Introduction
One of the most common congenital cardiac
abnormalities is coarctation of the aorta (CoA) (1). Around 85% of CoA patients also have a
bicuspid aortic valve (BAV) (2).
Following CoA repair, patients are at increased risk of developing
cardiovascular complications including hypertension, impaired left
ventricular function, aortic aneurysms and aortic dissection
(3-5).
Several studies have demonstrated that the risks are greater in CoA
patients with BAV compared to CoA patients with the normal
tricuspid valve (TAV) (6-8).
The co-existence of BAV and CoA is known to alter aortic blood flow
haemodynamics, and this may underlie the increased susceptibility
of CoA patients with BAV to cardiovascular complications (9,10).
A recent study by our group was the first to
investigate differences in the vascular proteome of CoA patients
with and without BAV (11). This
study focused on neonatal patients (less than 3 weeks old) and
demonstrated that the presence of BAV in neonatal CoA patients is
associated with altered expression of proteins involved in elastin
fibre formation and oxidative stress. In older CoA patients, there
will have been more time for aortic remodelling to occur in
response to the altered blood flow haemodynamics, and this will
likely effect protein expression. Therefore, molecular changes in
the coarctation area will be influenced not just by the valve and
blood flow haemodynamics, but also the effects of growth. The
current study therefore compares the proteome of CoA patients with
and without BAV in paediatric patients older than one month, thus
providing a unique insight into how the proteomic changes
associated with BAV evolve in an older group of CoA patients.
Materials and methods
Patients and sample collection
Tissue was collected from paediatric patients older
than one month undergoing congenital surgery including repair of an
aortic coarctation. Tissue was collected just proximal to the
coarctation site. The study was conducted in accordance with the
declaration of Helsinki, and the protocol was approved by the North
Somerset and South Bristol Research Ethics Committee (Health
Research Authority, Whitefriars, Level 3 Block B, Lewins Mead,
Bristol, BS1 2NT, REC 07/H0106/172). Full informed consent was
obtained from parents prior to admission for operation. The tissue
was snap frozen in liquid nitrogen before being stored at -80˚C
(BAV: n=6, patient age 1.9±1.7 years (mean ± SEM). TAV: n=4,
patient age 1.7±1.5 years).
Sample preparation
Proteins were extracted in
radio-immuno-precipitation assay buffer (RIPA; 1% Nonidet P-40,
0.5% sodium deoxycholate, 0.1% SDS, in PBS with phosphatase and
protease inhibitors), and quantified using Bradford's assay.
Aliquots of 100 µg were digested (2.5 µg trypsin, 37˚C, overnight),
labelled with Tandem Mass Tag (TMT) 10Plex reagents (Thermo Fisher
Scientific, Loughborough, UK) and the labelled samples pooled. For
total proteome analysis, aliquots of 50 µg of the pooled sample
were dried, re-suspended in buffer A (20 mM ammonium hydroxide, pH
10) and fractionated by high pH reversed-phase (RP) chromatography
(UltiMate 3000 liquid chromatography system (Thermo Fisher
Scientific) with XBridge BEH C18 Column (130 Å, 3.5 µm, 2.1x150 mm,
Waters, UK). The samples were loaded in buffer A and peptides
eluted with an increasing gradient of buffer B (20 mM ammonium
hydroxide in acetonitrile, pH 10, 0-95% over 60 min). The resulting
fractions were dried and re-suspended in 1% (v/v) formic acid. The
remainder of the TMT-labelled pooled sample was dried and enriched
using a TiO2 based phosphopeptide enrichment protocol
(Pierce), before further drying and re-suspension in 1% (v/v)
formic acid.
Nano-LC mass spectrometry
Mass spectrometry was performed using an Ultimate
3000 nano-HPLC system in line with an Orbitrap Fusion Tribrid mass
spectrometer (Thermo Scientific). The samples were injected onto an
Acclaim PepMap C18 nano-trap column (Thermo Scientific), washed
(0.5% v/v acetonitrile, 0.1% v/v formic acid) and resolved on a 250
mm x 75 µm Acclaim PepMap C18 reverse phase analytical column
(Thermo Scientific) over an organic gradient (150 min, flow rate
300 nl/min, solvent A: 0.1% formic acid, solvent B: Aqueous 80%
acetonitrile in 0.1% formic acid, seven gradient segments: 1-6% B
over 1 min, 6-15% B over 58 min, 15-32% B over 58 min, 32-40% B
over 5 min, 40-90% B over 1 min, held at 90% B for 6 min and then
reduced to 1% B over 1 min). Nano-electrospray ionization was used
to ionize the peptides (2.0 kV, stainless steel emitter internal
diameter 30 µm (Thermo Scientific), capillary temperature
275˚C).
Spectra were acquired using an Orbitrap Fusion
Tribrid mass spectrometer with Xcalibur 3.0 software (Thermo
Scientific), operated using an SPS-MS3 workflow and data-dependent
acquisition mode. For FTMS1 spectra a resolution of 120,000 was
used, alongside an automatic gain control (AGC) target of 200,000
and a maximum injection time of 50 ms. Precursors were filtered
with an intensity threshold of 5000, with monoisotopic peak
determination set to peptide and to include charge states 2-7.
Previously interrogated precursors were excluded with a dynamic
window (60s +/-10 ppm). The MS2 precursors were isolated with a
quadrupole mass filter (width of 1.2 m/z). ITMS2 spectra were
collected (AGC target 10000, max injection time 70ms, CID collision
energy 35%). FTMS3 analysis was then performed (resolution 50,000,
AGC target 50,000, max injection time 105 ms). Fragmentation of
precursors was achieved using high-energy collision dissociation at
normalised collision energy of 60%. Synchronous Precursor Selection
(SPS) was enabled to include up to five MS2 fragment ions in the
FTMS3 scan.
Data processing and analysis
Processing and quantification of the raw data files
was performed using Proteome Discoverer Software (Thermo
Scientific, version 1.4). Peptide sequences were searched against
the Uniprot human database (SEQUEST algorithm, peptide precursor
mass tolerance 10 ppm, MS/MS tolerance 0.6Da). Oxidation of
methionine (+15.9949) was included as a variable modification, and
both carbamido-methylation of cysteine (+57.0214) and the addition
of the TMT mass tag (+229.163) to peptide N-termini and lysine were
included as fixed modifications. Phosphorylation of serine,
threonine and tyrosine (+79.966) were also included as variable
modifications in the phosphoproteomic analysis. Searches were
performed with full tryptic digestion, allowing a maximum of one
missed cleavage. The reverse database search option was enabled,
and all the data filtered to satisfy false discovery rate of
5%.
