Introduction
Lower back pain (LBP) is one of the most common
complaints in adult patients, with total costs incurred by
complications associated with LBP amounting to ~$100 billion/year
in the US (1,2). Intervertebral disc degeneration (IDD)
is closely linked to LBP and IDD-associated conditions are among
the most common causes of disability among all workers aged 18-64
years in the US (3).
One of the characteristic features of IDD is the
loss of intervertebral disc (IVD) extracellular matrix (ECM). The
mechanisms of IDD are complex, involving numerous factors that may
interact with each other (4).
During the course of IDD, homeostasis of the ECM shifts toward a
degenerative state (5). ECM
degradation is caused by increases in the levels of destructive
enzymes and a reduction in ECM synthesis. Certain drugs, including
transforming growth factor-β1 (TGF-β1), which promote ECM synthesis
and regeneration, may restore homeostasis within the IVD and reduce
IDD (6-8).
Vascular ingrowth has been frequently observed in
degenerated IVDs, while healthy IVDs are avascular (9,10).
Intact aggrecan and notochordal cells are key inhibitors of neural
and vascular ingrowth in healthy IVDs. However, with the
degradation of ECM and the degeneration of nucleus pulposus (NP)
cells, these barriers may also become dysfunctional (11,12).
According to previous studies, degenerative NP cells secrete
vascular endothelial growth factor (VEGF), which binds to VEGF
receptor 2 (VEGFR-2) on the endothelial cell (EC) membrane surface
(13).
Melatonin is an endogenous molecule released from
the pineal gland that has a role in the regulation of the circadian
rhythm and anti-oxidative stress and has an anti-inflammatory
effect (14,15). Recent studies have reported that
melatonin may protect IVDs from degeneration by improving cell
survival and mitophagy induction (14,16).
In addition, melatonin has been reported to exert a potential
anti-angiogenic effect (17).
However, whether melatonin is able to exert chondroprotective and
anti-angiogenic effects in IVDs has yet to be established.
Current treatments for IDD are largely focused on
symptom management, rather than treatment of the underlying causes
of the disease (4,18). There is no satisfactory treatment
available that is able to completely regenerate the degenerated
disc and spinal fusion and disc arthroplasty may cause numerous
problems, including fusion failure, pain and loss of function
(19,20). In the present study, the potential
anabolic and anti-angiogenic effects mediated by melatonin were
examined in IVD cells. The study aimed to assess the biological
function of melatonin in IDD to determine its suitability as a
therapeutic agent.
Materials and methods
Cell culture
Human NP cells and degenerative human NP (DNP) cells
were isolated from surgical specimens at Ningbo No. 6 Hospital
(Ningbo, China). Normal NP cells were isolated from patients with
lumbar fracture or neurilemmoma that underwent spinal fusion
surgery and DNP cells were isolated from patients with IDD. Samples
were collected between January 2018 and June 2019. Written informed
consent was provided prior to sample collection. Institutional
review board approval was given by the Ethics Committee of the
Ningbo No. 6 Hospital (Ningbo, China; approval no. 2016031). A
total of 15 patients (age, 30-75 years; mean age, 51 years; female,
10; male, 5) were included in the present study. All samples were
used individually. NP and DNP cells were cultured in high-glucose
Dulbecco's modified Eagle's medium (DMEM; Gibco; Thermo Fisher
Scientific, Inc.) with 10% fetal bovine serum (Gibco; Thermo Fisher
Scientific, Inc.), 100 U/ml penicillin (HyClone; Cytiva) and 100
µg/ml streptomycin (HyClone; Cytiva). To achieve a better growth
environment for the cells, high-glucose DMEM was used in the
experiments. The concentration of glucose in the medium (4.5 g/l)
was far below the harmful concentration reported in the literature
(36 g/l) (21). Human umbilical
vein ECs (HUVEC) were purchased from the Cell Bank of Type Culture
Collection of the Chinese Academy of Science (Shanghai, China) and
cultured in DMEM/F12 media (Gibco; Thermo Fisher Scientific, Inc.)
with 10% fetal bovine serum, 100 U/ml penicillin and 100 µg/ml
streptomycin. All cells were cultured in an incubator at 37˚C in a
humidified atmosphere with 5% CO2 up to passages
2-3.
Cell proliferation assay
A commercial kit [Cell Counting Kit-8 (CCK-8);
Dojindo Molecular Technologies, Inc.] was used to assess cell
proliferation. NP cells were seeded in 96-well plates at a density
of 5x103 cells per well. The concentration of melatonin
was selected based on the result of preliminary experiments and
values reported in the literature (16). In the present study, cells were
incubated with melatonin (10 µM, 2.39 µg/ml) or TGF-β1 (10 ng/ml)
for 1-6 days, followed by treatment with 10 µl CCK-8 solution and
incubation at 37˚C for 2.5 h. The optical density (OD) of each well
was measured at 450 nm with a microplate reader (Thermo Fisher
Scientific, Inc.).
