Introduction
Systemic lupus erythematosus (SLE) is an autoimmune
disease that is characterized by the production of autoantibodies
and immune complexes that may affect the majority of organs in the
human body (1). Lupus nephritis
(LN) is the most common and serious complication of SLE and is the
major cause of morbidity and mortality in patients with this
condition (2). A previous survey
demonstrated that at least 33% of patients with SLE also have a
manifestation of significant glomerulonephritis (3).
The origins of SLE remain to be fully elucidated. It
is generally thought that the imbalance between cellular and
humoral immunity in patients with lupus is caused by a complex
interaction between genetic and environmental factors and the
abnormal function of T- and B-cells that ultimately destroy normal
immune tolerance mechanisms. During the initiation of a normal
immune response, T-cell activation, proliferation, differentiation,
cytokine secretion and avoidance of apoptosis require at least two
signals (4). These signals include
a specific signal that is delivered by the antigen peptide
presented by the major histocompatibility complex and a
non-specific signal produced by the interaction of co-stimulatory
molecules (5). Co-stimulatory
pathways were first proposed and confirmed on the basis of a T-cell
activation double signal theory (6).
B7, an important co-stimulatory molecule that is
expressed on the surface of antigen-presenting cells (APCs), exists
primarily in two forms, B7-1 and B7-2. The B7 receptor on the
surface of T-cells is CD28/cytotoxic T lymphocyte antigen-4
(CTLA-4) and the interaction of B7 with CD28 promotes T-cell
activation, proliferation and secretion of cytokines, as well as
regulating T-helper (TH)1/TH2 cell
differentiation and contributing to B-cell antibody production and
isotype switching (7). The
interaction of B7 with CTLA-4 inhibits T-cell activation and
proliferation (5,8). A previous study demonstrated that the
B7-CD28 co-stimulatory signal is associated with the development of
SLE (9). Therefore, blocking or
weakening of the induced signal may help reduce the associated
pathological damage or the development of other immune-mediated
diseases.
In this present study, a monoclonal antibody (mAb)
against human B7-1 was generated and characterized. The potential
efficacy against SLE-related pathologies was assessed using a
lupus-like nephritis mouse model, a generally-accepted animal model
induced by the injection of parental BALB/c lymphocytes into BALB/c
x C57BL/J6 F1 hybrids, leading to chronic graft-vs.-host
disease (cGVHD) (10). This model
has been previously verified and characterized as an SLE-like
disease model that displays lymphoid hyperplasia, the formation of
autoantibodies similar to those occurring in patients with SLE and
an increased presence of renal pathologies mediated in part by
immune complex formation and deposition (11).
Materials and methods
Cell lines
The human B7-1-transfected L929 cell line
(L929/B7-1) and mock-transfected L929 cells (L929/mock) were
previously generated and used in this present study (12). Daudi and Raji cell lines were
purchased from the American type culture collection. The cell lines
were cultured in RPMI-1640 medium (Gibco; Thermo Fisher Scientific,
Inc.) containing 10% fetal bovine serum (HyClone; GE Life Sciences)
100 U/ml penicillin and 100 mg/ml streptomycin (Gibco; Thermo
Fisher Scientific, Inc.). Cells were maintained at 37˚C in a
humidified atmosphere containing 5% CO2.
Animals
BALB/c mice (10; female; age, 6-8 weeks; weight,
25±2 g) and C57BL/J6 mice (10; male; age, 6-8 weeks; weight, 29±2
g) were purchased from the Department of Experimental Animals
(Shanghai Institute of Biological Products, Ministry of Health of
China). All mice were kept in specific pathogen-free animal
facilities maintained at 18-22˚C and 50-60% relative humidity, with
a 12-h light/dark cycle and provided ad libitum access to
standard rodent chow and filtered water. All experiments were
performed in accordance with the guidelines and approved by the
Ethics Committee of Soochow University (Suzhou, China; approval no.
201912A341).
Reagents and antibodies
Complete Freund's adjuvant (CFA), incomplete
Freund's adjuvant and hypoxanthine-aminopterin-thymidine (HAT)
selection medium were purchased from Sigma-Aldrich; Merck KGaA. The
rabbit anti-mouse antibodies used were as follows: Mouse isotype
immunoglobulin (Ig)G (cat. no. 555571), FITC-labeled anti-mouse
granulocyte receptor 1 antigen (GR-1; IgG2b; cat. no.
553127), FITC-labeled anti-mouse CD11b (IgG2b; cat. no.
553310), phycoerythrin (PE)-labeled anti-mouse CD23
(IgG2a; cat. no. 561773), PE-labeled anti-mouse
CD21(IgG2b; cat. no. 552957), PE-labeled anti-mouse CD86
(IgG2a; cat. no. 561963), PE-labeled anti-mouse CD80
(IgG2a; cat. no. 561955), FITC-labeled anti-mouse CD11c
(IgGl; cat. no. 561045), PE-labeled mouse IgG (cat. no.
555988), PE-labeled rat anti-mouse IgG1 (cat. no. 562027) and
allophycocyanin-labeled anti-mouse B220 (IgG2a; cat. no.
561880). All antibodies were purchased from BD Pharmingen.
Generation and characterization of
anti-human B7-1 mAbs
The hybridoma cell line from Shi et al
(12) has been observed to secrete
anti-human B7-1 antibodies. To generate anti-human B7-1, BALB/c
mice were immunized with L929/B7-1 cells (107 cells in
100 µl PBS) that were pretreated with 10 µg/ml mitomycin
(Sigma-Aldrich; Merck KGaA) for 2 h at 37˚C, by subcutaneous
injection into the right flank in conjunction with a separate
injection of 400 µl CFA. Mice then received three intraperitoneal
injections of the cells/CFA at 1-week intervals. Mice were
anesthetized (1% sodium pentobarbital; 50 mg/kg body weight) 4 days
after the final injection and euthanized by cervical dislocation.
The spleen was removed by opening the abdominal cavity and a single
cell suspension of spleen cells prepared by mechanized
homogenization with a syringe filled with PBS, filtration through a
filter screen (200 mesh), centrifugation of the suspension (233 x g
for 8 min at 4˚C) and removal of the supernatant. These cells were
then fused with murine myeloma SP2/0 cells using standard protocols
(13,14). The resulting hybridomas were
subsequently cultured at 37˚C for 10 days in HAT selection medium.
From the successful cultures, supernatants were evaluated for
antigen (B7-1) recognition. In brief, L929/B7-1 cells were
incubated with the supernatant at 4˚C for 45 min, then stained with
PE-labeled rat anti-mouse IgG1 (1:200) at 4˚C for 30 min. FACS
(Calibur system; BD Cell Quest Pro software; version 6.0; BD
Biosciences) was used in order to analyze the stained cells.
L929/B7-1 cells were also stained with PE-labeled rat anti-mouse
IgG1 (1:200) as a negative control.
The specific 4E5 mAb was purified from BALB/c mouse
ascites using Protein G-sepharose CL4B affinity columns (GE
Healthcare) following the manufacturer's protocols. Female BALB/c
mice were intraperitoneally injected with pristane (0.5 ml;
Sigma-Aldrich; Merck KGaA) and 1 week later, mice were injected
with a specific hybridoma line (intraperitoneally; 1x107
total cells/mouse). Mice were then injected with an equal volume of
a 1:1 (v/v) mixture of pristane and Freund's incomplete adjuvant.