Proteins or phosphoproteins with more than one
missing value per group were excluded from the analysis (12), as were putative uncharacterized
proteins and proteins with accession numbers representing cDNA with
weak similarity. Values are presented as a ratio to the internal
standard (a pool of all samples) and represent the median of the
measured peptide(s) for each protein. Fold change (BAV/TAV) ratios
were calculated, and log2 (fold-change) was plotted
against -log10 (P-value) on a volcano plot. A change in
protein expression greater than 1.3x or less than -1.3x and with
P<0.05 (Student's t-test) was considered significant. These cut
offs were chosen in accordance with other similar studies, for
example (13). A heatmap was
plotted using R software (version 3.5.1). QuickGo software was used
to analyse gene ontology (GO) enrichment. Differentially expressed
proteins and phosphoproteins were inputted into Ingenuity Pathway
Analysis software (IPA version 46901286, Qiagen, Aarhus, Denmark)
to determine significantly enriched canonical pathways (P-value of
overlap calculated by Fisher's exact test right tailed).
Results
The age range of patients in the study was from one
month to 10.5 years. The average age was not different
significantly between groups (TAV; 1.7±1.5 years, BAV; 1.9±1.7
years, P=0.92).
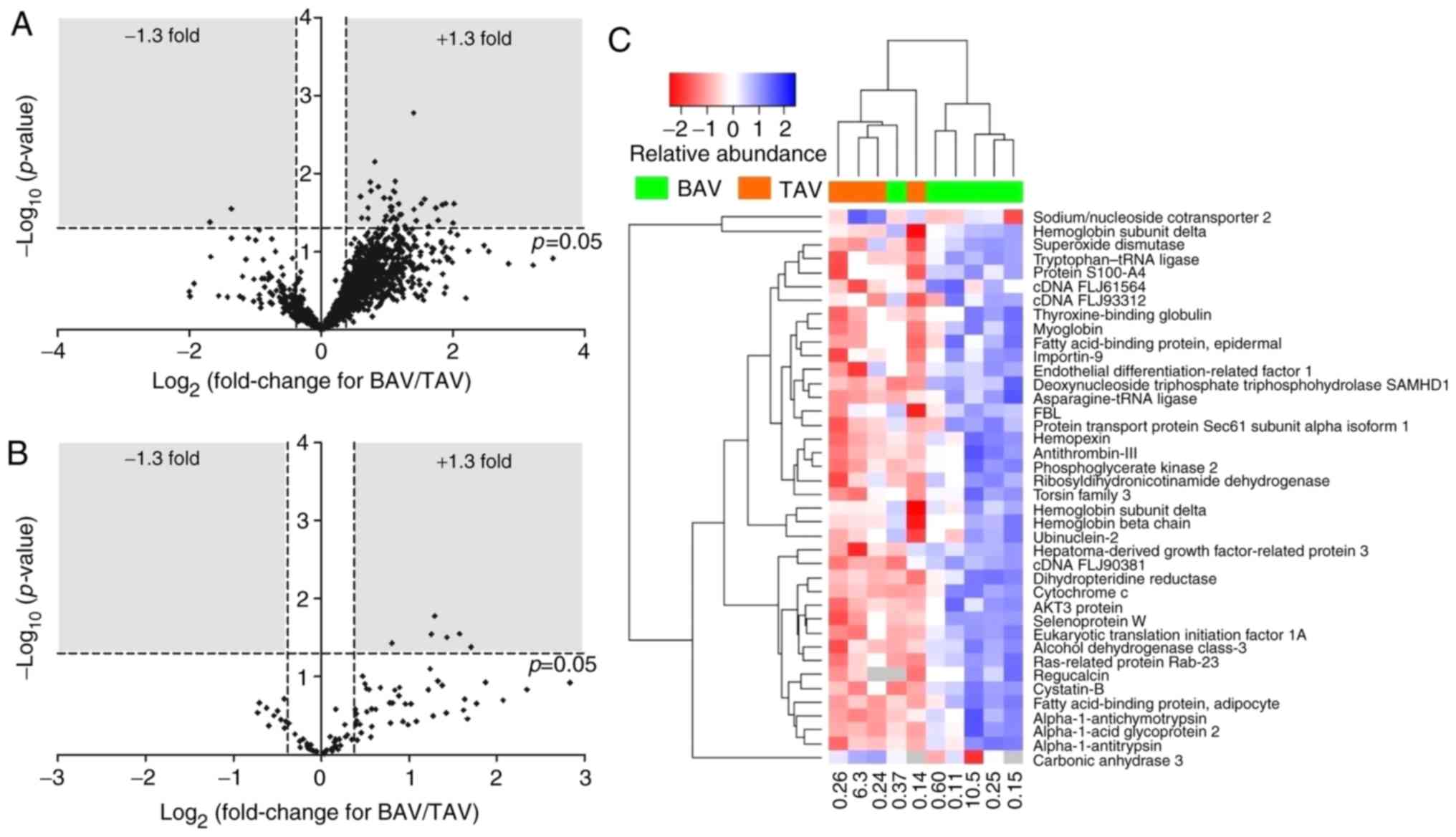
A total of 1811 protein accession numbers were
detected, of which 40 were significantly differentially expressed
between BAV and TAV patients (Fig.
1A and C, Table I); 38 proteins upregulated and two
proteins downregulated in BAV patients compared to TAV patients. A
total of 76 phosphorylated proteins were identified, some with
multiple phosphorylation sites, resulting in 92 phosphorylation
site matches. 6 phosphoproteins demonstrated significantly altered
expression in BAV patients compared to TAV patients; all 6
demonstrated increased phosphorylation in BAV patients (Fig. 1B, Table
II).
 | Table IProteins differentially (fold
increase >1.3 or <0.769) and significantly (P<0.05)
expressed in coarcted aorta from paediatric BAV patients compared
to paediatric TAV patients. - : gene ID not applicable. |
Table I
Proteins differentially (fold
increase >1.3 or <0.769) and significantly (P<0.05)
expressed in coarcted aorta from paediatric BAV patients compared
to paediatric TAV patients. - : gene ID not applicable.