Reverse transcription-quantitative PCR
(RT-qPCR)
NP cells were treated with melatonin (10 µM) or
TGF-β1 (10 ng/ml) for 24 h. Following treatment, total RNA was
extracted from NP cells using TRIzol® reagent
(Invitrogen; Thermo Fisher Scientific, Inc.), following the
manufacturer's protocol. A total of 1 µg of total RNA was then
incubated with the Prime Script™ Master Mix (Takara Bio, Inc.) to
synthesize complementary DNA by RT. Gene expression was determined
by qPCR using the SYBR Premix Ex Taq kit (Takara Bio, Inc.). The
thermocycling conditions were as follows: Initialization at 95˚C
for 3 min, followed by 40 cycles of 95˚C for 30 sec, 55˚C for 20
sec and 72˚C for 20 sec, with a final amplification at 95˚C for 15
sec. Gene expression was determined using the
2-ΔΔCq method (22). The primers were designed and
selected using BLAST (https://blast.ncbi.nlm.nih.gov). Primer sequences are
listed in Table I. GAPDH was used
as an internal control.
 | Table IPrimer sequences. |
Table I
Primer sequences.
| Gene | Primer sequences
(5'-3') |
|---|
| SOX9 | Forward
AGCGAACGCACATCAAGAC |
| | Reverse
CTGTAGGCGATCTGTTGGGG |
| ACAN | Forward
ACTCTGGGTTTTCGTGACTCT |
| | Reverse
ACACTCAGCGAGTTGTCATGG |
| COL2A1 | Forward
TGGACGCCATGAAGGTTTTCT |
| | Reverse
TGGGAGCCAGATTGTCATCTC |
| GAPDH | Forward
TGTGGGCATCAATGGATTTGG |
| | Reverse
ACACCATGTATTCCGGGTCAAT |
Immunohistochemistry
A total of 1x104/ml NP or DNP cells were
seeded onto coverslips and treated with melatonin (10 µM) or TGF-β1
(10 ng/ml) for 3 or 7 days. Immunohistochemistry was then performed
to determine the expression of collagen-II. Cells were fixed using
4% paraformaldehyde for 30 min at room temperature, prior to
incubation with 0.2% Triton X-100 for 10 min at room temperature.
After blocking with 2% bovine serum albumin (Sigma Aldrich; Merck
KGaA) at room temperature for 1 h, cells were incubated with
anti-collagen-II (cat. no. ab24118; 1:500; Abcam) antibody at 4˚C
overnight. Cells were then treated with secondary antibody (cat.
no. M00172; 1:2,000; Boster Biological Technology) at room
temperature for 1 h, followed by color development with
diaminobenzidine tetrahydrochloride (Dako; Agilent Technologies,
Inc.). The images were observed under a light microscope (Olympus
Corp.). The results of type II collagen staining were quantified by
determining the integral OD (IOD) using Image-Pro Plus software
(version 6.0; Media Cybernetics, Inc.)
Tube formation assay
NP cells were cultured in complete medium for 48 h.
The medium was then collected and centrifuged at 200 x g at 4˚C for
5 min and the supernatant was collected as the conditioned medium.
HUVECs were seeded at a density of 1x104/well in 96-well
plates precoated with Matrigel®. Matrigel Matrix was
coated at 12 mg/ml at 4˚C for 20 mins and then treated with
melatonin (10 µM) or TGF-β1 (10 ng/ml) for 4 h at 37˚C. HUVECs were
then incubated with NP conditioned medium in the presence or
absence of 25 ng/ml VEGF for 6 h. The groups were as follows:
HUVECs, HUVECs + melatonin (M), HUVECs + VEGF (V), HUVECs + V + M
and HUVECs + V + TGF-β1 (T). The formation of tube-like structures
was observed under a light microscope (Olympus Corp.) The images
were observed under x40 magnification, with 5 fields in each sample
randomly selected.
EC migration
A total of 1x105 HUVECs were seeded on
Transwell polycarbonate membrane inserts (8.0-µm pores; Corning,
Inc.) and maintained in 100 µl high-glucose DMEM without serum. NP
or DNP cells were cultured in complete medium with melatonin (10
µM) or TGF-β1 (10 ng/ml) in the lower chamber. The groups were as
follows (upper chamber/lower chamber): HUVECs/NP, HUVECs/DNP,
HUVECs/DNP + M, HUVECs/NP + V, HUVECs/NP + M + V, HUVECs/NP + V +
T. All inserts were incubated for 24 h at 37˚C and 5%
CO2 in an incubator. The inserts were washed with PBS
and the cells on the top surface of the inserts were carefully
removed using a cotton swab. The inserts were fixed with 4%
paraformaldehyde for 15 min at room temperature, followed by
staining with 0.1% crystal violet for 30 min at room temperature.
Migrated cells were observed and counted under x40 magnification
with 5 fields randomly selected in each sample using a light
microscope (Olympus Corp.).
VEGF binding assay
HUVECs were seeded at a density of
1x104/well in 96-well plates for 24 h. Cells were
pretreated with melatonin ((10 µM) or TGF-β1 (10 ng/ml) for 4 h,
then 25 ng/ml VEGF labeled with Alexa Fluor 647® (Abace
Biology) was added to the culture medium and the wells were
incubated at room temperature for 4 h. The HUVECs were then washed
with PBS 3 times and counterstained with DAPI (1:1,000 dilution)
for 5 min at room temperature. VEGF bound to the remaining
receptors and was observed under a confocal microscope (Olympus
Corp.). The positive rate was calculated as the IOD/cell number
using Image-Pro Plus software (version 6.0; Media Cybernetics,
Inc.)
Statistical analysis
Values are expressed as the mean ± standard
deviation from three independent repeats. Statistical analyses were
performed with unpaired t-test or one-way analysis of variance
(ANOVA) followed by Duncan's post-hoc test using SPSS 24.0 (IBM
Corp.). For experiments with >3 groups, Tukey's post-hoc test
was used following ANOVA. P<0.05 was considered to indicate a
statistically significant difference.