After a period of 1 week, which facilitated fluid accumulation in
the abdomen (15), the ascites in
each mouse were extracted with a syringe and processed using CL4B
affinity column purification. The antibody was eluted with pH 2.8
glycine-hydrochloric acid mixture and the concentration was
estimated spectrophotometrically. This was calculated as protein
concentration (mg/ml)=[optical density at 280 nm
(OD280)x1.55]-(OD260x0.76). The purified
antibody was then conjugated with PE using standard protocols
(16). Flow cytometry was
subsequently used to verify the recognition of the B7-1 antigen by
the clone 4E5 mAb with L929/mock, L929/B7-1, Daudi and Raji cells,
as well as naive BALB/c mouse splenocytes, and performed as
described earlier.
Induction of the murine LN model of
cGVHD and 4E5 antibody treatment
Female BALB/c mice were mated with male C57BL/J6
mice to obtain F1 generations. When F1
hybrids reached the age of 6-8 weeks, the mice (all females
selected) were randomly allocated into four groups: Normal control
group (Mock, mice did not receive any treatment), model group (MG,
cGVHD was induced in the mice), 4E5-treated group (4E5, cGVHD was
induced in the mice and they were treated with 4E5) and a random
IgG-treated group (IgG, mice cGVHD was induced in the mice and they
were treated with IgG), with 15 mice in each group. Excluding the
Mock mice, all other F1 hybrids were used as recipients of female
BALB/c donor cells.
To prepare the cells for each injection, two naive
female BALB/c mice were anesthetized and euthanized using cervical
dislocation. After death, the thymus, spleen and axillary lymph
nodes of each mouse were recovered and single-cell suspensions were
prepared by mechanical homogenization with a syringe filled with
PBS, filtering through a filter screen (200 mesh), centrifuging the
suspension (233 x g for 8 min at 4˚C) and removing the supernatant
PBS. Then these single-cell suspensions of different organs from
two mice were mixed together and counted [following standard
treatments to remove contaminating erythrocytes as required
(12)]. In each case, four
individual intravenous injections were performed at 3-day intervals
and a 100 µl volume of the solution, containing 5x107
cells/100 µl of the mixture of fresh donor cells, was injected into
each recipient by tail vein. In the 4E5-treated mice, 4E5 mAb (10
mg/kg weight) diluted in PBS, was injected intravenously into the
tail on days 15, 17, 19, 21, 44 and 74 after the first lymphocyte
injection. This dose was selected based upon previous work reported
by Shi et al (12). The
IgG-treated mice were treated in parallel with mouse isotype IgG
(BD Pharmingen; BD Biosciences). From 2 weeks after the first
lymphocyte inoculation, mice received intraperitoneal anesthesia
(1% sodium pentobarbital; 50 mg/kg body weight) and 100 µl of blood
was drawn from the retro-orbital plexus every 30 days. Serum was
isolated and frozen at -80˚C for subsequent analyses of
autoantibodies. At the same time as the blood was collected, urine
from each mouse was also collected, by pressing the mouse bladder
and inserting a syringe into the urethra, for analysis of potential
proteinuria. In all cases, urine and blood were collected at 10 am
on the experimental day. At 12 weeks after the final inoculation,
all mice were euthanized by intraperitoneal injections of 2%
pentobarbital (120 mg/kg body weight) and death was verified using
cervical dislocation. Tissues (including spleen and kidney) were
harvested for subsequent analysis.
Immune response evaluated by flow
cytometry
A total of 5 mice were selected randomly from each
group at 3 weeks after the first lymphocyte injection and their
splenocytes were isolated as mentioned above. After counting,
distinct sets of 107 splenocytes were incubated with a
specific fluorochrome-conjugated antibody (GR-1,1:500; CD11b,
1:500; CD23, 1:200; CD21, 1:200; CD86, 1:200; CD80, 1:200; CD11c,
1:500 or B220, 1:200) for 30 min on ice and washed in PBS (pH 7.4)
for 30 min at room temperature. All samples were then analyzed
using flow cytometry as mentioned above with analysis using Cell
Quest Pro software (BD Biosciences). The major APC populations were
macrophages, dendritic cells and granulocytes, which were pre-gated
based on the APC activation markers GR-1, CD11b and CD11c in the
present study.
Anti-nuclear antibody (ANA) and
anti-double-stranded DNA (anti-dsDNA) measurements
To measure the levels of ANA and anti-dsDNA in
isolated sera, immunofluorescent staining was performed using an
ANA analysis kit (cat. no. YZB/Jing 1373-2009) and a daDNA analysis
kit (cat. no. YZB/Jing 1372-2009, Beijing H&J NovoMed, Co.,
Ltd.) following the manufacturer's protocol. Sera were diluted
(1:100 in PBS) and placed on cell-bearing glass slides containing
ANA or dsDNA antigen for incubation (30 min at room temperature) in
a humidified chamber. After being washed gently with PBS, the
presence of any ANA or anti-dsDNA was determined using
FITC-conjugated goat anti-mouse IgG antibodies with incubation for
30 min in the dark. Slides were washed again with PBS, air-dried
and then sealed. Images were captured using a fluorescence
microscope (Olympus Corporation). Levels of the autoantibodies were
quantified indirectly using measurements of fluorescence intensity
assessed in a blinded manner by three individuals using Image-Pro
Plus software version 5.0 (BioRad Laboratories, Inc.). Intensities
were then reported based using a semi-quantitative scale: Absent to
mild (low) and moderate to very severe (high).
Proteinuria measurement
The fresh urine collected from each mouse was
evaluated using Albustix urine dip sticks (Bayer AG), following the
manufacturer's protocol. The extent of proteinuria was scored in a
blinded manner by three individuals. Scores were reported as
follows: Complexes absent (-), or present at mild (+), moderate
(++), severe (+++) or very severe (++++) intensity.
Histopathology
A section from each mouse kidney dissected after
mouse death was fixed in 10% buffered formalin for 2 h at room
temperature and then embedded in paraffin. Sections (5 µm) were
then prepared and stained with hematoxylin and eosin for 10 min at
room temperature. The stained sections were then examined by light
microscopy in a blinded manner.
A second section (100 nm) of each kidney was fixed
in 2.5% glutaraldehyde buffer (pH 7.4) for 2 h at room temperature,
washed twice with PBS for 15 min and dehydrated using gradient
dilutions of acetone (70% acetone for 15 min, 80% for 15 min, 90%
for 15 min and 100% twice for 10 min). Dehydration of samples was
then performed using liquid CO2. Samples were then
sputter-coated with gold and examined using an H-800 transmission
electron microscope (Hitachi, Ltd.) (17).
Detection of immune complexes in
kidney tissue
The remaining parts of the kidney samples were
frozen in optimal cutting temperature medium (Thermo Fisher
Scientific, Inc.) and cut into 10-µm sections, which were air-dried
and fixed in acetone at -20˚C for 20 min. After being washed in
PBS, specimens were incubated with FITC-conjugated goat anti-mouse
IgG (1:100 in PBS) for 30 min at 37˚C. Samples were then washed
with PBS, air-dried and examined using a fluorescence microscope
(Olympus Corp.). Each sample was scored for fluorescence intensity
in a blinded manner by three individuals. Scores were reported as
follows: Complexes absent (-), or present at mild (+), moderate
(++), severe (+++) or very severe (++++) intensity.
Statistical analysis
All values are expressed as the mean ± standard
deviation. The statistical significance of differences in values
and frequencies between groups was evaluated using Student's
t-tests (2 groups) or analysis of variance with Tukey's post-hoc
test (>2 groups). All analyses were performed using SPSS 19.0
software (IBM, Corp.). P<0.05 was considered to indicate a
statistically significant difference.
