Introduction
Lipopolysaccharide (LPS) is a component of
Gram-negative bacterial cell walls, which releases a variety of
inflammatory factors that lead to hepatic necrosis (1). Antioxidant enzyme levels and free
radical scavenging have been reported to decrease following LPS
exposure (2,3). Drug-induced liver injury (DILI) is a
leading limitation of therapeutic drug use and often leads to
post-marketing drug withdrawal and attrition due to toxicity during
drug development (4,5). Between 2004 and 2007 antibacterial
agents accounted for 45.5% of DILI in the USA, including
amoxicillin/clavulanate multiples, third generation cephalosporins,
fluoroquinolones and amidealcohols (6-8).
Enrofloxacin (ENR) is a fluoroquinolone, which is widely used for
the prevention and treatment of poultry-associated bacterial
infections, however the long-term use of ENR may induce liver
injury (9). Previous reports
(10,11) have confirmed that ENR induces
endotoxin release by disrupting the outer membrane of organisms
(12).
Composite ammonium glycyrrhizin (CAG) is composed of
ammonium glycyrrhizin, glycine and methionine (13). The primary bioactive components of
licorice root are glycyrrhizin and glycyrrhizic acid (GA), which
are commonly used in Asia to treat patients with chronic hepatitis
(14,15). As an ammonium salt of glycyrrhizic
acid, ammonium glycyrrhizin also has protective functions. GA may
promote liver cell proliferation, thereby promoting liver
regeneration (16). GA also
exhibits antiviral, anti-tumor and immunomodulatory activity
(17-19).
In previous studies, it has been demonstrated that GA may prevent
liver injury by suppressing lipid peroxidation reactions, which
enhances the ability of the liver to scavenge free radicals and
resist free radical damage in the liver (20). CAG has been reported to be effective
as an anti-inflammatory, anticancer, antihepatotoxic and
antioxidant agent (13,16). In addition, its antiradical activity
has been reported to be responsible for its anti-inflammatory
actions (13,21,22).
Although the beneficial effects of CAG have been
extensively studied, it remains unknown as to whether it protects
the liver from a combination of LPS- and ENR-induced injuries.
Therefore, the aim of the present study was to establish an in
vitro LPS/ENR-induced liver model and utilize this model to
evaluate the protective effects of CAG, whilst also examining the
molecular mechanisms underlying the protective effects.
Materials and methods
Reagents and materials
CAG (2 mg/ml) was produced by the authors in our
laboratory [ammonium glycyrrhizin (2.8 g; Shaanxi FUJIE
Pharmaceutical Co., Ltd., Xianyang, China), glycine (2 g) and
methionine (2 g, Tianjin Tianyao Pharmaceuticals Co., Ltd.,
Tianjin, China) filled with water to 1,000 g)]. ENR was purchased
from the National Institute for the Control of Pharmaceutical and
Biological Products (Beijing, China). LPS (E. coli L-2880;
Sigma-Aldrich; Merck KGaA, Darmstadt, Germany). Dulbecco's modified
Eagle's medium (DMEM) was purchased from Hyclone; GE Healthcare
Life Sciences (Logan, UT, USA). Primary antibodies against
apoptosis regulator Bcl-2 (Bcl-2; cat. no. 610538; 1:1,000; BD
Biosciences, Franklin Lakes, NJ, USA), Bcl-2 associated X-protein
(Bax; cat. no. orb334986; 1:1,000; Biorbyt Ltd., Cambridge, UK),
caspase-3 (cat. no. ab115183; 1:1,000, Abcam, Cambridge, USA),
β-actin (1:5,000; Sigma-Aldrich; Merck KGaA, A5441) and secondary
horseradish peroxidase-labeled goat anti-rabbit IgG (cat. no.
4201-100; 1:1,000; Shanghai Pufei Biotechnology Co., Ltd.,
Shanghai, China) were obtained.
Cell isolation and culture
A total of 12 30 day old male Hailan chickens (mean
weight, 1.5±0.2 kg) were purchased from Qinglong Mountain (Nanjing,
China) and housed at 22˚C with ventilation using 12-h light/dark
cycles. Animals had free access to food and water and were fasting
12 h prior to experiments. Hepatocytes (23,24)
were isolated from the chickens (using the two-stage, collagenase
type IV perfusion method via the portal vein. Briefly, the livers
were perfused with solution A [33 mM 4-hydroxyethylpiperazine
ethanesulfonic acid (HEPES), 127.8 mM NaCl, 3.15 mM KCl, 0.7 mM
Na2HPO4 x 12 H2O and 0.6 mM EGTA;
pH 7.6] and solution B (33 mM HEPES, 127.8 mM NaCl, 3.15 mM KCl,
0.7 mM Na2HPO4 x 12 H2O and 3 mM
CaCl2; pH 7.6) at room temperature at a flow rate of
10-20 ml/min for 15 min. A total of 100 ml 0.5% collagenase type IV
buffer (C0016; NanJing HoldBio E-Commerce Co., Ltd., Nanjing,
China) was perfused for 20-25 min at 37˚C at a flow rate of 20
ml/min. The hepatocytes were separated from the other cellular
components by centrifugation at 167.7 x g for 3 min at 37˚C.
Isolated cells were washed twice in solution B supplemented with
0.2% bovine serum albumin (BSA; Sigma Aldrich; Merck KGaA). MTT
stock solution (5 µl; 5 mg/ml) was added in each well and cells
were incubated in a humidified incubator with an atmosphere of 5%
CO2 for 4 h at 37˚C. Absorbance at 570 nm was measured
using a microplate reader. Cell yield varied between 4 and
5x108 cells/liver and the cell viability varied between
90 and 95% as determined by trypan blue exclusion (0.04%; Beijing
Solarbio Science & Technology Co., Ltd., Beijing, China).
Adherent cells were digested using trypsin to prepare single cell
suspension. Cell suspension was mixed with 0.4% trypan blue
solution (9:1). Following 3 min at room temperature, living and
dead cells were counted. Dead cells appeared blue and living cells
were colorless/transparent. The viable cell rate was defined as:
Viable cell rate (%)=total viable cells/total cells.
The present study was approved by the Animal Ethics
Committee of Nanjing Agricultural University (Nanjing, China).
In-vitro LPS/ENR-induced hepatocyte
injury model
Chicken hepatocytes were plated at a density of
5x105 cells in 96-well cell plates and cultured for 48 h
at 37˚C in DMEM. LPS (30 µg/ml) and different concentrations of ENR
(40-120 µg/ml) were added to determine the optimal concentration.
Cytotoxicity was measured using an MTT assay. MTT (0.1%) was added
to the cell plates and incubated at 37˚C for 4 h. The medium was
added to 150 µl dimethyl sulfoxide to dissolve the purple formazan
and the absorbance was measured at a wavelength of 570 nm using a
microplate reader.
