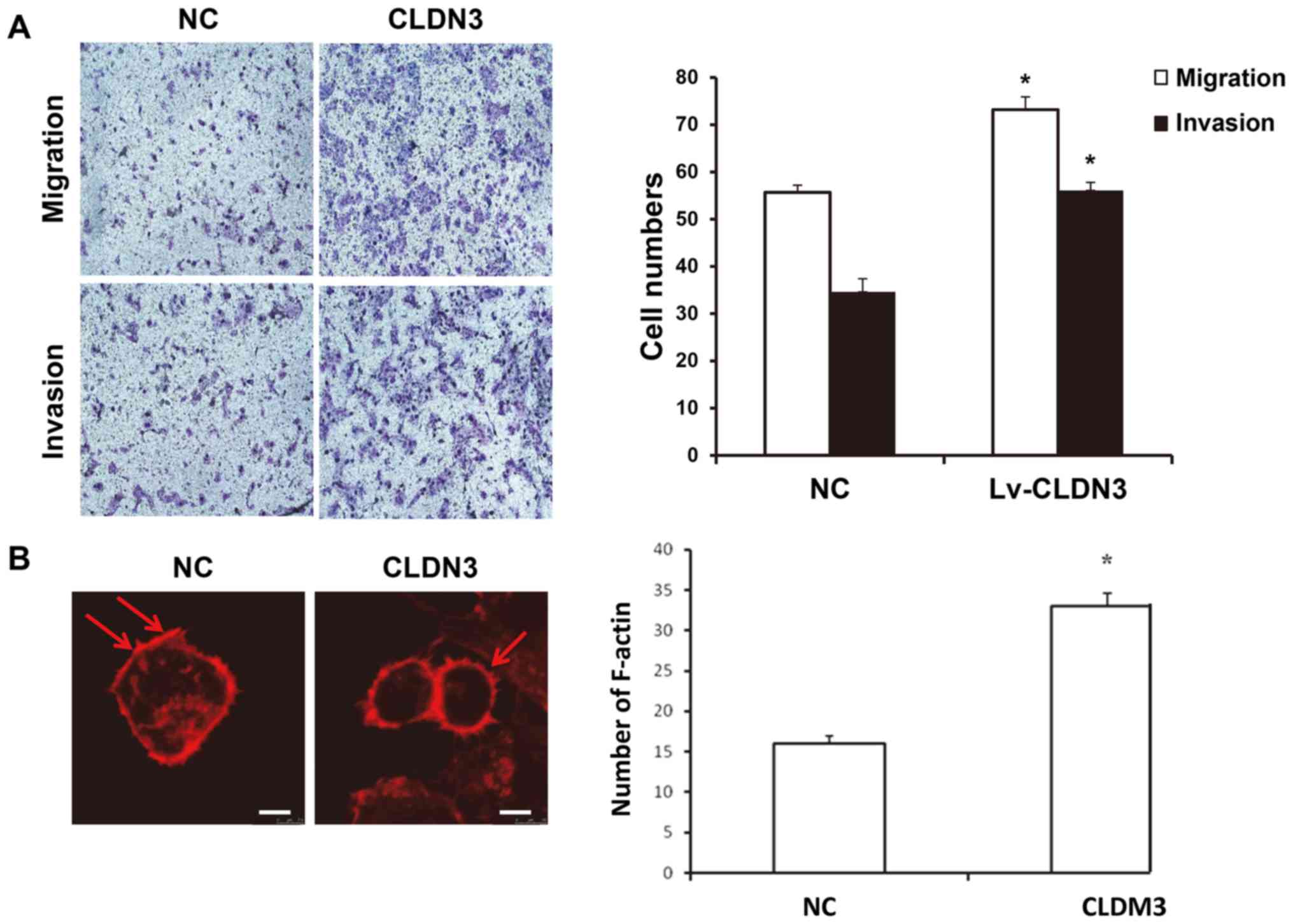
CLDN3 overexpression promotes the
invasive and migratory abilities of HTR8/SVneo cells
To investigate the effect of CLDN3 on the invasive
and migratory abilities of HTR8/SVneo cells, Transwell chamber
assays were performed. Compared with the control group, CLDN3
overexpression significantly promoted the migratory and invasive
capabilities of HTR8/SVneo cells (P<0.05; Fig. 3A). In addition, CLSM results showed
that in HTR8/SVneo cells overexpressing CLDN3, the number of
F-actin myofibers was significantly increased, indicating increased
migratory ability (P<0.05; Fig.