Results
Generation and characterization of
anti-human B7-1 mAbs
Based upon previously established procedures
(12), a novel mAb targeting human
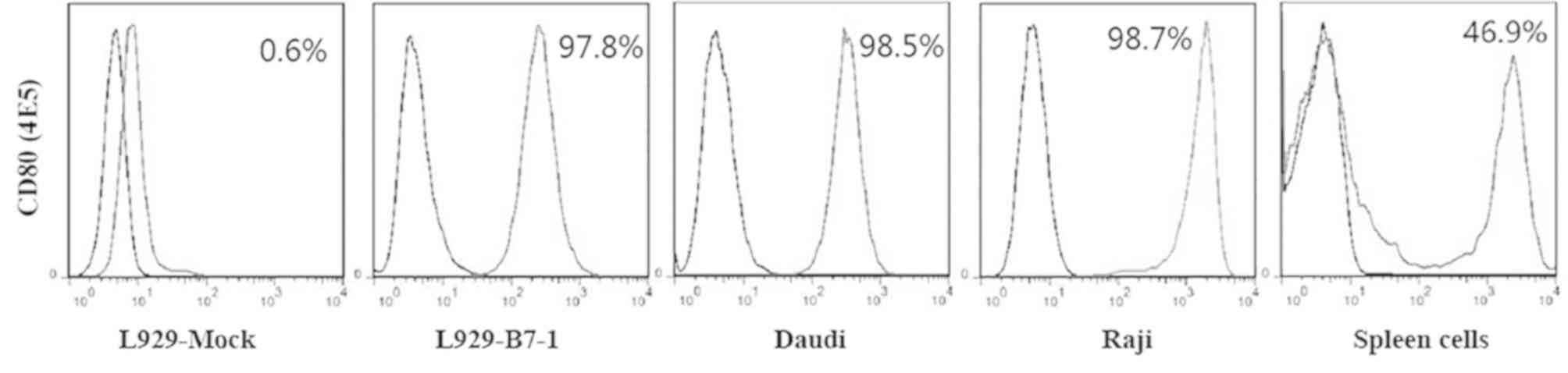
B7-1, 4E5, was generated (isotype IgG1). The flow cytometry results
showed that cells were 97.8% positive in the L929/B7-1 line, 98.5%
in the Daudi cell line and 98.7% positive in the Raji cell line,
(Fig. 1). As the spleen contains
several cell types, the spleen cells were only 46.9% positive.
These results indicated that 4E5 mAb was able to recognize B7-1 not
only on human cells (including L929/B7-1) but also on mouse
splenocytes.
4E5 mAb reduces the expression of
activation markers on mouse splenocytes
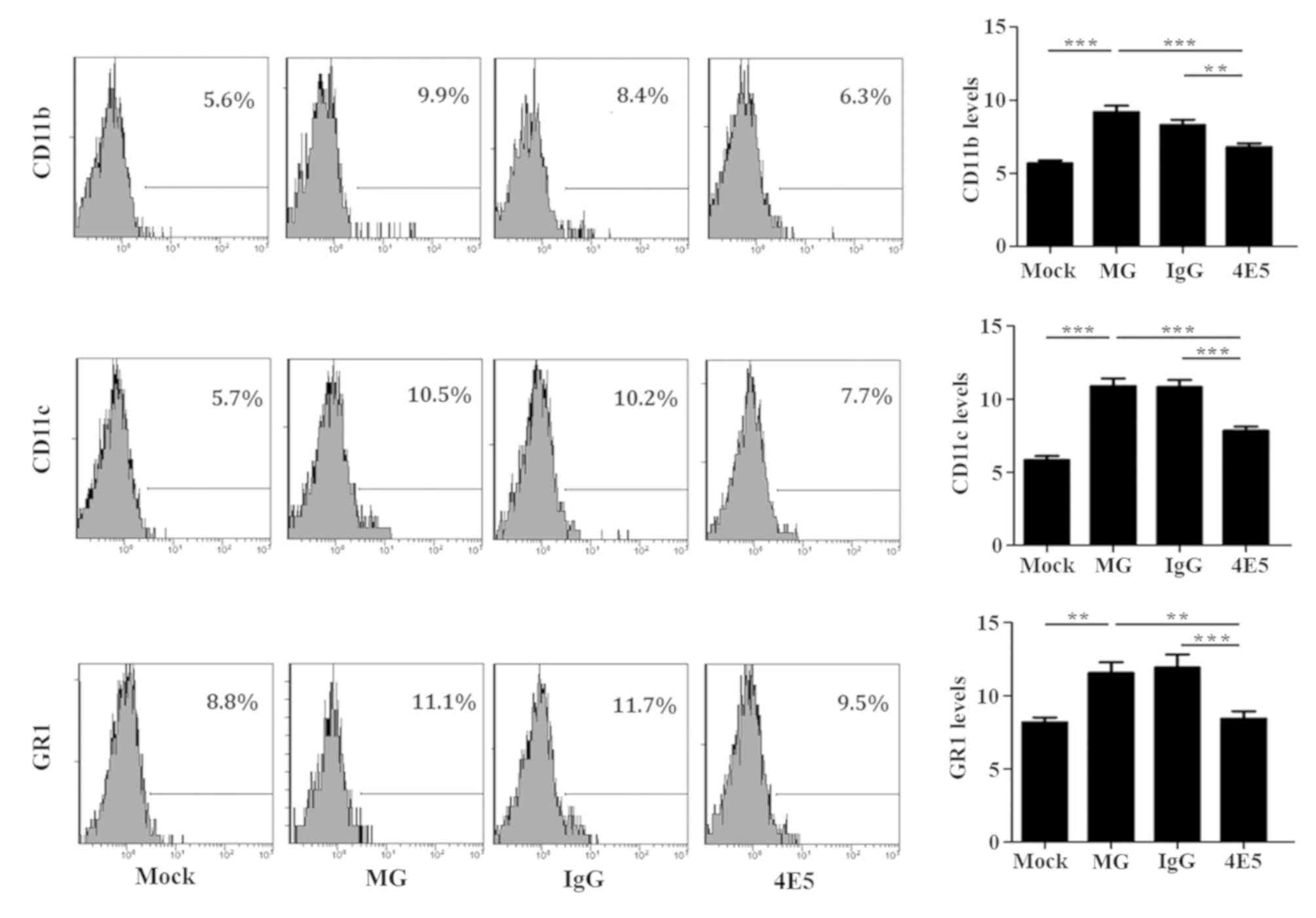
The expression of CD11b, CD11c and GR1 activation
markers on APC cells was higher among the mouse splenocytes from
the MG mice compared with that on cells from the Mock mice,
indicating that the spleen cells were abnormally activated. By
contrast, the frequency of the expression of each marker on cells
from the 4E5-treated mice was significantly lower compared with
that on cells from MG mice (Fig. 2,
P<0.05). There was no significant difference in the expression
levels of these markers between cells from MG mice and those from
IgG control mice, suggesting that the activation of APC cells in
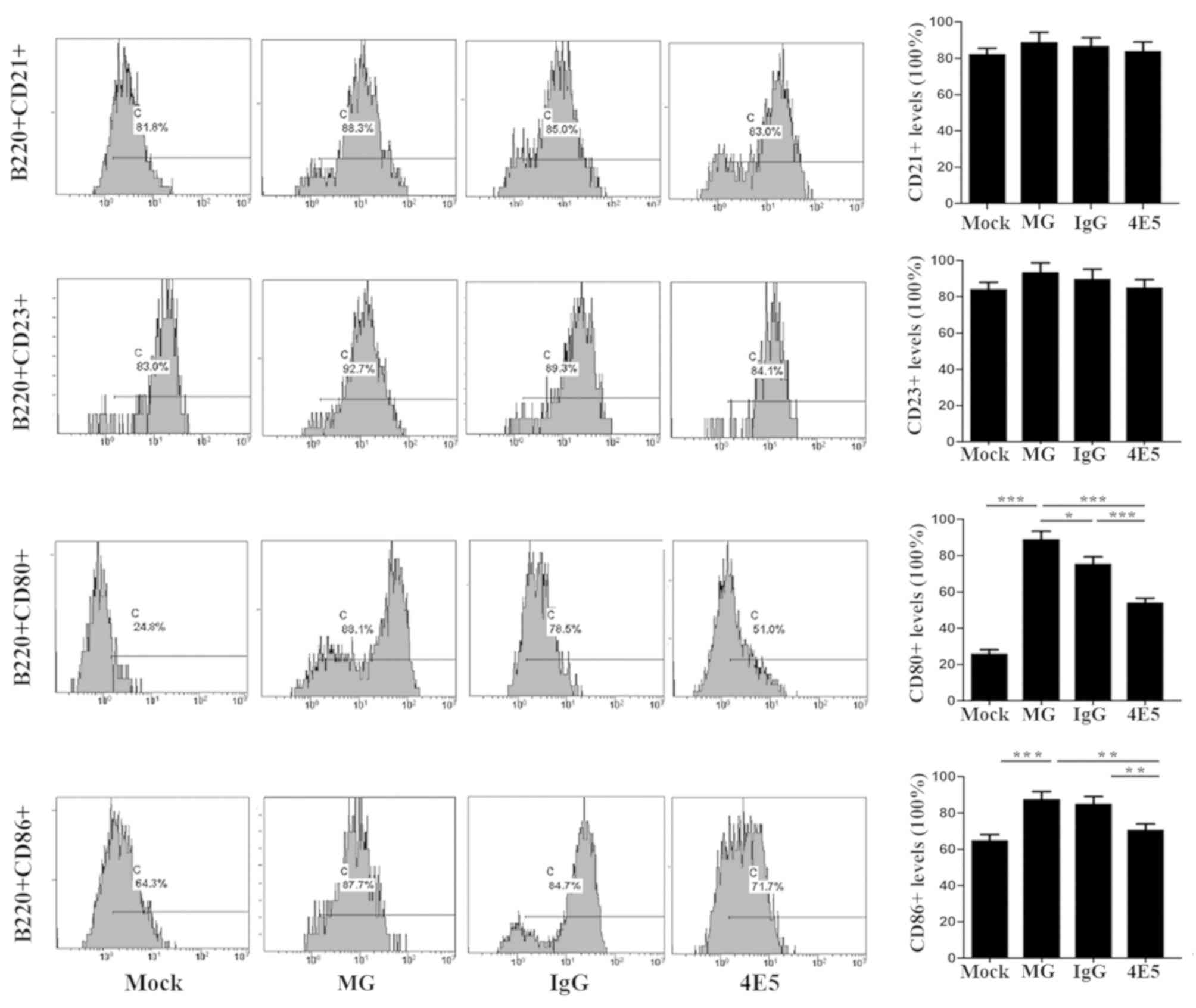
the spleens of mice that received 4E5 was inhibited. The expression