Treatment groups
The cells were divided into four groups: i) Control
group, not administered with CAG or exposed to LPS/ENR; ii) model
group, administered LPS/ENR (30/80 µg/ml) for 24 h; iii) CAG group,
administered CAG 400 µg/ml; and iv) combined group, pretreated with
CAG (25, 50, 100, 200 and 400 µg/ml) for 24 h prior to LPS/ENR
(30/80 µg/ml).
Analysis of alanine aminotransferase
(ALT) and aspartate aminotransferase (AST) enzyme levels in the
cell culture supernatant
ALT and AST are liver enzyme markers (1). These two enzymes were measured by
spectrophotometric analysis. Cell supernatants of the different
groups were obtained by centrifugation (1,006 x g; 10 min; 4˚C) and
the activity of ALT (C009-1; Jiancheng Bioengineering Institute
Co., Ltd., Nanjing, China) and AST (C010-1; Jiancheng
Bioengineering Institute Co., Ltd.) were determined according to
the manufacturer's protocol.
Enzyme activity in hepatocytes
The hepatocytes were lysed in a
radioimmunoprecipitation assay buffer (RIPA; Jiancheng
Bioengineering Institute Co., Ltd.) on ice and centrifuged at 7,280
x g for 15 min at 4˚C. The cell lysate supernatant was aspirated
and stored at -20˚C prior to glutathione (GSH) assays (reduced
glutathione assay kit; A006-1; Jiancheng Bioengineering Institute
Co., Ltd.), superoxide dismutatse (SOD) assays (SOD assay kit;
A001-3; Jiancheng Bioengineering Institute Co., Ltd.), catalase
(CAT) assays (CAT assay kit; A007-1-1; Jiancheng Bioengineering
Institute Co., Ltd.), glutathione peroxidase (GPx) assays (GPx
assay kit; A005; Jiancheng Bioengineering Institute Co., Ltd.) and
malondialdehyde (MDA) assays (MDA assay kit; A003-1; Jiancheng
Bioengineering Institute Co., Ltd.) according to the manufacturer's
protocols. The protein concentration of each sample was determined
using the bicinchoninic acid protein quantitation cassette.
Flow cytometry analysis
In normal viable cells propidium iodide (PI) and
Annexin V are negative, while in early apoptotic cells Annexin V is
positive and PI is negative. In late apoptotic cells Annexin V and
PI are positive, while in dead cells Annexin V is negative and PI
is positive. Cells following 24 h LPS/ENR treatment were collected
(560 x g; 5 min; 4˚C) and stained using an Annexin V-fluorescein
(FITC)/PI Apoptosis Detection kit (BD Biosciences) according to
manufacturer's protocol. The cells collected were then analyzed by
flow cytometry using a BD FACS Calibur Cell Sorting system (BD
Biosciences, Franklin Lakes, NJ, USA) and FlowJo v10 (FlowJo LLC,
Ashland, OR, USA).
Extraction of total RNA and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
Chicken hepatocytes following 24 h treatment with
LPS/ENR were collected (7,280 x g; 15 min, 4˚C) and total RNA was
extracted using an RNAiso Plus kit (Takara Biotechnology Co., Ltd.,
Dalian, China). RT was performed using the PrimeScript™
RT reagent Kit with gDNA Eraser (RR037A; Takara Biotechnology Co.,
Ltd.) at 37˚C for 15 min, followed by 85˚C for 5 sec. RNA
concentrations were detected using a Qubit RNA Assay kit and a
Qubit 2.0 fluorometer (Invitrogen; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA). The primer sequences of the genes used for PCR
are listed in Table I. SYBR green
premix (RR820A; Takara Biotechnology Co., Ltd.) was used for qPCR
analysis. The PCR thermocycling conditions were as follows: 95˚C
for 30 sec, followed by 40 cycles of 95˚C for 5 sec and 60˚C for 30
sec. Quantification was based on the
2-ΔΔCq method (25) and β-actin was used as the
housekeeping gene.
 | Table IPrimer sequences of the target genes
used for polymerase chain reaction. |
Table I
Primer sequences of the target genes
used for polymerase chain reaction.
| | Primer sequence
(5'→3') |
|---|
| Gene | Forward | Reverse |
|---|
| β-actin |
ATGTGGATCAGCAAGCAGGAGTA |
TTTATGCGCATTTATGGGTTTTGT |
| Bax |
GTGATGGCATGGGACATAGCTC |
TGGCGTAGACCTTGCGGATAA |
| Bcl-2 |
ATCGTCGCCTTCTTCGAGTT |
ATCCCATCCTCCGTTGTCCT |
| Caspase-3 |
AAGGCTCCTGGTTTATTC |
CTGCCACTCTGCGATTTA |
Western blotting analysis
Total protein was extracted from chicken hepatocytes
using a RIPA buffer. Protein concentrations were determined using
bicinchoninic acid assays. Equal amounts of protein (100 µg) were
separated on 10% SDS-PAGE and then transferred to polyvinylidene
difluoride membranes. Membranes were blocked with 5% BSA at room
temperature for 2 h followed by incubation with primary antibodies
overnight at 4˚C. Membranes were incubated with secondary
antibodies for 1 h at room temperature. Following, membranes were
washed with TBST and immunoreactive bands were detected by using
enhanced chemiluminescence (Vazyme, Piscataway, NJ, USA). The
western blot bands were analyzed using ImageJ software (1.46r;
National Institutes of Health, Bethesda, MD, USA).
Measurement of Caspase-3 activity
Caspase-3 activity was measured in all groups
following 24 h of LPS/ENR treatment using a commercial Caspase-3
Colorimetric Assay kit (C1115; Beyotime Institute of Biotechnology,
Shanghai, China) according to the manufacturer's protocol.
Transmission electron microscopy
(TEM)
Cells were fixed with 2.5% glutaraldehyde in 0.1 M
sodium phosphate buffer (pH 7.2) for 3 h at 4˚C. Fixed cells were
subsequently cut into sections for TEM (10-100 nm). The sections
were stained with 2% uranyl acetate at 37˚C for 30 min and embedded
in osmium tetroxide at 37˚C for 30 min and observed under a Hitachi
TEM (Hitachi, Ltd., Tokyo, Japan).
Mitochondrial membrane potential (MMP)
assay
Hepatocyte MMP was measured using the
membrane-sensitive JC-1 dye and analyzed with a fluorescence
microscope. JC-1 staining solution (1 ml; Mitochondrial Membrane
Potential kit; Beyotime Institute of Biotechnology) was added to
each well and mixed. Cells were incubated for 20 min at 37˚C in an
incubator with the JC-1 stain. Following incubation the supernatant
was aspirated, cells were washed twice with JC-1 buffer solution at
room temperature for 20 sec, 2 ml DMEM were added and samples were
observed under a fluorescence microscope (magnification, x100).