Results
Melatonin promotes ECM-associated gene
expression and NP cell proliferation
NP cells were treated with 10 µM melatonin or 10
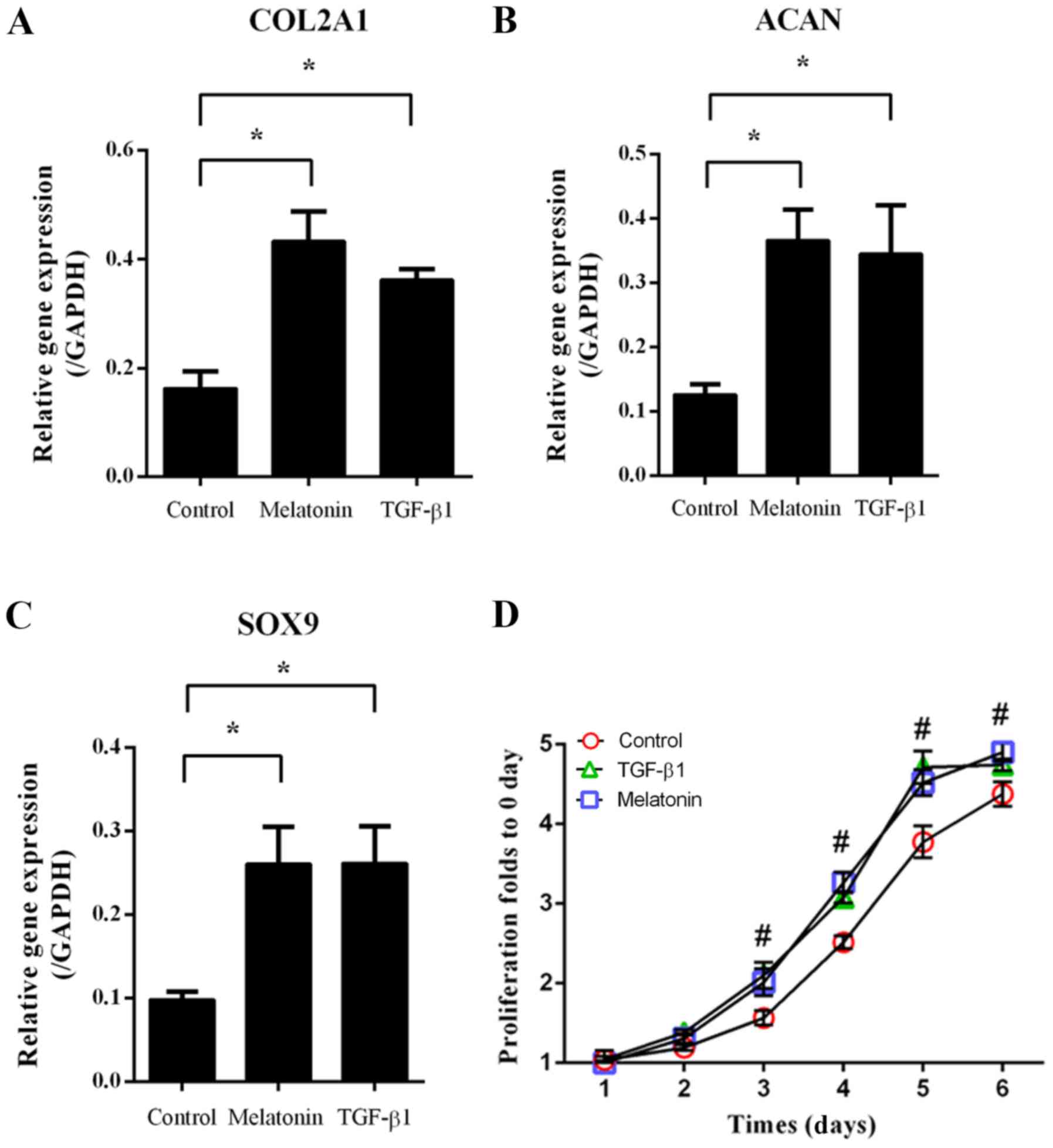
ng/ml TGF-β1, followed by RT-qPCR to assess the expression of
ECM-associated genes. As indicated in Fig. 1, melatonin and TGF-β1 significantly
increased type II collagen AI (COL2A1), aggrecan (ACAN) and
SRY-related HMG box-9 (SOX9) expression compared with that in the
group subjected to the control treatment (P<0.05; Fig. 1A-C). In addition, the cell
proliferation in the melatonin- and TGF-β1-treated groups was
increased from day 3 onwards when compared with that in the control
group (P<0.05; Fig. 1D). These
results indicated that melatonin and TGF-β1 promoted NP cell
proliferation and ECM-associated gene expression.