of the activation markers CD80 and CD86 on B220+ B-cells
from the 4E5-treated mice was also significantly lower compared
with that on B-cells from MG mice (Fig.
3, P<0.05). These results suggested that 4E5 mAb may
suppress immune responses that are normally induced by cGVHD.
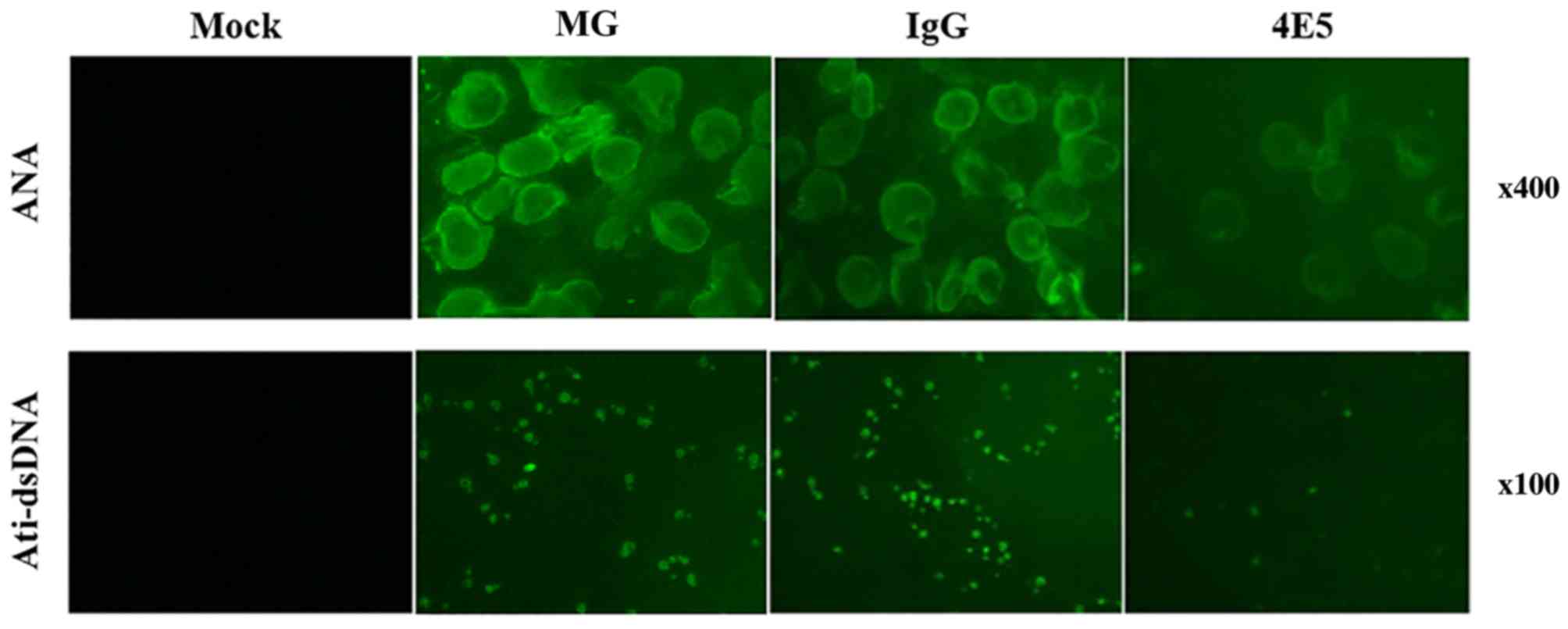
4E5 mAb treatment reduces the
production of ANA and anti-dsDNA
ANA and anti-dsDNA are cGVHD-induced lupus
autoantibodies (18). To assess the
severity of LN, the levels of each antibody in the sera were
evaluated (Fig. 4). The results
demonstrated that at 3 months after the initiation of the cell
injections, 30% of the MG mice and 10% of the 4E5-treated mice
exhibited high levels of ANA antibodies (P<0.05, Table I). At the 4-month time-point, 90% of
the MG mice and 50% of the 4E5-treated mice exhibited high levels
of ANA antibodies (P<0.05). At 3 months, 40% of the MG mice and
10% of the 4E5-treated mice exhibited high levels of circulating
anti-dsDNA antibodies (P<0.05). At 4 months, 100% of the MG mice
and 40% of the 4E5-treated mice expressed high levels of anti-dsDNA
antibodies (P<0.05). No significant differences were indicated
in the amount of serum ANA and anti-dsDNA antibodies at either
time-point between the MG mice and IgG control mice. ANA and
anti-dsDNA antibodies were not identified in the serum of the mock
mice at any time-point.
