Introduction
The incidence of RAS gene mutations in all
known human cancers is ~30% worldwide over the last 30 years
(1). Although the mutation in the
RAS gene is not the direct cause of all cancers, it has been
reported to promote tumor occurrence, development and metastasis
(2). Ras protein GTPase activity
was reported to serve a key role in the process of human
carcinogenesis more than 30 years ago (3,4).
Furthermore, the RAS gene has been demonstrated to be one of
the most mutated oncogenes in humans and to promote further
development of tumors (5). A
previous study demonstrated that mutant RAS gene stimulated
cell proliferation and inhibited cell apoptosis, thereby promoting
tumor occurrence, survival, migration, diffusion and angiogenesis
(6). Therefore, Ras protein is used
as a target for drug treatment; however, due to the complexity of
the Ras signaling pathway and its unique structure, drugs cannot
actively target Ras to bind directly of the protein (7).
In recent years, naphthoquinone derivatives have
become a focus of interest due to their ability to effectively
cause death in cancer cells, including those in lung cancer
(8), breast cancer (9) and colon cancer (10). However, naphthoquinone reportedly
exerts low cytotoxic effects on healthy cells compared with
currently used, existing clinical drugs, including 5-fluorouracil,
cisplatin and sorafenib (11).
Naphtoquinone derivatives are primarily constructed through
multiple additions and substitutions with shikonin as the primary
substrate to obtain a variety of derivatives with different
structures (12). The
5,8-dimethoxy-1,4-naphthoquinone (DMNQ) compound is commonly used
to synthesize naphthoquinone derivatives. It has been reported that
DMNQ derivatives can inhibit tumor activity in lung (13) and breast cancers (14). Since each derivative has different
substituents, the resulting functions of each derivative are
subsequently different (15).
According to previous research, naphthoquinone derivatives
primarily aim to target two different physiological processes to
cause cell death: i) To induce cytotoxicity by increasing
intracellular reactive oxygen species (ROS) levels, which causes
damage to mitochondrial function in cells, endoplasmic reticulum
stress and autophagy that lead to cell death (16); and ii) to directly cause
cytotoxicity, DNA strand breakage and cell cycle arrest (17). In hepatocarcinoma cells,
naphthoquinone derivatives were reported to suppress
hepatocarcinoma liver cancer cell proliferation, inhibit
angiogenesis and promote cellular apoptosis by inhibiting DNA
synthesis (18,19). Research into drugs targeting Ras has
made progress (20). Certain
research demonstrated that 2,3-dichloro-1,4-naphthoquinone
inhibited the growth of N-Ras-mutant melanoma (21), while
2-(2',4'-dihydroxyphenyl)-8-hydroxy-1,4-naphthoquinone and
5-hydroxy-2-(2,4-dihydroxyphenyl)naphthalene-1,4-dione inhibited
Apc/K-Ras-mutant mouse organoid differentiation capacity (22). These results indicate that
naphthoquinone derivatives have a certain inhibitory effect on the
proliferation and differentiation of cancer cells and Ras
signaling.
Therefore, the present study used shikonin as the
main backbone to which to add different substituents and to design
a variety of naphtoquinone derivatives, to select and investigate
the anticancer mechanism of drugs with lower cytotoxicity for
normal cells and higher lethality for cancer cells. Furthermore,
the present study aimed to establish novel structural modification
sites, while retaining the structure of derivatives for targeting
Ras signaling to maximize the anticancer effect and reduce
cytotoxicity, which may have practical significance for the
development of targeted drugs.
Materials and methods
Materials
Solvents of reagent grade and were used without
further purification. 1H NMR and 13C NMR
spectra were recorded using a JNM-AL 400 (400 MHz) spectrometer,
Chemical shift ppm (δ) were relative to internal standard
tetramethylsilane. MS spectra were recorded with an AB SCIEX API
2000 LC/MS/MS (Applied Biosystems; Thermo Fisher Scientific, Inc.)
and LCMS-IT-TOF (Shimadzu Corporation).
Synthesis of
5,8-dimethoxy-1,4-naphthoquinone (DMNQ) derivatives
Synthetic schemes for 12 DMNQ derivatives are
summarized in Fig. 1.
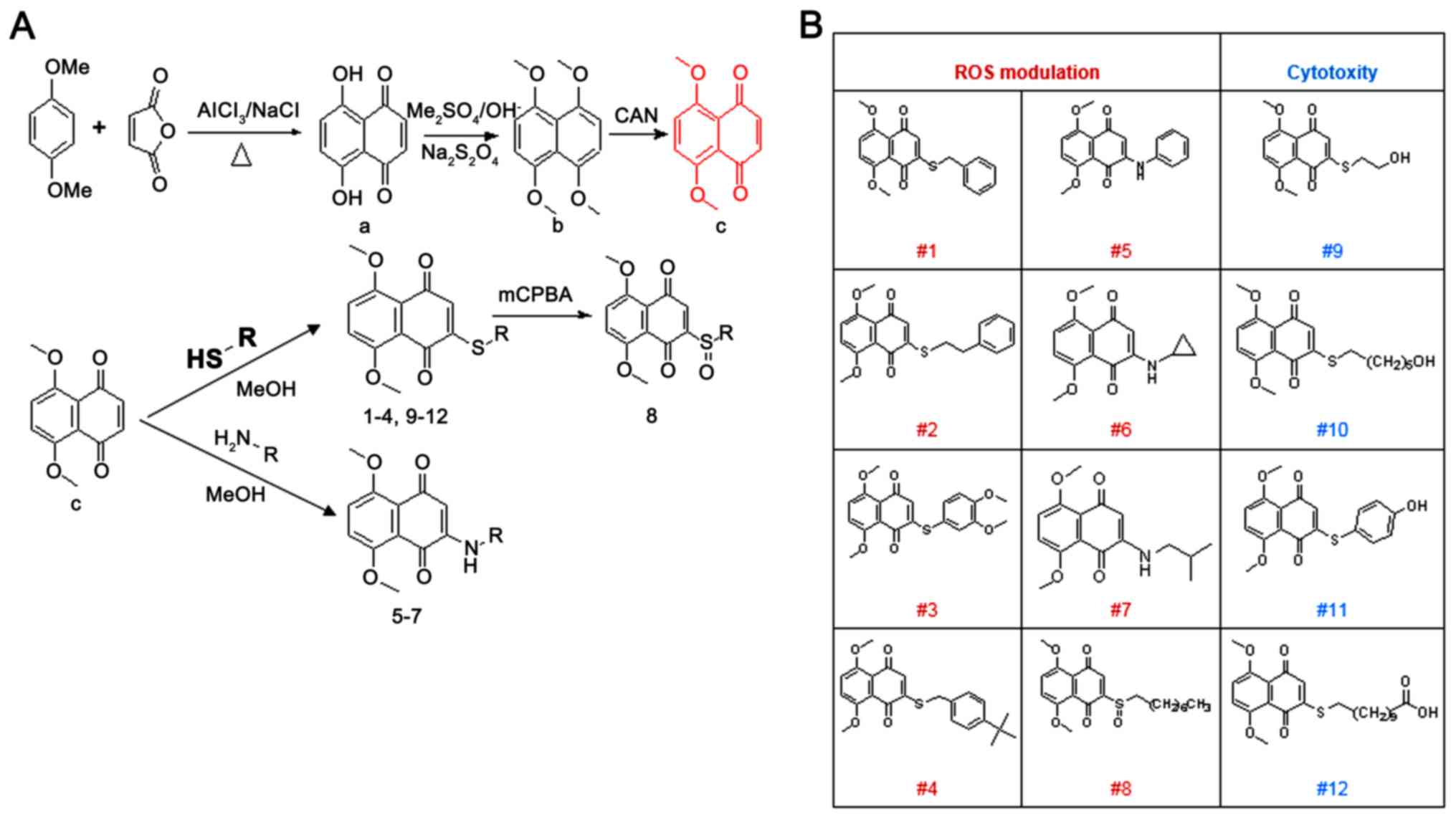 | Figure 1Synthetic schemes and structures of
the 12 different 5,8-dimethoxy-1,4-naphthoquinone derivatives. (A)
Synthetic schemes of 5,8-dimethoxy-1,4-naphthoquinone derivatives.
