Introduction
There are ~1 million new cases of gastric cancer
diagnosed every year worldwide, and the malignancy has become a
leading contributor to cancer-induced deaths in East Asia,
prompting the World Health Organization to declare gastric cancer a
public health concern (1,2). There is a wide geographical variation
in the incidence of gastric cancer, with the highest incidence
rates observed in China, Japan, Latin America and Eastern Europe,
whereas the lowest incidence rates are reported in North America,
parts of Africa and Northern Europe (3,4).
Despite advancements being made in the clinical treatment, the
5-year survival rate of patients with gastric cancer is <30% in
most countries due to a majority of patients being only being
diagnosed at an advanced stage (5).
Increasing research has focused on determining the molecular
mechanisms underlying the development of gastric cancer, which may
provide a basis for discovering novel therapeutic targets.
Tac2-N (TC2N), located on human chromosome 14q32.12,
belongs to the carboxyl-terminal type tandem C2 protein family, and
is a putative C2 domain-containing protein (6). Although the function of the C2 domain
was originally related to calcium-dependent phospholipid binding, a
previous study has also indicated that the C2 domain may be
involved in cellular signal transduction and protein-protein
interactions (7). Furthermore,
other proteins containing C2 domains were also reported to be
involved in the modulation of tumorigenesis. For example, in a
previous study, the double C2-like domain β (DOC2B) gene was
identified to be closely associated with cervical carcinogenesis,
as the downregulated expression levels of DOC2B led to selective
cervical tumor growth, which was dependent on the methylation of
its DNA promoter (8). In addition,
copines were demonstrated to effectively increase cellular
migration in breast cancer through binding with c-Jun activation
domain binding protein-1(9). It was
also reported that TC2N served as an oncogene and regulated tumor
metastasis in lung cancer, while serving as a tumor suppressor in
breast cancer (6,10,11).
Therefore, due to various genes serving a dual role in cancer, the
role of TC2N in different types of cancer should also be
investigated.
The present study aimed to investigate the precise
role of TC2N in gastric cancer occurrence and development, and to
determine the regulatory mechanism of TC2N in gastric cancer. The
results revealed a previously unknown role of TC2N in gastric
cancer, and uncovered its potential mechanism underlying the
therapeutic targeting of TC2N to inhibit gastric cancer
progression.
Materials and methods
Bioinformatics analysis of TC2N
expression levels in gastric cancer
Data from The Cancer Genome Atlas (TCGA; https://portal.gdc.cancer.gov) database, including 375
gastric tumor tissues and 32 normal gastric tissues (adjacent
cancer samples), were used to analyze the expression levels of TC2N
(12). TC2N expression levels and
clinical data, including age, stage and overall survival, were
downloaded from the TCGA data portal. Patients with known TC2N
expression and complete clinical data were included in the present
study, while those with missing clinical data were excluded.
Cell culture
The normal gastric mucosa cell line GES1 was
obtained from Beijing Beina Chuanglian Biotechnology Institute and
the gastric cancer cell line AGS was obtained from The Cell Bank of
Type Culture Collection of the Chinese Academy of Sciences. Cells
were cultured in RPMI-1640 medium (Invitrogen; Thermo Fisher
Scientific, Inc.), supplemented with 10% FBS (Invitrogen; Thermo
Fisher Scientific, Inc.), and maintained in a humidified atmosphere
with 5% CO2 at 37˚C.
Cell transfection
For the overexpression and knockdown of TC2N,
pcDNA3.1-TC2N [overexpression (OE)-TC2N], the specific short
hairpin (sh)RNA vector targeting TC2N-1/2 (shRNA-TC2N-1/2), and
their corresponding negative control (NC) vectors (OE-NC empty
vector and scrambled shRNA-NC, respectively) were purchased from
Shanghai GenePharma Co., Ltd. AGS cells (1x104
cells/well) were seeded into a six-well culture plate and cultured
to 60-70% confluence. The vectors (500 ng/µl) were transfected into
AGS cells at a dose of 10 nM using Lipofectamine™ 2000 (Invitrogen;
Thermo Fisher Scientific, Inc.), according to the manufacturer's
protocol. Following 48 h of transfection at 37˚C, cells were
harvested for reverse transcription-quantitative PCR (RT-qPCR) and
western blotting analysis to determine the transfection
efficiency.
Cell treatment
To determine the function of TC2N on the sensitivity
of AGS cells to chemotherapy, following transfection for 48 h, AGS
cells were collected. Subsequently, the transfected AGS cells were
re-seeded into 96-well plates (1x104 cells/well).
Following 24 h, the cells were treated with different
concentrations of cisplatin (DPP; 0, 10, 20 or 50 µmol/l),
paclitaxel (0, 10, 20 or 50 nmol/l) and 5-fluorouracil (5-FU; 0,
10, 20 or 50 µmol/l; all from Sigma-Aldrich; Merck KGaA) for 24 h
in a humidified atmosphere with 5% CO2 at 37˚C,
respectively. The survival rate (viability) of each group was then
determined using CCK-8 assay via measuring the absorbance values at
450 nm, as described below.
Cell proliferation assay
To determine the cell proliferation rate of AGS
cells, a Cell Counting Kit-8 (CCK-8; Dojindo Molecular
Technologies, Inc.) assay was performed using transfected and
non-transfected (control) cells, according to the manufacturer's
protocol. Briefly, the cells were seeded into 96-well plates at a
density of 1x104 cells/well and incubated in a
humidified atmosphere with 5% CO2 at 37˚C for 24, 48 and
72 h. Subsequently, 10 µl CCK-8 solution was added/well at each
time point following incubation for 4 h at 37˚C, and the absorbance
(optical density) values were measured using a microplate reader at
a wavelength of 450 nm.
Wound healing assay
A wound healing assay was performed to analyze the
cell migration rate. Following 48 h of transfection, AGS cells were
seeded into 6-well plates (2x105 cells/well) and
cultured in a humidified atmosphere with 5% CO2 at 37˚C.
Subsequently, straight lines were scratched into the 100% confluent
cell monolayer using a 20-µl pipette tip. The cell debris was
removed by washing with PBS and then the cells were incubated with
fresh serum-free RPMI-1640 medium. Following 24 h of incubation at
37˚C, the wound healing process was analyzed under a light
microscope (magnification, x100; CKX41, Olympus Corporation). The
migratory distance was measured using ImageJ software version 1.46
(National Institutes of Health). Migration rate was determined
using representative images obtained at 0 and 24 h using the
following formula: Migration rate (%) = (migration
distance/original distance) x100. The original distance represented
the width of the wound at 0 h, and the migration distance
represented the original width of the wound at 0 h, minus the width
of the wound at 24 h. The relative migration rate was presented as
the fold change relative to the control group.