Melatonin increases collagen-II
expression in NP cells
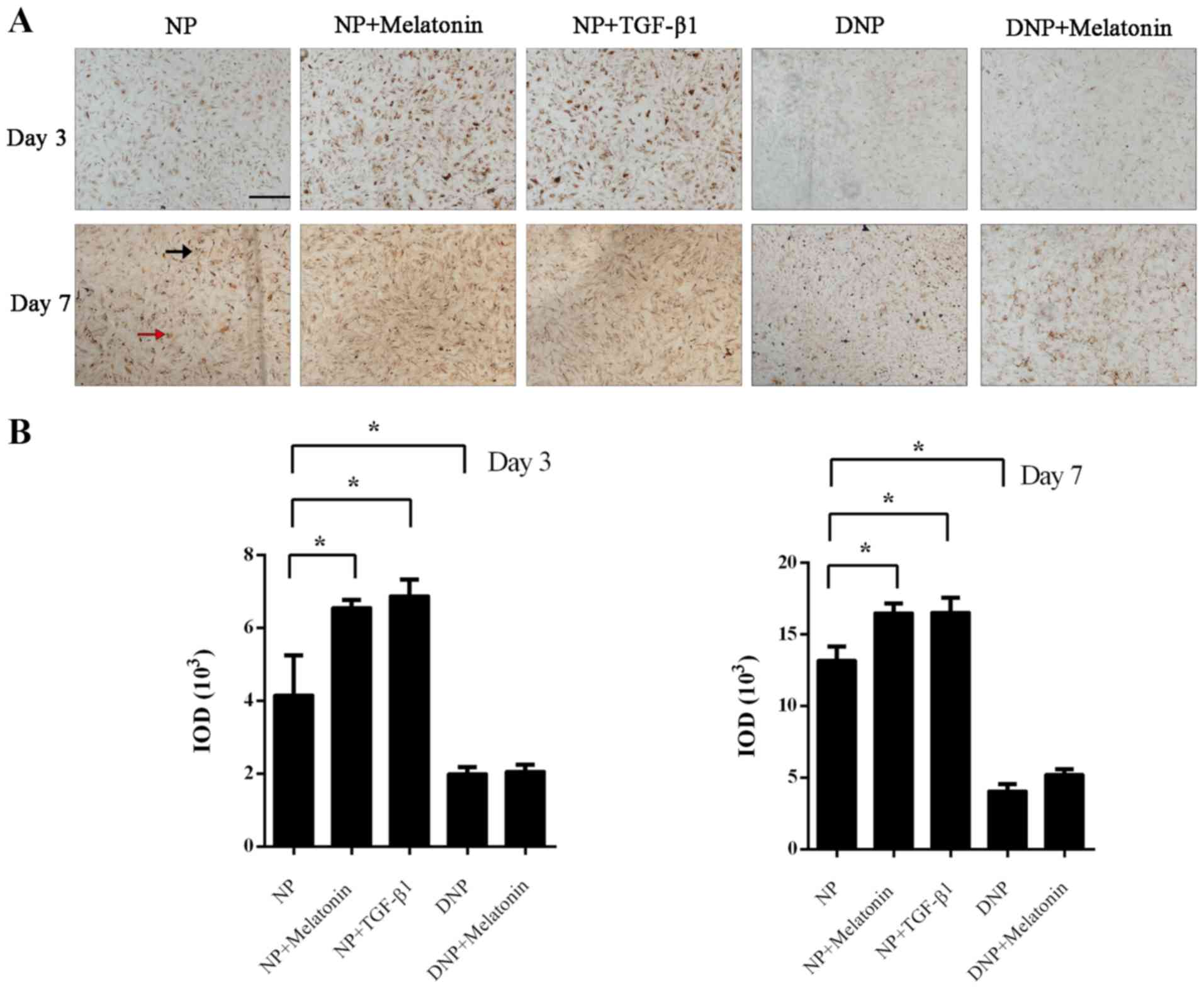
Immunohistochemistry was performed to assess
collagen-II expression in NP cells after incubation with melatonin
or TGF-β1. On day 3, positive staining was mainly present inside
the NP cells, while at day 7, staining for collagen-II was observed
both intracellularly (red arrow) and extracellularly (black arrow).
The immunohistochemistry staining for collagen-II was markedly
enhanced in the melatonin and TGF-β1 groups compared with that in
the control group (P<0.05; Fig.
2A). Quantification of the IOD also indicated that the
melatonin and TGF-β1 groups had increased collagen-II staining at
day 3 and day 7 (P<0.05; Fig.
2B). There was less collagen-II staining in DNP cells and the
rate of its increase was also markedly slower. Treatment of DNP
cells with melatonin did not increase the collagen-II content.
Although positive staining was increased on day 7 in the DNP +
melatonin group, this was not statistically significant (P>0.05;
Fig. 2B).