Compounds were synthesized by condensation, methylation, oxidation,
and Michael addition reactions. (B) structures of 12 different
5,8-dimethoxy-1,4-naphthoquinone derivatives. OMe, methoxy; MeOH,
Methanol; (CH2)5OH, hydroxy pentyl;
(CH2)6CH3, heptyl;
(CH2)9, nonyl;O, oxygen;
AlCl3, aluminum chloride; NaCl, sodium chloride; OH,
hydroxide; Me2SO4, dimethyl sulfate;
Na2S2O4, sodium dithionite; CAN,
Cerium (IV) diammonium nitrate; mCPBA, meta-Chloroperoxybenzoic
acid; S, sulphur; HS, hydrosulfide; R, functional group;
H2N, nitrogen hydride; NH, amine; ROS, reactive oxygen
species. |
Synthesis of
5,8-dihydoroxy-1,4-naphthoquinone
1,4-Dimethoxybenzene (1 equivalent) (Sigma-Aldrich;
Merck KGaA) and maleic anhydride (2 equivalent) (Sigma-Aldrich;
Merck KGaA) were added to the molten material of NaCl (4
equivalent) (Tianjin Damao Chemical Reagent Co., Ltd.) and aluminum
chloride (8 equivalent, 171-177˚C) (Tianjin Damao Chemical Reagent
Co., Ltd.) to give the condensed mixture. The derivatives were then
treated with 1.2 liters H2O and 120 ml 95% HCl (Tianjin
Damao Chemical Reagent Co., Ltd.) to generate DMNQ (Fig. 1A-a).
Synthesis of
1,4,5,8-tetramethoxynaphthalene
A solution of 25 ml methyl p-toluene
sulfonate (Sigma-Aldrich; Merck KGaA), 10.6 g sodium dithionite
(Tianjin Damao Chemical Reagent Co., Ltd.) in 40 ml 70% NaOH
(Tianjin Damao Chemical Reagent Co., Ltd.) was added to a solution
of 20.1 g (Fig. 1A-a) in 200 ml
tetrahydrofuran (Tianjin Damao Chemical Reagent Co., Ltd.), 220 ml
H2O and 2.2 g tetrabutylammonium bromide (Sigma-Aldrich;
Merck KGaA). The mixture was stirred at 25˚C for 20 h until a red
precipitate formed, which was filtered and recrystallized in
petroleum ether (Tianjin Damao Chemical Reagent Co., Ltd.)
(90-120˚C) to produce an intermediate
1,4,5,8-tetramethoxynaphthalene product (Fig. 1A-b).
Synthesis of DMNQ
An amount 5.5 g cerium (IV) diammonium nitrate
(Tianjin Damao Chemical Reagent Co., Ltd.) was dissolved in 250 ml
H2O and added drop-wise to a solution of 15 g
1,4,5,8-tetramethoxynaphthalene in 160 ml trichloromethane (Tianjin
Damao Chemical Reagent Co., Ltd.) and 480 ml acetonitrile (Tianjin
Damao Chemical Reagent Co., Ltd.). This mixture was stirred at 25˚C
for 1.5 h. The key compound DMNQ (Fig.
1A-c) was obtained by this oxidation reaction.
Synthesis of the DMNQ derivatives
#1
DMNQ solution (300 mg; 1.37 mmol) in 40 ml ethyl
alcohol (EtOH, Tianjin Damao Chemical Reagent Co., Ltd.) was added
to benzyl mercaptan (204.9 mg; 1.65 mmol; Sigma-Aldrich; Merck
KGaA; Fig. 1B-#1) for derivative
#1, 2-phenylethyl mercaptan (228.1 mg; 1.65 mmol; Sigma-Aldrich;
Merck KGaA; Fig. 1B-#2) for
derivative #2, 3,4-dimethoxybenzenethiol (280.9 mg; 1.65 mmol;
Sigma-Aldrich; Merck KGaA; Fig.
1B-#3) for derivative #3, 4-tert-butylphenylmethanethiol
(297.5 mg; 1.65 mmol; Sigma-Aldrich; Merck KGaA; Fig. 1B-#4) for derivative #4, phenyl amine
(148.7 mg; 1.65 mmol; Sigma-Aldrich; Merck KGaA; Fig. 1B-#5) for derivative #5,
cyclo-propyl amine (84.0 mg; 1.65 mmol; Sigma-Aldrich; Merck
KGaA; Fig. 1B-#6) for derivative
#6, 2-methylpropan-1-amine (120.6 mg; 1.65 mmol; Sigma-Aldrich;
Merck KGaA; Fig. 1B-#7) for
derivative #7, 2-mercaptoethanol (128.9 mg; 1.65 mmol;
Sigma-Aldrich; Merck KGaA; Fig.
1B-#9) for derivative #9, 6-mercaptohexan-1-ol (221.5 mg; 1.65
mmol; Sigma-Aldrich; Merck KGaA; Fig.
1B-#10) for derivative #10, 4-mercaptophenol (208.2 mg; 1.65
mmol; Sigma-Aldrich; Merck KGaA; Fig.
1B-#11) for derivative #11 and 10-mercaptodecanoic acid (337.1
mg; 1.65 mmol; Sigma-Aldrich; Merck KGaA; Fig. 1B-#12) for derivative #12. The
reaction mixture was stirred at room temperature for 4.5 h. The
resulting solution was added dropwise to a solution of 0.24 mmol
sodium dichromate (Tianjin Damao Chemical Reagent Co., Ltd.) and
0.77 mmol sulfuric acid (Tianjin Damao Chemical Reagent Co., Ltd.)
in 10 ml H2O. The resulting mixture was stirred for a
few minutes, extracted with methylene chloride
(CH2Cl2; Tianjin Damao Chemical Reagent Co.,
Ltd.), concentrated and recrystallized in EtOH to generate the
target compound #1 (Fig.
1B-#1).
Synthesis of DMNQ derivative #8
2-Heptyltho-DMNQ solution (300 mg, 1.37 mmol) in 40
ml dichloromethane was added to 1.65 mmol 3-chloroperoxybenzoic
acid (Shanghai Aladdin Bio-Technology Co., Ltd.). The reaction
mixture was stirred at room temperature for 2.5 h. The resulting
solution was added to 40 ml saturated sodium chloride solution. The
resulting mixture was stirred for a few minutes, extracted with
CH2Cl2, concentrated and recrystallized in
EtOH to produce the target compound #8 (Fig. 1B-#8).
The structures of compounds #1 were characterized by
nuclear magnetic resonance (NMR) and mass spectroscopy.
Chemicals
The following chemicals were used in the current
study: DMEM (Invitrogen; Thermo Fisher Scientific, Inc.), FBS
(Cytiva), penicillin (100 U/ml) and streptomycin (100 mg/ml;
Beijing Solarbio Science & Technology, Co., Ltd.), MTT
(Sigma-Aldrich; Merck KGaA), an Annexin V-FITC and propidium iodide
(PI) detection kit (Beijing Solarbio Science & Technology, Co.,
Ltd.), dihydroethidium (DHE; Beyotime Institute of Biotechnology),
Hoechst 32258 (Thermo Fisher Scientific, Inc.), N-acetyl-L-cysteine
(NAC; Beijing Solarbio Science & Technology, Co., Ltd.),
horseradish peroxidase-conjugated goat anti-rabbit immunoglobulin
(Ig)G and anti-mouse IgG (Sangon Biotech, Co., Ltd.).
Cell culture
The following cell lines were used: NIH3T3 mouse
embryonic fibroblasts (NC), H-RasG12V-transfected NIH3T3
mouse embryonic fibroblasts (NR), HepG2 human liver cancer (HC)
cells and H-RasG12V-transfected HepG2 human liver cancer
cells (HR). All cell lines were provided by Dr Dae-Yeul Yu (Aging
Intervention Research Center, Korea Research Institute of
Bioscience and Biotechnology, Daejeon, Korea). Cells were all
cultured in DMEM with 10% (v/v) FBS and maintained in a culture
environment of 37˚C and 5% CO2. The culture medium was
replaced once a day and cells were sub-cultured every 3 days. Cells
were treated with 5 mM NAC for 30 min prior to treatment with 8 µM
BZNQ.
Cytotoxicity analysis of the 12
different 5,8-dimethoxy-1,4-naphthoquinone derivatives
NC and NR cells were seeded on 96-well plates
(10,000 cells/well) and cultured for 24 h at 37˚C in the presence
of the 12 different 5,8-dimethoxy-1,4-naphthoquinone derivatives
(0, 10, 25 and 50 µM). A total of 10 µl (0.5 mg/ml) MTT reagent was
added and absorbance was measured at a wavelength of 490 nm using a
UV MAX kinetic microplate reader (Molecular Devices, LLC.) to
determine the accumulation of formazan. The purple formazan was
subsequently dissolved using DMSO. The IC50 values were
obtained using SigmaPlot Version 10.0 software (Systat Software
Inc.).
Cell viability assay
MTT assays were used to analyze cell viability. NC,
NR, HC and HR cells were seeded on 96-well plates (seeding density,
10,000 cells/well) and cultured for 24 h at 37˚C in the presence of
2-benzylthio-5,8-dimethoxynaphthalene-1,4-dione (BZNQ) at
concentrations of 0, 2, 4, 6, 8 and 10 µM in NC and NR cells. For
HC and HR cells, BZNQ was administered at 0, 2, 4, 6, 8, 10 and 20
µM. The 0 µM (untreated) group was used as the control. Following
this, 10 µl (0.5 mg/ml) of MTT reagent was added and the absorbance
was measured at the wavelength of 490 nm using a UV MAX kinetic
microplate reader (Molecular Devices, LLC.) to determine the
accumulation of formazan. The MTT reagent was dissolved with
DMSO.