| | Mean | SEM | |
|---|
| Accession
number | Gene ID | Description | BAV | TAV | BAV | TAV | Fold change
(BAV/TAV) | P-value | log2
(fold-change) | -log10
(P-value) |
|---|
| P15090 | FABP4 | Fatty acid-binding
protein, adipocyte | 1.51 | 0.37 | 0.33 | 0.04 | 4.05 | 0.024 | 2.02 | 1.62 |
| P19652 | ORM2 | α-1-acid
glycoprotein 2 | 1.45 | 0.36 | 0.36 | 0.02 | 4.01 | 0.043 | 2.00 | 1.37 |
| A0A024R943 | TOR3A | Torsin family 3,
member A, isoform CRA_b | 2.04 | 0.55 | 0.42 | 0.14 | 3.72 | 0.024 | 1.89 | 1.62 |
| Q6VFQ6 | HBB | Hemoglobin beta
chain (Fragment) | 2.53 | 0.71 | 0.52 | 0.19 | 3.58 | 0.025 | 1.84 | 1.60 |
| C9J6Y5 | UBN2 | Ubinuclein-2
(Fragment) | 3.78 | 1.10 | 0.84 | 0.35 | 3.43 | 0.039 | 1.78 | 1.41 |
| P01009 | SERPINA1 |
α-1-antitrypsin | 1.23 | 0.37 | 0.25 | 0.05 | 3.34 | 0.028 | 1.74 | 1.56 |
| Q15493 | RGN | Regucalcin | 1.14 | 0.37 | 0.23 | 0.07 | 3.10 | 0.047 | 1.63 | 1.33 |
| P02042 | HBD | Hemoglobin subunit
delta | 3.13 | 1.05 | 0.55 | 0.30 | 2.98 | 0.021 | 1.57 | 1.67 |
| P02790 | HPX | Hemopexin | 2.03 | 0.71 | 0.38 | 0.09 | 2.87 | 0.024 | 1.52 | 1.62 |
| P01011 | SERPINA3 |
α-1-antichymotrypsin | 1.31 | 0.48 | 0.27 | 0.07 | 2.72 | 0.040 | 1.44 | 1.40 |
| Q9ULC3 | RAB23 | Ras-related protein
Rab-23 | 1.29 | 0.49 | 0.23 | 0.10 | 2.66 | 0.029 | 1.41 | 1.54 |
| E9PEW8 | HBD | Hemoglobin subunit
delta (Fragment) | 7.99 | 3.02 | 0.72 | 0.73 | 2.65 | 0.002 | 1.41 | 2.78 |
| P01008 | SERPINC1 |
Antithrombin-III | 1.65 | 0.69 | 0.33 | 0.06 | 2.41 | 0.047 | 1.27 | 1.33 |
| P04080 | CSTB | Cystatin-B | 0.96 | 0.41 | 0.17 | 0.06 | 2.32 | 0.033 | 1.22 | 1.48 |
| P09417 | QDPR | Dihydropteridine
reductase | 1.36 | 0.60 | 0.24 | 0.06 | 2.29 | 0.034 | 1.19 | 1.47 |
| Q56A86 | AKT3 | AKT3 protein
(Fragment) | 1.21 | 0.54 | 0.19 | 0.06 | 2.26 | 0.024 | 1.18 | 1.61 |
| B2R773 | - | cDNA, FLJ93312,
highly similar to Homo sapiens adipose most abundant gene
transcript 1 (ADIPOQ), mRNA | 1.92 | 0.87 | 0.32 | 0.20 | 2.22 | 0.038 | 1.15 | 1.42 |
| Q5TD07 | NQO2 |
Ribosyldihydronicotinamide dehydrogenase
[quinone] | 1.54 | 0.70 | 0.25 | 0.21 | 2.20 | 0.047 | 1.14 | 1.33 |
| Q96P70 | IPO9 | Importin-9 | 1.53 | 0.70 | 0.19 | 0.14 | 2.18 | 0.013 | 1.12 | 1.90 |
| O14602 | EIF1AY | Eukaryotic
translation initiation factor 1A, Y-chromosomal | 1.18 | 0.55 | 0.16 | 0.08 | 2.17 | 0.017 | 1.12 | 1.76 |
| A0A024R0V8 | SEPW1 | Selenoprotein W, 1
(Fragment) | 1.37 | 0.64 | 0.18 | 0.11 | 2.14 | 0.017 | 1.10 | 1.78 |
| P05543 | SERPINA7 | Thyroxine-binding
globulin | 1.79 | 0.84 | 0.29 | 0.14 | 2.14 | 0.036 | 1.10 | 1.44 |
| B4E367 | - | cDNA FLJ61564,
highly similar to Plexin domain-containing protein 2 | 2.18 | 1.03 | 0.33 | 0.13 | 2.11 | 0.027 | 1.08 | 1.57 |
| B3KQF5 | - | cDNA FLJ90381 fis,
clone NT2RP2005035, highly similar to Calumenin | 0.96 | 0.46 | 0.14 | 0.09 | 2.09 | 0.030 | 1.07 | 1.53 |
| C9JFR7 | CYCS | Cytochrome c
(Fragment) | 1.28 | 0.62 | 0.19 | 0.02 | 2.08 | 0.027 | 1.05 | 1.57 |
| P11766 | ADH5 | Alcohol
dehydrogenase class-3 | 1.12 | 0.54 | 0.15 | 0.07 | 2.06 | 0.021 | 1.04 | 1.69 |
| P07205 | PGK2 | Phosphoglycerate
kinase 2 | 1.52 | 0.74 | 0.25 | 0.09 | 2.06 | 0.040 | 1.04 | 1.40 |
| Q01469 | FABP5 | Fatty acid-binding
protein, epidermal | 1.75 | 0.88 | 0.26 | 0.14 | 1.98 | 0.035 | 0.99 | 1.46 |
| Q9Y3E1 | HDGFRP3 | Hepatoma-derived
growth factor-related protein 3 | 1.02 | 0.52 | 0.13 | 0.16 | 1.95 | 0.040 | 0.96 | 1.40 |
| P02144 | MB | Myoglobin | 1.67 | 0.86 | 0.21 | 0.14 | 1.94 | 0.021 | 0.96 | 1.69 |
| P26447 | S100A4 | Protein
S100-A4 | 2.36 | 1.30 | 0.21 | 0.29 | 1.82 | 0.015 | 0.86 | 1.83 |
| P61619 | SEC61A1 | Protein transport
protein Sec61 subunit α isoform 1 | 1.16 | 0.66 | 0.11 | 0.06 | 1.75 | 0.007 | 0.81 | 2.15 |
| Q96BS4 | FBL | FBL protein
(Fragment) | 1.25 | 0.74 | 0.09 | 0.16 | 1.69 | 0.017 | 0.75 | 1.78 |
| P23381 | WARS | Tryptophan-tRNA
ligase, cytoplasmic | 1.74 | 1.04 | 0.15 | 0.15 | 1.68 | 0.013 | 0.74 | 1.89 |
| O43776 | NARS | Asparagine-tRNA
ligase, cytoplasmic | 1.18 | 0.72 | 0.15 | 0.05 | 1.64 | 0.037 | 0.71 | 1.43 |
| Q9Y3Z3 | SAMHD1 | Deoxynucleoside
triphosphate triphosphohydrolase SAMHD1 | 1.27 | 0.78 | 0.15 | 0.04 | 1.64 | 0.027 | 0.71 | 1.56 |
| P00441 | SOD1 | Superoxide
dismutase [Cu-Zn] | 1.92 | 1.26 | 0.15 | 0.24 | 1.53 | 0.037 | 0.62 | 1.43 |
| O60869 | EDF1 | Endothelial
differentiation-related factor 1 | 1.30 | 0.86 | 0.07 | 0.16 | 1.51 | 0.020 | 0.60 | 1.71 |
| P07451 | CA3 | Carbonic anhydrase
3 | 1.47 | 3.78 | 0.45 | 0.73 | 0.39 | 0.028 | -1.37 | 1.55 |
| O43868 | SLC28A2 | Sodium/nucleoside
cotransporter 2 | 5.72 | 18.47 | 1.44 | 6.25 | 0.31 | 0.042 | -1.69 | 1.38 |
| Table IIPhosphoproteins differentially (fold
increase >1.3 or <0.769) and significantly (P<0.05)
expressed in coarcted aorta from paediatric BAV patients compared
to paediatric TAV patients. - : gene ID not applicable. |
Table II
Phosphoproteins differentially (fold
increase >1.3 or <0.769) and significantly (P<0.05)
expressed in coarcted aorta from paediatric BAV patients compared
to paediatric TAV patients. - : gene ID not applicable.