Statistical analysis
All data are expressed as the mean ± standard
deviation, from ≥3 repeats. Data were analyzed using SPSS software
version 19.0 (IBM Corp., Armonk, NY, USA). One-way analysis of
variance with Dunnett's post hoc test was used for the comparison
of multiple groups. P<0.05 was considered to indicate a
statistically significant difference.
Results
Establishing of an LPS/ENR-induced
model of liver injury
Assessment of the variations in hepatocyte cell
viability and morphological evaluation following LPS/ENR treatment
were performed. The results demonstrated that LPS/ENR treatment
decreased the viability of cells in a dose-dependent manner
(Fig. 1A). When cells were treated
with LPS/ENR 30/80-30/120 µg/ml they demonstrated evident reduced
cell viability. When the concentration of LPS/ENR was 30/100 µg/ml,
the cell viability was 22.57±0.43%. Exposure of cells to 30/120
µg/ml LPS/ENR injury led to a notable reduction in cell number and
these cells exhibited morphological alterations, including the
disruption of cell membranes and nuclear fragmentation compared
with the control cells. At LPS/ENR 30/80 µg/ml the cell viability
was 48.27±2.50% and the cell shape was irregular with clear
disruption of the cell membrane. Therefore, LPS/ENR 30/80 µg/ml was
determined to be the most appropriate concentration and was used in
the model groups for all further experimentation.
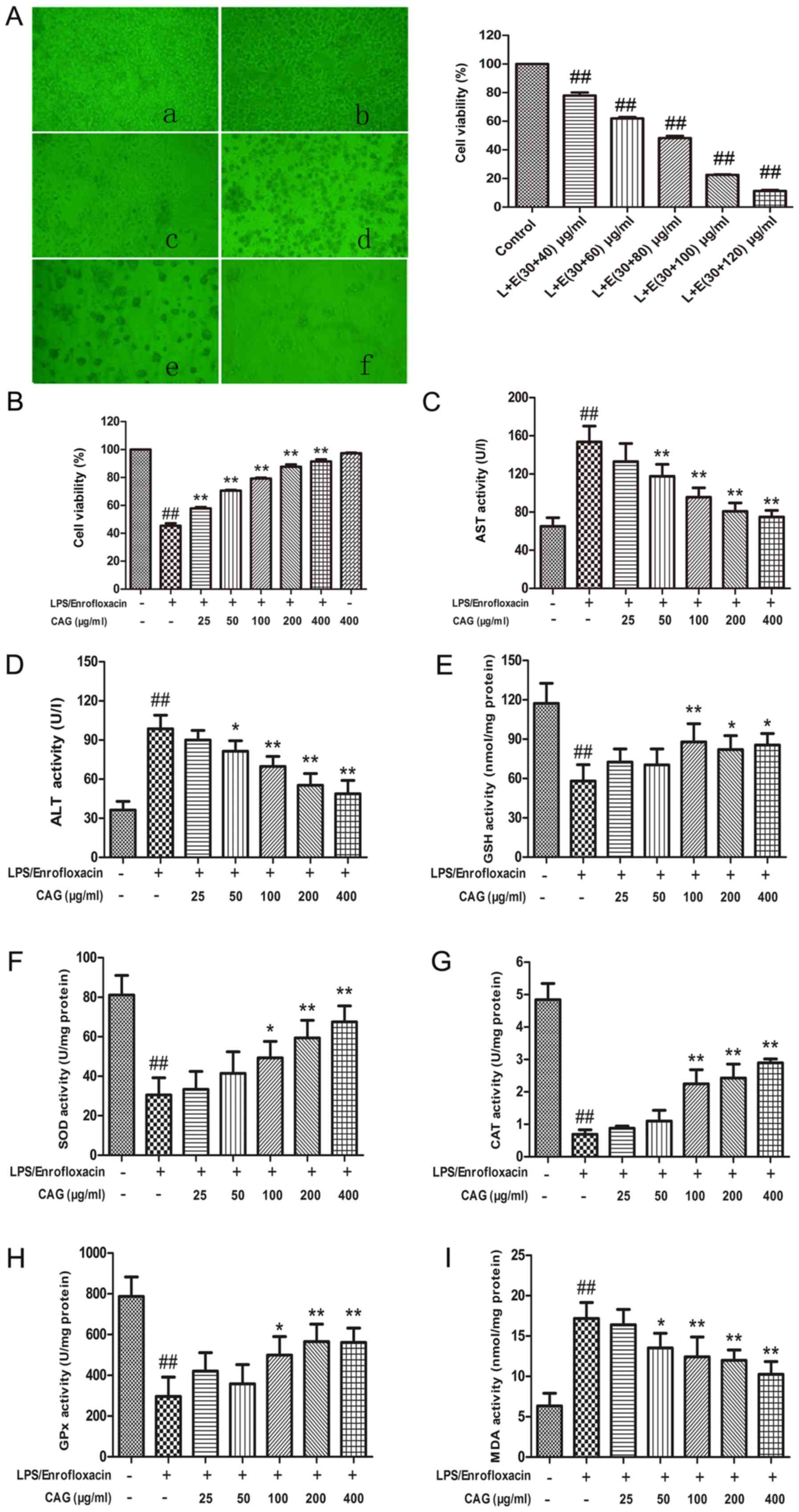 | Figure 1(A) Effect of different
concentrations of LPS/ENR on cytomorphology (magnification, x20)
and hepatocyte viability. Images of (a) normal cells; and cells
treated with (b) 30/40 µg/ml; (c) 30/60 µg/ml; (d) 30/80 µg/ml; (e)
30/100 µg/ml; and (f) 30/120 µg/ml LPS/ENR. The effect of CAG on
(B) cell viability, (C) AST, (D) ALT, (E) GSH, (F) SOD and (G) CAT
activity was The effect of CAG on (H) GPx and (I) MDA activity was
determined. LPS/enrofloxacin was given at a dose of 30/80 µg/ml.
Hepatocytes were collected following CAG treatment for 24 h. Data
are expressed as the mean ± standard deviation (n=3).
##P<0.01 vs. the control group;
*P<0.05, **P<0.01 vs. the model group
treated with 30/80 µg/ml LPS/ENR. LPS or L, lipopolysaccharide;
CAG, composite ammonium glycyrrhizin; AST, aspartate
aminotransferase; ALT, alanine aminotransferase; GSH, glutathione;
SOD, superoxide dismutase; CAT, catalase; GPx, glutathione
peroxidase; MDA, malondialdehyde; ENR or E, enrofloxacin. |
CAG attenuates LPS/ENR-induced
hepatocyte injury
ALT and AST activities were measured in the
supernatant and the cell viability was evaluated using an MTT
assay. Following treatment with CAG (400 µg/ml) the cell viability
was 97.39±0.75%, which was similar to the negative control group
and indicated the non-toxic nature of CAG (Fig. 1B). Treatment with LPS/ENR (30/80
µg/ml) induced a significant elevation in ALT (Fig. 1C) and AST (Fig. 1D) concentrations (P<0.01)
compared with the control, while treatment with CAG (50-400 µg/ml)
caused a dose-dependent decrease compared with the model group
(P<0.05 and P<0.01).