Early and late apoptosis detection
using Annexin V-FITC/PI
NC, NR, HC and HR cells treated with BZNQ were
harvested by trypsin and then suspended in PBS. Apoptosis was
measured by staining with Annexin V-FITC/PI for 15 min at room
temperature. The number and proportion of positively-labeled cells
were analyzed by flow cytometry (FACSCalibur; BD Biosciences). The
results were analyzed using the WinMDI software (version 2.9; BD
Biosciences).
Detection of ROS production
HC and HR cells were treated with BZNQ for 12 h.
Cells were subsequently fixed with 4% paraformaldehyde for 10 min
at room temperature. The changes in cellular ROS levels were
determined using 5 µM DHE for 15 min at 37˚C. Nuclei were
visualized using Hoechst 32258 staining. Nuclear staining was
observed qualitatively under a fluorescence microscopy
(EVOS®xl core cell culture microscope; Advanced
Microscopy Group) following 20 min of incubation with the Hoechst
dye at 37˚C.
Western blotting
For the isolation of proteins, cells was lysed in
ice-cold cell extraction buffer [20 mM HEPES (pH 7.0), 50 mM NaCl,
10% Triton X-100, 10% Glycerol, 1 mM β-ME and a protease inhibitor
cocktail tablet; Roche Diagnostics GmbH] for 30 min. Lysates were
centrifugated at 13,200 x g and 4˚C for 20 min. Total protein (2
µg/µl) quantification was determined by a Bradford protein assay
using an ultraviolet spectrophotometer (Aoyi Instruments Co.,
Ltd.). Denatured protein of cell lysates were separated using 12%
SDS-PAGE and transferred to nitrocellulose membranes (EMD
Millipore). The membranes were blocked with 5% skim milk (BD
Biosciences) for 1 h at room temperature. The membranes were
incubated with primary antibodies against H-Ras (cat. no bs-1071;
BIOSS; 1:1,000), protein kinase B (AKT; cat. no. sc-8044; Santa
Cruz Biotechnology, Inc.; 1:2,000), p-AKT (cat. no. sc-101629;
Santa Cruz Biotechnology, Inc.; 1:2,000), extracellular regulated
kinase (ERK; cat. no. sc-135900; Santa Cruz Biotechnology, Inc.;
1:2,000), p-ERK (cat. no. sc-7383; Santa Cruz Biotechnology, Inc.;
1:2,000), glycogen synthase kinase 3β (GSK3β; cat. no. AF1543;
Beyotime Institute of Biotechnology, Inc.; 1:2,000). p-GSK3β (Ser9;
cat. no. AF5830; Beyotime Institute of Biotechnology, Inc.;
1:2,000), β-catenin (cat. no. AF1189; Beyotime Institute of
Biotechnology, Inc.; 1:2,000), B-cell lymphoma-2 (Bcl-2; cat. no.
sc-7382; Santa Cruz Biotechnology, Inc.; 1:2,000), Bcl-2 associated
agonist of cell death (Bad; cat. no. 8044; Santa Cruz
Biotechnology, Inc.; 1:2,000), β-actin (cat. no. 3700; Cell
Signaling Technology, Inc.; dilution, 1:2,000) at 4˚C overnight.
The membranes were washed five times with TBST [10 mM Tris HCl (pH
7.5), 150 mM NaCl and 0.2% Tween-20] to remove excess antibodies
and then incubated for 1 h at room temperature with horseradish
peroxidase conjugated goat anti mouse and rabbit IgG secondary
antibodies(cat. no. D110087 and D110058; Shanghai Sangon Biotech
Co., Ltd.; 1:5,000). Bands of antibody-specific binding proteins
were visualized by a chemiluminescence detection system (GE
Healthcare) and densitometry was analyzed using ImageJ version
1.52a software (National institutes of Health).
Statistical analysis
Data are presented as the mean ± SEM from at least
three independent repeated experiments. Two-way ANOVA was used to
analyze the changes across time and differences between groups with
the Tukey's honestly significant difference test used as the
post-hoc test (α=0.05). P<0.05 was considered to indicate a
statistically significant difference.
Results
Effect of BZNQ on cell viability
According to the synthetic methodology provided
above, a total of 9 derivatives with a redox center S atom and 3
alkylamine derivatives that were able to clear ROS in cells were
designed (Fig. 1A). The structure
of each 5,8-dimethoxy-1,4-naphthoquinone derivative is provided in
Fig. 1B. The results indicated that
compounds #1-8 may be associated with ROS modulations, while
compounds #9-12 may be involved in cell cytotoxicity (Fig. 1B). To examine the cytotoxicity of
the synthesized compounds, the NC and NR NIH3T3 cells were treated
with derived compounds #1-12 and the concentrations of
IC50 for each compound were evaluated using MTT assays
(Table I). The results demonstrated
that at low concentrations, BZNQ exhibited higher cytotoxic
activity in both NC and NR cells compared with the rest of the
compounds. Compared with #11, #1 demonstrated lower dose targeting
to HRAS. Additionally, the lower dosage produced fewer side
effects. #1 was therefore selected for further study. Furthermore,
compounds #2-4 and #8-10 demonstrated significant activities in NC
and NR cells; however, there were no significant differences in
cytotoxicity effects between NC and NR cells (data not shown). The
results indicated that the different modulation of the substitution
groups at the specified positions exhibited various inhibiting
activities in cells. Therefore, the current study focused
subsequent experiments on BZNQ to elucidate the possible mechanisms
of action.
 | Table IIC50 values of the derived
compounds on NC and NR cells. |
Table I
IC50 values of the derived
compounds on NC and NR cells.
| Compound | NC | NR |
|---|
| #1 (BZNQ) | 12.40 | 4.73a |
| #2 | 39.31 | 35.86 |
| #3 | <10.00 | <10.00 |
| #4 | 16.42 | 24.37 |
| #5 | >50.00 | >50.00 |
| #6 | >50.00 | >50.00 |
| #7 | >100.00 | >100.00 |
| #8 | <1.00 | <1.00 |
| #9 | <1.00 | <1.00 |
| #10 | 5.37 | 7.53 |
| #11 | >30.00 | 21.52a |
| #12 | 22.50 | 12.39 |
BZNQ induces apoptosis in
Ras-transfected NIH3T3 cells
To investigate the structure of BZNQ, NMR spectra
were evaluated. According to the NMR spectra examination, the #1
compound as BZNQ: 1H-NMR (CDCl3, 400 MHz)
δ7.40-7.28 [multiplet (m), 5 hydrogen (H)], 6.98-6.80 (m, 2H), 5.99
[singlet (s), 1H] 3.94 (s, 6H), 3.63 (s, 2H); 13C NMR
(CDCl3, 100 MHz) δ 187.7 [C (carbon)-1], 180.7 (C-4),
154.3 (C-5), 154.1 (C-8), 153.7 (C-2), 135.2 (C-1'), 134.5 (C-3),
129.0 (C-3'), 129.0 (C-5'), 128.9 (C-2'), 128.9 (C-6'), 128.7
(C-4'), 128.1 (C-7), 127.5 (C-6), 121.1 (C-9), 119.6 (C-10), 56.9
(OCH3), 56.7 (OCH3), 35.4 (CH2)and
m/z 362.63 [Molecule (M)+Na]+ (Fig. 2A).