| | Mean | SEM | |
|---|
| Accession
number | Gene ID | Description | Modification | BAV | TAV | BAV | TAV | Fold-change
(BAV/TAV) | P-value | log2
(fold change) | -log10
(P-value) |
|---|
| B7Z556 | - | cDNA FLJ56822,
highly similar to α-2-HS-glycoprotein (AHSG) | S7(Phospho) | 0.88 | 0.27 | 0.20 | 0.04 | 3.27 | 0.042 | 1.709 | 1.381 |
| K7EKZ3 | VPS4B | Vacuolar protein
sorting-associated protein 4B | S4(Phospho) | 2.02 | 0.68 | 0.39 | 0.18 | 2.98 | 0.028 | 1.577 | 1.550 |
| Q6PK50 | HSP90AB1 | HSP90AB1 protein
(Fragment) | S6(Phospho) | 0.95 | 0.35 | 0.17 | 0.11 | 2.70 | 0.031 | 1.433 | 1.504 |
| Q09666 | AHNAK | Neuroblast
differentiation-associated protein AHNAK | S8(Phospho) | 1.05 | 0.43 | 0.15 | 0.11 | 2.45 | 0.017 | 1.292 | 1.777 |
| Q53TN4 | CYBRD1 | Cytochrome b
reductase 1 | S2(Phospho) | 1.88 | 0.79 | 0.31 | 0.18 | 2.39 | 0.028 | 1.255 | 1.547 |
| B1AKZ5 | PEA15 | Astrocytic
phosphoprotein PEA-15 | S3(Phospho) | 1.30 | 0.74 | 0.14 | 0.17 | 1.75 | 0.037 | 0.807 | 1.428 |
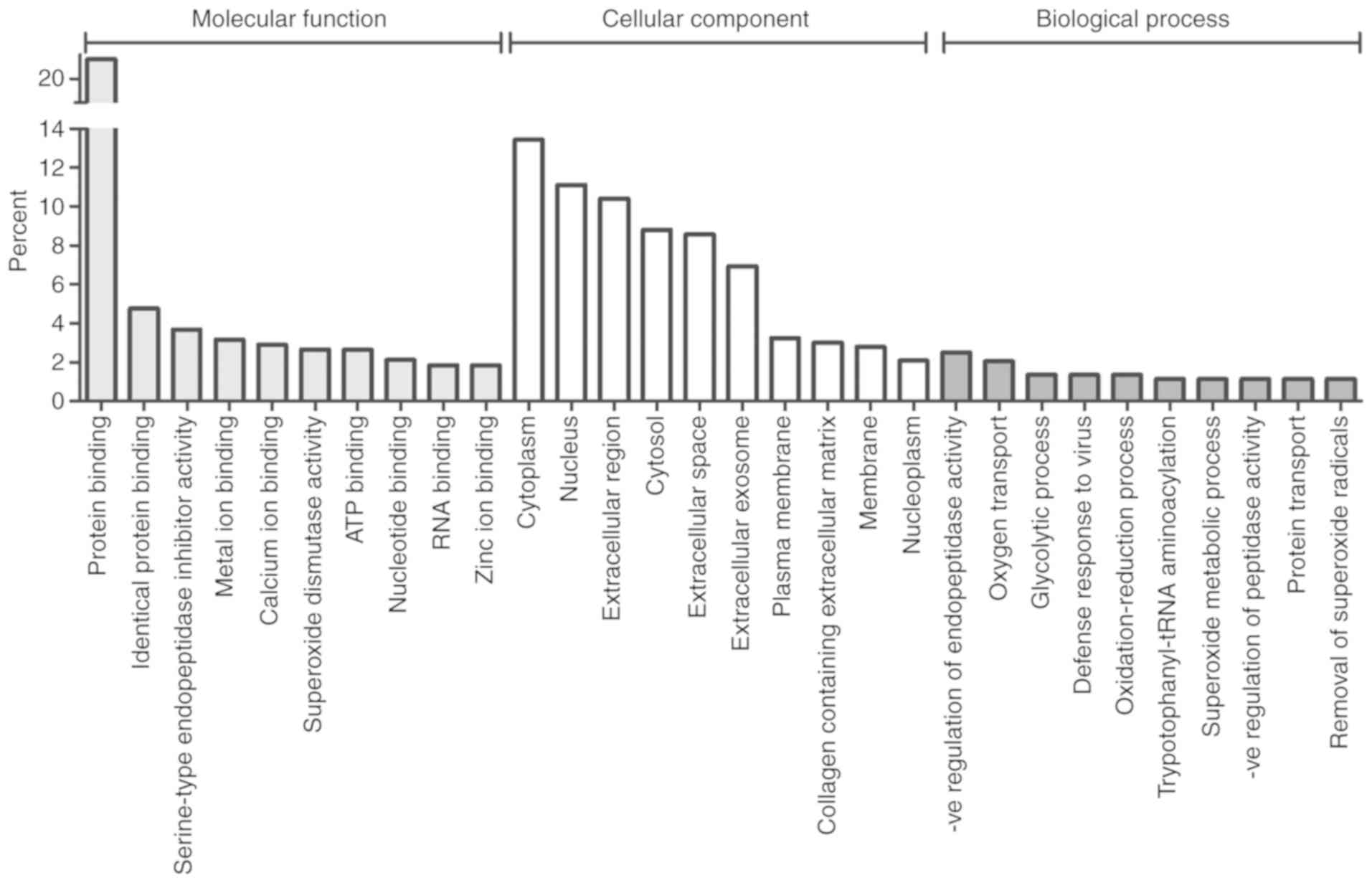
Gene Ontology (GO) analysis was used to functionally
classify the GO annotations associated with the differentially
expressed proteins under the three main categories of GO analysis
(molecular function, biological process and cellular component;
Fig. 2). Numerous molecular
functions relating to protein and ion binding were present in the
top ten molecular functions, however endopeptidase and superoxide
dismutase activities were also featured.