Cell invasion assay
A Transwell Matrigel assay was performed to
determine the cell invasion rate. At 48 h post-transfection,
2x104 cells were suspended in serum-free RPMI-1640
medium and plated into the upper chambers of Transwell plates,
which were precoated with 40 µl Matrigel (BD Biosciences) at 37˚C
for 30 min. Complete RPMI-1640 medium supplemented with 10% FBS was
plated in the lower chambers. Following incubation for 24 h at
37˚C, non-invasive cells remaining in the upper chamber were
removed using a cotton swab, while the invasive cells in the lower
chamber were fixed with 4% formaldehyde for 10 min at room
temperature and stained with 0.05% crystal violet for 10 min at
room temperature. Stained cells were visualized and counted using a
light microscope (magnification, x100).
Cell cycle assay
Flow cytometric analysis was performed to determine
whether TC2N regulated the growth phase of AGS cells. Following 48
h of transfection, cells were harvested by centrifugation (4˚C;
1,000 x g; 5 min) and fixed with 70% ethanol at -20˚C overnight.
Cells were washed twice with PBS and stained with a Cell Cycle
Detection Kit (Nanjing KeyGen Biotech Co., Ltd.) containing 200 µl
PBS, 1 µl propidium iodide (1 mg/ml) and 1 µl RNase (10 mg/ml) for
20 min at 37˚C in the dark. Cell cycle distribution was analyzed
using flow cytometry (CYTOMICS FC 500; Beckman Coulter, Inc.) and
BD CellQuest Pro version 5.1 software (BD Biosciences).
RNA extraction and RT-qPCR
Total RNA from the cells was extracted using
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.). The concentration and purity of the RNA was detected using
the A260/A280 ratio of 1.8-2.0 with NanoDrop
2000 (Thermo Fisher Scientific, Inc.). A total of 1 µg total RNA
was reverse transcribed into cDNA using the PrimeScript RT Master
mix kit (Takara Bio, Inc.). The following conditions were used for
the reverse transcription: 42˚C for 2 min, 37˚C for 15 min and 85˚C
for 5 sec. qPCR was subsequently performed using the SYBR Premix Ex
Taq kit (Takara Bio, Inc.). The following primer pairs were used
for qPCR: TC2N forward, 5'-TGGCTGTACTGAGGATTATTTGC-3' and reverse,
5'-TGTGAAGGAGTTTCTTGTGTCC-3'; and GAPDH forward,
5'-CCATCTTCCAGGAGCGAGAT-3' and reverse, 5'-TGCTGATGATCTTGAGGCTG-3'.
The following thermocycling conditions were used for the qPCR:
Initial denaturation at 95˚C for 10 min; 40 cycles at 94˚C for 2
min and 60˚C for 50 sec; and a final extension at 60˚C for 1 min.
The relative mRNA expression levels were determined using the
2-ΔΔCq method (13) and normalized to the GAPDH loading
control. All experiments were repeated ≥3 times.
Western blotting
Total protein was extracted from cells in six-well
plates on ice using RIPA lysis buffer (Invitrogen; Thermo Fisher
Scientific, Inc.). The lysate was centrifuged at 12,000 x g at 4˚C
for 10 min and the supernatant was collected. Total protein was
quantified using a Bradford assay and equal amounts of protein (25
µg per lane) were separated via 12% SDS-PAGE at 120 V. The
separated proteins were subsequently transferred onto PVDF
membranes (EMD Millipore) and blocked at room temperature with 5%
skimmed milk for 1 h. The membranes were then incubated overnight
at 4˚C with the following primary antibodies (all diluted 1:1,000):
Anti-TC2N (cat. no. PA5-32086; Invitrogen; Thermo Fisher
Scientific, Inc.), anti-cyclin D1 (cat. no. ab134175; Abcam),
anti-CDK4 (cat. no. ab108357; Abcam), anti-cyclin E1 (cat. no.
ab33911; Abcam), anti-matrix metalloproteinase (MMP)2 (cat. no.
ab92536; Abcam), anti-MMP9 (cat. no. ab38898; Abcam), anti-N-Myc
downstream regulated gene 1 (NDRG1; cat. no. ab124689; Abcam),
anti-P-glycoprotein (P-gp; cat. no. ab170904; Abcam) and anti-GAPDH
(cat. no. ab9485; Abcam). Following the primary antibody
incubation, the membranes were washed three times with TBS-0.05%
Tween-20 (Sigma-Aldrich; Merck KGaA) and incubated with a mouse
anti-rabbit horseradish peroxidase-conjugated secondary antibody
(1:3,000; cat. no. sc-2357; Santa Cruz Biotechnology, Inc.).
Protein bands were visualized using an ECL western blotting
substrate (GE Healthcare) and semi-quantitatively analyzed using
Quantity One version 4.4.0 software (Bio-Rad Laboratories,
Inc.).
Statistical analysis
All data are presented as the mean ± SD from ≥3
independent experiments. Statistical analysis was performed using
SPSS version 18.0 software (SPSS, Inc.). Statistical significance
was analyzed using a one-way ANOVA, followed by a Tukey's post hoc
test for multiple comparisons (Figs.
2-5), whereas the expression levels of TC2N in normal tissues
and tumors tissues were analyzed using a U-Mann Whitney test
(Fig. 1A). Low and high expression
levels of TC2N in gastric cancer samples were defined according to
the median, and those above the median were defined as high
expression and those under the median were defined as low
expression. The prognosis was analyzed using a Kaplan-Meier test,
followed by a log-rank test. The association between TC2N
expression levels and clinical parameters were analyzed using a
U-Mann Whitney test between 2 groups, or a Kruskal Wallis test for
>2 groups (Fig. 1B and D). P<0.05 was considered to indicate a
statistically significant difference.