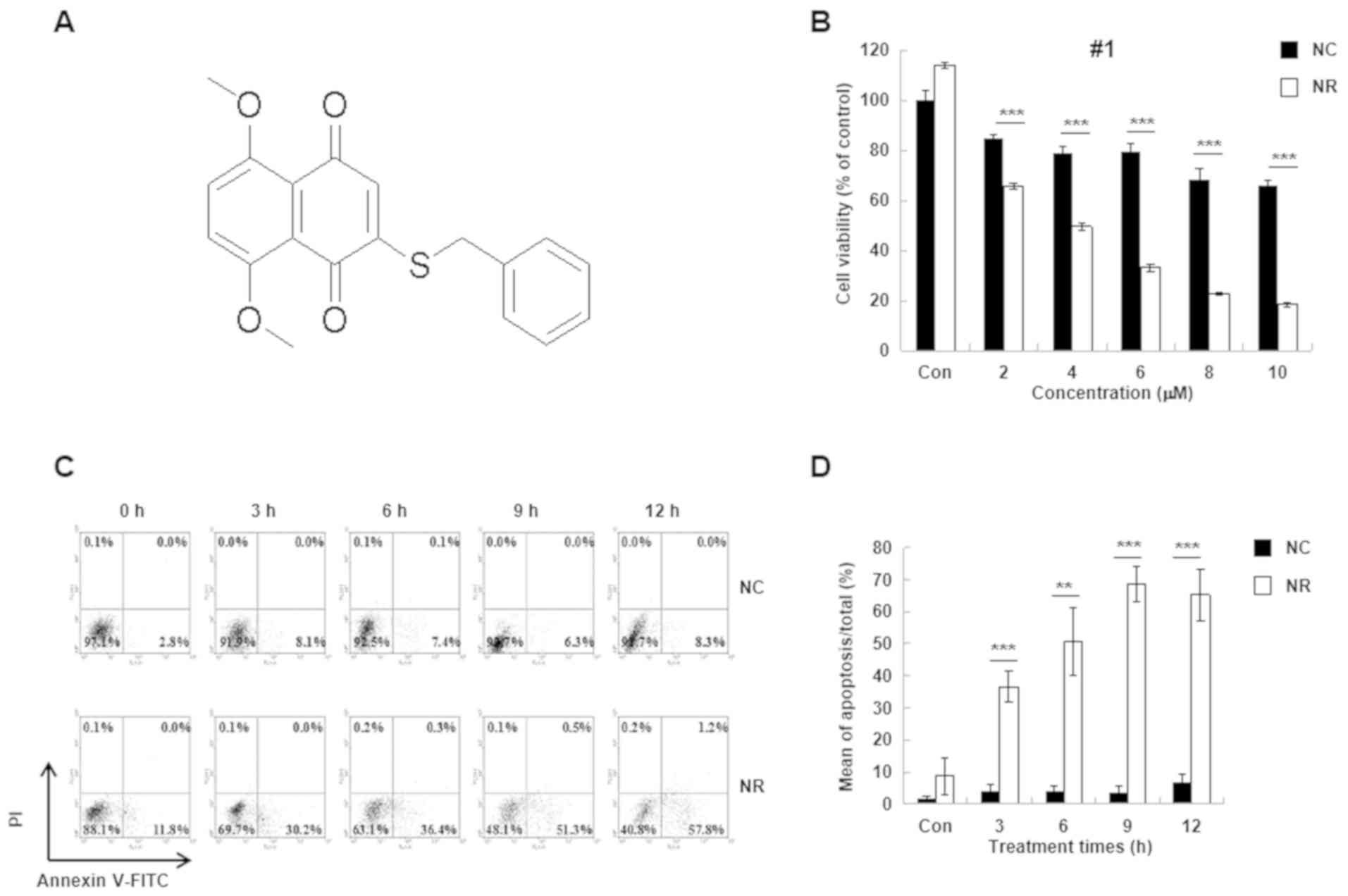 | Figure 2Effect of BZNQ on cellular viability
and apoptosis. (A) The structure of BZNQ as obtained from nuclear
magnetic resonance analysis. (B) The effect of BZNQ on NC (black
bars) and NR (white bars) cell viability was detected using MTT
assays. The untreated NC group was used as the control group. (C)
NC and NR cells were treated with BZNQ and cellular apoptosis was
examined with Annexin V-FITC/PI kit using flow cytometry. (D) The
level of apoptosis was presented as the mean ± standard deviation.
Experiments were performed in triplicate. **P<0.01,
***P<0.001. BZNQ,
2-benzylthio-5,8-dimethoxynaphthalene-1,4-dione; NC, NIH3T3 mouse
embryonic fibroblast cell line; NR,
H-RasG12V-transfected NIH3T3 mouse embryonic fibroblast
cell line; PI, propidium iodide; O, oxygen; S, sulphur; Con,
control. |
Following this, the effect of BZNQ on cellular
apoptosis in NC and NR cells was examined. The results indicated
that BZNQ induced NR cell apoptosis in a dose-dependent manner
(Fig. 2B). Furthermore, flow
cytometry analysis indicated that BZNQ treatment significantly
increased NR cell apoptosis compared with NC cells (Fig. 2C and D).
BZNQ induces cellular apoptosis
through the AKT, ERK and GSK signaling pathways
To elucidate the possible signaling pathways by
which BZNQ induces NR cell apoptosis, H-Ras protein expression in
cells treated with BZNQ for 24 h was evaluated. The results
demonstrated that BZNQ treatment significantly downregulated H-Ras
protein expression levels in NR cells (Fig. 3A). Due to these results, the known
Ras-mediated downstream signaling pathways, including AKT, ERK and
GSK, as well as β-catenin activation, were evaluated. The results
revealed that BZNQ treatment significantly downregulated AKT, ERK
and GSK3β protein phosphorylation in NR cells (white bar) compared
with NC cells (black bar; Fig.
3B).
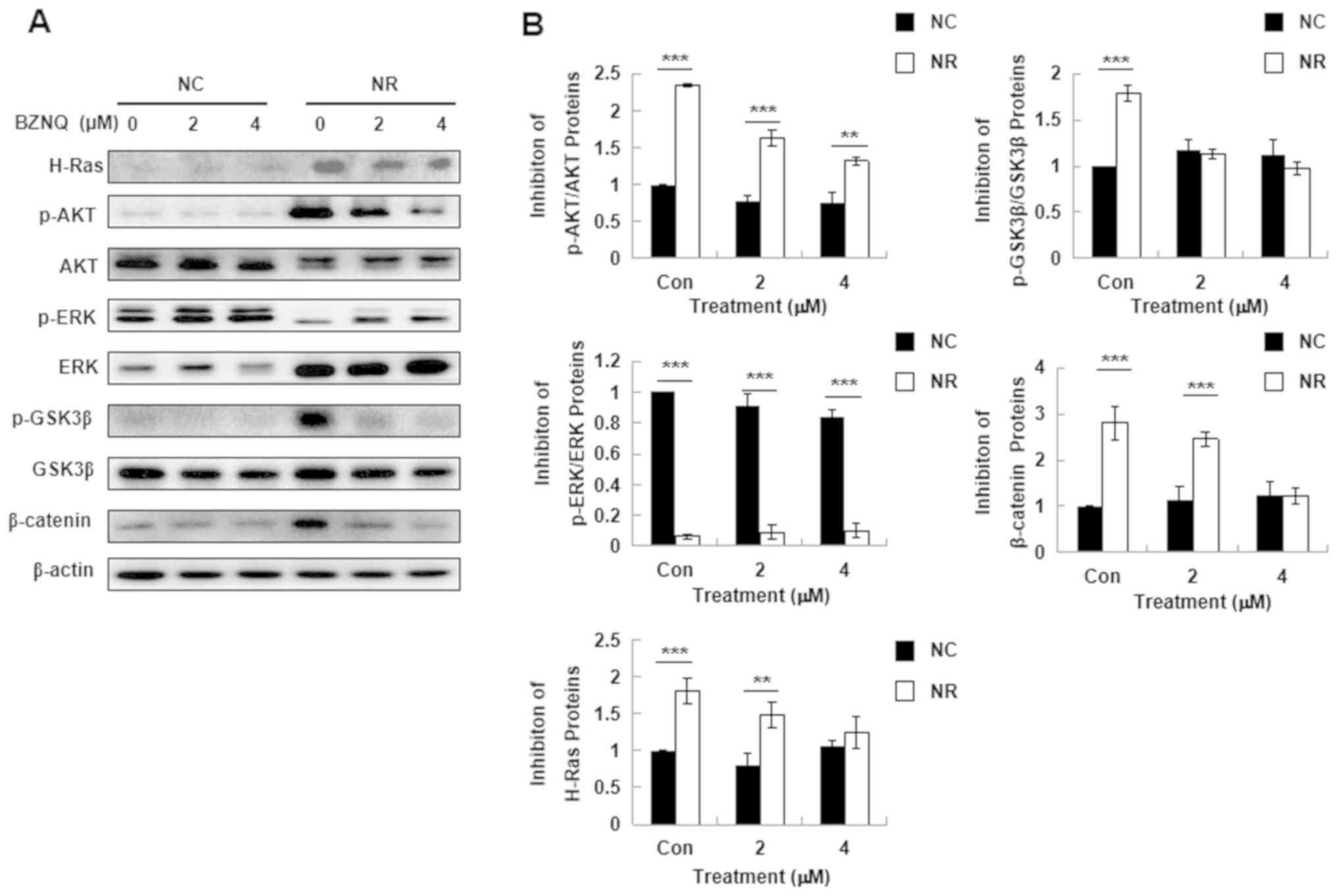 | Figure 3Effect of BZNQ on H-Ras protein
expression and Ras-mediated downstream signaling. (A) NC and NR
cells were treated with indicated concentrations of BZNQ for 24 h.
H-Ras, p-AKT, AKT, p-ERK, ERK, p-GSK3β, GSK3β, β-catenin and
β-actin protein expression levels were detected via western
blotting. (B) Fold increase in protein expression levels.