Melatonin suppresses tube formation
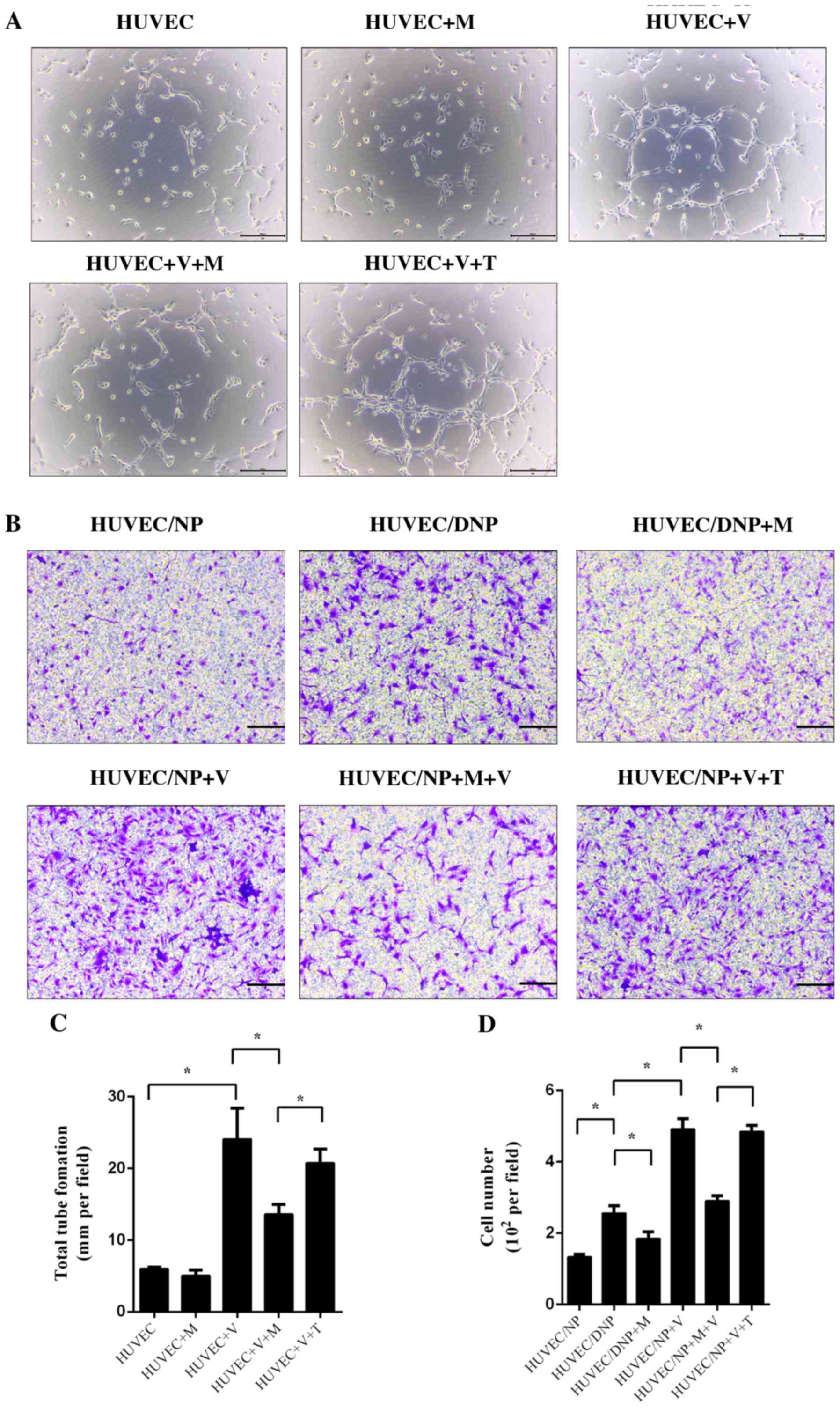
and migration of HUVECs
HUVECs were treated with conditioned medium with and
without VEGF for 6 h. Tube formation was observed under a light
microscope. After culture for 6 h, a small number of tube-like
structures was observed. Treatment with melatonin alone did not
influence tube formation of HUVECs (P>0.05; Fig. 3). Incubation with VEGF significantly
promoted tube formation of HUVECs; however, this effect was
inhibited by melatonin (P<0.05; Fig.
3A and C). TGF-β1 did not exert
any significant influence on the tube formation of HUVECs after
treatment with VEGF (P>0.05; Fig.
3A and C). An NP (DNP)/HUVEC
co-culture model was set up to assess the migration of HUVECs. When
compared with normal NP cells, DNP cells promoted increased HUVEC
migration, which suggested that degeneration may be a cause of
vascular ingrowth (P<0.05; Fig.
3B and D). The migration of
HUVECs was also promoted by VEGF in NP/HUVEC models (P<0.05;
Fig. 3B and C). Melatonin prevented the effect of DNP
and VEGF-induced migration of HUVECs (P<0.05; Fig. 3B and C), but TGF-β1 did not (P>0.05; Fig. 3B and C).
Melatonin inhibits the binding of VEGF
to its receptor in HUVECs
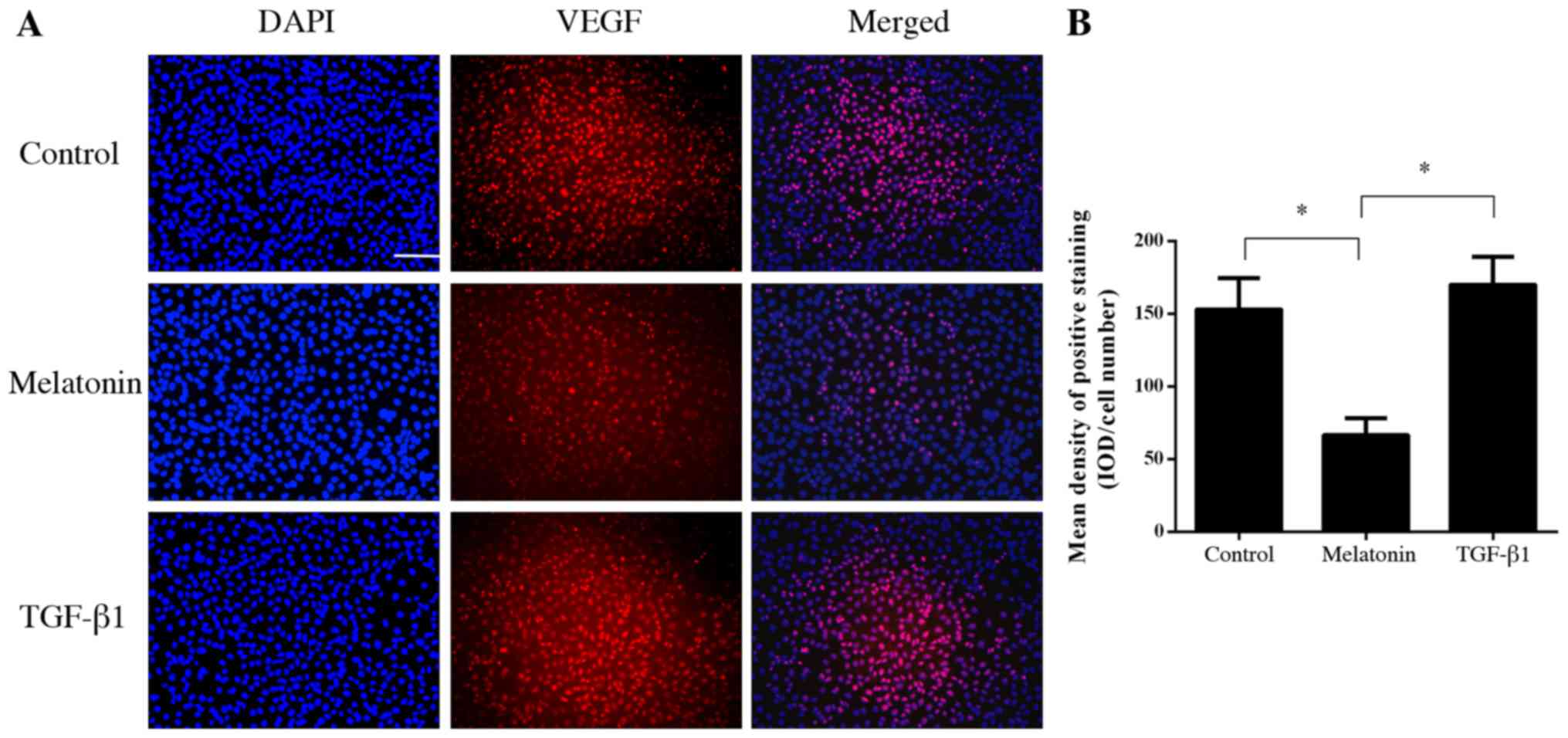
To further investigate the mechanism of the
anti-angiogenic effect of melatonin, fluorescence-labeled VEGF was
used to bind to VEGFR on HUVECs. The results indicated that
melatonin was able to inhibit the binding of VEGF to its receptor
on HUVECs (P<0.05; Fig. 4).
However, TGF-β1 did not influence the binding of VEGF to the VEGFR
(P>0.05; Fig. 4).
Discussion
Melatonin is not only prevalent inside the human
body but also in various plants and their products, including nuts,
tomatoes, olive oil and wine (17,23,24).
In addition to its function to synchronize the circadian rhythm
(25) and its utility in the
treatment of cardiovascular diseases and cancers (26), a beneficial application of melatonin
for its anti-angiogenic effect has been previously reported
(27,28). Melatonin has been suggested to
inhibit angiogenesis indirectly by downregulating VEGF expression
and secretion in various types of tumor cell (28,29).
VEGF is an important growth factor in ECs. Overexpression of VEGF
was identified in NP cells under inflammatory or degenerative
conditions and was considered to be one of the major causes of
vascular ingrowth in IVDs (30).