The top 20 significantly enhanced canonical pathways
for protein expression are shown in Table III. The most significantly
enhanced pathway was the acute phase response, a systemic response
to inflammation. IPA analysis predicted activation of the acute
phase response in BAV patients (z-score=2). Several other of the
top 20 canonical pathways also relate to inflammation, including
EIF2 signalling and macrophage production of IL12 and reactive
oxygen species (ROS). Superoxide radical degradation was also
highlighted as a significantly enriched canonical pathway. Finally,
a number of canonical pathways involved in apoptosis, including Myc
mediated apoptosis signalling, ceramide signalling and lymphotoxin
β receptor signalling, were all significantly enriched. Sixteen
canonical pathways were found to be significantly enriched from the
phosphoproteomic data (Table IV),
including two pathways relating to inflammation, namely the acute
phase response and Th17 activation.
| Table IIITop twenty significant canonical
pathways with enriched protein expression. IPA: Ingenuity Pathway
Analysis. |
Table III
Top twenty significant canonical
pathways with enriched protein expression. IPA: Ingenuity Pathway
Analysis.
| IPA canonical
pathway | P-value | Molecule(s) |
|---|
| Acute Phase
Response Signalling |
1.52x10-5 | HPX, AKT3, ORM2,
SERPINA1, SERPINA3 |
| FXR/RXR
Activation |
7.10x10-5 | HPX, AKT3, ORM2,
SERPINA1 |
| Amyotrophic Lateral
Sclerosis Signalling |
1.04x10-3 | AKT3, CYCS,
SOD1 |
| LXR/RXR
Activation |
1.27x10-3 | HPX, ORM2,
SERPINA1 |
| Coagulation
System |
1.74x10-3 | SERPINC1,
SERPINA1 |
| Iron homeostasis
signalling pathway |
1.81x10-3 | HPX, HBD, HBB |
| tRNA Charging |
2.16x10-3 | NARS, WARS |
| IL-12 Signalling
and Production in Macrophages |
2.26x10-3 | ORM2, AKT3,
SERPINA1 |
| Formaldehyde
Oxidation II (Glutathione-dependent) |
3.53x10-3 | ADH5 |
| Docosahexaenoic
Acid (DHA) Signalling |
4.10x10-3 | AKT3, CYCS |
| Production of
Nitric Oxide and Reactive Oxygen Species in Macrophages |
5.55x10-3 | ORM2, AKT3,
SERPINA1 |
| Lymphotoxin β
Receptor Signalling |
6.61x10-3 | AKT3, CYCS |
| Phenylalanine
Degradation I (Aerobic) |
7.04x10-3 | QDPR |
| EIF2
Signalling |
7.90x10-3 | WARS, AKT3,
EIF1AY |
| Myc Mediated
Apoptosis Signalling |
8.79x10-3 | AKT3, CYCS |
| Small Cell Lung
Cancer Signalling |
1.03x10-2 | AKT3, CYCS |
| Superoxide Radicals
Degradation |
1.40x10-2 | SOD1 |
| Ceramide
Signalling |
1.45x10-2 | AKT3, CYCS |
| VEGF
Signalling |
1.73x10-2 | AKT3, EIF1AY |
| Glucose and
Glucose-1-phosphate Degradation |
1.92x10-2 | RGN |
| Table IVSignificant canonical pathways with
enriched phosphoprotein expression. IPA: Ingenuity Pathway
Analysis. |
Table IV
Significant canonical pathways with
enriched phosphoprotein expression. IPA: Ingenuity Pathway
Analysis.
| IPA canonical
pathway | P-value | Molecule |
|---|
| Mitotic Roles of
Polo-Like Kinase |
1.77x10-2 | HSP90AB1 |
| Hypoxia Signalling
in the Cardiovascular System |
1.99x10-2 | HSP90AB1 |
| Th17 Activation
Pathway |
2.44x10-2 | HSP90AB1 |
| Prostate Cancer
Signalling |
2.47x10-2 | HSP90AB1 |
| Neuregulin
Signalling |
2.57x10-2 | HSP90AB1 |
| Nitric Oxide
Signalling in the Cardiovascular System |
2.68x10-2 | HSP90AB1 |
| PPAR
Signalling |
2.78x10-2 | HSP90AB1 |
| Telomerase
Signalling |
2.89x10-2 | HSP90AB1 |
| LXR/RXR
Activation |
3.23x10-2 | AHSG |
| FXR/RXR
Activation |
3.36x10-2 | AHSG |
| PI3K/AKT
Signalling |
3.52x10-2 | HSP90AB1 |
| Iron homeostasis
signalling pathway |
3.65x10-2 | CYBRD1 |
| Aryl Hydrocarbon
Receptor Signalling |
3.81x10-2 | HSP90AB1 |
| Aldosterone
Signalling in Epithelial Cells |
4.23x10-2 | HSP90AB1 |
| eNOS
Signalling |
4.26x10-2 | HSP90AB1 |
| Acute Phase
Response Signalling |
4.75x10-2 | AHSG |
Discussion
Our previous study (11) demonstrated that aortic tissue from
neonatal CoA patients with BAV have proteomic and histological
differences compared to TAV CoA patients of the same age. The
changes included increased elastin content and altered expression
of genes involved in elastin formation, inositol signalling and
oxidative stress. The current study demonstrates that in older
children (>one month) with CoA, there are still significant
differences between the proteomes of BAV and TAV patients, but
different proteins and pathways are involved, suggesting that the
pathology associated with CoA not only develops with age, but also
develops differently in BAV patients compared to TAV patients. Each
group of patients contains one patient older than one year, whilst
the others are aged between 1 month and one year. Interestingly,
the protein expression profile of the two oldest patients are not
the most extreme in either group (see Fig. 1C); this may suggest that the
alterations in protein expression are similar in children between
one month and ten years of age.
The presence of BAV affects the
expression of proteins involved in inflammatory pathways
The most significantly enriched proteomic canonical
pathway was acute phase response signalling, which was predicted to
be activated in BAV patients. This pathway was also significantly
enriched in the phosphoproteomics data. The acute phase response is
a systemic response triggered by major local inflammation and
cytokine release. Several other inflammatory canonical pathways
were also significantly enriched. Within the proteomic data,
production of NO, ROS and the pro-inflammatory cytokine IL-12 by
macrophages were enriched, as was EIF2 signalling [a pathway
involved in the regulation of pro-inflammatory cytokine expression
(14)]. In the phosphoproteome
analysis, the third most significant canonical pathway was
activation of Th17 cells, a subset of pro-inflammatory T helper
cells (15).
These changes in canonical pathways related to
inflammation are driven by significant upregulation of the
abundance of several proteins and phosphoproteins involved in
inflammation in BAV patients compared to TAV patients.
α-1-antitrypsin (AAT), α-1-antichymotrypsin (ACT) and α-1-acid
glycoprotein 2 (AGP) were all overexpressed in BAV patients
compared to TAV patients. These proteins are all known to regulate
macrophage function (16-18)
and are all positive acute phase proteins (19,20)
(proteins whose expression increases during an inflammatory
episode), as is hemopexin (21),
another protein overexpressed in BAV patients. AAT and ACT both
belong to the serpin family of protease inhibitors and provide
negative feedback of the acute phase response via their inhibition
of inflammatory cells including neutrophils and mast cells
(22,23). Another member of the serpin family
with marked anti-inflammatory properties is anti-thrombin III
(24), and this was also
overexpressed in BAV patients. BAV patients also had overexpression
of phosphorylated α-2 Heremans-Schmid glycoprotein [AHSG; a
negative acute phase protein (25,26)]
and phosphorylated HSP90AB1 [a member of the heat shock protein 90
family, which have been demonstrated to increase secretion of
pro-inflammatory cytokines (27)].