LPS/ENR (30/80 µg/ml) treatment of hepatocytes
significantly reduced the cell viability of the hepatocytes
compared with the control group (P<0.01; Fig. 1B). Hepatocytes treated with CAG
demonstrated a dose dependent improvement in cell growth and all
groups treated with CAG had significantly improved cell viability
compared with the model group (all P<0.01).
MDA and assays of the antioxidant
enzymes GSH, SOD, CAT and GPx
Treating hepatocytes with LPS/ENR (30/80 µg/ml) for
24 h caused a significant reduction in activity of GSH (P<0.01;
Fig. 1E), SOD (P<0.01; Fig. 1F), CAT (P<0.01; Fig. 1G) and GPx (P<0.01; Fig. 1H) compared with the control group.
Treatment with LPS/ENR caused a significant elevation in MDA levels
(P<0.01; Fig. 1I) compared with
the control group. CAG treatment provided a protective effect as
evidenced by the restoration of these biomarkers. The highest dose
of CAG (400 µg/ml) attenuated protein MDA levels and caused an
increase in GSH protein levels and the activities of the SOD, CAT
and GPx antioxidant enzymes (P<0.01; Fig. 1E-I).
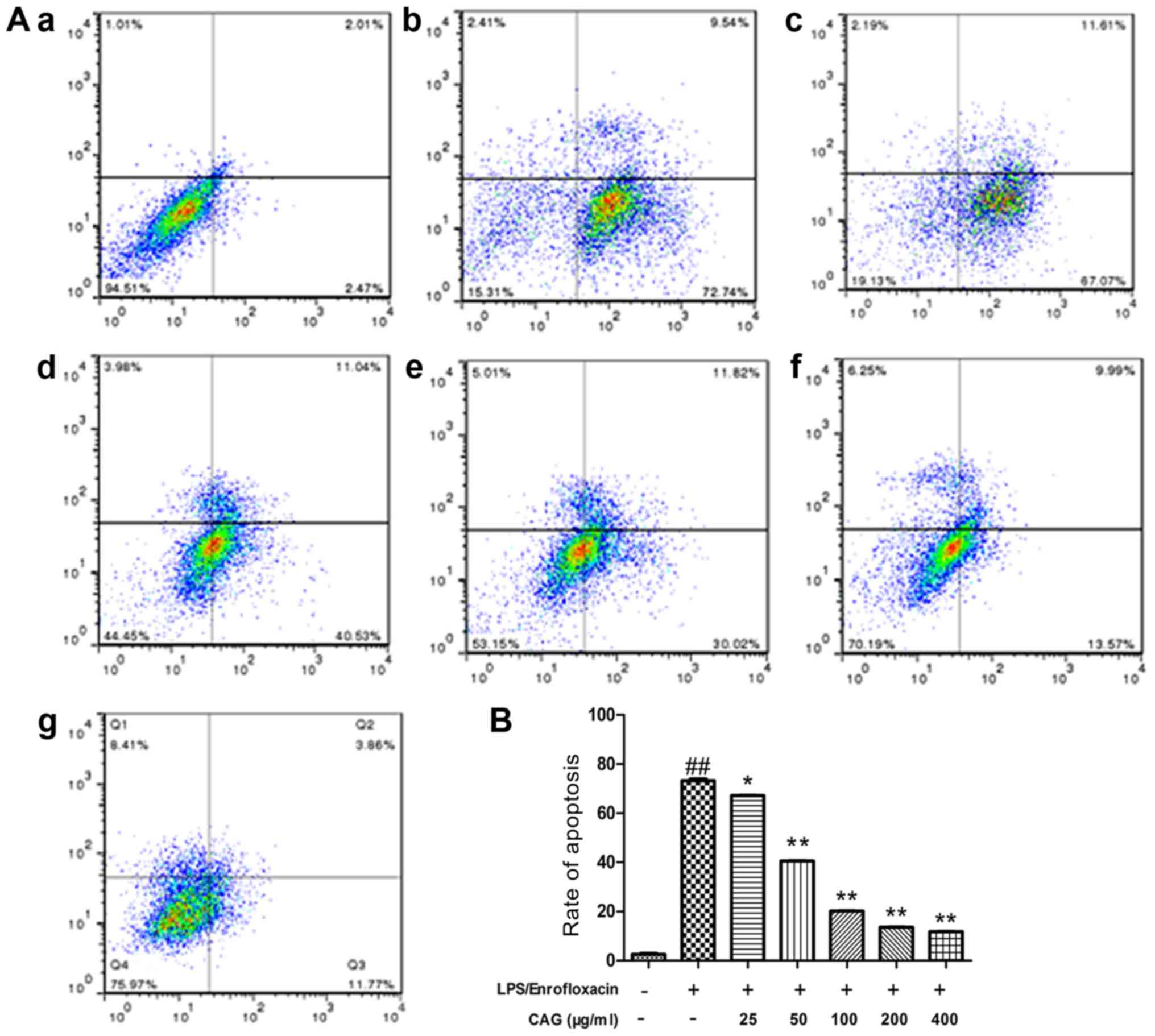
Apoptosis assay using Annexin
V-FITC/PI staining and flow cytometry
To confirm apoptotic activity, flow cytometry
analysis of hepatocytes was performed using dual stain Annexin
V-FITC/PI (Fig. 2). In the control
group there was a high percentage of surviving cells (91.40±3.74%)
and a low percentage of early apoptotic (2.69±0.79%) and late
apoptotic (3.05±1.81%) cells. In the LPS/ENR group the percentage
of surviving cells (17.26±3.98%) was significantly decreased
compared with the control group. The number of early apoptotic
(73.03±3.36%) and late apoptotic (10.42±2.15%) cells were
significantly increased following LPS/ENR (30/80 µg/ml) treatment
in comparison with the control group. The number of early apoptotic
cells (66.92±2.73, 41.73±4.07, 31.32±2.74%, 13.82±2.07 and
11.53±1.17%) notably reduced in a dose-dependent manner with
increasing concentrations of CAG (25, 50, 100, 200 and 400 µg/ml,
respectively). All groups treated with CAG had a significantly
reduced number of apoptotic cells compared with the model group
(73.03±3.36%; P<0.05).