Results
TC2N expression levels are upregulated
in gastric cancer tissues and cell lines
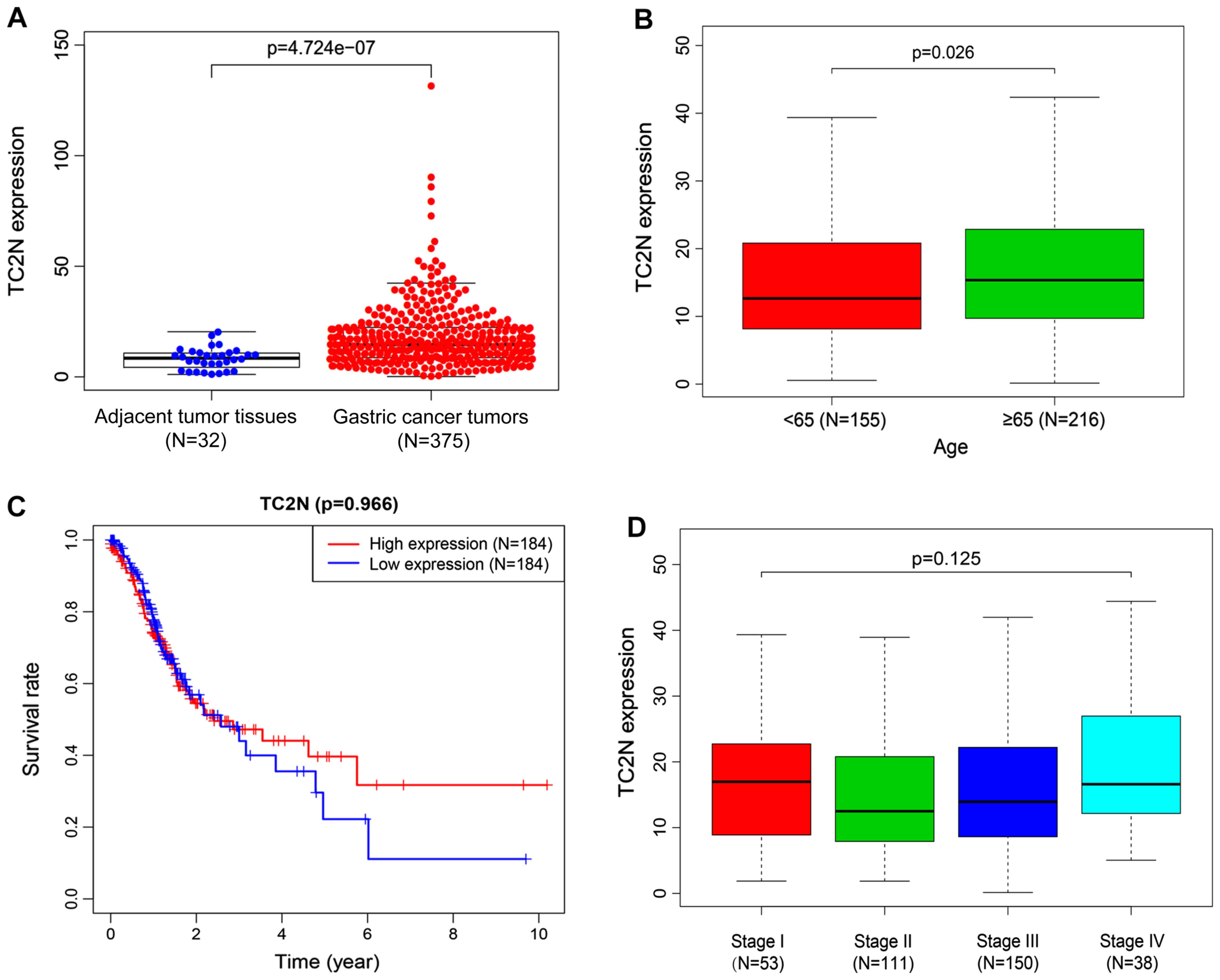
The data from TCGA database revealed that the
expression levels of TC2N were significantly upregulated in gastric
cancer tissues compared with the adjacent tumor tissues (Fig. 1A). Notably, TC2N was discovered to
be differentially expressed in patients with gastric cancer
depending on the age; the expression levels of TC2N were
significantly upregulated in patients of ≥65 years old compared
with patients that were <65 years old, excluding those without
clinical information on age (Fig.
1B). In addition, the survival rate in patients with high
expression levels of TC2N was observed to be increased compared
with patients with low expression levels of TC2N, excluding the
patients without clinical information on survival (Fig. 1C); however, there was no significant
difference identified between the two groups in the overall
survival rate. Furthermore, the expression levels of TC2N were
indicated to be independent of the tumor stage of patients, as no
significant differences were observed between patients with
different stages of gastric cancer, excluding the patients without
information on the clinical stage (Fig.
1D). These findings indicated that the expression levels of
TC2N may be upregulated in gastric cancer; however, this was likely
to be independent of the degree of malignancy.
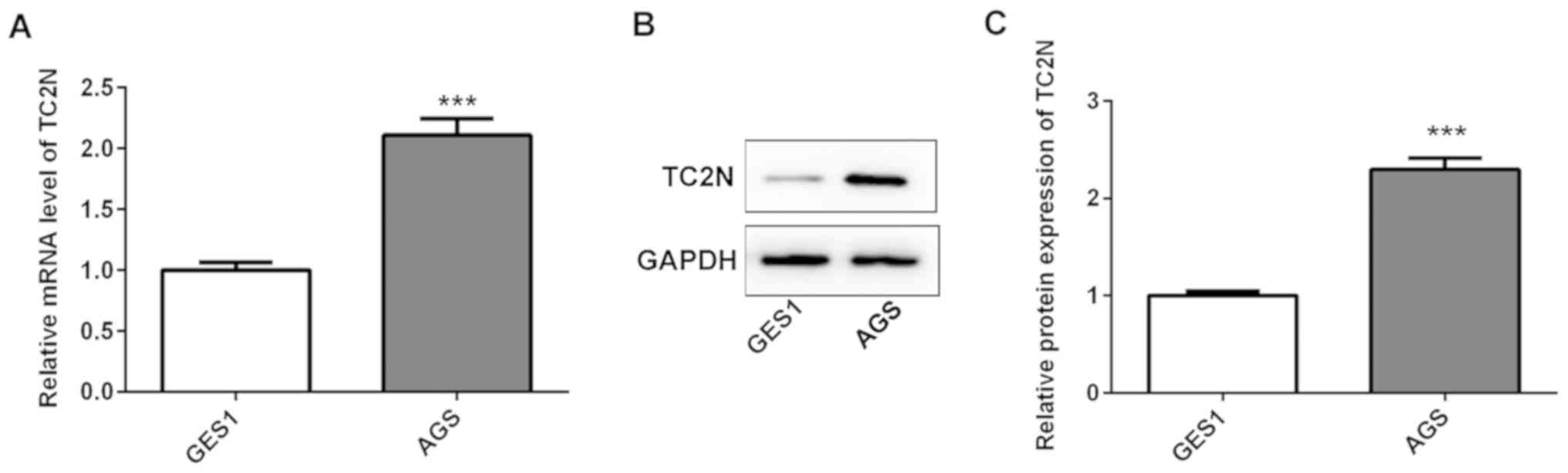
To further investigate the role of TC2N, TC2N
expression levels in GES1 and AGS cells were analyzed using RT-qPCR
and western blotting. The present data demonstrated that compared
with the GES1 cells, the mRNA and protein expression levels of TC2N
were significantly upregulated in AGS cells (Fig. 2A-C).