Several studies have previously suggested that
differences in basal inflammation may exist between BAV and TAV
patients. Local inflammation is known to occur around the abnormal
BAV valve (28), however whether
the presence of BAV has an effect on aortic or systemic
inflammation is less clear. The combination of CoA and BAV has been
demonstrated to cause changes in aortic blood flow haemodynamics
including altered indices of shear stress (9), factors which are, in turn, known to
affect vascular inflammatory pathways (29). However, studies comparing
inflammatory markers such as MMPs, myeloperoxidase or measures of
macrophage infiltration in BAV vs. TAV patients have mixed
conclusions: Some studies found an increase in inflammatory markers
associated with BAV whilst others found no difference or reduced
inflammation (30-34).
The trauma of cardiac surgery is known to activate
the complement system even when no cardiopulmonary bypass is
involved (35). Although the full
immunological response takes hours to days to materialise, markers
of complement activation and some markers of inflammation start
increasing measurably during surgery itself (35), and therefore some inflammatory
proteomic changes are likely to already be occurring at the
timepoint our samples were taken. It is possible that the
differences between BAV and TAV patients in the current study
indicate that BAV patients have a greater inflammatory response to
surgery than TAV patients. Possibly changes in both basal
immunological status and in the response to surgery exist. One
recent paper found that BAV patients have decreased T and B
lymphocyte levels and the authors suggested that BAV patients have
‘an old immune system’ which is ‘more easily vulnerable to internal
and external stressors’ (31).
The presence of BAV affects the
expression of proteins involved in oxidative stress and
apoptosis
The antioxidant enzyme cytosolic superoxide
dismutase (SOD1) was significantly overexpressed in BAV CoA
patients compared to CoA patients with a normal aortic valve, and
superoxide radical degradation and superoxide dismutase activity
were highlighted in canonical pathway analysis and gene ontology
analysis respectively (Fig. 2 and
Table III). These findings
suggest there may be differences in the levels of oxidative stress
between BAV and TAV CoA patients. Previous studies have
demonstrated that adult BAV patients have increased aortic
oxidative stress compared to similar patients with a
morphologically normal aortic valve (36). This may be the result of altered
mechanical stretch in the aortic walls of BAV patients as vascular
mechanical stretch is a known stimulant of superoxide production
(37). A previous study in adults
found increased SOD1 expression in the aorta of some subsets of BAV
patients (38); to the best of our
knowledge the data in the current study is the first to suggest
that an association of increased SOD1 expression with BAV also
exists in paediatric CoA patients. However it is interesting that
in our neonatal data set (11)
there is no significant difference in SOD1 expression, suggesting
the change in expression may only occur in older children.
Previous studies in neonates with BAV associated
with CoA and adults with BAV have demonstrated decreased expression
of the extracellular form of superoxide dismutase (SOD3) associated
with BAV (11,38,39),
however there was no significant change in SOD3 expression in the
current data set. One of only two proteins to be significantly
downregulated in BAV patients was however carbonic anhydrase III,
which is known to have an antioxidant role under conditions of
oxidative stress (40). Clearly
much more work is required to understand the changes in oxidative
stress and antioxidant defence mechanisms which occur at different
ages in BAV patients with and without CoA.
Two proteins involved in apoptotic pathways, AKT3
and cytochrome c, were also significantly overexpressed in BAV CoA
patients compared to TAV CoA patients. This resulted in the
enrichment of a number of canonical pathways (Myc mediated
apoptosis signalling, ceramide signalling and lymphotoxin β
receptor signalling) which are involved in apoptosis (41-43).
Several previous studies have reported an increase in vascular
smooth muscle cell (VSMC) apoptosis in BAV patients, and have
linked this to the increased incidence of in aortic dilation and
aortic aneurysms in BAV patients (44-47).
The degree to which this is a genetic effect or secondary to
changes in blood flow haemodynamics, or a combination of both
factors, is controversial (44,48-50).
Interestingly, there were also alterations in the
ratio of haemoglobin chains. Various haemoglobin subunits are
expressed in a number of non-erythroid cells, including lungs,
neurons, endometrium and blood vessel walls (51,52).
In the current study there was overexpression of haemoglobins beta
and delta in the aorta of BAV CoA patients compared to TAV CoA
patients. The role of the beta and delta subunits in the aortic
wall is unclear however it has been suggested that haemoglobin α
acts from within blood vessel walls to help regulate nitric oxide
release and vascular tone (51). It
is possible therefore that the observed changes relate to the
altered blood flow haemodynamics in BAV patients, however much more
work is needed to understand this.
Paediatric CoA patients with BAV have a greater
long-term risk of cardiovascular complications than similar
patients with TAV (6-8).
This study demonstrates significant differences between the
proteome of the coarcted aortic tissue of young CoA patients with
and without BAV, indicating that inflammation, oxidative stress and
apoptotic pathways could contribute to future complications
associated with BAV patients. The structural changes observed in
our previous study in neonates are no longer apparent. In the
long-term, improved understanding of the molecular differences
between these patients coupled with an understanding of how the
changes evolve with age should help to optimise treatment
strategies.
Limitations of the study include the lack of tissue
from a healthy control group; this would help to elucidate how the
proteomic differences observed in BAV patients in this study are
affected by the presence of the CoA. However, such tissues would be
very difficult to obtain. Using tissues from only diseased
individuals may also increase the likelihood that pathways are
involved which deviate from the canonical paradigms, and these may
not have been fully uncovered by our analysis. Slight regional
differences in the exact location of the tissue collection may also
have introduced some variation into our analyses. Additionally,
validation of the changes in protein expression using western
blotting would be informative, however very small sample sizes made
this difficult.
Acknowledgements
Not applicable.
Funding
The present study was funded by a grant from the
British Heart Foundation (grant no. CH/1/32804.)
Data availability
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Author contributions
Conceptualization: MC, MTG, MSS; investigation and
analysis: KLS, AJB, SA-G, DI, KJH, MCW, MTG, MGB; sample
collection: MC, RM, S-LK, SS, writing-original draft preparation:
MSS, KLS; writing-review and editing: KLS, MGB, MSS, MC; funding
acquisition: MTG, MC, MSS. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
The study was conducted in accordance with the
declaration of Helsinki, and the protocol was approved by the North
Somerset and South Bristol Research Ethics Committee (Health
Research Authority, Whitefriars, Level 3 Block B, Lewins Mead,
Bristol, BS1 2NT, REC 07/H0106/172). Full informed consent was
obtained from parents prior to admission for operation.