Healthy IVDs are aneural and avascular, while neovascularization is
considered to be an important pathological mechanism of symptomatic
IVD degeneration (4). Vascular
ingrowth has also been reported to be linked to nerve ingrowth and
back pain (10,13). Vascular cells are able to produce
nerve growth factor and brain-derived growth factor, the levels of
which were indicated to be associated with increasing degeneration
of the IVD (4,10). As the major angiogenic factor in
IVD, VEGF binds to receptors on the surface of ECs (mainly VEGFR-2)
to trigger proliferation and migration and promote the survival of
ECs, ultimately resulting in vascular ingrowth in IVDs (31,32).
Previous studies have also indicated that melatonin was able to
inhibit HUVEC proliferation via modulating P53, Bax/Bcl-2
expression and ERK1/2/PI3K/AKT/protein kinase C/NF-Κb (33,34).
Another study suggested that melatonin significantly inhibited
VEGFR-2 phosphorylation (30%) and HUVEC migration (87%) at a high
concentration of 1 mM (239 µg/ml) (17). Plants and their products contain
melatonin at concentrations of up to 230 µg/g or ml (17,23,24).
Calculation of the daily intake of melatonin and total plasma
volume of adults indicated that the concentration of circulating
melatonin would be 0.15-21 ng/ml, which is far below the
therapeutic dose (17). The results
of the present study demonstrated that melatonin exerted a
potential anti-angiogenic effect at a relatively low concentration
of 10 µM (2.39 µg/ml), which is comparatively easier to
achieve.
Recently, the protective effect of melatonin in IDD
was also reported. Ge et al (14) revealed that the melatonin was able
to upregulate the expression of collagen-II and aggrecan via
elevating the activity of the ERK signaling pathway in a rabbit
model of IDD. Melatonin also ameliorated IDD via mitophagy
induction and apoptosis inhibition (16). The present study provided similar
results, in that melatonin promoted collagen-II and aggrecan
expression and cell proliferation. Treatment with melatonin may
repair degraded ECM and restore the normal morphology and
biomechanics of IVDs. As a growth factor, TGF-β1 has been studied
for decades and is thought to promote ECM synthesis in NP cells
(8,35,36).
In the present study, TGF-β1 was used as a positive control for
promoting ECM synthesis and the results revealed that melatonin had
a similarly promoting effect to TGF-β1 at current
concentration.