 | Table ISerum ANA and anti-dsDNA positivity
at 3 and 4 months after initiation of inoculations. |
Table I
Serum ANA and anti-dsDNA positivity
at 3 and 4 months after initiation of inoculations.
|
Time-point/autoantibody | Mock | MG | IgG-treated |
4E5-treateda |
|---|
| 3 months | | | | |
|
ANA | 0 (0) | 3(30) | 4(40) | 1(10) a |
|
Anti-dsDNA | 0 (0) | 4(40) | 4(40) | 1(10) a |
| 4 months | | | | |
|
ANA | 0 (0) | 9(90) | 9(90) | 5(50) a |
|
Anti-dsDNA | 0 (0) | 10(100) | 9(90) | 4(40) a |
4E5 mAb treatment reduces the
incidence of proteinuria
Proteinuria is a marker of renal lesions, and in the
present study, it was used as a criterion for the successful
induction of the LN model. Analysis of the urine collected monthly
following the initiation of lymphocyte inoculations indicated that
proteinuria appeared later in the 4E5 treatment group and was less
common compared with the frequencies in MG mice (Table II, P<0.05). At 3 months after
initiation of inoculation, 50% of MG mice and none of the
4E5-treated mice exhibited high proteinuria (moderate to very
severe; data not shown). The frequency of high proteinuria remained
higher in MG mice (100%) at 4 months after initiation of the
inoculations. By contrast, 50% of the 4E5-treated mice developed
high proteinuria at this time-point. The frequency of proteinuria
was not significantly different between the MG mice and IgG control
mice and no proteinuria was present in the mock mice at any
time-point.
| Table IIComparison of proteinuria status in
mice at 4 months after initiation of inoculations. |
Table II
Comparison of proteinuria status in
mice at 4 months after initiation of inoculations.
| Group | - | + | ++ | +++ | ++++ |
|---|
| Mock | 7 | 3 | 0 | 0 | 0 |
| MG | 0 | 0 | 4 | 5 | 1 |
| IgG-treated | 0 | 0 | 3 | 6 | 1 |
|
4E5-treateda | 1 | 4 | 5 | 0 | 0 |
Effects of 4E5 mAb treatment on the
formation of renal lesions
Histological analysis revealed that kidneys from the
MG mice exhibited a compensatory increase in the glomerular volume
at 4 months after the initiation of inoculation. The analysis also
revealed leukocytic infiltration in perivascular and internal
areas, as well as endothelial and mesangial hypercellularity. By
contrast, specimens from 4E5-treated mice exhibited slight
histopathological changes, including glomerular enlargement and
minor congestion of the glomerular vasculature (Fig. 5A). The IgG control mice also
demonstrated a number of histological changes compared with mock
mice.
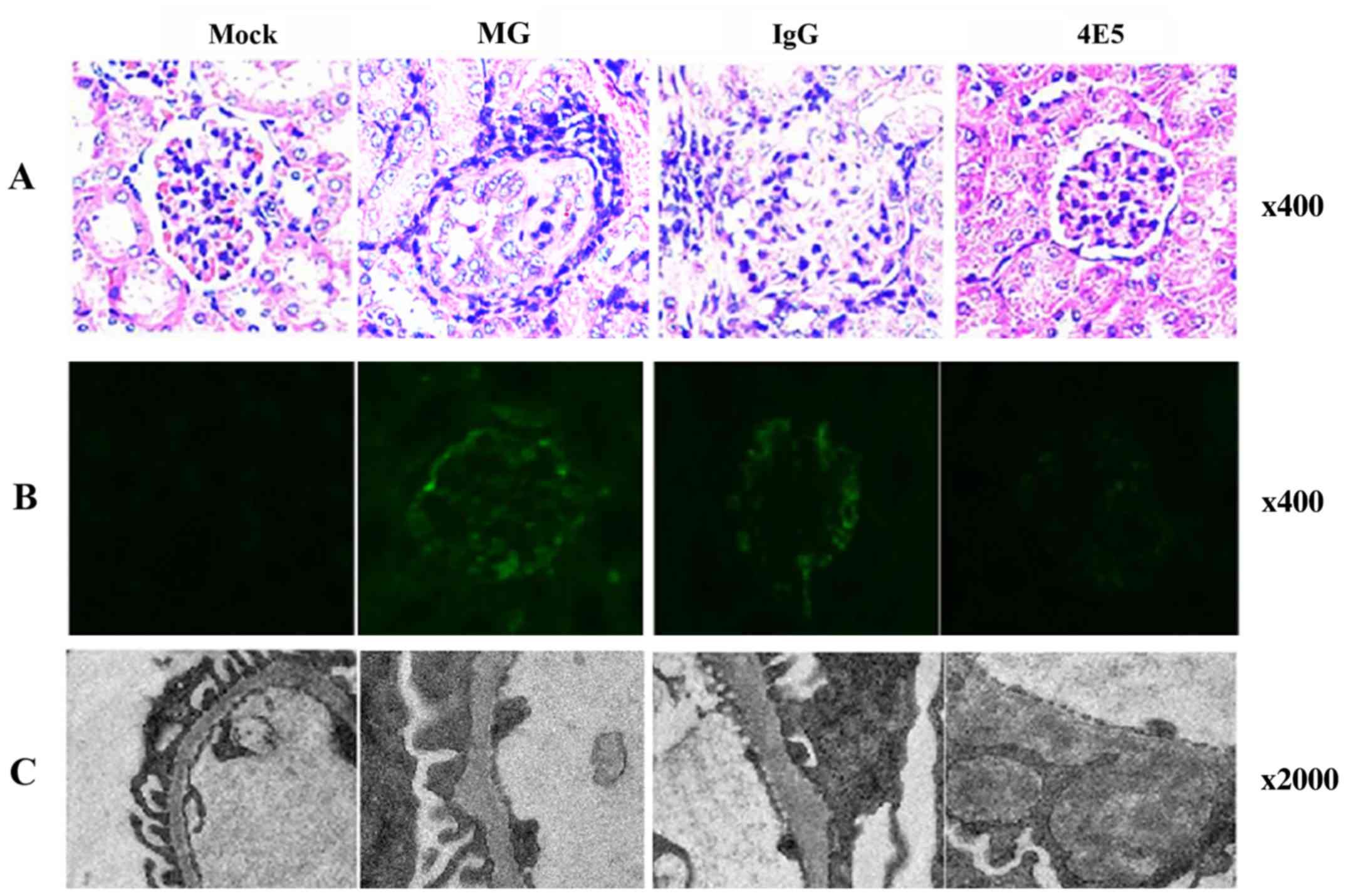 | Figure 5Analysis of kidneys at 4 months after
initiation of inoculations. (A) H&E staining was used to
evaluated the kidneys. The glomeruli of MG mice exhibited
glomerular volume compensatory enlargement and leukocytic
infiltration in perivascular and internal areas, as well as
endothelial and mesangial cell hyper-cellularity. Only minor
changes were noted in the organs of 4E5 mice when compared with
mock mice (magnification, x400). (B) Representative fluorescence
microscopy images of kidney sections from MG mice exhibited a
granular linear staining pattern of IgG deposits along glomerular
capillary loops; tissues of 4E5 mice had little-to-no staining
intensity, suggesting minimal deposits (magnification, x400). (C)
Representative transmission electron microscopy images revealing
electron-dense deposits localized in subepithelial lesions of
glomerular basement membrane in kidneys of MG mice; in glomeruli of
4E5 mice, the basement membrane layer was clear and intact, the
‘humps’ were less and smaller than MG group (magnification,
x2,000). ANA, anti-nuclear antibody; dsDNA, double-stranded DNA;
Ig, immunoglobulin; MG, model group. |
Effects of 4E5 mAb treatment on immune
complex deposition
Fluorescence analysis of kidney sections from the MG
mice at 4 months following the initiation of the inoculations
revealed a granular linear staining pattern of IgG deposits along
the glomerular capillary loops, indicating that immune complexes
were deposited in/on the loops of glomeruli. Although the
fluorescence intensity of mice treated with 4E5 decreased
significantly at the same time-point compared with MG group
(P<0.05), no glomerulonephritis was observed and significant
differences in the levels of immune complexes between the MG mice
and IgG control mice were indicated at 4 months after the
initiation of the inoculations (Table
III; Fig. 5B). No immune
complexes were observed in mock mice at 4 months after the
initiation of the inoculations.