3B). These results suggest that CLDN3 overexpression promotes
HTR8/SVneo cell migration and invasion.
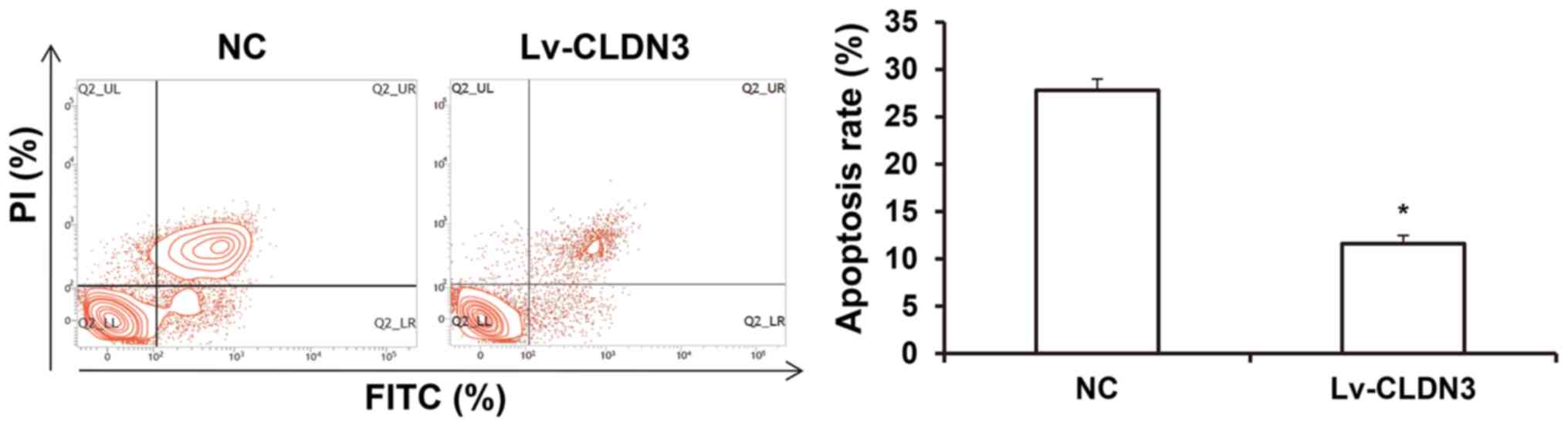
Effects of CLDN3 overexpression on
HTR8/SVneo cell apoptosis
Apoptosis is also an important factor affecting the
migratory and invasive abilities of HTR8/SVneo cells (29). To investigate the effects of CLDN3 on
apoptosis in HTR8/SVneo cells, flow cytometry was performed.
Compared with the control group, the overexpression of CLDN3
significantly reduced HTR8/SVneo cell apoptosis (P<0.05;
Fig. 4). These results suggest that
CLDN3 overexpression can suppress apoptosis in HTR8/SVneo
cells.