Compared with healthy NP cells, treatment of DNP
cells with melatonin did not produce any significant therapeutic
effects in the present study. This may be attributed to two
reasons: First, DNP cells were isolated from surgical specimens,
which meant that the DNP cells were probably at an advanced stage
of degeneration and were difficult to recover. Furthermore, DNP
cells differed from healthy NP cells in numerous aspects, including
gene expression, proliferative ability, ECM synthesis and
morphology (37). Relatively
long-term treatment may be required to gradually restore the
biological function of DNP cells. The results of the present study
also indicated that the collagen-II content in DNP cells was
increased at day 7, although there was no statistical significance.
Gene therapy may be effectively used to restore the function of DNP
cells (38); however, the
application of a widely used non-genetic agent may be safer and
more acceptable for patients.
In conclusion, the results of the present study
indicated that melatonin inhibited IVD degeneration by promoting
ECM synthesis and suppressing angiogenesis in NP cells. However,
the present study had certain limitations. To make the grouping
clearer, DNP media was not used in the tube formation assays, which
makes the results less persuasive. In addition, delivering agents
precisely to the avascular IVD without iatrogenic damage while
ensuring agents remain at an appropriate concentration in the long
term is challenging. Therapeutic agents may also lose their
activity because of the adverse microenvironment in degenerative
IVD. Further research is required in order to address these
problems.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
CS, YL, LH, FZ and WW designed the study. CS, YL,
YC, LH, FZ and WW performed the experiments. CS and YL analyzed
data. CS and WW wrote the manuscript. All authors read and approved
the final manuscript.
Ethics approval and consent to
participate
The current study was given institutional review
board approval by the Ethics Committee of the Ningbo No. 6 Hospital
(Ningbo, China; approval no. 2016031).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Hart LG, Deyo RA and Cherkin DC: Physician
office visits for low back pain. Frequency, clinical evaluation,
and treatment patterns from a U.S. national survey. Spine (Phila Pa
1976). 20:11–19. 1995.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Katz JN: Lumbar disc disorders and
low-back pain: Socioeconomic factors and consequences. J Bone Joint
Surg Am. 88: (Suppl 2):S21–S24. 2006.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Vo NV, Hartman RA, Yurube T, Jacobs LJ,
Sowa GA and Kang JD: Expression and regulation of
metalloproteinases and their inhibitors in intervertebral disc
aging and degeneration. Spine J. 13:331–341. 2013.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Kepler CK, Ponnappan RK, Tannoury CA,
Risbud MV and Anderson DG: The molecular basis of intervertebral
disc degeneration. Spine J. 13:318–330. 2013.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Lee S, Moon CS, Sul D, Lee J, Bae M, Hong
Y, Lee M, Choi S, Derby R, Kim BJ, et al: Comparison of growth
factor and cytokine expression in patients with degenerated disc
disease and herniated nucleus pulposus. Clin Biochem. 42:1504–1511.
2009.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Kim JS, Ellman MB, Yan D, An HS, Kc R, Li
X, Chen D, Xiao G, Cs-Szabo G, Hoskin DW, et al: Lactoferricin
mediates anti-inflammatory and anti-catabolic effects via
inhibition of IL-1 and LPS activity in the intervertebral disc. J
Cell Physiol. 228:1884–1896. 2013.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Masuda K: Biological repair of the
degenerated intervertebral disc by the injection of growth factors.
Eur Spine J. 17 (Suppl 4):S441–S451. 2008.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Yang H, Cao C, Wu C, Yuan C, Gu Q, Shi Q
and Zou J: TGF-βl suppresses inflammation in cell therapy for
intervertebral disc degeneration. Sci Rep. 5(13254)2015.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Boos N, Weissbach S, Rohrbach H, Weiler C,
Spratt KF and Nerlich AG: Classification of age-related changes in
lumbar intervertebral discs: 2002 Volvo Award in basic science.
Spine (Phila Pa 1976). 27:2631–2644. 2002.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Freemont AJ, Peacock TE, Goupille P,
Hoyland JA, O'Brien J and Jayson MI: Nerve ingrowth into diseased
intervertebral disc in chronic back pain. Lancet. 350:178–181.
1997.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Johnson WE, Caterson B, Eisenstein SM,
Hynds DL, Snow DM and Roberts S: Human intervertebral disc aggrecan
inhibits nerve growth in vitro. Arthritis Rheum. 46:2658–2664.
2002.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Kwon WK, Moon HJ, Kwon TH, Park YK and Kim
JH: Influence of rabbit notochordal cells on symptomatic
intervertebral disc degeneration: Anti-angiogenic capacity on human
endothelial cell proliferation under hypoxia. Osteoarthritis
Cartilage. 25:1738–1746. 2017.PubMed/NCBI View Article : Google Scholar
|
|
13
|
He M, Pang J, Sun H, Zheng G, Lin Y and Ge
W: Overexpression of TIMP3 inhibits discogenic pain by suppressing
angiogenesis and the expression of substance P in nucleus pulposus.
Mol Med Rep. 21:1163–1171. 2020.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Ge J, Zhou Q, Niu J, Wang Y, Yan Q, Wu C,
Qian J, Yang H and Zou J: Melatonin protects intervertebral disc
from degeneration by improving cell survival and function via
activation of the ERK1/2 signaling pathway. Oxid Med Cell Longev.
2019(5120275)2019.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Chang CC, Huang TY, Chen HY, Huang TC, Lin
LC, Chang YJ and Hsia SM: Protective effect of melatonin against
oxidative stress-induced apoptosis and enhanced autophagy in human
retinal pigment epithelium cells. Oxid Med Cell Longev.
2018(9015765)2018.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Chen Y, Wu Y, Shi H, Wang J, Zheng Z, Chen
J, Chen X, Zhang Z, Xu D, Wang X and Xiao J: Melatonin ameliorates
intervertebral disc degeneration via the potential mechanisms of
mitophagy induction and apoptosis inhibition. J Cell Mol Med.