Patients consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Teo LL, Cannell T, Babu-Narayan SV, Hughes
M and Mohiaddin RH: Prevalence of associated cardiovascular
abnormalities in 500 patients with aortic coarctation referred for
cardiovascular magnetic resonance imaging to a tertiary center.
Pediatr Cardiol. 32:1120–1127. 2011.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Sinning C, Zengin E, Kozlik-Feldmann R,
Blankenberg S, Rickers C, von Kodolitsch Y and Girdauskas E:
Bicuspid aortic valve and aortic coarctation in congenital heart
disease-important aspects for treatment with focus on aortic
vasculopathy. Cardiovasc Diagn Ther. 8:780–788. 2018.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Preventza O, Livesay JJ, Cooley DA,
Krajcer Z, Cheong BY and Coselli JS: Coarctation-associated
aneurysms: A localized disease or diffuse aortopathy. Ann Thorac
Surg. 95:1961–1967. 2013.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Webb G: Treatment of coarctation and late
complications in the adult. Semin Thorac Cardiovasc Surg.
17:139–142. 2005.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Yin Z, Yang JR, Wei YS, Liang BL, Wei YB,
Zhou KQ, Wang Z, Yan B and Gao YL: Ischemia-reperfusion injury in
an aortic dissection patient. Am J Emerg Med. 33:987.e5–e6.
2015.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Bambul Heck P, Pabst von Ohain J,
Kaemmerer H, Ewert P and Hager A: Survival and cardiovascular
events after coarctation-repair in long-term follow-up (COAFU):
Predictive value of clinical variables. Int J Cardiol. 228:347–351.
2017.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Jashari H, Lannering K, Ibrahimi P, Djekic
D, Mellander M, Rydberg A and Henein MY: Persistent reduced
myocardial deformation in neonates after CoA repair. Int J Cardiol.
221:886–891. 2016.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Oliver JM, Gallego P, Gonzalez A, Aroca A,
Bret M and Mesa JM: Risk factors for aortic complications in adults
with coarctation of the aorta. J Am Coll Cardiol. 44:1641–1647.
2004.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Keshavarz-Motamed Z, Garcia J and Kadem L:
Fluid dynamics of coarctation of the aorta and effect of bicuspid
aortic valve. PLoS One. 8(e72394)2013.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Pedersen TA: Late morbidity after repair
of aortic coarctation. Dan Med J. 59(B4436)2012.PubMed/NCBI
|
|
11
|
Skeffington KL, Bond AR, Abdul-Ghani S,
Iacobazzi D, Kang SL, Heesom KJ, Wilson MC, Ghorbel M, Stoica S and
Martin R: Bicuspid aortic valve alters aortic protein expression
profile in neonatal coarctation patients. J Clin Med 8:
5172019.
|
|
12
|
Valikangas T, Suomi T and Elo LL: A
comprehensive evaluation of popular proteomics software workflows
for label-free proteome quantification and imputation. Brief
Bioinform. 19:1344–1355. 2018.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Bond AR, Iacobazzi D, Abdul-Ghani S,
Ghorbel MT, Heesom KJ, George SJ, Caputo M, Suleiman MS and Tulloh
RM: The cardiac proteome in patients with congenital ventricular
septal defect: A comparative study between right atria and right
ventricles. J Proteomics. 191:107–113. 2019.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Shrestha N, Bahnan W, Wiley DJ, Barber G,
Fields KA and Schesser K: Eukaryotic initiation factor 2 (eIF2)
signaling regulates proinflammatory cytokine expression and
bacterial invasion. J Biol Chem. 287:28738–28744. 2012.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Sandquist I and Kolls J: Update on
regulation and effector functions of Th17 cells. F1000Res.
7(205)2018.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Bories PN, Guenounou M, Feger J, Kodari E,
Agneray J and Durand G: Human alpha 1-acid glycoprotein-exposed
macrophages release interleukin 1 inhibitory activity. Biochem
Biophys Res Commun. 147:710–715. 1987.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Krotova K, Marek GW, Wang RL, Aslanidi G,
Hoffman BE, Khodayari N, Rouhani FN and Brantly ML: Alpha-1
antitrypsin-deficient macrophages have increased
matriptase-mediated proteolytic activity. Am J Respir Cell Mol
Biol. 57:238–247. 2017.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Skeel A and Leonard EJ: alpha
1-Antichymotrypsin is the human plasma inhibitor of macrophage
ectoenzymes that cleave pro-macrophage stimulating protein. J Biol
Chem. 276:21932–21937. 2001.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Fournier T, Medjoubi NN and Porquet D:
Alpha-1-acid glycoprotein. Biochim Biophys Acta. 1482:157–171.
2000.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Jain S, Gautam V and Naseem S: Acute-phase
proteins: As diagnostic tool. J Pharm Bioallied Sci. 3:118–127.
2011.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Rolla S, Ingoglia G, Bardina V, Silengo L,
Altruda F, Novelli F and Tolosano E: Acute-phase protein hemopexin
is a negative regulator of Th17 response and experimental
autoimmune encephalomyelitis development. J Immunol. 191:5451–5459.
2013.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Janciauskiene S, Wrenger S, Immenschuh S,
Olejnicka B, Greulich T, Welte T and Chorostowska-Wynimko J: The
multifaceted effects of Alpha1-antitrypsin on neutrophil functions.
Front Pharmacol. 9(341)2018.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Kalsheker NA: Alpha 1-antichymotrypsin.
Int J Biochem Cell Biol. 28:961–964. 1996.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Levy JH, Sniecinski RM, Welsby IJ and Levi
M: Antithrombin: Anti-inflammatory properties and clinical
applications. Thromb Haemost. 115:712–728. 2016.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Haglund AC, Ek B and Ek P: Phosphorylation
of human plasma alpha2-Heremans-Schmid glycoprotein (human fetuin)
in vivo. Biochem J. 357:437–445. 2001.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Lebreton JP, Joisel F, Raoult JP, Lannuzel
B, Rogez JP and Humbert G: Serum concentration of human alpha 2 HS
glycoprotein during the inflammatory process: Evidence that alpha 2
HS glycoprotein is a negative acute-phase reactant. J Clin Invest.
64:1118–1129. 1979.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Tukaj S, Zillikens D and Kasperkiewicz M:
Inhibitory effects of heat shock protein 90 blockade on
proinflammatory human Th1 and Th17 cell subpopulations. J Inflamm
(Lond). 11(10)2014.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Mathieu P, Bosse Y, Huggins GS, Della
Corte A, Pibarot P, Michelena HI, Limongelli G, Boulanger MC,
Evangelista A, Bédard E, et al: The pathology and pathobiology of
bicuspid aortic valve: State of the art and novel research
perspectives. J Pathol Clin Res. 1:195–206. 2015.PubMed/NCBI View
Article : Google Scholar
|
|
29
|
Helderman F, Segers D, de Crom R, Hierck
BP, Poelmann RE, Evans PC and Krams R: Effect of shear stress on
vascular inflammation and plaque development. Curr Opin Lipidol.