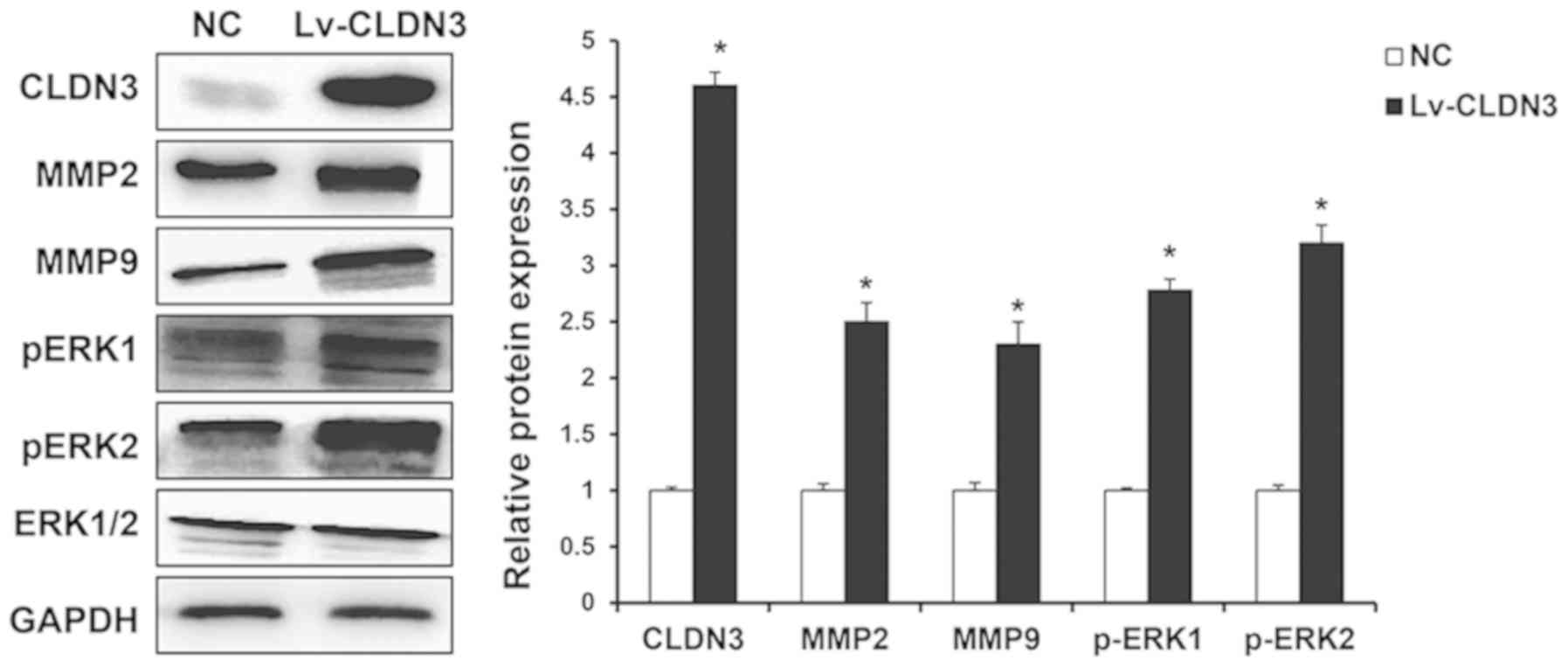
Effects of CLDN3 on the ERK1/2
signaling pathway and MMP expression in HTR8/SVneo cells
The ERK1/2/MMP pathway serves an important role in
the cell migration and is regulated by tight junction proteins
(6). To investigate the effects of
CLDN3 on the ERK1/2 signaling pathway (30) in addition to MMP-2 and MMP-9
expression in HTR8/SVneo cells, western blot analysis was performed
following CLDN3 overexpression. The overexpression of CLDN3
significantly elevated the expression levels of MMP-2 and MMP-9 in
HTR8/SVneo cells. ERK1/2 phosphorylation was also significantly
increased following CLDN3 overexpression (Fig. 5). These results suggest that CLDN3
can significantly affect the expression of MMPs and ERK activation
in HTR8/SVneo cells.
Discussion
Pregnancy-induced hypertension is a common
complication in obstetrics (31).