23:2136–2148. 2019.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Cerezo AB, Hornedo-Ortega R,
Álvarez-Fernández MA, Troncoso AM and García-Parrilla MC:
Inhibition of VEGF-induced VEGFR-2 activation and HUVEC migration
by melatonin and other bioactive indolic compounds. Nutrients.
9(249)2017.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Buser Z, Chung AS, Abedi A and Wang JC:
The future of disc surgery and regeneration. Int Orthop.
43:995–1002. 2019.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Freeman BJ and Davenport J: Total disc
replacement in the lumbar spine: A systematic review of the
literature. Eur Spine J. 15: (Suppl 3):S439–S447. 2006.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Van den Eerenbeemt KD, Ostelo RW, van
Royen BJ, Peul WC and van Tulder MW: Total disc replacement surgery
for symptomatic degenerative lumbar disc disease: A systematic
review of the literature. Eur Spine J. 19:1262–1280.
2010.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Wang W, Li P, Xu J, Wu X, Guo Z, Fan L,
Song R, Wang J, Wei L and Teng H: Resveratrol attenuates high
glucose-induced nucleus pulposus cell apoptosis and senescence
through activating the ROS-mediated PI3K/Akt pathway. Biosci Rep.
38(BSR20171454)2018.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Murch SJ, Simmons CB and Saxena PK:
Melatonin in feverfew and other medicinal plants. Lancet.
350:1598–1599. 1997.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Chen G, Huo Y, Tan DX Liang Z, Zhang W and
Zhang Y: Melatonin in Chinese medicinal herbs. Life Sci. 73:19–26.
2003.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Reiter RJ, Tan DX and Galano A: Melatonin:
Exceeding expectations. Physiology (Bethesda). 29:325–333.
2014.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Reiter RJ, Tan DX and Korkmaz A: The
circadian melatonin rhythm and its modulation: Possible impact on
hypertension. J Hypertens (Suppl 27). S17–S20. 2009.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Alvarez-García V, González A,
Alonso-González C, Martínez-Campa C and Cos S: Antiangiogenic
effects of melatonin in endothelial cell cultures. Microvasc Res.
87:25–33. 2013.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Cui P, Yu M, Peng X, Dong L and Yang Z:
Melatonin prevents human pancreatic carcinoma cell PANC-1-induced
human umbilical vein endothelial cell proliferation and migration
by inhibiting vascular endothelial growth factor expression. J
Pineal Res. 52:236–243. 2012.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Alvarez-García V, González A,
Alonso-González C, Martínez-Campa C and Cos S: Regulation of
vascular endothelial growth factor by melatonin in human breast
cancer cells. J Pineal Res. 54:373–380. 2013.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Binch AL, Cole AA, Breakwell LM, Michael
AL, Chiverton N, Cross AK and Le Maitre CL: Expression and
regulation of neurotrophic and angiogenic factors during human
intervertebral disc degeneration. Arthritis Res Ther.
16(416)2014.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Holmes K, Roberts OL, Thomas AM and Cross
MJ: Vascular endothelial growth factor receptor-2: Structure,
function, intracellular signalling and therapeutic inhibition. Cell
Signal. 19:2003–2012. 2007.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Ohba T, Haro H, Ando T, Wako M, Suenaga F,
Aso Y, Koyama K, Hamada Y and Nakao A: TNF-alpha-induced NF-kappaB
signaling reverses age-related declines in VEGF induction and
angiogenic activity in intervertebral disc tissues. J Orthop Res.
27:229–235. 2009.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Cui P, Yu M, Lou Z, Dai M, Han J, Xiu R
and Yang Z: Intracellular signaling pathways involved in cell
growth inhibition of human umbilical vein endothelial cells by
melatonin. J Pineal Res. 44:107–114. 2008.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Cui P, Luo Z, Zhang H, Su Y, Li A, Li H,
Zhang J, Yang Z and Xiu R: Effect and mechanism of melatonin's
action on the proliferation of human umbilical vein endothelial
cells. J Pineal Res. 41:358–362. 2006.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Zhang J, Li Z, Chen F, Liu H, Wang H, Li
X, Liu X, Wang J and Zheng Z: TGF-β1 suppresses CCL3/4 expression
through the ERK signaling pathway and inhibits intervertebral disc
degeneration and inflammation-related pain in a rat model. Exp Mol
Med. 49(e379)2017.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Cai F, Zhu L, Wang F, Shi R, Xie XH, Hong
X, Wang XH and Wu XT: The paracrine effect of degenerated disc
cells on healthy human nucleus pulposus cells is mediated by MAPK
and NF-κB pathways and can be reduced by TGF-β1. DNA Cell Biol.
36:143–158. 2017.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Liu Y, Li Y, Huang ZN, Wang ZY, Nan LP,
Wang F, Zhou SF, Wang JC, Feng XM and Zhang L: The effect of
intervertebral disc degenerative change on biological
characteristics of nucleus pulposus mesenchymal stem cell: An in
vitro study in rats. Connect Tissue Res. 60:376–388.
2019.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Luo XW, Liu K, Chen Z, Zhao M, Han XW, Bai
YG and Feng G: Adenovirus-mediated GDF-5 promotes the extracellular
matrix expression in degenerative nucleus pulposus cells. J
Zhejiang Univ Sci B. 17:30–42. 2016.PubMed/NCBI View Article : Google Scholar
|