**P<0.01, ***P<0.001. Experiments were
performed in triplicate. BZNQ,
2-benzylthio-5,8-dimethoxynaphthalene-1,4-dione; NC, NIH3T3 mouse
embryonic fibroblast cell line; NR,
H-RasG12V-transfected NIH3T3 mouse embryonic fibroblast
cell line; AKT, protein kinase B; ERK, extracellular regulated
kinase; GSK3β, glycogen synthase kinase 3β. |
BZNQ increases cellular apoptosis and
ROS accumulation in HepG2 liver cancer cells
Since BZNQ increased cell apoptosis by targeting Ras
signaling proteins, the cytotoxic effects of BZNQ on HC and HR
liver cancer cells was evaluated. BZNQ exhibited highly marked
cytotoxicity in both HC and HR cells, as shown by MTT assays. After
BZNQ treatment, fluorescence intensity was increased when compared
with untreated cells, as observed by fluorescent microscopy
(Fig. 4C) and flow cytometry
analysis (Fig. 4D) by detecting DHE
fluorescence intensity. The results demonstrated that BZNQ
treatment elevated cellular ROS accumulations in HC and HR
cells.
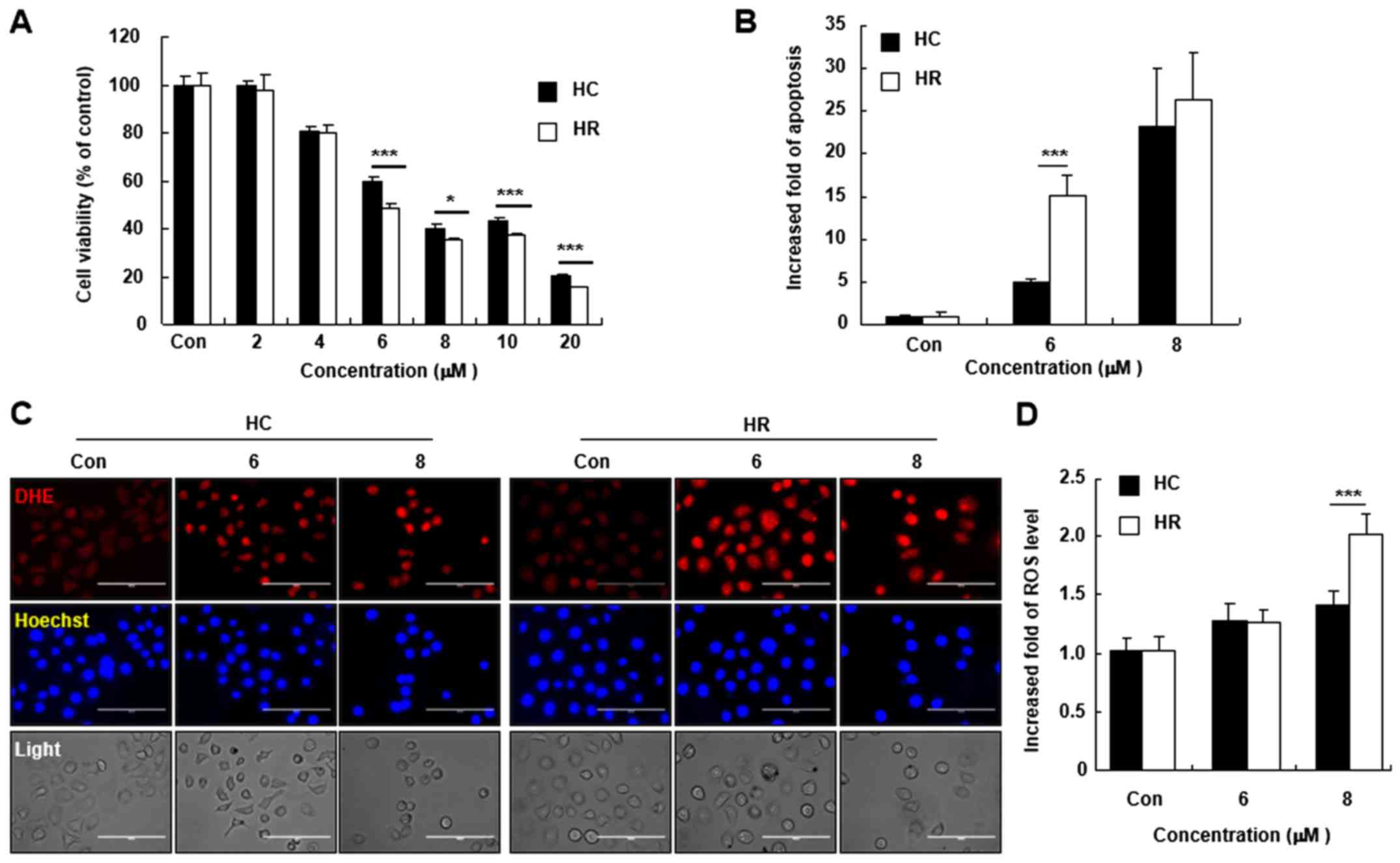 | Figure 4Effect of BZNQ on cancer cell
viability, apoptosis and ROS levels. (A) HC and HR cells were
treated with the indicated concentrations of BZNQ and cell
viability was examined using MTT assays. (B) Cellular apoptosis was
assayed using an Annexin V/propidium iodide staining kit and
measured using flow cytometry. The increased fold of apoptosis was
presented as mean ± SD. (C) Cellular ROS levels were detected using
DHE staining and observed with fluorescent microcopy. Scale bar,
100 µM. (D) Fold increase of ROS levels was presented as mean ± SD.
All experiments were performed in triplicate.
*P<0.05, ***P<0.001. BZNQ,
2-Benzylthio-5,8-dimethoxynaphthalene-1,4-dione; ROS, reactive
oxygen species; HC, HepG2 human liver cancer cell line; HR,
H-RasG12V-transfected HepG2 human liver cancer cell
line; DHE, dihydroethidium; SD, standard deviation; Con,
control. |
Scavenging ROS levels decreases
BZNQ-induced cellular apoptosis by upregulating the protein
expression of Bad
To understand the role of ROS on BZNQ-induced
cellular apoptosis, HC and HR cells were pretreated with NAC, a ROS
scavenger, for 30 min, followed with BZNQ treatment. NAC
pre-treatment decreased cellular ROS levels and apoptosis in HC and
HR cells (Fig. 5A-C). Furthermore,
apoptosis-related protein expression levels, including Bcl-2
(anti-apoptotic) and Bad (pro-apoptotic), were evaluated following
the BZNQ treatment. The results indicated that BZNQ treatment
upregulated Bad protein expression and slightly downregulated the
Bcl-2 expression in HC and HR cells (Fig. 5D).