Clinical manifestations of pregnancy-induced hypertension include
proteinuria and fetal growth restriction after 20 weeks of
gestation (32). At present, the
pathogenesis of this disease remains poorly characterized (33). A previous study has demonstrated that
in most women with pregnancy-induced hypertension, the symptoms of
hypertension typically disappear following the delivery of
placenta, suggesting that the placenta may serve an important role
in the development of pregnancy-induced hypertension (34). Trophoblast cells are important for
the structure and function of placenta, where the tight junction
proteins serve important roles in maintaining the cellular polarity
and integrity of cell barriers. However, whether or not the tight
junction proteins regulate the biological function of trophoblast
cells remains unclear. The present study demonstrated that the
expression levels of the tight junction protein CLDN3 were
significantly downregulated in the placental tissues of patients
with pregnancy-induced hypertension compared with healthy pregnant
controls. Supporting this, in vitro experiments using human
trophoblast cell line HTR8/SVneo cells showed that CLDN3
overexpression promoted cell proliferation and invasion, possibly
by activating the ERK1/2/MMP-2/MMP-9 signaling pathway.
The placenta is the location where gas and nutrient
exchange occurs and where metabolic products are eliminated between
the mother and the fetus. As part of the main placental structure,
invasion of trophoblast cells into the endometrium is an important
physiological process in the formation of the placenta (35,36).
Studies have shown that extravillous trophoblast cells invade into
the uterine decidua and myometrial spiral artery, where they
gradually replace the endothelial cell layer and part of the smooth
muscle tissue in the maternal spiral arteries, resulting in
thickened blood vessels to provide sufficient blood supply to the
fetus (37,38). Trophoblast cell invasion is a key
factor to this process, and factors hindering this physiological
process may lead to placental dysplasia, pregnancy-induced
hypertension and eclampsia (39). In
particular, one of the pathological changes in the placenta during
pre-eclampsia is the shallow and limited invasion of trophoblast
cells into the uterine spiral artery (40).
Tight junction proteins, of which the CLDN family of
proteins is an important example, serve important roles in the
maintenance of cell barriers and polarity. In recent years, a large
number of studies have shown that CLDNs are closely associated with
tumor cell invasion and metastasis (41). Indeed, CLDN2 has been reported to
promote the self-renewal of colon cancer stem cells (42), whereas the downregulation of CLDN12
expression by interleukin (IL)-18 has been demonstrated to activate
the p38 pathway to promote the invasion and metastasis of breast
cancer in another recent study (43). CLDN3 is an important member of the
CLDN family. Studies have confirmed the involvement of CLDN3 in the
regulation of tumor cell invasion and metastasis through multiple
signaling pathways. Ahmad et al (44) showed that the loss of CLDN3
expression activated the IL-6/glycoprotein 130/STAT3 pathway to
promote the malignant transformation of colon cancer. In lung
squamous cell carcinoma, CLDN3 could inhibit the invasion and
metastasis of lung cancer cells (45). However, it remains to be elucidated
whether CLDN3 is involved in the regulation of human trophoblast
cell invasion. Results from the present study showed that the mRNA
expression levels of CLDN3 in the placental tissues and peripheral
blood of patients with pregnancy-induced hypertension were
significantly reduced compared with those in healthy controls,
indicating a negative association with disease pathogenesis. The
in vitro experiments of the present study indicated that the
overexpression of CLDN3 promoted HTR8/SVneocell proliferation,
migration and invasion. During invasion, human trophoblast cells
would detach from adjacent cells or the extracellular matrix.
Normal cells undergo apoptosis to maintain the stability of the
tissue environment, whereas tumor cells or other migratory cells
may suppress apoptosis to promote their migration. In the present
study, CLDN3 overexpression significantly inhibited the apoptosis
of HTR8/SVneo cells. These observations suggest that CLDN3 promotes
the proliferation, in addition to the invasive and migratory
abilities of HTR8/SVneo cells in vitro. The present study
demonstrated changes in CLDM3 expression in human tissues and
cells, and also detected similar changes in the levels of CLDN3 in
the peripheral blood. From these observations it could be
hypothesized that the placental tissue released CLDM3 into the
circulation; however, there isno direct evidence that the placenta
is the sole source of CLDM3.