TC2N regulates the cell proliferation
of AGS cells
To determine the function of TC2N in gastric cancer,
the effects of TC2N knockdown or overexpression on AGS cell
proliferation were analyzed. Both the mRNA and protein expression
levels of TC2N were significantly downregulated following the
transfection with shRNA-TC2N-1/2 compared with the shRNA-NC group
(Fig. 3A and B), and shRNA-TC2N-1 was selected for
experiments due to its higher knockdown efficacy. The expression
levels of TC2N were significantly upregulated following the
transfection with OE-TC2N compared with the OE-NC group (Fig. 3C and D). Flow cytometric analysis revealed a
significant increase in the percentage of cells in the G0/G1 phase
and a decrease in the percentage of cells in the S phase following
TC2N knockdown compared with the shRNA-NC group (Fig. 3E and F). By contrast, the percentage of cells in
the G0/G1 phase was significantly decreased, while the percentage
in the S phase was significantly increased following TC2N
overexpression compared with the OE-NC group. The effect of TC2N
knockdown or TC2N overexpression on G2/M was not significantly
different (Fig. 3E and F). These results indicated that TC2N
knockdown may induce G1/S phase arrest, whereas TC2N overexpression
may inhibit G1/S phase arrest. CCK-8 analysis also revealed that
the cell proliferative ability was significantly decreased in the
shRNA-TC2N group compared with the shRNA-NC group at 72 h, whereas
the proliferative ability was significantly increased following
TC2N overexpression compared with the OE-NC group at 72 h (Fig. 3G). Moreover, the protein expression
levels of cyclin D1, CDK4 and cyclin E1 were significantly
downregulated following TC2N knockdown and upregulated following
TC2N overexpression, compared with their respective NCs (Fig. 3H), which is consistent with the
previous results from CCK-8 and flow cytometry assays. These
findings indicated that TC2N may have a regulatory role over cell
proliferation, as the knockdown of TC2N inhibited cell
proliferation, whereas the overexpression of TC2N promoted cell
proliferation.
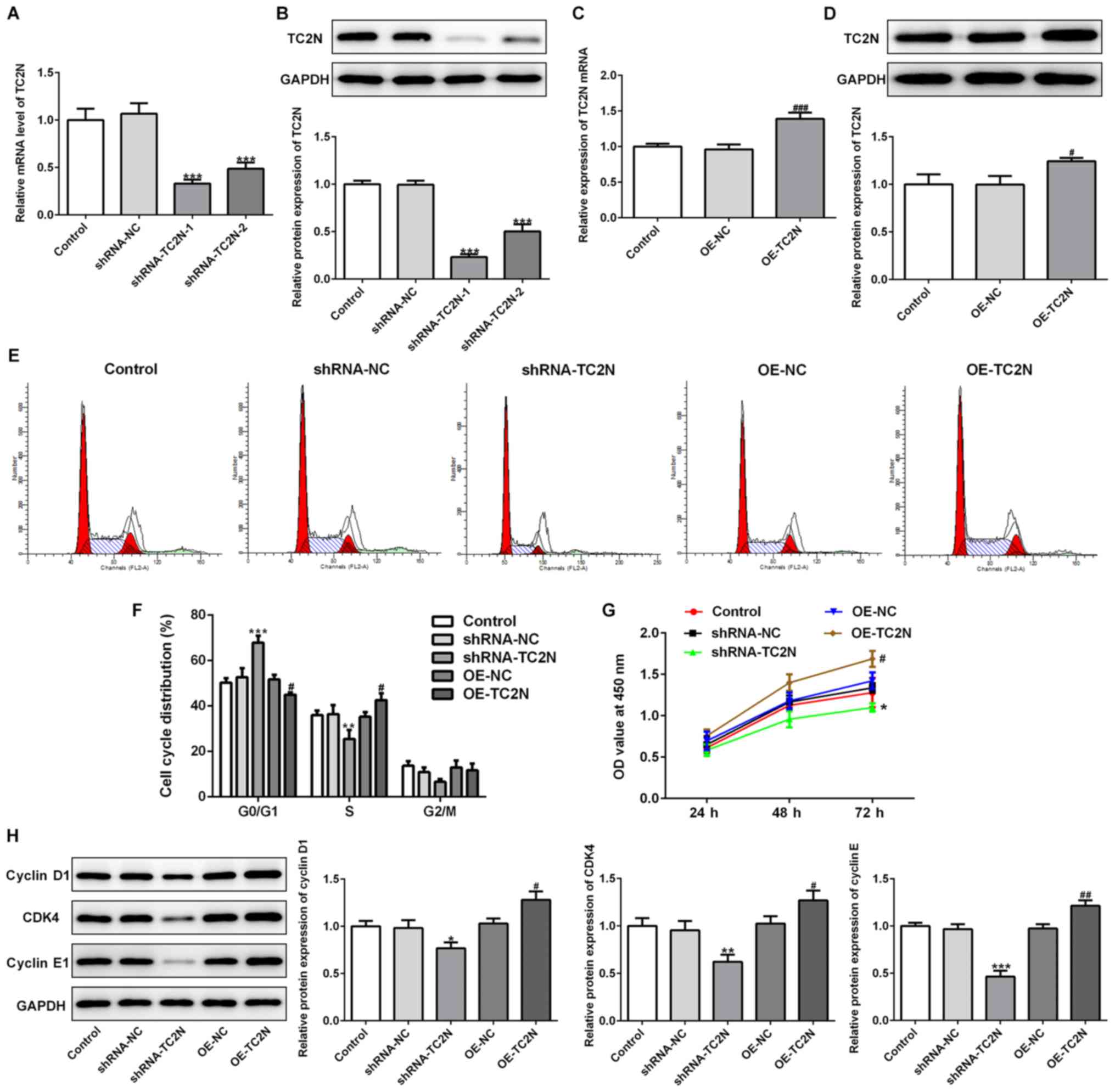 | Figure 3TC2N regulates the cell proliferation
of AGS cells. To determine the function of TC2N in gastric cancer,
AGS cells were transfected with shRNA-TC2N and the (A) mRNA and (B)
protein expression levels of TC2N were analyzed using RT-qPCR and
western blotting, respectively. AGS cells were transfected with
OE-TC2N and the (C) mRNA and (D) protein expression levels of TC2N
were determined using RT-qPCR and western blotting, respectively.