| Table IIIComparison of fluorescence intensity
in the mice at 4 months after initiation of inoculations. |
Table III
Comparison of fluorescence intensity
in the mice at 4 months after initiation of inoculations.
| Group | - | + | ++ | +++ | ++++ |
|---|
| Mock | 5 | 5 | 0 | 0 | 0 |
| MG | 0 | 0 | 5 | 3 | 2 |
| IgG | 0 | 0 | 5 | 2 | 3 |
| 4E5a | 2 | 3 | 5 | 0 | 0 |
Effects of 4E5 mAb treatment on renal
ultrastructure
TEM analysis demonstrated that in the kidneys of the
MG and IgG control mice, electron-dense deposits were localized in
subepithelial lesions of the glomerular basement membrane that were
segmental and thickened. These deposits were also observed in
visceral epithelial cells that exhibited effaced foot processes
(humps). By contrast, in the glomerulus from 4E5-treated mice, the
basement membrane layer was clear and intact and there were fewer,
smaller ‘humps’ than in the MG group (Fig. 5C). No abnormal renal ultrastructure
was indicated in the kidneys isolated from mock mice.
Discussion
cGVHD, which is induced in recipient F1 hybrid mice
following the inoculation of parental lymphocytes, resembles SLE
with autoantibody production and glomerulonephritis (19). cGVHD is a sex-biased GVHD model in
which the female donor makes the female host more vulnerable to SLE
than a male (20-22).
cGVHD has been revealed to be due to donor alloreactive
CD4+ T-cell activation of host B-cells (23) and is characterized by lymphocyte
proliferation, the production of autoantibodies that resemble SLE
and serious renal pathology that is mediated by immune complexes.
The pathological characteristics mentioned above are similar to
those in human SLE; furthermore, it has a rapid onset (~2 months)
and is easy to reproduce with a low cost (24). The most commonly used model of SLE
worldwide is the MRL/lpr mouse model (25). These mice were produced by a series
of complex hybridizations to the 12th generation from a number of
different strains of mice, including LG/J, AKR/J, C3H/D and
C57BL/6. The Fas gene of these mice became mutated and the
lymphoproliferative gene appeared, leading to T-cell proliferation
and lymph node swelling, so that the autoreactive lymphocytes are
not eliminated. The symptoms of this model are similar to those of
human SLE, which is characterized by the presence of ANA,
anti-dsDNA, anti-single stranded DNA and other auto-antibodies. The
disadvantage of this model is that it is expensive to create,
therefore it was not used in the present study. In future studies,
this model should be used to confirm the results of the present
study.
B7 molecules (including B7-1 and B7-2) are important
co-stimulatory molecules, which may promote or inhibit T-cell
activation, proliferation and differentiation, depending on the
interactions with the receptors CD28 and CTLA-4 (26-29).
B7-CD28 co-stimulatory signals serve important roles in SLE
occurrence/development. It has previously been indicated that
preferential expression of B7 on B-cells is essential for anti-DNA
autoantibody production in patients with SLE (30). Bijl et al (31) revealed that the expression of CD86
on CD19+ B-cells was increased and associated with
disease activity, B-cell activation and levels of anti-dsDNA in
patients with SLE. The percentage of CD80+ cells in the
large activated B-cell (CD19+) subset of an SLE patient
population was also significantly higher compared with subset
populations from normal controls and patients with allergies
(31). Folzenlogen et al
(32) demonstrated that the B7
protein family may reflect immunologic dysregulation in patients
with autoimmune diseases and may also indicate a state of increased
B-cell activity and hypergammaglobulinemia that occurs during
active SLE. Laurent et al (33) demonstrated that a CD28 blockade
prevented the development of LN in NZB/NZW) F1 mice, which have
typical lupus symptoms and are one of the most recognized animal
models of lupus nephritis. In the present study, flow cytometry
revealed that the expression of activation markers CD80 and CD86 on
B220+ B-cells in the MG group were significantly higher
compared with those in the Mock group.
A number of therapies that block or activate
co-stimulation molecules have been indicated to be efficient for
the treatment of autoimmune diseases. Inhibiting the B7/CD28
co-stimulatory pathways using anti-B7 antibodies has been revealed
to promote corneal allograft survival by inhibiting CD4+
T-cell secretion of interferon-γ, inhibiting further cellular
immune responses and inflammatory reactions (34). Another study demonstrated that
anti-CD80+ or anti-CD86 mAb infusion was effective in
preventing GVHD-associated mortality by inhibiting donor
CD4+ or CD8+ T-cell expansion in mice
(35). Other avenues of
investigation have examined the potential use of CTLA-4 as an
immune checkpoint to inhibit T cell activation (36). CTLA-4 Ig is a soluble protein that
is composed of an extracellular portion of CTLA-4 and an
Fc fragment of IgG1 (37). The essential component of Abatacept
is CTLA-4 Ig, which was approved by the US Food and Drug
Administration in December 2005 for the treatment of rheumatoid
arthritis. Abatacept therapy has been demonstrated to be successful
in murine SLE models and in early human clinical trials (38). In the previous study, it was
observed that the clinical effects of Abatacept provided a possible
benefit in patients with refractory disease, particularly articular
or cutaneous involvement requiring medium- to high-dose
corticosteroids (39). Previous
studies have also indicated that CTLA4-Ig was more effective than
cyclophosphamide in preventing glomerular sclerosis and tubular
damage in murine SLE models (39,40).
In the present study, a mAb targeting the human CD80
molecule, 4E5, was successfully generated. This mAb was observed to
not only recognize human CD80 on a variety of cells but also mouse
splenocyte CD80. DNA sequencing analysis of the mouse CD80
complementary DNA has revealed that the mouse CD80 gene is closely
associated with the human CD80 gene and shares a 63% identity in
the protein-coding region (41).
4E5 was applied to the LN model that was induced by cGVHD and the
present results demonstrated that 4E5 was able to prevent the
development of cGVHD-induced lupus, as well as significantly
inhibit immune-cell activation and autoantibody production. The
onset of proteinuria, renal histopathologic changes and immune
complex deposition in the kidneys were also inhibited. The
expression of CD11b, CD11c and GR1 in spleen cells indicates the
activation of macrophages, dendritic cells and granulocytes in the
spleen respectively (12). In the
present study, the activation of splenocytes (including
macrophages, dendritic cells and granulocytes) in 4E5-treated mice
was also significantly lower compared with that in the MG mice,
suggesting that 4E5 exerted a suppressive effect during the early
stages of immune activation.
Owing to their ability to promote the onset of SLE,
B-cells are now considered to serve a key role in the pathogenesis
of the disease by providing co-stimulatory signals necessary for
T-cell activation, cytokine secretion and immune complex
deposition. Treatments aimed at B-cells have therefore become a
major therapeutic focus (42). In
the present study, the expression of activation markers of B220+
B-cells CD86 and CD80 on cells in the 4E5-treated mice were lower
compared with those on the cells of control mice, which may
indicate a potential use in anti-B-cell therapy for treating SLE.