It has previously been found that human trophoblast
cells secrete MMPs to degrade the extracellular matrix during
invasion (46). MMP-2 and MMP-9 are
the main enzymes involved in the degradation of the basal membrane
of the uterus (47). It has been
reported that MMP-2 and MMP-9 could assist in the invasion of human
trophoblast cells, and MMP-2 and MMP-9 can degrade extracellular
matrix, making cells more susceptible to metastasis and
infiltration (48,49). In the present study, results from
western blot analysis showed that CLDN3 overexpression
significantly elevated the expression levels of MMP-2 and MMP-9 in
human trophoblast cells. Regulation of MMPs involves a number of
different factors, including cytokine profiles, signaling pathway
activation and MMP-inhibiting factors. A previous study found that
both the expression and activity of MMP-9 are regulated by the
ERK1/2 signaling pathway (50).
Indeed, the present study showed that the overexpression of CLDN3
significantly elevated the phosphorylation levels of ERK1 and ERK2
in HTR8/SVneo cells, suggesting that CLDN3 activates the ERK1/2
signaling pathway.
However, in the present study, although the
expression levels of CLDN3 in the placental tissue and the effects
on the biological function of trophoblast cells were investigated,
only a single trophoblast cell line was used, and further in-depth
studies on primary trophoblast cells are required to confirm the
regulatory effects of CLDN3.
In conclusion, the present study showed that CLDN3
can promote human trophoblast cell proliferation, migration and
invasion, with the underlying mechanism possibly involving the
upregulation of MMP-2 and MMP-9 expression levels via the ERK1/2
signaling pathway. These findings suggest that downregulation of
CLDN3 may be associated with the pathogenesis of pregnancy-induced
hypertension, which may serve to assist in the design of
therapeutic interventions for the treatment of this disease in the
clinic.
Acknowledgements
The authors thank Director Ailan Wang from the Laiwu
Maternal and Child Health Hospital (Shandong, China) for kind
assistance in the study design, experiment performance, data
analysis and manuscript preparation.
Funding
No funding was received.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
AZ, YQ and KL contributed to the study design,
experiment performance, data collection and analysis, and
manuscript preparation. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
The study was approved by the ethics review board of
Laiwu Maternal and Child Health Hospital (Shandong, China). Prior
written informed consent was obtained from every patient.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Ali AM, Alobaid A, Malhis TN and Khattab
AF: Effect of vitamin D3 supplementation in pregnancy on risk of
pre-eclampsia-randomized controlled trial. Clin Nutr. 38:557–563.
2018.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Leikin JB: Renal complications during
pregnancy: In the hypertension spectrum. Dis Mon.
65(24)2018.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Ray JG, Diamond P, Singh G and Bell CM:
Brief overview of maternal triglycerides as a risk factor for
pre-eclampsia. BJOG. 113:379–386. 2006.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Tripathy K, Chawla R, Mutha V and Selvan
H: Spontaneous suprachoroidalhaemorrhage with exudative retinal
detachment in pregnancy-induced hypertension. BMJ Case Rep.
9(223907)2018.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Shawkat E, Mistry H, Chmiel C, Webster L,
Chappell L, Johnstone ED and Myers JE: The effect of labetalol and
nifedipine MR on blood pressure in women with chronic hypertension
in pregnancy. Pregnancy Hypertens. 11:92–98. 2018.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Rhodes CA, Beevers DG and Churchill D: A
randomized trial of ambulatory blood pressure monitoring versus
clinical blood pressure measurement in the management of
hypertension in pregnancy. A feasibility study. Pregnancy
Hypertens. 11:142–144. 2018.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Braga F, Ferraro S, Borille S and
Panteghini M: Biological variation of two serum markers for
preeclampsia prediction. Clin Chem Lab Med. 28:e27–e28.