(E) Cell cycle distribution was analyzed using flow cytometric
analysis. (F) Quantification of cell cycle distribution presented
in part (E). (G) Cell Counting Kit-8 assay was performed to
determine the cell proliferation rate. (H) Protein expression
levels of cyclin D1, CDK4 and cyclin E1 were analyzed using western
blotting. *P<0.05, **P<0.01,
***P<0.001 vs. shRNA-NC; #P<0.05,
##P<0.01, ###P<0.001 vs. OE-NC. TC2N,
Tac2-N; shRNA, short hairpin RNA; RT-qPCR, reverse
transcription-quantitative PCR; OE, overexpression; NC, negative
control; OD, optical density. |
TC2N regulates the cell migration and
invasion of AGS cells
The effects of TC2N knockdown or overexpression on
the migratory and invasive ability of AGS cells were further
investigated. The wound healing and Transwell Matrigel assays
demonstrated that TC2N knockdown significantly inhibited both cell
migration and invasion compared with the shRNA-NC group, while TC2N
overexpression significantly promoted cell migration and invasion
compared with the OE-NC group (Fig.
4A-C). MMP2 and MMP9 are known to be involved in cell invasion
and tumor metastasis (14). NDRG1
has been demonstrated to repress tumor metastasis, and downregulate
MMP2 and MMP9 expression levels (15,16).
Therefore, MMP2, MMP9 and NDRG1 were investigated in the present
study to determine whether they were involved in the effects of
TC2N on cell migration and invasion. The results revealed that TC2N
knockdown significantly downregulated the protein expression levels
of MMP2 and MMP9, whereas the protein expression levels of NDRG1
were significantly upregulated, compared with the shRNA-NC group
(Fig. 4D). Conversely, the
overexpression of TC2N exhibited the opposite effects in AGS cells
(Fig. 4D), indicating that TC2N may
affect the gastric cancer migrative and invasive abilities partly
via regulating MMP2 and MMP9 expression levels.
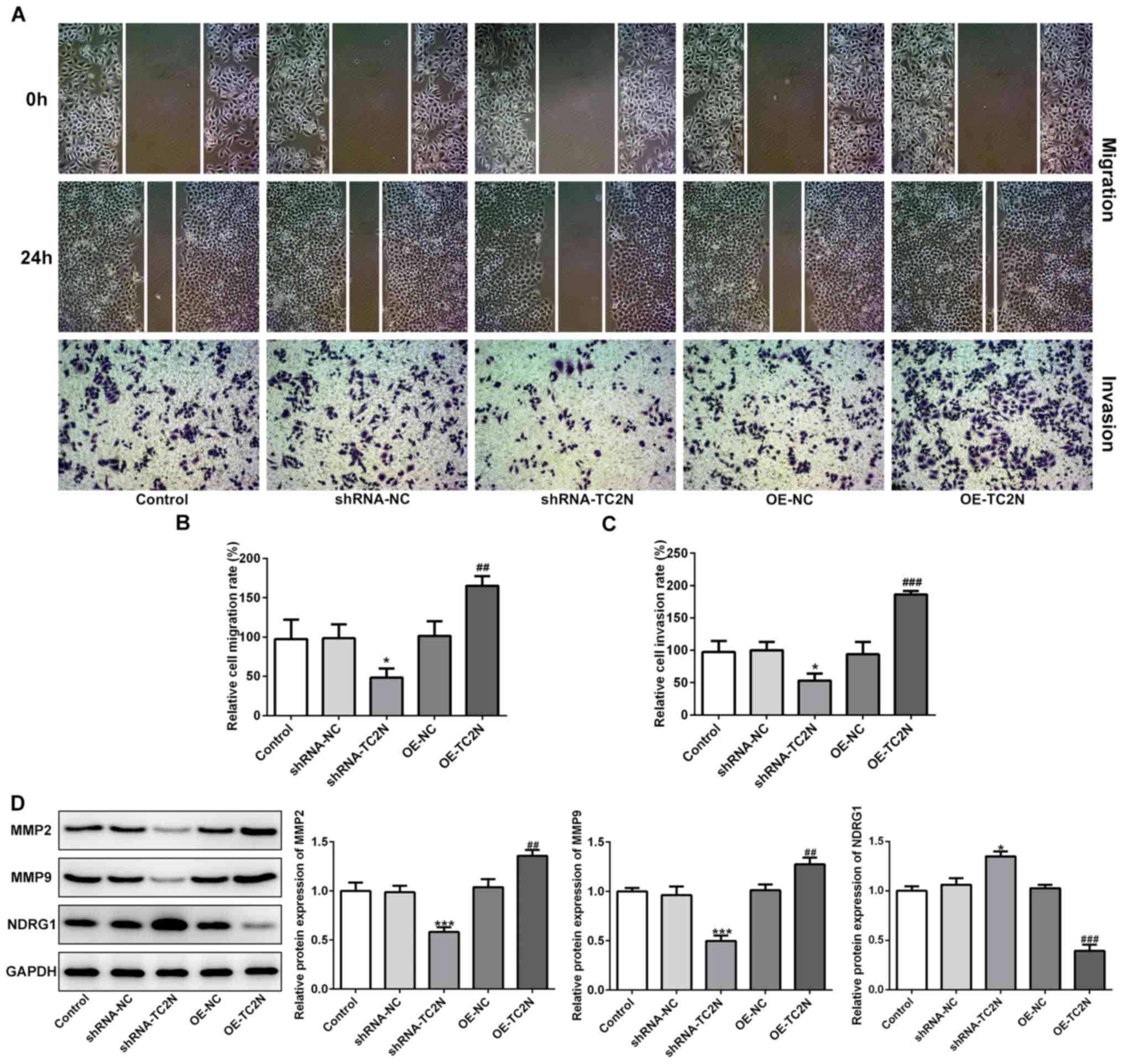 | Figure 4TC2N regulates the cell migration and
invasion of AGS cells. (A) Wound healing and Transwell Matrigel
assays were performed to analyze the cell migratory and invasive
abilities, respectively (magnification, x 100). (B)
Semi-quantification of the cell migration rate from part (A). (C)
Semi-quantification of the cell invasion rate from part (A). (D)
Protein expression levels of MMP2, MMP9 and NDRG1 were analyzed
using western blotting. *P<0.05,
***P<0.001 vs. shRNA-NC; ##P<0.01,
###P<0.001 vs. OE-NC. TC2N, Tac2-N; shRNA, short
hairpin RNA; OE, overexpression; NC, negative control; MMP, matrix
metalloproteinase; NDRG1, N-Myc downstream regulated gene 1. |
TC2N mediates drug resistance of AGS
cells
To determine the function of TC2N on the sensitivity
of AGS cells to chemotherapy, cells were treated with different
concentrations of DPP, paclitaxel and 5-FU for 24 h and the
survival rate of each group was detected using a CCK-8 assay. The
cell survival rate was discovered to be closely associated with the
drug concentration, as the survival rate of the cells reduced upon
increasing the drug concentration (Fig.