Since T-B cell activation was inhibited and early immune responses
were reduced in the cGVHD mice by treatment with the 4E5 mAb, the
inhibition of CD80/CD28 signaling may be useful in B-cell-directed
therapies against autoimmune diseases, including SLE. The model
selected in this present study is characterized by the production
of autoantibodies and immune complexes that are able to affect the
majority of human organs (43). The
autoantibodies are produced by the activated B cells, and thus, the
present study focused on whether B cells were activated, while T
cells were not assessed. However, the T-cell pathway is important
and future research should determine its role in SLE. Since
proteinuria is a marker of renal lesions and a criterion for
determining the induction of a successful LN model, the detection
of proteinuria is regarded as an important index for the diagnosis
and prognostic evaluation of LN. However, it is easily affected by
the compliance of patients (44). A
previous study (44) demonstrated
that urinary albumin may be used to monitor renal damage due to the
following: i) Albumin is the major component of urinary protein;
ii) epidemiological data indicated that elevated levels of urinary
albumin are associated with the risk of renal and cardiovascular
disease; iii) according to the latest guidelines, the level of
urinary albumin is one of the bases for the classification of
chronic kidney disease. Overall, the determination of urinary
albumin level is an important indicator for monitoring the
development of nephropathy and should be performed in future
experiments (45,46).
The primary cause of renal damage, LN, in patients
with SLE is overproduction of pathogenic autoantibodies, including
ANA and anti-dsDNA (18). The
present study revealed that the production of ANA and anti-dsDNA in
4E5-treated mice was lower compared with that in the MG mice and
that the extent of kidney damage, reflected by proteinuria, was
also reduced. Although the present study demonstrated that B7-CD28
co-stimulatory signaling was associated with the development of
SLE, inhibiting or weakening the signal may reduce the pathological
damage associated with the disease. Further exploration of the
roles of anti-CD80 antibodies in humans with SLE is required. A
number of therapeutic strategies for SLE are currently under
investigation (47). The novel 4E5
mAb identified in the present study may be useful for immune
therapies for the treatment of SLE, as well as the treatment of
other autoimmune diseases and potentially in organ
transplantation.
Acknowledgements
Not applicable.
Funding
This study was supported by the National Natural
Science Foundation of China (grant nos. 81802341 and 81373236), the
Jiangsu Provincial Medical Youth Talent (grant no. QNRC2016235) and
the Suzhou Administration of Science and Technology (grant no.
SYS201571).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request
Authors' contributions
YHQ and YX designed the experiments and drafted the
manuscript. LJS, YZ, LHH, YYW, TMY and YK performed the experiments
and analyzed the data. STZ analyzed the data. LJS, YX and STZ
revised the manuscript and gave final approval of the version to be
published . All authors read and approved the final manuscript.
Ethics and approval and consent to
participate
The present study was approved by the Ethics
Committee of Soochow University (Suzhou, China; approval no.
201912A341).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Tenbrock K, Juang YT, Kyttaris VC and
Tsokos GC: Altered signal transduction in SLE T cells. Rheumatology
(Oxford). 46:1525–1530. 2007.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Saxena R, Mahajan T and Mohan C: Lupus
nephritis: Current update. Arthritis Res Ther.
13(240)2011.PubMed/NCBI View
Article : Google Scholar
|
|
3
|
Caltik A, Demircin G, Bulbul M, Erdogan O,
Akyuz SG and Arda N: An unusual case of ANA negative systemic lupus
erythematosus presented with vasculitis, long-standing serositis
and full-house nephropathy. Rheumatol Int. 33:219–222.
2013.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Appleman LJ and Boussiotis VA: T cell
anergy and costimulation. Immunol Rev. 192:161–180. 2003.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Lenschow DJ, Walunas TL and Bluestone JA:
CD28/B7 system of T cell costimulation. Annu Rev Immunol.
14:233–258. 1996.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Bretscher P and Cohn M: A theory of
self-nonself discrimination. Science. 169:1042–1049.
1970.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Bour-Jordan H and Blueston JA: CD28
function: A balance of costimulatory and regulatory signals. J Clin
Immunol. 22:1–7. 2002.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Suvas S, Singh V, Sahdev S, Vohra H and
Agrewala JN: Distinct role of CD80 and CD86 in the regulation of
the activation of B cell and B cell lymphoma. J Biol Chem.
277:7766–7775. 2002.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Reynolds J, Tam FW, Chandraker A, Smith J,
Karkar AM, Cross J, Peach R, Sayegh MH and Pusey CD: CD28-B7
blockade prevents the development of experimental autoimmune
glomerulonephritis. J Clin Invest. 105:643–651. 2000.PubMed/NCBI View
Article : Google Scholar
|
|
10
|
Lewis RM, Armstrong MY, Andre-Schwartz J,
Muftuoglu A, Beldotti L and Schwartz RS: Chronic allogeneic
disease. I. Development of glomerulonephritis. J Exp Med.
128:653–679. 1968.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Sekine H, Watanabe H and Gilkeson GS:
Enrichment of anti-glomerular antigen antibody-producing cells in
the kidneys of MRL/MpJ-Fas(lpr) mice. J Immunol. 172:3913–3921.
2004.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Shi Q, Gao ZY, Xie F, Wang LF, Gu YP, Yang
TJ, Huang L, Qian QH and Qiu YH: A novel monoclonal antibody
against human CD80 and its immune protection in a mouse lupus-like
disease. Int J Immunopathol Pharmacol. 24:583–593. 2011.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Zhang GB, Zhou H, Chen YJ, Ge Y, Xie F,
Shi Q, Ma HB, Fei M and Zhang XG: Characterization and application
of two novel monoclonal antibodies against 2IgB7-H3: Expression
analysis of 2IgB7-H3 on dendritic cells and tumor cells. Tissue
Antigens. 66:83–92. 2005.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Sun WP, Wang FM, Xie F, Wang GQ, Sun J, Yu
GH, Qiu YH and Zhang XG: A novel anti-human syndecan-1 (CD138)
monoclonal antibody 4B3: Characterization and application. Cell Mol
Immunol. 4:209–214. 2007.PubMed/NCBI
|
|
15
|
Kints JP, Manouvriez P and Bazin H: Rat
monoclonal antibodies. VII. Enhancement of ascites production and
yield of monoclonal antibodies in rats following pretreatment with
pristane and Freund's adjuvant. J Immunol Methods. 119:241–245.
1989.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Kronick MN: The use of phycobiliproteins
as fluorescent labels in immunoassay. J Immunol Methods. 92:1–13.
1986.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Han L, Shen L, Zhu Y and Qiu Y: A
monoclonal antibody against CD86 and its protection in a murine
lupus nephritis model of chronic graft-versus-host disease.
Immunopharmacol Immunotoxicol. 39:285–291. 2017.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Bruns A, Blass S, Hausdorf G, Burmester GR
and Hiepe F: Nucleosomes are major T and B cell autoantigens in
systemic lupus erythematosus. Arthritis Rheum. 43:2307–2315.
2000.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Murphy WJ: Revisiting graft-versus-host
disease models of autoimmunity: New insights in immune regulatory
processes. J Clin Invest. 106:745–747. 2000.PubMed/NCBI View
Article : Google Scholar
|
|
20
|
Howie JB and Helyer BJ: The immunology and
pathology of NZB mice. Adv Immunol. 9:215–266. 1968.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Ka SM, Rifai A, Chen JH, Cheng CW, Shui
HA, Lee HS, Lin YF, Hsu LF and Chen A: Glomerular crescent-related
biomarkers in a murine model of chronic graft versus host disease.