2019.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Chatterjee P, Chiasson VL, Seerangan G, De
Guzman E, Milad M, Bounds KR, Gasheva O, Tobin RP, Hatahet M,
Kopriva S, et al: Depletion of MHC class II invariant chain peptide
or γ-δ T-cells ameliorates experimental preeclampsia. Clin Sci
(Lond). 131:2047–2058. 2017.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Webster LM, Gill C, Seed PT, Bramham K,
Wiesender C, Nelson-Piercy C, Myers JE and Chappell LC: Chronic
hypertension in pregnancy: The impact of ethnicity and superimposed
preeclampsia on placental, endothelial and renal biomarkers. Am J
Physiol Regul Integr Comp Physiol. 1:R36–R47. 2018.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Sava RI, March KL and Pepine CJ:
Hypertension in pregnancy: Taking cues from pathophysiology for
clinical practice. Clin Cardiol. 41:220–227. 2018.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Gingrich J, Pu Y, Roberts J, Karthikraj R,
Kannan K, Ehrhardt R and Veiga-Lopez A: Gestational bisphenol S
impairs placental endocrine function and the fusogenic trophoblast
signaling pathway. Arch Toxicol. 92:1861–1876. 2018.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Chang TA, Bondarenko GI, Gerami-Naini B,
Drenzek JG, Durning M, Garthwaite MA, Schmidt JK and Golos TG:
Trophoblast differentiation, invasion and hormone secretion in a
three-dimensional in vitro implantation model with rhesus monkey
embryos. Reprod Biol Endocrinol. 16(24)2018.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Lee CQE, Turco MY, Gardner L, Simons BD,
Hemberger M and Moffett A: Integrin α2 marks a niche of trophoblast
progenitor cells in first trimester human placenta. Development.
16(145)2018.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Qin X, Liang Y, Guo Y, Liu X, Zeng W, Wu
F, Lin Y and Zhang Y: Eukaryotic initiation factor 5A and
Ca2+ /calmodulin-dependent protein kinase 1D modulate
trophoblast cell function. Am J Reprod Immunol.
80(e12845)2018.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Xu J, Xia Y, Zhang H, Guo H, Feng K and
Zhang C: Overexpression of long non-coding RNA H19 promotes
invasion and autophagy via the PI3K/AKT/mTOR pathways in
trophoblast cells. Biomed Pharmacother. 101:691–697.
2018.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Midic U, Goheen B, Vincent KA, VandeVoort
CA and Latham KE: Changes in gene expression following long-term in
vitro exposure of macacamulatta trophoblast stem cells to
biologically relevant levels of endocrine disruptors. Reprod
Toxicol. 77:154–165. 2018.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Zheng J, Wang H and Zhou W: Modulatory
effects of trophoblast-secreted CXCL12 on the migration and
invasion of human first-trimester decidual epithelial cells are
mediated by CXCR4 rather than CXCR7. Reprod Biol Endocrinol.
16(17)2018.PubMed/NCBI View Article : Google Scholar
|
|
18
|
HudonThibeault AA, Vaillancourt C and
Sanderson JT: Profile of CYP19A1 mRNA expression and aromatase
activity during syncytialization of primary human villous
trophoblast cells at term. Biochimie. 148:12–17. 2018.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Tian H, Miao J, Zhang F, Xiong F, Zhu F,
Li J, Wang X, Chen S, Chen J, Huang N and Wang Y: Non-histone
nuclear protein HMGN2 differently regulates the urothelium barrier
function by altering expression of antimicrobial peptides and tight
junction protein genes in UPEC J96-infected bladder epithelial cell
monolayer. Acta Biochim Pol. 65:93–100. 2018.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Matsuoka H, Tamura A, Kinehara M, Shima A,
Uda A, Tahara H and Michihara A: Levels of tight junction protein
CLDND1 are regulated by microRNA-124 in the cerebellum of
stroke-prone spontaneously hypertensive rats. Biochem Biophys Res
Commun. 498:817–823. 2018.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Altunbulakli C, Costa R, Lan F, Zhang N,
Akdis M, Bachert C and Akdis CA: Staphylococcus aureus enhances the
tight junction barrier integrity in healthy nasal tissue, but not
in nasal polyps. J Allergy Clin Immunol. 142:665–668.