5A-C). Furthermore, TC2N knockdown significantly decreased the
survival rate of the cells following 50 µmol/l DPP or 5-FU or 50
nmol/l paclitaxel treatment compared with the shRNA-NC group,
whereas TC2N overexpression exerted the opposite effects,
indicating that TC2N knockdown may improve the sensitivity of AGS
cells to DDP, paclitaxel and 5-FU. The inhibition of P-gp has been
considered to be an effective approach for reversing cancer-related
drug resistance (17). In the
present study, TC2N knockdown significantly downregulated the
expression levels of P-gp compared with the shRNA-NC group, whereas
TC2N overexpression significantly upregulated the expression levels
of P-gp compared with the OE-TC2N group (Fig. 5D and E), indicating that TC2N knockdown may
improve the sensitivity of AGS cells to chemotherapeutics via the
inhibition of P-gp expression.
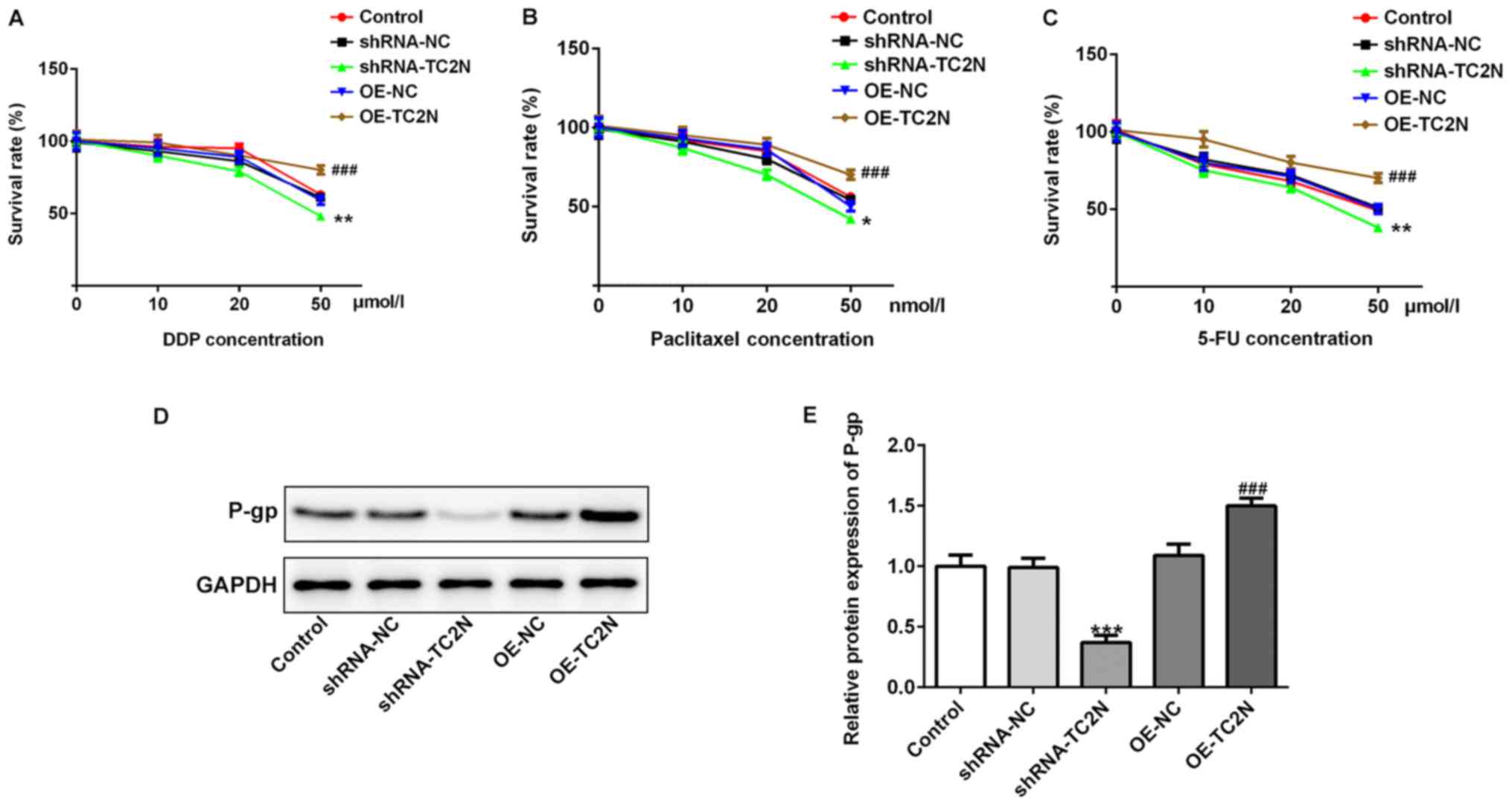 | Figure 5TC2N mediates the drug resistance of
AGS cells. AGS cells were treated with different concentrations of
DPP (0, 10, 20 or 50 µmol/l), paclitaxel (0, 10, 20 or 50 nmol/l)
and 5-FU (0, 10, 20 or 50 µmol/l) for 24 h. The survival rate of
each group following (A) DDP, (B) paclitaxel and (C) 5-FU treatment
was analyzed using Cell Counting Kit-8 assays. (D) Protein
expression levels of P-gp were determined using western blotting.
(E) Semi-quantification of the P-gp expression levels from part
(D). *P<0.05, **P<0.01,
***P<0.001 vs. shRNA-NC; ###P<0.001 vs.
OE-NC. TC2N, Tac2-N; shRNA, short hairpin RNA; OE, overexpression;
NC, negative control; DPP, cisplatin; 5-FU, 5-fluorouracil; P-gp,
P-glycoprotein. |
Discussion
Gastric cancer is one of the most common types of
malignant tumor worldwide (18). To
date, surgical resection remains the only curative modality for
localized gastric cancer, accompanied with perioperative
chemotherapy or postoperative chemoradiotherapy to improve the
outcomes (18). For advanced and
metastatic disease, the combination of a fluoropyrimidine and a
platinum compound-containing chemotherapy, with the incorporation
of trastuzumab for the HER2-enriched population, remains the
standard care for first-line therapy (3). However, due to the high degree of
biological malignancy of gastric cancer itself, especially at the
advanced stage, the 5-year survival rate of gastric cancer remains
poor and does not exceed 30% in the USA and Europe (18). Therefore, determining the molecular
mechanism underlying the development of gastric cancer is required.