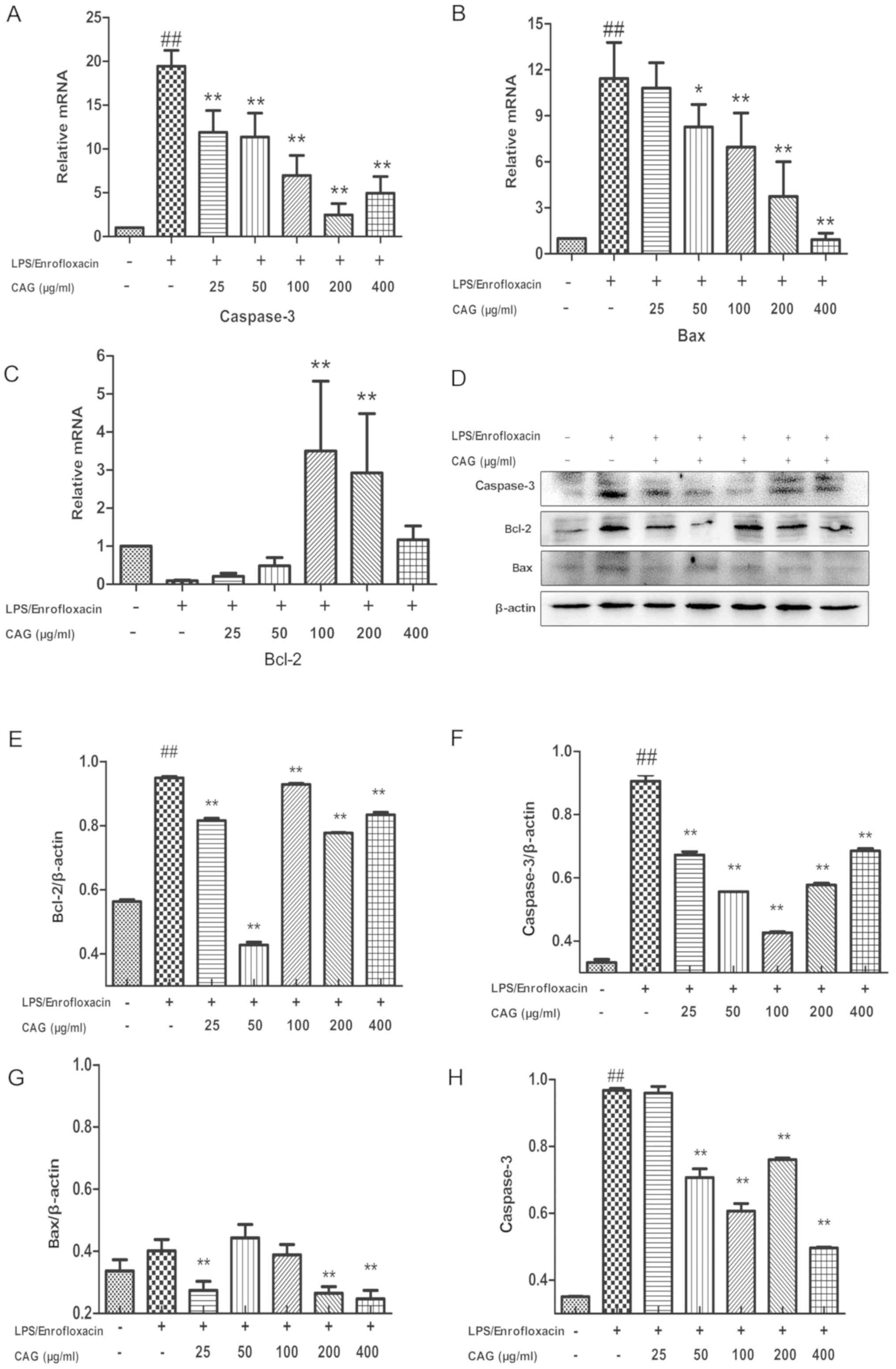
CAG modifies caspase-3, Bax and Bcl-2
mRNA expression
Treatment with LPS/ENR (30/80 µg/ml) significantly
increased hepatocyte caspase-3 (Fig.
3A) and Bax (Fig. 3B) mRNA
expression 19.45±1.87- and 11.43±2.35-fold, respectively compared
with the control group (P<0.01). Treatment with 25-400 µg/ml CAG
significantly inhibited Caspase-3 expression (P<0.01; Fig. 3A). Treatment with ≥50 µg/ml CAG
significantly inhibited Bax mRNA expression (P<0.05; Fig. 3B). LPS/ENR (30/80 µg/ml) treatment
markedly suppressed Bcl-2 expression, while CAG treatment at doses
of 100 and 200 µg/ml significantly increased Bcl-2 mRNA expression
compared with the control (P<0.01; Fig. 3C). Interestingly, at 400 µg/ml Bcl-2
mRNA was not significantly decreased (P>0.05).
CAG treatment affects the protein
expression of caspase-3, Bax and Bcl-2
Western blot analysis was performed to confirm the
apoptotic data (Fig. 3D). Bcl-2 and
caspase-3 protein expression levels were significantly upregulated
in the LPS/ENR group compared with the control group (P<0.01;
Fig. 3E and F). The results also revealed that Bax
protein expression was notably increased in the LPS/ENR group,
while this was significantly reversed by CAG treatment (25, 200 and
400 µg/ml) for 24 h prior to LPS/ENR treatment (P<0.01; Fig. 3G). Caspase-3 activity was further
examined using a colorimetric assay; the results demonstrated that
24 h exposure to LPS/ENR caused a significant increase in caspase-3
activity compared with the control (P<0.01). In addition, when
the hepatocytes were exposed to CAG (50-400 µg/ml) the caspase-3
activity significantly decreased compared with the LPS/ENR group
(all P<0.01; Fig.
3H).