18:527–533. 2007.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Ali OA, Chapman M, Nguyen TH, Chirkov YY,
Heresztyn T, Mundisugih J and Horowitz JD: Interactions between
inflammatory activation and endothelial dysfunction selectively
modulate valve disease progression in patients with bicuspid aortic
valve. Heart. 100:800–805. 2014.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Balistreri CR, Buffa S, Allegra A, Pisano
C, Ruvolo G, Colonna-Romano G, Lio D, Mazzesi G, Schiavon S, Greco
E, et al: A typical immune T/B subset profile characterizes
bicuspid aortic valve: In an old status? Oxid Med Cell Longev.
2018(5879281)2018.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Balistreri CR, Pisano C, Candore G, Maresi
E, Codispoti M and Ruvolo G: Focus on the unique mechanisms
involved in thoracic aortic aneurysm formation in bicuspid aortic
valve versus tricuspid aortic valve patients: Clinical implications
of a pilot study. Eur J Cardiothorac Surg. 43:e180–e186.
2013.PubMed/NCBI View Article : Google Scholar
|
|
33
|
LeMaire SA, Wang X, Wilks JA, Carter SA,
Wen S, Won T, Leonardelli D, Anand G, Conklin LD, Wang XL, et al:
Matrix metalloproteinases in ascending aortic aneurysms: bicuspid
versus trileaflet aortic valves. J Surg Res. 123:40–48.
2005.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Tzemos N, Lyseggen E, Silversides C,
Jamorski M, Tong JH, Harvey P, Floras J and Siu S: Endothelial
function, carotid-femoral stiffness, and plasma matrix
metalloproteinase-2 in men with bicuspid aortic valve and dilated
aorta. J Am Coll Cardiol. 55:660–668. 2010.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Gu YJ, Mariani MA, Boonstra PW, Grandjean
JG and van Oeveren W: Complement activation in coronary artery
bypass grafting patients without cardiopulmonary bypass: The role
of tissue injury by surgical incision. Chest. 116:892–898.
1999.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Billaud M, Phillippi JA, Kotlarczyk MP,
Hill JC, Ellis BW, St Croix CM, Cantu-Medéllin N, Kelley EE and
Gleason TG: Elevated oxidative stress in the aortic media of
patients with bicuspid aortic valve. J Thorac Cardiovasc Surg.
154:1756–1762. 2017.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Birukov KG: Cyclic stretch, reactive
oxygen species, and vascular remodeling. Antioxid Redox Signal.
11:1651–1667. 2009.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Phillippi JA, Hill JC, Billaud M, Green
BR, Kotlarczyk MP and Gleason TG: Bicuspid aortic valve morphotype
correlates with regional antioxidant gene expression profiles in
the proximal ascending aorta. Ann Thorac Surg. 104:79–87.
2017.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Arcucci A, Ruocco MR, Albano F, Granato G,
Romano V, Corso G, Bancone C, De Vendittis E, Della Corte A and
Montagnani S: Analysis of extracellular superoxide dismutase and
Akt in ascending aortic aneurysm with tricuspid or bicuspid aortic
valve. Eur J Histochem. 58(2383)2014.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Di Fiore A, Monti DM, Scaloni A, De Simone
G and Monti SM: Protective role of carbonic anhydrases iii and vii
in cellular defense mechanisms upon redox unbalance. Oxid Med Cell
Longev. 2018(2018306)2018.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Browning JL, Miatkowski K, Sizing I,
Griffiths D, Zafari M, Benjamin CD, Meier W and Mackay F: Signaling
through the lymphotoxin beta receptor induces the death of some
adenocarcinoma tumor lines. J Exp Med. 183:867–878. 1996.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Haimovitz-Friedman A, Kolesnick RN and
Fuks Z: Ceramide signaling in apoptosis. Br Med Bull. 53:539–553.
1997.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Hoffman B and Liebermann DA: Apoptotic
signaling by c-MYC. Oncogene. 27:6462–6472. 2008.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Bonderman D, Gharehbaghi-Schnell E,
Wollenek G, Maurer G, Baumgartner H and Lang IM: Mechanisms
underlying aortic dilatation in congenital aortic valve
malformation. Circulation. 99:2138–2143. 1999.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Cecconi M, Nistri S, Quarti A, Manfrin M,
Colonna PL, Molini E and Perna GP: Aortic dilatation in patients
with bicuspid aortic valve. J Cardiovasc Med (Hagerstown). 7:11–20.
2006.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Nataatmadja M, West M, West J, Summers K,
Walker P, Nagata M and Watanabe T: Abnormal extracellular matrix
protein transport associated with increased apoptosis of vascular
smooth muscle cells in marfan syndrome and bicuspid aortic valve
thoracic aortic aneurysm. Circulation. 108 (Suppl 1):II329–II334.
2003.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Schmid FX, Bielenberg K, Schneider A,
Haussler A, Keyser A and Birnbaum D: Ascending aortic aneurysm
associated with bicuspid and tricuspid aortic valve: Involvement
and clinical relevance of smooth muscle cell apoptosis and
expression of cell death-initiating proteins. Eur J Cardiothorac
Surg. 23:537–543. 2003.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Hahn RT, Roman MJ, Mogtader AH and
Devereux RB: Association of aortic dilation with regurgitant,
stenotic and functionally normal bicuspid aortic valves. J Am Coll
Cardiol. 19:283–288. 1992.PubMed/NCBI View Article : Google Scholar
|
|
49
|
Harrison OJ, Visan AC, Moorjani N, Modi A,
Salhiyyah K, Torrens C, Ohri S and Cagampang FR: Defective NOTCH
signaling drives increased vascular smooth muscle cell apoptosis
and contractile differentiation in bicuspid aortic valve
aortopathy: A review of the evidence and future directions. Trends
Cardiovasc Med. 29:61–68. 2019.PubMed/NCBI View Article : Google Scholar
|
|
50
|
Niwa K, Perloff JK, Bhuta SM, Laks H,
Drinkwater DC, Child JS and Miner PD: Structural abnormalities of
great arterial walls in congenital heart disease: Light and
electron microscopic analyses. Circulation. 103:393–400.
2001.PubMed/NCBI View Article : Google Scholar
|
|
51
|
Butcher JT, Johnson T, Beers J, Columbus L
and Isakson BE: Hemoglobin alpha in the blood vessel wall. Free
Radic Biol Med. 73:136–142. 2014.PubMed/NCBI View Article : Google Scholar
|
|
52
|
Saha D, Patgaonkar M, Shroff A, Ayyar K,
Bashir T and Reddy KV: Hemoglobin expression in nonerythroid cells:
Novel or ubiquitous? Int J Inflam. 2014(803237)2014.PubMed/NCBI View Article : Google Scholar
|