TC2N was recently identified as a novel oncogene in lung cancer
(10,11); however, the role of TC2N in gastric
cancer remains unclear. In the present study, upregulated
expression levels of TC2N were identified in gastric cancer cells
and tumors of patients with gastric cancer via TCGA analysis. To
determine the role of TC2N in gastric cancer, further studies
revealed that TC2N regulated the cell proliferation, migration and
invasion of gastric cancer cells. Furthermore, the knockdown of
TC2N strengthened the sensitivity of AGS cells to DPP, paclitaxel
and 5-FU, and downregulated P-gp expression levels, which indicated
that TC2N may be an important regulator in gastric cancer.
The overexpression of TC2N in AGS cells
significantly increased the cell proliferation, and promoted cell
migration and invasion, whereas the knockdown of TC2N exerted the
opposite effects, which is consistent with previous studies, which
reported an oncogenic function of TC2N (10,11).
Cell proliferation is predominantly regulated by the cell cycle,
which is a complex network of regulatory process involving multiple
signaling pathways (19). For
example, T-Box transcription factor TBX3 has been indicated to
function as an immortalizing gene promoting proliferation via
actively repressing negative cell cycle regulators, which has been
associated with the c-Myc-mediated pathway (20). Moreover, xanthoxyletin has been
revealed to inhibit proliferation and induce cell cycle arrest via
modulating the MEK/ERK signaling pathway (20). Thus, the effect of TC2N on cell
cycle progression and cell cycle-related proteins was further
determined The experimental results revealed that TC2N
overexpression increased the percentage of cells in the S phase,
and upregulated the expression levels of cyclin D1, cyclin E1 and
CDK4, while TC2N knockdown achieved the opposite effects. Cyclin
D1, cyclin E1 and CDK4 all serve important roles during G1-S phase
cell cycle progression, and cyclins and CDKs can directly
participate in cell cycle regulation (21). In particular, cyclin D1 and cyclin
E1 are important members of the cyclin family, which are
responsible for cell transformation from the G1 to S phase
(22). Therefore, these results
implied that TC2N knockdown may result in the downregulation of
cell cycle-related proteins to prolong the cell cycle, regulating
the progression of gastric cancer.
To investigate the role of TC2N in drug resistance
and determine the potential mechanism of action, three different
types of chemotherapeutic drugs (DPP, paclitaxel and 5-FU) were
used to treat AGS cells following TC2N overexpression or knockdown.
The survival rate of AGS cells varied depending on the expression
levels of TC2N; TC2N knockdown significantly decreased the cell
survival rate, reflecting a higher sensitivity of AGS cells to DPP,
paclitaxel and 5-FU when TC2N expression was inhibited. Acquired
drug resistance is one of the predominant reasons for the low cure
rate and high recurrence rate in patients with gastric cancer,
especially at an advanced stage (23). Thus, improving the sensitivity of
cancer cells to chemotherapeutics may be an important strategy for
cancer treatment. Reducing the resistance of cancer cells to
chemotherapeutics has been indicated as a potential strategy to
improve the therapeutic effects in numerous types of cancer
(24-26).
In the present study, the findings suggested that TC2N may serve as
an oncogene, which not only promoted gastric cancer cell
proliferation, migration and invasion, but also regulated the
sensitivity of gastric cancer cells to DPP, paclitaxel and 5-FU.
The predominant mechanism underlying cancer cell resistance
involves the drug efflux pump, P-gp (27). P-gp pumps intracellular drugs out of
cells as an energy-dependent drug efflux pump, resulting in drug
resistance caused by a reduced intracellular concentration of drugs
(28). In a previous study, P-gp
expression levels were discovered to be upregulated in gastric
cancer cells presenting with drug resistance (29). In the present study, the expression
levels of P-gp were significantly upregulated following TC2N
overexpression and AGS cells had a higher sensitivity to DPP,
paclitaxel and 5-FU, whereas the opposite effects were observed
when TC2N expression was knocked down. These results indicated that
TC2N may influence the sensitivity of AGS cells to drugs via
regulating P-gp expression levels.
To the best of our knowledge, this study was the
first to determine a previously unknown role of TC2N in gastric
cancer and uncover its potential mechanism; however, further
experiments and clinical data are required to validate and make the
conclusions reliable. However, there are some limitations to the
present study. Firstly, the knockdown of TC2N significantly
improved the sensitivity of AGS cells to chemotherapeutics;
however, whether the improvement of drug sensitivity is also
reflected in the processes of cell migration, invasion and cell
cycle distribution remains to be elucidated. Secondly, the data in
this study were predominantly based on cell lines, and data from an
animal model were lacking; thus, in vivo experiments are
required to verify the therapeutic effect of TC2N in gastric
cancer. Finally, further clinical data are also required to
determine the diagnostic and prognostic value of TC2N in gastric
cancer. These limitations are worthy of being studied and our
future work aims to investigate these points further.
In conclusion, the present study provided evidence
that suggested that TC2N expression levels may be upregulated in
AGS cells. The knockdown of TC2N expression inhibited the
proliferation, migration and invasion of cells, in addition to
inducing cell cycle G1/S phase arrest. Furthermore, the knockdown
of TC2N partially reversed the cell resistance to DPP, paclitaxel
and 5-FU, which was indicated to be partly through inhibiting P-gp
expression levels, whereas TC2N overexpression exerted the opposite
effects on AGS cells. Thus, TC2N may serve as an important
regulator in gastric cancer.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
PJ conceptualized and designed the study, wrote and
proof-read the manuscript. PZ created the figures and provided
important comments during the revision of the manuscript. LS, PZ
and JW participated in the literature collection, experiment
conduction and analysis of the data. All authors read and approved
the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Song Z, Wu Y, Yang J, Yang D and Fang X:
Progress in the treatment of advanced gastric cancer. Tumour Biol.
39(1010428317714626)2017.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Ferro A, Peleteiro B, Malvezzi M, Bosetti
C, Bertuccio P, Levi F, Negri E, La Vecchia C and Lunet N:
Worldwide trends in gastric cancer mortality (1980-2011), with
predictions to. 2015, and incidence by subtype. Eur J Cancer.
50:1330–1344. 2014.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Charalampakis N, Economopoulou P,
Kotsantis I, Tolia M, Schizas D, Liakakos T, Elimova E, Ajani JA
and Psyrri A: Medical management of gastric cancer: A 2017 update.