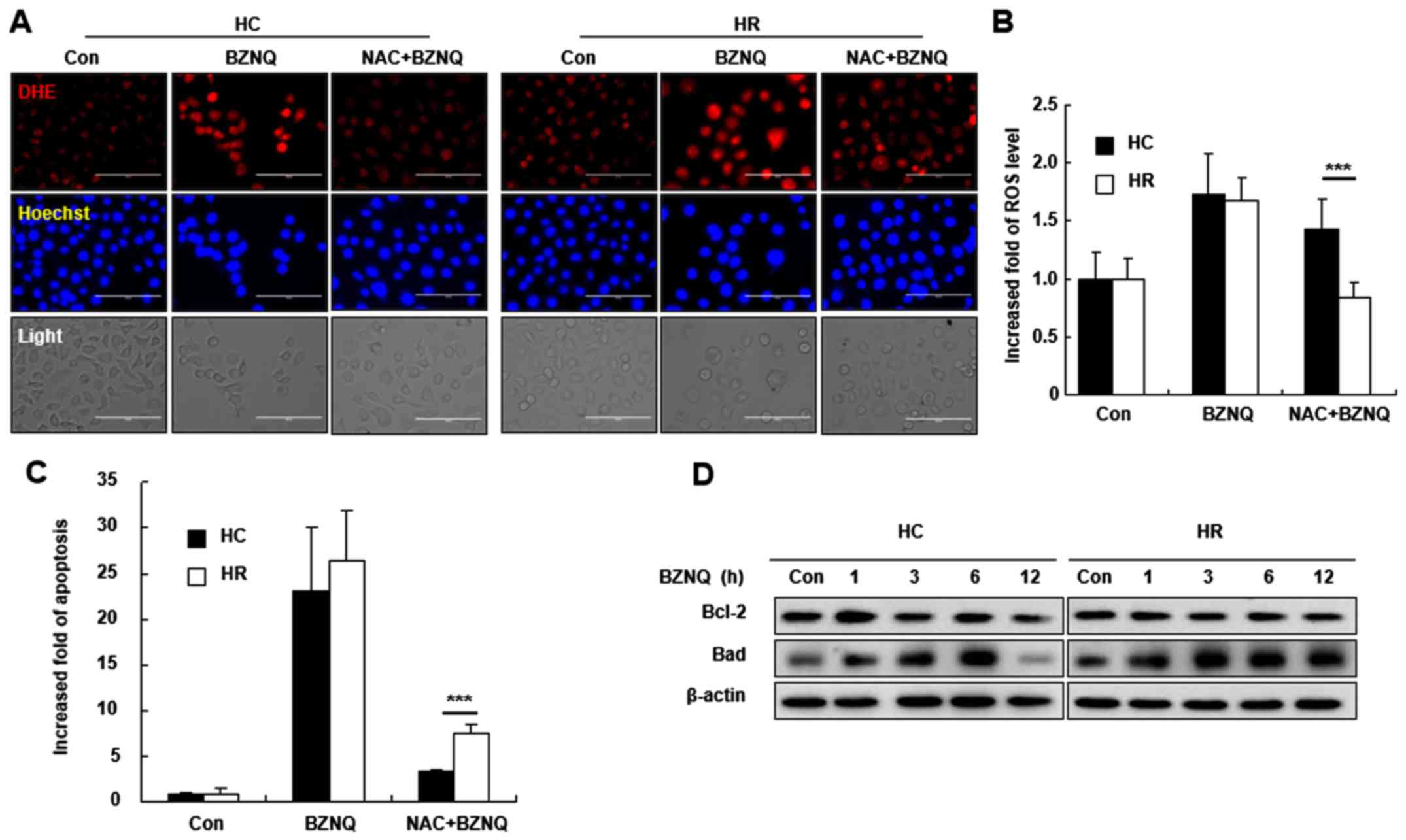 | Figure 5ROS scavenging inhibited BZNQ-induced
cellular apoptosis. (A) HC and HR cells were pre-treated with 5 mM
NAC, an ROS scavenger, for 30 min, followed by BZNQ treatment. ROS
levels were detected by DHE staining and observed with fluorescent
microcopy. Scale bar, 100 µM. (B) Cellular ROS levels were assayed
with DHE staining following NAC and BZNQ treatment and measured
using flow cytometry. Fold increase in ROS levels was presented as
mean ± SD. (C) Cellular apoptosis was assayed with Annexin
V/propidium iodide (staining kit following NAC and BZNQ treatment
and measured using flow cytometry assay. Fold increase in apoptosis
was presented as mean ± SD. (D) The protein expression levels of
Bcl-2 and Bad were examined following BZNQ treatment in HC and HR
cells by western blotting. Experiments were performed in
triplicate. ***P<0.001. ROS, reactive oxygen species;
BZNQ, 2-benzylthio-5,8-dimethoxynaphthalene-1,4-dione; HC, HepG2
human liver cancer cell line; HR, H-RasG12V-transfected
HepG2 human liver cancer cell line; NAC, N-acetyl-L-cysteine; DHE,
dihydroethidium; SD, standard deviation; Bcl-2, B-cell lymphoma 2;
Bad, Bcl-2 associated agonist of cell death; Con, control. |
Discussion
Shikonin is a small-molecule compound extracted from
the natural plant Arnebia euchroma and induces tumor cell
apoptosis (23-25),
inhibits DNA topoisomerase (26)
and protein tyrosine kinase (26),
represses human telomerase activity (27), suppresses tumor angiogenesis
(28), alters tumor cell signal
transduction and activates mitogen activated protein kinase (MAPK)
through chemical protection and induces antimitotic effects
(29). Furthermore, shikonin
influences the metabolism, proliferation, differentiation, signal
transmission, gene expression, cell growth, invasion and metastasis
of tumor cells (30-32).
However, due to the strong cytotoxicity and side effects of
shikonin, including nausea, vomiting and loss of appetite, it has
not been widely used in clinical practice (33). With further research into the
mechanisms of action of shikonin, it has been reported that the
toxic side effects of its derivatives are greatly reduced following
the addition of the methylated 5' and 8' hydroxyl groups to methoxy
groups (34). Moreover,
naphthoquinone derivatives are more convenient for the latter
modification and therefore, has more structural advantages in the
synthesis of new compounds. Previous results have demonstrated that
naphthoquinone derivatives specifically inhibit the p53-Snail
binding in K-RasG12D mutant A549 cells to induce cell
apoptosis (35), demonstrating that
naphthoquinone derivatives have the potential to target the Ras
signaling pathway.
1,4-Naphthoquinone is the main component of
Arnebia purpurea and widely exists in nature as a small
molecular compound with quinone moiety (36). Generally, when two kinds of
substituents contain the same hydrocarbon group, the cytotoxicity
of mercaptan derivatives is stronger compared with amino
derivatives (37). While amino
derivatives have anti-inflammatory effects (38), mercaptan derivatives have better
anti-tumor (39,40) and anti-malaria activity (41,42).
Bielitza et al (43)
reported that benzyl carbon in naphthoquinone derivatives
participates in the oxidation-reduction cycle of NADPH and that
benzyl carbon can be oxidized to carbonyl by hydroxylation or
peroxidation. If the para position of the benzene ring contains a
tert-butyl group, it can reduce the toxicity of compounds (44). The substituent alkyl terminal
containing hydroxyl or carboxyl group exhibits strong cytotoxicity
(45). The current study
synthesized and reported several groups of these kinds of compounds
which exhibited significant anticancer property. In order to
further investigate this series of DMNQ compounds, group compounds
that contain a sulfur atom linked between naphthnoquinone core and
linear substitutes were designed. The sulfur atom can be used as an
oxidation-reduction center to eliminate ROS in cells (46). Several substitutes, including
benzyl, butyl and cyclo propyl, which are normally selected to
optimize the property of designed compounds, were used in the
current study. The results demonstrated that benzyl substituted
compound BZNQ demonstrated excellent effects, with increased
cytotoxity in H-RasG12V mutated cells and a lower
concentration.
In the current study, 12 DMNQ derivatives were
designed, including BZNQ. Among the 12 kinds of derivatives, BZNQ
had the highest cytotoxic effect in NR cells compared with NC cells
compared with all other compounds from the IC50 value.
Furthermore, compounds #2-4 and #8-10 demonstrated significant
activities in NC and NR cells; however, there were no significant
differences in cytotoxicity effects between NC and NR cells (data
not shown), indicating that the different modulation of the
substitution groups at the positions exhibited various inhibiting
activities in cells. The possible molecular mechanisms that rely on
structure-activity relationships should be studied further.
The intracellular signaling pathways that regulate
apoptosis are diverse and complex. Generally, when Ras signaling
has been activated, the downstream MAPK signaling pathway must also
be activated (47). However, it has
been reported that H-RasG12V preferentially activates
the PI3K/AKT signaling pathway to promote cell survival following
starvation and activated AKT inhibits s621 phosphorylation of RAF
and downregulates ERK phosphorylation (48-50).
Furthermore, NF-κB was revealed to be an important downstream
signal molecule for AKT to regulate cell anti-apoptotic signaling
and inhibiting NF-κB blocked the anti-apoptotic signals regulated
by PDG/PI3K/AKT (51,52). Furthermore, BZNQ inhibited AKT and
GSK3β S9 phosphorylation, which resulted in promotion of β-catenin
in NIH3T3 cells These changes in the activity of signaling pathways
promoted apoptosis-related protein expression levels (Bad
upregulation and Bcl-2 downregulation), which led to cellular
apoptosis. Additionally, Ras, a member of MAPK upstream signal, can
cause the activation of downstream signals (53). It has also been reported that AKT
phosphorylates Raf, which activates ERK phosphorylation (54) and that there have alternative active
phenomenon between ERK and AKT, such as ERK upregulation and AKT
downregulation (55). The current
study did not result in direct evidence to clarify the differences
in the activation of H-Ras and p-ERK expression levels. This should
be further investigated in future studies.
Since BZNQ exhibited strong cytotoxic effects by
targeting Ras signaling, the current study hypothesized that it may
also have a toxic effect on cancer cells, particularly in Ras
mutated cancer cells. Therefore, the effect of BZNQ in HC and HR
cells was examined. The observed results demonstrated that there
was a high toxic effect on BZNQ on HC and HR cells, there were
significant differences in cellular apoptosis and ROS levels
between HC and HR cells. This is closely associated with the
5,8-dimethoxynaphthalene-1,4-dione core and benzyl group of BZNQ as
these subunits may serve an important role in the increase of
intracellular ROS levels and cytotoxicity (56). The current study speculates that
BZNQ promoted the apoptosis of Ras mutant cell lines by inhibiting
the expression of Ras proteins and increasing the level of ROS in
H-RasG12V transfected cells. The results indicated that
the Ras signaling pathway may serve different roles in healthy
cells and cancer cells. The possible mechanisms should be studied
further.