2018.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Kim SO, Choi BT, Choi IW, Cheong J, Kim
GY, Kwon TK, Kim ND and Choi YH: Anti-Invasive activity of histone
deacetylase inhibitors via the induction of Egr-1 and the
modulation of tight junction-related proteins in human
hepatocarcinoma cells. BMB Rep. 42:655–660. 2009.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Jaaskelainen A, Soini Y, Jukkola-Vuorinen
A, Auvinen P, Haapasaari KM and Karihtala P: High-Level cytoplasmic
claudin 3 expression is an independent predictor of poor survival
in triple-negative breast cancer. BMC Cancer.
18(223)2018.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Piontek A, Rossa J, Protze J, Wolburg H,
Hempel C, Günzel D, Krause G and Piontek J: Polar and charged
extracellular residues conserved among barrier-forming claudins
contribute to tight junction strand formation. Ann N Y Acad Sci.
1397:143–156. 2017.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Torres-Martinez AC, Gallardo-Vera JF,
Lara-Holguin AN, Montano LF and Rendon-Huerta EP: Claudin-6
enhances cell invasiveness through claudin-1 in AGS human
adenocarcinoma gastric cancer cells. Exp Cell Res. 350:226–235.
2017.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Akizuki R, Shimobaba S, Matsunaga T, Endo
S and Ikari A: Claudin-5, -7, and -18 suppress proliferation
mediated by inhibition of phosphorylation of akt in human lung
squamous cell carcinoma. Biochim Biophys Acta. 1864:293–302.
2017.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Deng Q, Yin N, Chen Y, Shan N, Liu X and
Qi H: Downregulated N-acetylglucosaminyltransferase III is involved
in attenuating trophoblast migration and invasion under
hypoxia-reoxygenation condition. J Matern Fetal Neonatal Med.
32:1–7. 2018.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Li X, Ma XL, Tian FJ, Wu F, Zhang J, Zeng
WH, Lin Y and Zhang Y: Downregulation of CCNA2 disturbs trophoblast
migration, proliferation, and apoptosis during the pathogenesis of
recurrent miscarriage. Am J Reprod Immunol.
82(e13144)2019.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Mishra JS, Te Riele GM, Qi QR, Lechuga TJ,
Gopalakrishnan K, Chen DB and Kumar S: Estrogen receptor-β mediates
estradiol-induced pregnancy-specific uterine artery endothelial
cell angiotensin type-2 receptor expression. Hypertension.
74:967–974. 2019.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Rashidi F and Sate H: Pregnancy outcome in
a pregnant patient with idiopathic pulmonary arterial hypertension:
A case report and review of the literature. J Med Case Rep.
12(31)2018.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Liao QP, Xu Q and Yan JY: SHH expression
in placental tissues and trophoblast cell oxidative stress injury
during preeclampsia. Eur Rev Med Pharmacol Sci. 23:6026–6034.
2019.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Chen S, Li N, Mei Z, Ye R, Li Z, Liu J and
Serdula MK: Micronutrient supplementation during pregnancy and the
risk of pregnancy-induced hypertension: A randomized clinical
trial. Clin Nutr. 38:146–151. 2018.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Tateishi A, Ohira S, Yamamoto Y and Kanno
H: Histopathological findings of pregnancy-induced hypertension:
Histopathology of early-onset type reflects two-stage disorder
theory. Virchows Arch. 472:635–642. 2018.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Yang C, Lim W, Bazer FW and Song G: Butyl
paraben promotes apoptosis in human trophoblast cells through
increased oxidative stress-induced endoplasmic reticulum stress.
Environ Toxicol. 33:436–445. 2018.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Nikolaou S, Hadjikypri X, Ioannou G, Elia
A and Georgiades P: Functional and phenotypic distinction of the
first two trophoblast subdivisions and identification of the border
between them during early postimplantation: A prerequisite for
understanding early patterning during placentogenesis. Biochem
Biophys Res Commun. 496:64–69. 2018.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Trinh QD, Pham NTK, Takada K,
Komine-Aizawa S and Hayakawa S: Myelin oligodendrocyte
glycoprotein-independent rubella infection of keratinocytes and
resistance of first-trimester trophoblast cells to rubella virus in
vitro. Viruses. 10(E23)2018.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Xu P, Zhao Y, Liu K, Lin S, Liu X, Wang M,
Yang P, Tian T, Zhu YY and Dai Z: Prognostic role and clinical
significance of trophoblast cell surface antigen 2 in various
carcinomas. Cancer Manag Res. 9:821–837. 2017.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Cheng JC, Yi Y, Chang HM and Leung PCK:
TGF-β1 upregulates cadherin-11 expression through snail: A
potential mechanism for human trophoblast cell differentiation.