Cancer Med. 7:123–133. 2018.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Russo A, Li P and Strong VE: Differences
in the multimodal treatment of gastric cancer: East versus west. J
Surg Oncol. 115:603–614. 2017.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Hamashima C: Current issues and future
perspectives of gastric cancer screening. World J Gastroenterol.
20:13767–13774. 2014.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Hao XL, Han F, Zhang N, Chen HQ, Jiang X,
Yin L, Liu WB, Wang DD, Chen JP, Cui ZH, et al: TC2N, a novel
oncogene, accelerates tumor progression by suppressing p53
signaling pathway in lung cancer. Cell Death Differ. 26:1235–1250.
2019.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Farah CA and Sossin WS: The role of C2
domains in PKC signaling. Adv Exp Med Biol. 740:663–683.
2012.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Kabekkodu SP, Bhat S, Radhakrishnan R,
Aithal A, Mascarenhas R, Pandey D, Rai L, Kushtagi P, Mundyat GP
and Satyamoorthy K: DNA promoter methylation-dependent
transcription of the double C2-like domain β (DOC2B) gene regulates
tumor growth in human cervical cancer. J Biol Chem.
289:10637–10649. 2014.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Choi HY, Park N, Na JB, Ko ES, Park JY and
Yoo JC: Direct binding of Copine3 with Jab1 activates downstream
ErbB2 signaling and motility in SKBr3 breast cancer cells. Oncol
Rep. 35:1147–1152. 2016.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Hao X, Gao LY, Zhang N, Chen H, Jiang X,
Liu W, Ao L, Cao J, Han F and Liu J: Tac2-N acts as a novel
oncogene and promotes tumor metastasis via activation of NF-κB
signaling in lung cancer. J Exp Clin Cancer Res.
38(319)2019.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Hao XL, Gao LY, Deng XJ, Han F, Chen HQ,
Jiang X, Liu WB, Wang DD, Chen JP, Cui ZH, et al: Identification of
TC2N as a novel promising suppressor of PI3K-AKT signaling in
breast cancer. Cell Death Dis. 10(424)2019.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Tomczak K, Czerwińska P and Wiznerowicz M:
The cancer genome atlas (TCGA): An immeasurable source of
knowledge. Contemp Oncol (Pozn). 19:A68–A77. 2015.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Wang X, Yang B, She Y and Ye Y: The lncRNA
TP73-AS1 promotes ovarian cancer cell proliferation and metastasis
via modulation of MMP2 and MMP9. J Cell Biochem. 119:7790–7799.
2018.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Tian S, Wang X and Proud CG: Oncogenic MNK
signalling regulates the metastasis suppressor NDRG1. Oncotarget.
8:46121–46135. 2017.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Fu Y, Wei J, Dai X and Ye Y: Increased
NDRG1 expression attenuate trophoblast invasion through ERK/MMP-9
pathway in preeclampsia. Placenta. 51:76–81. 2017.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Battistella C and Klok HA: Reversion of
P-gp-mediated drug resistance in ovarian carcinoma cells with
PHPMA-zosuquidar conjugates. Biomacromolecules. 18:1855–1865.
2017.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Ciesielski M, Kruszewski WJ, Walczak J,
Szajewski M, Szefel J, Wydra J, Buczek T and Czerepko M: Analysis
of postoperative morbidity and mortality following surgery for
gastric cancer. Surgeon volume as the most significant prognostic
factor. Prz Gastroenterol. 12:215–221. 2017.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Willmer T, Peres J, Mowla S, Abrahams A
and Prince S: The T-Box factor TBX3 is important in S-phase and is
regulated by c-Myc and cyclin A-CDK2. Cell Cycle. 14:3173–3183.
2015.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Wen Q, Luo K, Huang H, Liao W and Yang H:
Xanthoxyletin inhibits proliferation of human oral squamous
carcinoma cells and induces apoptosis, autophagy, and cell cycle
arrest by modulation of the MEK/ERK signaling pathway. Med Sci
Monit. 25:8025–8033. 2019.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Galderisi U, Jori FP and Giordano A: Cell
cycle regulation and neural differentiation. Oncogene.
22:5208–5219. 2003.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Liu W, Hua S, Dai Y, Yuan Y, Yang J, Deng
J, Huo Y, Chen X, Teng B, Yu X and Zhang Y: Roles of Cx43 and
AKAP95 in ovarian cancer tissues in G1/S phase. Int J Clin Exp
Pathol. 8:14315–14324. 2015.PubMed/NCBI
|
|
23
|
Yao Y, Yang X, Sun L, Sun S, Huang X, Zhou
D, Li T, Zhang W, Abumrad NA, Zhu X, et al: Fatty acid
2-hydroxylation inhibits tumor growth and increases sensitivity to
cisplatin in gastric cancer. EBioMedicine. 41:256–267.
2019.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Luo Y, Zha L, Luo L, Chen X, Zhang Q, Gao
C, Zhuang X, Yuan S and Qiao T: [6]-Gingerol enhances the cisplatin
sensitivity of gastric cancer cells through inhibition of
proliferation and invasion via PI3K/AKT signaling pathway.
Phytother Res. 33:1353–1362. 2019.PubMed/NCBI View
Article : Google Scholar
|
|
25
|
Wei X, Gao M, Ahmed Y, Gao M, Liu W, Zhang
Y, Xie X, Zhao Q, Wang H and Gu K: MicroRNA-362-5p enhances the
cisplatin sensitivity of gastric cancer cells by targeting
suppressor of zeste 12 protein. Oncol Lett. 18:1607–1616.
2019.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Yang Q, Zhu C, Zhang Y, Wang Y, Wang Y,
Zhu L, Yang X, Li J, Nie H, Jiang S, et al: Molecular analysis of
gastric cancer identifies genomic markers of drug sensitivity in
Asian gastric cancer. J Cancer. 9:2973–2980. 2018.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Liu F, Yin R, Chen X, Chen W, Qian Y, Zhao
Y, Jiang Y, Ma D, Hu T, Yu T, et al: Over-expression of miR-206
decreases the Euthyrox-resistance by targeting MAP4K3 in papillary
thyroid carcinoma. Biomed Pharmacother. 114(108605)2019.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Cantore M, Capparelli E, Berardi F,
Perrone R and Colabufo NA: Clinical pharmacokinetic and metabolism
of PET radiotracers for imaging P-glycoprotein in chemoresistant
tumor of colorectal cancer. Curr Drug Metab. 12:985–988.
2011.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Tan B, Li Y, Zhao Q, Fan L and Wang D:
ZNF139 increases multidrug resistance in gastric cancer cells by
inhibiting miR-185. Biosci Rep. 38(BSR20181023)2018.PubMed/NCBI View Article : Google Scholar
|