In conclusion, based on the analysis of the
structure-activity relationship of DMNQ derivatives, the current
study designed, synthesize and screened compounds that specifically
regulated the Ras mutation signal. The results confirmed the
effects of the compounds on the apoptosis signaling pathway
AKT/GSK3β/β-catenin, which is downstream of the Ras signaling
pathway. Although there was no direct evidence of how BZNQ induced
cell death in Ras-mutated cells, the results demonstrated that the
apoptotic levels of NC, NR, HC and HR cells were significantly
different following BZNQ treatment, resulting in significant
changes in the downstream signal of Ras. This indicated that
H-RasG12V transfected cells were more sensitive to BZNQ.
The specific mechanism needs further study. These results indicated
that the novel derivative synthesized based on DMNQ, BZNQ, may be a
therapeutic drug for treating Ras-mediated tumorigenesis through
the downregulation of H-Ras protein expression and Ras-mediated
signaling pathways.
Acknowledgements
Not applicable.
Funding
The current study was supported by the Natural
Science Foundation of Heilongjiang Province of China (grant no.
QC2016012), the University Nursing Program for Young Scholars with
Creative Talents in Heilongjiang Province (grant no. CXRC2017016)
the Heilongjiang Undergraduate Training Programs for Innovation and
Entrepreneurship (grant no. 201910223027), and the Basic Science
Research Program through the National Research Foundation of Korea
(NRF) funded by the Ministry of Education (2020R1I1A2052417) and
the grants from the Korean Research Institute of Bioscience and
Biotechnology Research Initiative Program (KRIBB; KGM5162021,
RBM0112011).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
GNS, JL and YHJ analyzed the data and wrote the
manuscript. GNS, TK and YHH designed experiments. HNS, YYH, MHJ,
RL, WLL and YQZ performed the experiments. JBY, NNY, WDW, LYY and
JSK developed novel software and performed simulations. TK and YHH
conceived and supervised the study. All authors read and approved
the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Prior IA, Hood FE and Hartley JL: The
frequency of ras mutations in cancer. Cancer Res.
80(2969)2020.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Yaeger R, Cowell E, Chou JF, Gewirtz AN,
Borsu L, Vakiani E, Solit DB, Rosen N, Capanu M, Ladanyi M and
Kemeny N: RAS mutations affect pattern of metastatic spread and
increase propensity for brain metastasis in colorectal cancer.
Cancer. 121:1195–1203. 2015.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Maertens O and Cichowski K: An expanding
role for RAS GTPase activating proteins (RAS GAPs) in cancer. Adv
Biol Regul. 55:1–14. 2014.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Grewal T, Koese M, Tebar F and Enrich C:
Differential regulation of RasGAPs in cancer. Genes Cancer.
2:288–297. 2011.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Maurya HK, Gautam SK, Pratap R, Tandon VK,
Kumar A, Kumar B, Saxena S, Tripathi D, Rajwanshi M, Das M and Ram
VJ: Regioselective synthesis of polycyclic aza-oxa and aza-oxa-thia
heteroarenes as Colo-205 and HepG2 carcinoma cells growth
inhibitors. Eur J Med Chem. 81:367–377. 2014.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Inamdar GS, Madhunapantula SV and
Robertson GP: Targeting the MAPK pathway in melanoma: Why some
approaches succeed and other fail. Biochem Pharmacol. 80:624–637.
2010.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Ledford H: Cancer: The Ras renaissance.
Nature. 520:278–280. 2015.PubMed/NCBI View
Article : Google Scholar
|
|
8
|
Zhang Y, Luo YH, Piao XJ, Shen GN, Wang
JR, Feng YC, Li JQ, Xu WT, Zhang Y, Zhang T, et al: The design of
1,4-naphthoquinone derivatives and mechanisms underlying apoptosis
induction through ROS-dependent MAPK/Akt/STAT3 pathways in human
lung cancer cells. Bioorg Med Chem. 27:1577–1587. 2019.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Lin HY, Han HW, Wang YS, He DL, Sun WX,
Feng L, Wen ZL, Yang MK, Lu GH, Wang XM, et al: Shikonin and
4-hydroxytamoxifen synergistically inhibit the proliferation of
breast cancer cells through activating apoptosis signaling pathway
in vitro and in vivo. Chin Med. 15(23)2020.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Seetha A, Devaraj H and Sudhandiran G:
Indomethacin and juglone inhibit inflammatory molecules to induce
apoptosis in colon cancer cells. J Biochem Mol Toxicol.
34(e22433)2020.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Vukic MD, Vukovic NL, Obradovic A, Matic
M, Djukic M and Avdovic E: Redox status, DNA and HSA binding study
of naturally occurring naphthoquinone derivatives. EXCLI J.
19:48–70. 2020.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Guo C, He J, Song X, Tan L, Wang M, Jiang
P, Li Y, Cao Z and Peng C: Pharmacological properties and
derivatives of shikonin-A review in recent years. Pharmacol Res.
149(104463)2019.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Lim ES, Rhee YH, Park MK, Shim BS, Ahn KS,
Kang H, Yoo HS and Kim SH: DMNQ S-64 induces apoptosis via caspase
activation and cyclooxygenase-2 inhibition in human nonsmall lung
cancer cells. Ann N Y Acad Sci. 1095:7–18. 2007.PubMed/NCBI View Article : Google Scholar
|
|
14
|
PLOS ONE Editors. Retraction:
Ramentaceone, a naphthoquinone derived from drosera sp., induces
apoptosis by suppressing PI3K/Akt signaling in breast cancer cells.
PLoS One. 14(e0226703)2019.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Zhang Q, Dong J, Cui J, Huang G, Meng Q
and Li S: Cytotoxicity of synthesized 1,4-naphthoquinone oxime
derivatives on selected human cancer cell lines. Chem Pharm Bull
(Tokyo). 66:612–619. 2018.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Shahsavari Z, Karami-Tehrani F and Salami
S: Shikonin induced necroptosis via reactive oxygen species in the
T-47D breast cancer cell line. Asian Pac J Cancer Prev.
16:7261–7266. 2015.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Qiao C, Bi S, Sun Y, Song D, Zhang H and
Zhou W: Study of interactions of anthraquinones with DNA using
ethidium bromide as a fluorescence probe. Spectrochim Acta A Mol
Biomol Spectrosc. 70:136–143. 2008.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Kobayashi K, Nishiumi S, Nishida M, Hirai
M, Azuma T, Yoshida H, Mizushina Y and Yoshida M: Effects of
quinone derivatives, such as 1,4-naphthoquinone, on DNA polymerase
inhibition and anti-inflammatory action. Med Chem. 7:37–44.
2011.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Liu C, Shen GN, Luo YH, Piao XJ, Jiang XY,
Meng LQ, Wang Y, Zhang Y, Wang JR, Wang H, et al: Novel
1,4-naphthoquinone derivatives induce apoptosis via ROS-mediated
p38/MAPK, Akt and STAT3 signaling in human hepatoma Hep3B cells.
Int J Biochem Cell Biol. 96:9–19. 2018.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Moore AR, Rosenberg SC, McCormick F and
Malek S: RAS-targeted therapies: Is the undruggable drugged? Nat
Rev Drug Discov. 19:533–552. 2020.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Aly AA, El-Sheref EM, Bakheet MEM, Mourad
MAE, Bräse S, Ibrahim MAA, Nieger M, Garvalov BK, Dalby KN and
Kaoud TS: Design, synthesis and biological evaluation of fused
naphthofuro[3,2-c] quinoline-6,7,12-triones and
pyrano[3,2-c]quinoline-6,7,8,13-tetraones derivatives as ERK
inhibitors with efficacy in BRAF-mutant melanoma. Bioorg Chem.
82:290–305. 2019.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Kabakci Z, Käppeli S, Cantù C, Jensen LD,
König C, Toggweiler J, Gentili C, Ribaudo G, Zagotto G, Basler K,
et al: Pharmacophore-guided discovery of CDC25 inhibitors causing
cell cycle arrest and tumor regression. Sci Rep.
9(1335)2019.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Jeung YJ, Kim HG, Ahn J, Lee HJ, Lee SB,
Won M, Jung CR, Im JY, Kim BK, Park SK, et al: Shikonin induces
apoptosis of lung cancer cells via activation of FOXO3a/EGR1/SIRT1
signaling antagonized by p300. Biochim Biophys Acta.
1863:2584–2593. 2016.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Gara RK, Srivastava VK, Duggal S, Bagga
JK, Bhatt M, Sanyal S and Mishra DP: Shikonin selectively induces
apoptosis in human prostate cancer cells through the endoplasmic
reticulum stress and mitochondrial apoptotic pathway. J Biomed Sci.
22(26)2015.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Chandimali N, Sun HN, Kong LZ, Zhen X, Liu
R, Kwon T and Lee DS: Shikonin-induced apoptosis of colon cancer
cells is reduced by peroxiredoxin V expression. Anticancer Res.