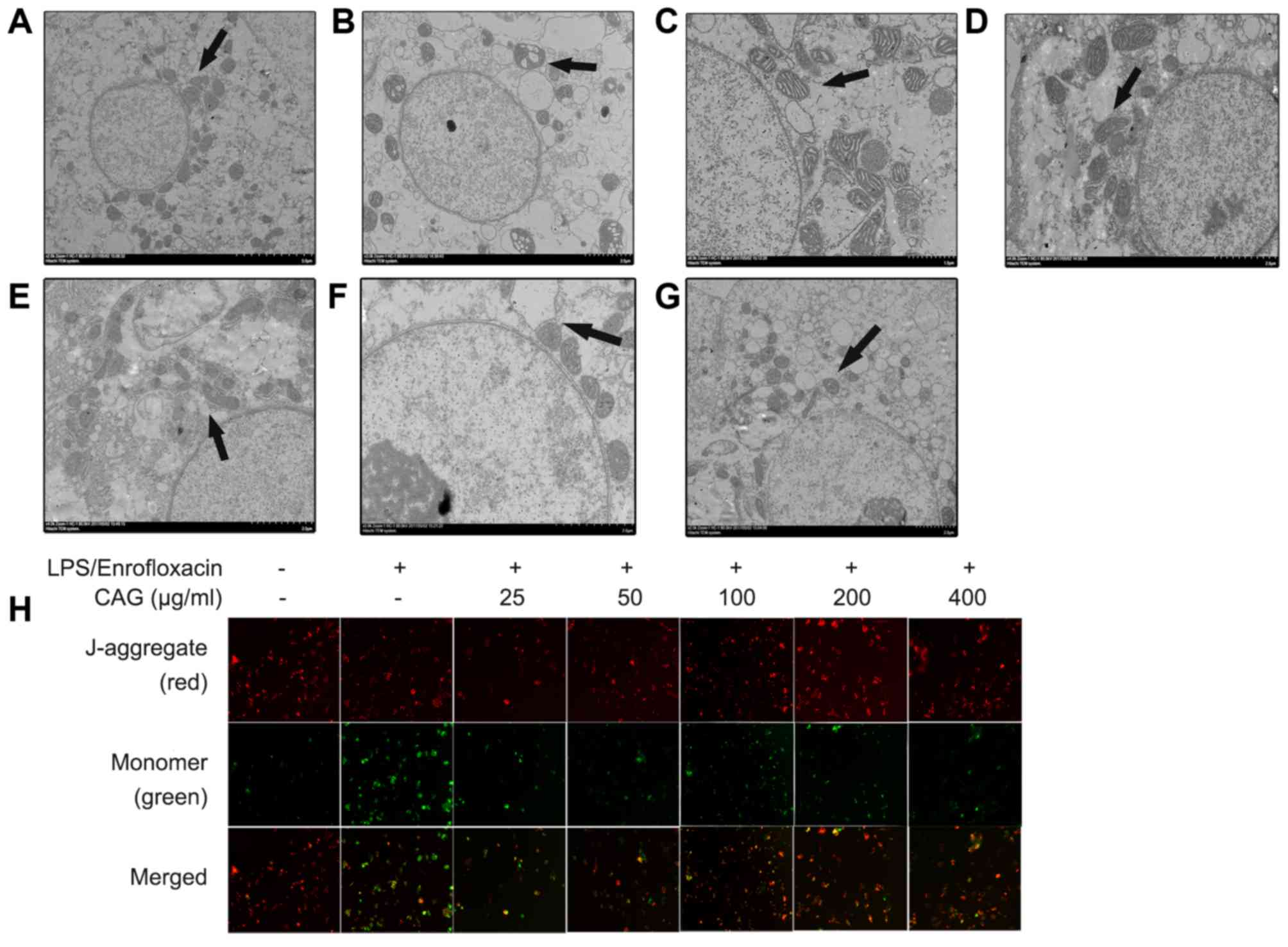
Alterations in mitochondrial
ultrastructure and MMP
Mitochondria serve an important role in the
apoptotic-signaling pathway (26).
In the present study, normal ultrastructure features were observed
in the control group, including a round nucleus, nuclear membrane
integrity and normal mitochondrial cristae (Fig. 4A). In the LPS/ENR treatment group,
the mitochondria were swollen with clear vacuolization and loss of
the crest (Fig. 4B). Following
treatment with CAG (25-400 µm/ml) for 24 h these modifications were
alleviated (Fig. 4C-G). The MMP was
also investigated using JC-1 dye. In normal mitochondria, JC-1
accumulates in the mitochondrial matrix to form polymers and the
polymer emits intense red fluorescence. Unhealthy mitochondria may
be present in the cytoplasm in monomeric form due to a decrease or
loss of membrane potential and produces green fluorescence
(10). In the model group green
fluorescence was increased compared with red fluorescence. However,
in the CAG group, red fluorescence was increased compared with
green fluorescence (Fig. 4H).
Discussion
LPS- and drug-induced liver toxicity are common
causes of liver injury (1). Common
compound liver injury models use Bacille Calmette Guerin and LPS or
D-galactosamine and LPS (1) and to
the best of the authors' knowledge, there are no previous reports
investigating antimicrobial drugs in combination with LPS-induced
liver injury. In the present study, primary cells were separated
using a modified two-step collagenase type IV ex-situ
perfusion method. Primary hepatocytes displayed a high level of
viability and purity. Unpublished results suggested the LPS and ENR
doses required to induce hepatocyte injury in addition to their
respective IC50 values, with LPS at 60 µg/ml and ENR at
180 µg/ml. LPS/ENR treatment resulted in a dose-dependent increase
in cell death; therefore a lower concentration of each component
was used than required for modeling separately. In the present
study it was observed that CAG relieved liver injury. Preliminary
experiments demonstrated that LPS/ENR treatment significantly
decreased cell viability and increased ALT and AST levels in the
supernatant, and treatment with CAG improved these indicators.
These results indicate that LPS/ENR induced in vitro liver
injury and CAG may represent a promising therapeutic tool for the
treatment of acute hepatic damage. Further investigation was also
performed into the mechanisms behind LPS/ENR hepatotoxicity and the
hepatoprotective effects of CAG, including hepatocyte apoptosis,
hepatic oxidative stress and the expression of apoptosis-associated
genes.
Protective effects on hepatocytes may be associated
with antioxidant capacity via scavenging of reactive oxygen species
(ROS) (27). The balance of
intracellular ROS depends on the production of aerobic metabolism
in normal cells in addition to non-enzymatic substances, such as
MDA and enzyme antioxidants, including GSH, SOD, CAT and GPx
(28). In the present study, high
levels of MDA were collected from cell lysates in the model group,
which indicated that LPS/ENR triggered the oxidation of liver cell
membranes. The results demonstrated that CAG has the ability to
restore lipid peroxidation of liver cells induced by LPS/ENR. It
has been previously established that cellular antioxidant enzymes
are the most important biomolecules for the prevention of oxidative
stress (29). Antioxidants balance
the cellular antioxidant system by inhibiting free radical
production, thereby preventing toxin-induced hepatocyte injury
(30). A previous study
demonstrated that glycyrrhizin is effective at inhibiting lipid
peroxidation and enhancing the capacity to eliminate free-radicals,
revealing the antioxidative effects of CAG (20). The present study has demonstrated
that LPS/ENR may initiate ROS overproduction and cause a reduction
in the cellular anti-oxidant enzymes GSH, SOD, CAT and GPx in
hepatocytes. The results also revealed that LPS/ENR treatment
reduced antioxidant enzyme activity and increased lipid
peroxidation. CAG pretreatment increased antioxidant enzyme
levels.
Apoptosis is essential for maintaining multicellular
organism development and the stability of the internal environment
by eliminating excess or unwanted cells (31). Under pathological conditions
(including chemically-induced lesions), the apoptotic balance may
be disrupted, which leads to excessive and persistent apoptosis
(32). Hepatocyte apoptosis has
been revealed to serve an important role in viral and
non-viral-induced acute liver injury (33,34).