Nephrol Dial Transplant. 21:288–298. 2006.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Foster AD, Soloviova K, Puliaeva I,
Puliaiev M, Puliaev R, Finkelman F and Via CS: Donor CD8 T cells
and IFN-gamma are critical for sex-based differences in donor CD4 T
cell engraftment and lupus-like phenotype in short-term chronic
graft-versus-host disease mice. J Immunol. 186:6238–6254.
2011.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Rus V, Svetic A, Nguyen P, Gause WC and
Via CS: Kinetics of Th1 and Th2 cytokine production during the
early course of acute and chronic murine graft-versus-host disease.
Regulatory role of donor CD8+ T cells. J Immunol. 155:2396–2406.
1995.PubMed/NCBI
|
|
24
|
Bergijk EC, Munaut C, Baelde JJ, Prins F,
Foidart JM, Hoedemaeker PJ and Bruijn JA: A histologic study of the
extracellular matrix during the development of glomerulosclerosis
in murine chronic graft-versus-host disease. Am J Pathol.
140:1147–1156. 1992.PubMed/NCBI
|
|
25
|
Dhaher YY, Chan K, Greenstein BD, de
Fougerolles Nunn E, Khamashta MA and Hughes GR: Impaired estrogen
priming of progesterone receptors in uterus of MRL/MP-lpr/lpr mice,
a model of systemic lupus erythematosus (SLE). Int J
Immunopharmacol. 22:537–545. 2000.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Kuroda Y, Akaogi J, Nacionales DC, Wasdo
SC, Szabo NJ, Reeves WH and Satoh M: Distinctive patterns of
autoimmune response induced by different types of mineral oil.
Toxicol Sci. 78:222–228. 2004.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Kim J, Park K, Kim HJ, Kim J, Kim HA, Jung
D, Kim HJ, Choi HJ, Choi SY, Seo KW, et al: Breaking of CD8+ T cell
tolerance through in vivo ligation of CD40 results in inhibition of
chronic graft-versus-host disease and complete donor cell
engraftment. J Immunol. 181:7380–7389. 2008.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Kothlow S, Morgenroth I, Tregaskes CA,
Kaspers B and Young JR: CD40 ligand supports the long-term
maintenance and differentiation of chicken B cells in culture. Dev
Comp Immunol. 32:1015–1026. 2008.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Leitner J, Grabmeier-Pfistershammer K and
Steinberger P: Receptors and ligands implicated in human T cell
costimulatory processes. Immunol Lett. 128:89–97. 2010.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Nagafuchi H, Shimoyama Y, Kashiwakura J,
Takeno M, Sakane T and Suzuki N: Preferential expression of B7.2
(CD86), but not B7.1 (CD80), on B cells induced by CD40/CD40L
interaction is essential for anti-DNA autoantibody production in
patients with systemic lupus erythematosus. Clin Exp Rheumatol.
21:71–77. 2003.PubMed/NCBI
|
|
31
|
Bijl M, Horst G, Limburg PC and Kallenberg
CG: Expression of costimulatory molecules on peripheral blood
lymphocytes of patients with systemic lupus erythematosus. Ann
Rheum Dis. 60:523–526. 2001.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Folzenlogen D, Hofer MF, Leung DY, Freed
JH and Newell MK: Analysis of CD80 and CD86 expression on
peripheral blood B lymphocytes reveals increased expression of CD86
in lupus patients. Clin Immunol Immunopathol. 83:199–204.
1997.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Laurent L, Le Fur A, Le Bloas RL, Néel M,
Mary C, Moreau A, Poirier N, Vanhove B and Fakhouri F: Prevention
of lupus nephritis development in NZB/NZW mice by selective
blockade of CD28. Eur J Immunol. 47:1368–1376. 2017.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Asai T, Choi BK, Kwon PM, Kim WY, Kim JD,
Vinay DS, Gebhardt BM and Kwon BS: Blockade of the 4-1BB
(CD137)/4-1BBL and/or CD28/CD80/CD86 costimulatory pathways
promotes corneal allograft survival in mice. Immunology.
121:349–358. 2007.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Blazar BR, Sharpe AH, Taylor PA,
Panoskaltsis-Mortari A, Gray GS, Korngold R and Vallera DA:
Infusion of anti-B7.1 (CD80) and anti-B7.2 (CD86) monoclonal
antibodies inhibits murine graft-versus-host disease lethality in
part via direct effects on CD4+ and CD8+ T cells. J Immunol.
157:3250–3259. 1996.PubMed/NCBI
|
|
36
|
Brunner-Weinzierl MC, Hoff H and Burmester
GR: Multiple functions for CD28 and cytotoxic T lymphocyte
antigen-4 during different phases of T cell responses: Implications
for arthritis and autoimmune diseases. Arthritis Res Ther. 6:45–54.
2004.PubMed/NCBI View
Article : Google Scholar
|
|
37
|
Finck BK, Linsley PS and Wofsy D:
Treatment of murine lupus with CTLA-Ig. Science. 265:1225–1227.
1994.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Furie R, Nicholls K, Cheng TT, Houssiau F,
Burgos-Vargas R, Chen SL, Hillson JL, Meadows-Shropshire S,
Kinaszczuk M and Merrill JT: Efficacy and safety of abatacept in
lupus nephritis: A twelve-month, randomized, double-blind study.
Arthritis Rheumatol. 66:379–389. 2014.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Danion F, Rosine N, Belkhir R, Gottenberg
JE, Hachulla E, Chatelus E, Pugnet G, Pers YM, Mariette X, Sibilia
J, et al: Efficacy of abatacept in systemic lupus erythematosus: A
retrospective analysis of 11 patients with refractory disease.
Lupus. 25:1440–1447. 2016.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Cunnane G, Chan OT, Cassafer G, Brindis S,
Kaufman E, Yen TSB and Daikh DI: Prevention of renal damage in
murine lupus nephritis by CTLA-4Ig and cyclophosphamide. Arthritis
Rheum. 50:1539–1548. 2004.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Freeman GJ, Gray GS, Gimmi CD, Lombard DB,
Zhou LJ, White M, Fingeroth JD, Gribben JG and Nadler LM:
Structure, expression, and T cell costimulatory activity of the
murine homologue of the human B lymphocyte activation antigen B7. J
Exp Med. 174:625–631. 1991.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Tieng AT and Peeva E: B-cell-directed
therapies in systemic lupus erythematosus. Semin Arthritis Rheum.
38:218–227. 2008.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Mills JA: Systemic lupus erythematosus. N
Engl J Med. 330:1871–1879. 1994.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Satoh M, Kumar A, Kanwar YS and Reeves WH:
Anti-nuclear antibody production and immune-complex
glomerulonephritis in BALB/c mice treated with pristane. Proc Natl
Acad Sci USA. 92:10934–10938. 1995.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Nagata M, Ninomiya T, Kiyohara Y, Murakami
Y, Irie F, Sairenchi T, Miura K, Okamura T and Ueshima H:
EPOCH-JAPAN Research Group. Prediction of cardiovascular disease
mortality by proteinuria and reduced kidney function: Pooled
analysis of 39,000 individuals from 7 cohort studies in Japan. Am J
Epidemiol. 178:1–11. 2013.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Currie G and Delles C: Proteinuria and its
relation to cardiovascular disease. Int J Nephrol Renovasc Dis.
7:13–24. 2013.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Holdsworth SR, Gan PY and Kitching AR:
Biologics for the treatment of autoimmune renal diseases. Nat Rev
Nephrol. 12:217–231. 2016.PubMed/NCBI View Article : Google Scholar
|