39:6115–6123. 2019.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Ahn BZ, Baik KU, Kweon GR, Lim K and Hwang
BD: Acylshikonin analogues: Synthesis and inhibition of DNA
topoisomerase-I. J Med Chem. 38:1044–1047. 1995.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Lu Q, Liu W, Ding J, Cai J and Duan W:
Shikonin derivatives: Synthesis and inhibition of human telomerase.
Bioorg Med Chem Lett. 12:1375–1378. 2002.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Li W, Zhang C, Ren A, Li T, Jin R, Li G,
Gu X, Shi R and Zhao Y: Shikonin suppresses skin carcinogenesis via
inhibiting cell proliferation. PLoS One.
10(e0126459)2015.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Wiench B, Eichhorn T, Paulsen M and
Efferth T: Shikonin directly targets mitochondria and causes
mitochondrial dysfunction in cancer cells. Evid Based Complement
Alternat Med. 2012(726025)2012.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Wang F, Yao X, Zhang Y and Tang J:
Synthesis, biological function and evaluation of Shikonin in cancer
therapy. Fitoterapia. 134:329–339. 2019.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Andújar I, Ríos JL, Giner RM and Recio MC:
Pharmacological properties of shikonin-a review of literature since
2002. Planta Med. 79:1685–1697. 2013.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Liang W, Cui J, Zhang K, Xi H, Cai A, Li
J, Gao Y, Hu C, Liu Y, Lu Y, et al: Shikonin induces ROS-based
mitochondria-mediated apoptosis in colon cancer. Oncotarget.
8:109094–109106. 2017.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Boulos JC, Rahama M, Hegazy MF and Efferth
T: Shikonin derivatives for cancer prevention and therapy. Cancer
Lett. 459:248–267. 2019.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Ali A, Assimopoulou AN, Papageorgiou VP
and Kolodziej H: Structure/antileishmanial activity relationship
study of naphthoquinones and dependency of the mode of action on
the substitution patterns. Planta Med. 77:2003–2012.
2011.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Lee SH, Shen GN, Jung YS, Lee SJ, Chung
JY, Kim HS, Xu Y, Choi Y, Lee JW, Ha NC, et al: Antitumor effect of
novel small chemical inhibitors of Snail-p53 binding in
K-Ras-mutated cancer cells. Oncogene. 29:4576–4587. 2010.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Chen X, Yang L, Oppenheim JJ and Howard
MZ: Cellular pharmacology studies of shikonin derivatives.
Phytother Res. 16:199–209. 2020.
|
|
37
|
Elferink JG and Deierkauf M: Inhibition of
polymorphonuclear leukocyte functions by fluorinated nitrobenzenes.
Chem Biol Interact. 52:163–72. 1984.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Sun HN, Shen GN, Jin YZ, Jin Y, Han YH,
Feng L, Liu L, Jin MH, Luo YH, Kwon TH, et al:
2-Cyclohexylamino-5,8-dimethoxy-1,4-naphthoquinone inhibits
LPS-induced BV2 microglial activation through MAPK/NF-kB signaling
pathways. Heliyon. 2(e00132)2016.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Gaascht F, Teiten MH, Cerella C, Dicato M,
Bagrel D and Diederich M: Plumbagin modulates leukemia cell redox
status. Molecules. 19:10011–10032. 2014.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Zhang J, Peng S, Li X, Liu R, Han X and
Fang J: Targeting thioredoxin reductase by plumbagin contributes to
inducing apoptosis of HL-60 cells. Arch Biochem Biophys. 619:16–26.
2017.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Sharma A, Santos IO, Gaur P, Ferreira VF,
Garcia CR and da Rocha DR: Addition of thiols to o-quinone methide:
New 2-hydroxy-3-phenylsulfanylmethyl[1,4]naphthoquinones and their
activity against the human malaria parasite Plasmodium falciparum
(3D7). Eur J Med Chem. 59:48–53. 2013.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Pandey SK, Naware NB, Trivedi P and Saxena
AK: Molecular modeling and 3D-QSAR studies in 2-aziridinyl-and
2,3-bis(aziridinyl)-1,4-naphthoquinonyl sulfonate and acylate
derivatives as potential antimalarial agents. SAR QSAR Environ Res.
12:547–564. 2001.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Bielitza M, Belorgey D, Ehrhardt K, Johann
L, Lanfranchi DA, Gallo V, Schwarzer E, Mohring F, Jortzik E,
Williams DL, et al: Antimalarial NADPH-consuming redox-cyclers as
superior glucose-6-phosphate dehydrogenase deficiency copycats.
Antioxid Redox Signal. 22:1337–1351. 2015.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Edenharder R and Grünhage D: Free radical
scavenging abilities of flavonoids as mechanism of protection
against mutagenicity induced by tert-butyl hydroperoxide or cumene
hydroperoxide in Salmonella typhimurium TA102. Mutat Res. 540:1–18.
2003.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Fukushi S, Yoshino H, Yoshizawa A and
Kashiwakura I: p53-independent structure-activity relationships of
3-ring mesogenic compounds' activity as cytotoxic effects against
human non-small cell lung cancer lines. BMC Cancer.
16(521)2006.
|
|
46
|
Gao M, Wang R, Yu F and Chen L: Evaluation
of sulfane sulfur bioeffects via a mitochondria-targeting
selenium-containing near-infrared fluorescent probe. Biomaterials.
160:1–14. 2018.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Santarpia L, Lippman SM and El-Naggar AK:
Targeting the MAPK-RAS-RAF signaling pathway in cancer therapy.
Expert Opin Ther Targets. 16:103–119. 2012.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Zhu J, Blenis J and Yuan J: Activation of
PI3K/Akt and MAPK pathways regulates Myc-mediated transcription by
phosphorylating and promoting the degradation of Mad1. Proc Natl
Acad Sci USA. 105:6584–6589. 2008.PubMed/NCBI View Article : Google Scholar
|
|
49
|
Steelman LS, Chappell WH, Abrams SL, Kempf
RC, Long J, Laidler P, Mijatovic S, Maksimovic-Ivanic D, Stivala F,
Mazzarino MC, et al: Roles of the Raf/MEK/ERK and
PI3K/PTEN/Akt/mTOR pathways in controlling growth and sensitivity
to therapy-implications for cancer and aging. Aging (Albany NY).
3:192–222. 2011.PubMed/NCBI View Article : Google Scholar
|
|
50
|
Asati V, Mahapatra DK and Bharti SK:
PI3K/Akt/mTOR and Ras/Raf/MEK/ERK signaling pathways inhibitors as
anticancer agents: Structural and pharmacological perspectives. Eur
J Med Chem. 109:314–341. 2016.PubMed/NCBI View Article : Google Scholar
|
|
51
|
Romashkova JA and Makarov SS: NF-kappaB is
a target of AKT in anti-apoptotic PDGF signalling. Nature.
401:86–90. 1999.PubMed/NCBI View
Article : Google Scholar
|
|
52
|
Gingery A, Bradley EW, Pederson L, Ruan M,
Horwood NJ and Oursler MJ: TGF-beta coordinately activates
TAK1/MEK/AKT/NFkB and SMAD pathways to promote osteoclast survival.
Exp Cell Res. 314:2725–2738. 2008.PubMed/NCBI View Article : Google Scholar
|
|
53
|
Delire B and Stärkel P: The Ras/MAPK
pathway and hepatocarcinoma: Pathogenesis and therapeutic
implications. Eur J Clin Invest. 45:609–63. 2015.PubMed/NCBI View Article : Google Scholar
|
|
54
|
Rommel C, Clarke BA, Zimmermann S, Nuñez
L, Rossman R, Reid K, Moelling K, Yancopoulos GD and Glass DJ:
Differentiation stage-specific inhibition of the Raf-MEK-ERK
pathway by Akt. Science. 286:1738–1741. 1999.PubMed/NCBI View Article : Google Scholar
|
|
55
|
Balmanno K, Chell SD, Gillings AS, Hayat S
and Cook SJ: Intrinsic resistance to the MEK1/2 inhibitor AZD6244
(ARRY-142886) is associated with weak ERK1/2 signalling and/or
strong PI3K signalling in colorectal cancer cell lines. Int J
Cancer. 125:2332–2341. 2009.PubMed/NCBI View Article : Google Scholar
|
|
56
|
Gholampour M, Ranjbar S, Edraki N,
Mohabbati M, Firuzi O and Khoshneviszadeh M: Click
chemistry-assisted synthesis of novel
aminonaphthoquinone-1,2,3-triazole hybrids and investigation of
their cytotoxicity and cancer cell cycle alterations. Bioorg Chem.
88(102967)2019.PubMed/NCBI View Article : Google Scholar
|