The positive effect of CAG may be due to its ability to scavenge
free radicals. Hepatocyte apoptosis is the most important molecular
mechanism associated with liver failure (35,36).
Therefore, exploring the underlying mechanisms of apoptosis
provides a basis for LPS/ENR-induced liver injury. Animal
experiments have demonstrated that hepatocyte apoptosis
significantly increased following LPS treatment (37). Conversely, glycyrrhizin was revealed
to inhibit apoptosis in CCl4-induced liver injury in
rats (38). In the present study,
the results demonstrated an increase in the number of apoptotic
hepatocytes in the LPS/ENR-treated group; these cells primarily
comprised early apoptotic cells. In the CAG-treated group the
number of early apoptotic cells decreased in a dose-dependent
manner, indicating that CAG relieved liver injury caused by
LPS/ENR.
The current study focused primarily on a variety of
genes associated with cell death and the protease cascade system.
Caspase-3 is an apoptosis effector, which has been reported to be a
key factor in the apoptotic process (39). In addition, following examination of
apoptotic chromatin condensation and DNA fragmentation, which are
essential for the process of disassembling cells and the formation
of apoptotic bodies, it was revealed that caspase-3 was required
(40). The results of the present
study demonstrated that mRNA and caspase-3 activity increased
following treatment with LPS/ENR, while CAG pretreatment
significantly attenuated caspase-3 activity and its mRNA levels. In
addition, mitochondria were observed to be swollen and vacuolated
in the model group as revealed by TEM. The effect of LPS/ENR on MMP
will be included in further studies.
The Bcl-2 family contains pro-apoptotic (Bax and
Bid) and anti-apoptotic (Bcl-2 and Bcl-xl) proteins, which regulate
the mitochondrial apoptotic signaling pathway (41). In addition, cell survival and death
may be regulated by blocking death receptor formation and binding
with the mitochondrial outer membrane (42). Bcl-2 and Bax have opposite effects
on cell death; when Bcl-2 is higher than Bax it causes the
inhibition of cell death, whereas Bax levels higher than Bcl-2
cause the acceleration of apoptosis (43). The results of the present study
suggest that the Bcl-2 mRNA level was decreased in the
LPS/ENR-treatment group results of the present study indicated that
the protective effect of CAG against LPS/ENR-induced liver injury
may be partly mediated by regulation of the apoptotic signaling
pathway.
In conclusion, the present study demonstrated that
ENR in combination with LPS exacerbate toxicity in chicken
hepatocytes, whereas CAG relieves this toxicity. Therefore, CAG may
have potential as a novel prophylactic agent for the treatment of
hepatic liver disease. However, further studies are required to
confirm the clinical viability of this treatment.
Acknowledgements
Not applicable.
Funding
The present study was supported by the National
Natural Science Foundation (grant no. 31572569).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
XG and ZY conceived and designed the experiments. WL
and RA performed the experiments. WL, ZY and MH analyzed the data.
WL drafted and revised the manuscript. All authors read and
approved the final manuscript.
Ethics approval and consent to
participate
The present study was approved by the Animal Ethics
Committee of Nanjing Agricultural University (IACUC).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Xia X, Su C, Fu J, Zhang P, Jiang X, Xu D,
Hu L, Song E and Song Y: Role of α-lipoic acid in LPS/d-GalN
induced fulminant hepatic failure in mice: Studies on oxidative
stress, inflammation and apoptosis. Int Immunopharmacol.
22:293–302. 2014.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Jeong YI, Jung ID, Lee CM, Chang JH, Chun
SH, Noh KT, Jeong SK, Shin YK, Lee WS, Kang MS, et al: The novel
role of platelet-activating factor in protecting mice against
lipopolysaccharide-induced endotoxic shock. PLoS One.
4(e6503)2009.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Thirunavukkarasu C, Uemura T, Wang LF,
Watkins SC and Gandhi CR: Normal rat hepatic stellate cells respond
to endotoxin in LBP-independent manner to produce inhibitor(s) of
DNA synthesis in hepatocytes. J Cell Physiol. 204:654–665.
2005.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Kaplowitz N: Drug-induced liver disorders:
Implications for drug development and regulation. Drug Saf.
24:483–490. 2001.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Hirashima R, Itoh T, Tukey RH and Fujiwara
R: Prediction of drug-induced liver injury using keratinocytes. J
Appl Toxicol. 37:863–872. 2017.PubMed/NCBI View
Article : Google Scholar
|
|
6
|
Ghabril M, Chalasani N and Bjornsson E:
Drug-induced liver injury: A clinical update. Curr Opin
Gastroenterol. 26:222–226. 2010.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Leitner JM, Graninger W and Thalhammer F:
Hepatotoxicity of antibacterials: Pathomechanisms and clinical.
Infection. 38:3–11. 2010.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Yilmaz B, Ekiz F, Coban S, Yüksel I and
Yüksel O: Cefixime-induced hepatotoxicity. Turk J Gastroenterol.
22(445)2011.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Orman ES, Conjeevaram HS, Vuppalanchi R,
Freston JW, Rochon J, Kleiner DE and Hayashi PH: DILIN Research
Group: Clinical and histopathologic features of
fluoroquinolone-induced liver injury. Clin Gastroenterol Hepatol.
9:517–523 e3. 2011.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Liu B, Cui Y, Brown PB, Ge X, Xie J and Xu
P: Cytotoxic effects and apoptosis induction of enrofloxacin in
hepatic cell line of grass carp (Ctenopharyngodon idellus). Fish
Shellfish Immunol. 47:639–644. 2015.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Kalpana S, Aggarwal M, Srinivasa Rao G and
Malik JK: Effects of aflatoxin B1 on tissue residues of
enrofloxacin and its metabolite ciprofloxacin in broiler chickens.
Environ Toxicol Pharmacol. 33:121–126. 2012.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Shinozuka Y, Uematsu K, Takagi M and Taura
Y: Comparison of the amounts of endotoxin released from Escherichia
coli after exposure to antibiotics and ozone: An in vitro
evaluation. J Vet Med Sci. 70:419–422. 2008.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Oh HM, Lee S, Park YN, Choi EJ, Choi JY,
Kim JA, Kweon JH, Han WC, Choi SC, Han JK, et al: Ammonium
glycyrrhizinate protects gastric epithelial cells from hydrogen
peroxide-induced cell death. Exp Biol Med (Maywood). 234:263–277.
2009.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Sato H, Goto W, Yamamura J, Kurokawa M,
Kageyama S, Takahara T, Watanabe A and Shiraki K: Therapeutic basis
of glycyrrhizin on chronic hepatitis B. Antiviral Res. 30:171–177.