Cell Signal. 43:55–61. 2018.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Chang CW, Wakeland AK and Parast MM:
Trophoblast lineage specification, differentiation and their
regulation by oxygen tension. J Endocrinol. 236:R43–R56.
2018.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Hashimoto Y, Fukasawa M, Kuniyasu H, Yagi
K and Kondoh M: Claudin-Targeted drug development using
anti-claudin monoclonal antibodies to treat hepatitis and cancer.
Ann N Y Acad Sci. 1397:5–16. 2017.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Paquet-Fifield S, Koh SL, Cheng L, Beyit
LM, Shembrey C, Mølck C, Behrenbruch C, Papin M, Gironella M,
Guelfi S, et al: Tight junction protein claudin-2 promotes
self-renewal of human colorectal cancer stem-like cells. Cancer
Res. 78:2925–2938. 2018.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Yang Y, Cheon S, Jung MK, Song SB, Kim D,
Kim HJ, Park H, Bang SI and Cho D: Interleukin-18 enhances breast
cancer cell migration via downregulation of claudin-12 and
induction of the p38 MAPK pathway. Biochem Biophys Res Commun.
459:379–386. 2015.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Ahmad R, Kumar B, Chen Z, Chen X, Müller
D, Lele SM, Washington MK, Batra SK, Dhawan P and Singh AB: Loss of
claudin-3 expression induces IL6/gp130/Stat3 signaling to promote
colon cancer malignancy by hyperactivating wnt/β-catenin signaling.
Oncogene. 36:6592–6604. 2017.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Che J, Yue D, Zhang B, Zhang H, Huo Y, Gao
L, Zhen H, Yang Y and Cao B: Claudin-3 inhibits lung squamous cell
carcinoma cell epithelial-mesenchymal transition and invasion via
suppression of the wnt/β-catenin signaling pathway. Int J Med Sci.
15:339–351. 2018.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Chaudhary P, Babu GS, Sobti RC and Gupta
SK: HGF regulate HTR-8/SVneo trophoblastic cells migration/invasion
under hypoxic conditions through increased HIF-1α expression via
MAPK and PI3K pathways. J Cell Commun Signal. 13:503–521.
2019.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Liu Y, Shan N, Yuan Y, Tan B, He C, Tong C
and Qi H: Knockdown of activated Cdc42-associated kinase inhibits
human extravillous trophoblast migration and invasion and decreases
protein expression of pho-akt and matrix metalloproteinase. J
Matern Fetal Neonatal Med. 33:1–9. 2018.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Zhong T, Chen J, Ling Y, Yang B, Xie X, Yu
D, Zhang D, Ouyang J and Kuang H: Down-Regulation of neuropathy
target esterase in preeclampsia placenta inhibits human trophoblast
cell invasion via modulating MMP-9 levels. Cell Physiol Biochem.
45:1013–1022. 2018.PubMed/NCBI View Article : Google Scholar
|
|
49
|
Zong L, Wei X, Gou W, Huang P and Lv Y:
Zinc improves learning and memory abilities of fetal growth
restriction rats and promotes trophoblast cell invasion and
migration via enhancing STAT3-MMP-2/9 axis activity. Oncotarget.
8:115190–115201. 2017.PubMed/NCBI View Article : Google Scholar
|
|
50
|
Fu Y, Wei J, Dai X and Ye Y: Increased
NDRG1 expression attenuate trophoblast invasion through ERK/MMP-9
pathway in preeclampsia. Placenta. 51:76–81. 2017.PubMed/NCBI View Article : Google Scholar
|