1996.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Yamamura Y, Kotaki H, Tanaka N, Aikawa T,
Sawada Y and Iga T: The pharmacokinetics of glycyrrhizin and its
restorative effect on hepatic function in patients with chronic
hepatitis and in chronically carbon-tetrachloride-intoxicated rats.
Biopharm Drug Dispos. 18:717–725. 1997.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Lin JC, Cherng JM, Hung MS, Baltina LA,
Baltina L and Kondratenko R: Inhibitory effects of some derivatives
of glycyrrhizic acid against Epstein-Barr virus infection:
Structure-activity relationships. Antiviral Res. 79:6–11.
2008.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Fiore C, Eisenhut M, Krausse R, Ragazzi E,
Pellati D, Armanini D and Bielenberg J: Antiviral effects of
glycyrrhiza species. Phytother Res. 22:141–148. 2008.PubMed/NCBI View
Article : Google Scholar
|
|
18
|
Seeff LB, Lindsay KL, Bacon BR, Kresina TF
and Hoofnagle JH: Complementary and alternative medicine in chronic
liver disease. Hepatology. 34:595–603. 2001.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Stickel F and Schuppan D: Herbal medicine
in the treatment of liver diseases. Dig Liver Dis. 39:293–304.
2007.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Visavadiya NP and Narasimhacharya AV:
Hypocholesterolaemic and antioxidant effects of glycyrrhiza glabra
(Linn) in rats. Mol Nutr Food Res. 50:1080–1086. 2006.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Racková L, Jancinová V, Petríková M,
Drábiková K, Nosál R, Stefek M, Kostálová D, Prónayová N and
Kovácová M: Mechanism of anti-inflammatory action of liquorice
extract and glycyrrhizin. Nat Prod Res. 21:1234–1241.
2007.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Yim SB, Park SE and Lee CS: Protective
effect of glycyrrhizin on 1-methyl-4-phenylpyridinium-induced
mitochondrial damage and cell death in differentiated PC12 cells. J
Pharmacol Exp Ther. 321:816–822. 2007.PubMed/NCBI View Article : Google Scholar
|
|
23
|
LaBrecque DR and Howard RB: The
preparation and characterization of intact isolated parenchymal
cells from rat liver. Methods Cell Biol. 14:327–340.
1976.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Morikawa A, Sugiyama T, Kato Y, Koide N,
Jiang GZ, Takahashi K, Tamada Y and Yokochi T: Apoptotic cell death
in the response of D-galactosamine-sensitized mice to
lipopolysaccharide as an experimental endotoxic shock model. Infect
Immun. 64:734–738. 1996.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Shiota M, Sugai N, Tamura M, Yamaguchi R,
Fukushima N, Miyano T and Miyazaki H: Correlation of
mitogen-activated protein kinase activities with cell survival and
apoptosis in porcine granulosa cells. Zoolog Sci. 20:193–201.
2003.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Szeto HH: Mitochondria-targeted peptide
antioxidants: Novel neuroprotective agents. AAPS J. 8:E521–E531.
2006.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Lowry OH, Rosebrough NJ, Farr AL and
Randall RJ: Protein measurement with the Folin phenol reagent. J
Biol Chem. 193:265–275. 1951.PubMed/NCBI
|
|
29
|
Mohammadi M and Yazdanparast R: Methoxy
VO-salen complex: In vitro antioxidant activity, cytotoxicity
evaluation and protective effect on CCl4-induced oxidative stress
in rats. Food Chem Toxicol. 47:716–721. 2009.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Song E, Fu J, Xia X, Su C and Song Y:
Bazhen decoction protects against acetaminophen induced acute liver
injury by inhibiting oxidative stress, inflammation and apoptosis
in mice. PLoS One. 9(e107405)2014.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Srivastava M, Ma LQ, Singh N and Singh S:
Antioxidant responses of hyper-accumulator and sensitive fern
species to arsenic. J Exp Bot. 56:1335–1342. 2005.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Shamsabadi FT, Eidgahi MR, Mehrbod P,
Daneshvar N, Allaudin ZN, Yamchi A and Shahbazi M: Survivin, a
promising gene for targeted cancer treatment. Asian Pac J Cancer
Prev. 17:3711–3719. 2016.PubMed/NCBI
|
|
33
|
Danial NN and Korsmeyer SJ: Cell death:
Critical control points. Cell. 116:205–219. 2004.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Jaeschke H, Gujral JS and Bajt ML:
Apoptosis and necrosis in liver disease. Liver Int. 24:85–89.
2004.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Kasahara I, Saitoh K and Nakamura K:
Apoptosis in acute hepatic failure: Histopathological study of
human liver tissue using the tunel method and immunohistochemistry.
J Med Dent Sci. 47:167–175. 2000.PubMed/NCBI
|
|
36
|
Eichhorst ST: Modulation of apoptosis as a
target for liver disease. Expert Opin Ther Targets. 9:83–99.
2005.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Togo S, Kubota T, Matsuo K, Shimizu T,
Momiyama N, Takeda K, Tanaka K, Endo I, Sekido H and Shimada H:
Mechanism of liver failure after hepatectomy. Nihon Geka Gakkai
Zasshi. 105:658–663. 2004.(In Japanese). PubMed/NCBI
|
|
38
|
Liu LM, Zhang JX, Luo J, Guo HX, Deng H,
Chen JY and Sun SL: A role of cell apoptosis in lipopolysaccharide
(LPS)-induced nonlethal liver injury in D-galactosamine
(D-GalN)-sensitized rats. Dig Dis Sci. 53:1316–1324.
2008.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Wang XJ, Kong KM, Qi WL, Ye WL and Song
PS: Interleukin-1 beta induction of neuron apoptosis depends on p38
mitogen-activated protein kinase activity after spinal cord injury.
Acta Pharmacol Sin. 26:934–942. 2005.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Guo XL, Liang B, Wang XW, Fan FG, Jin J,
Lan R, Yang JH, Wang XC, Jin L and Cao Q: Glycyrrhizic acid
attenuates CCl4-induced hepatocyte apoptosis in rats via
a p53-mediated pathway. World J Gastroenterol. 19:3781–3791.
2013.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Chen XC, Zhu YG, Wang XZ, Zhu LA and Huang
C: Protective effect of ginsenoside Rg1 on dopamine-induced
apoptosis in PC12 cells. Acta Pharmacol Sin. 22:673–678.
2001.PubMed/NCBI
|
|
42
|
Porter AG and Jänicke RU: Emerging roles
of caspase-3 in apoptosis. Cell Death Differ. 6:99–104.
1999.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Wolter KG, Hsu YT, Smith CL, Nechushtan A,
Xi XG and Youle RJ: Movement of Bax from the cytosol to
mitochondria during apoptosis. J Cell Biol. 139:1281–1292.
1997.PubMed/NCBI View Article : Google Scholar
|