Introduction
Osteoarthritis (OA) is the main cause of
mobility-related disability caused by cartilage degeneration. Due
to the rising average age of the global population in the United
States, the prevalence of OA is expected to double by 2030
(1,2). OA could lead to a decreased quality of
life, the occurrence and formation of OA is because that articular
cartilage has limited regenerative capacity due to poor cell
migratory ability and low density (3). Once damaged, articular cartilage has a
limited potential to repair (4,5). The
expression levels of matrix metalloproteinases (MMPs), including
MMP9 and MMP13, in damaged articular chondrocytes are increased
(6,7). The extracellular matrix (ECM) of
articular cartilage contains collagen type 2 (Col2α) and aggrecan
(AGG) synthesized by chondrocytes (8). Imbalances in ECM function lead to
degenerative diseases, such as OA and cartilage injury (9), chondrocytes are stimulated to release
MMPs, including MMP9, MMP13 and a disintegrin and
metalloproteinase with thrombospondin motifs-5 (ADAMTS5) (10). It is well known that ADAMTS5
and MMPs serve essential roles in degradation of cartilage
ECM during the progression of OA (11).
Chondrogenesis is affected by a variety of factors,
including growth factors such as TGF-β (12) and BMP4(13) and oxygen levels (14,15).
HIF-1α in chondrocytes responds to oxygen concentration during
cartilage differentiation and formation in a hypoxic
microenvironment (16). When the
oxygen concentration is close to normal, specific proline residues
of HIF-1α are rapidly hydroxylated and degraded by a series of
proteasomes, including prolyl hydroxylase and asparaginyl
hydroxylase (17). When the oxygen
concentration is lower than the normal level, HIF-1α is stable and
binds to its counterpart HIF-1α to form a heterodimer and relevant
genes related to glucose metabolism, collagen production,
angiogenesis and differentiation are activated and expressed
(18,19). For chondrocytes, the expression of
SOX9, Col2α and AGG can be activated and upregulated by
stabilization and accumulation of HIF-1α (16).
Baicalin (BA) is a flavonoid compound extracted from
the roots and stems of Scutellaria baicalensis Georgi and
has a number of therapeutic effects, including antibacterial,
diuretic, anti-inflammatory, anti-metamorphosis and antispasmodic
effects, similar to those of other natural Chinese medicines, such
as icariin and Salvianolic acid (20,21).
According to a report by Chen et al (21), flavonoids including BA are effective
in the treatment of OA. Another study demonstrated that BA may
maintain the phenotype of chondrocytes isolated from New Zealand
rabbits (22). Previous studies
have also demonstrated that BA inhibits the expression of
inflammatory factors in articular chondrocytes by blocking the
NF-κB pathway (23). It has also
been confirmed that Bone Gla Protein (BGP, osteocalcin) promotes
cartilage formation and glucose metabolism by activating HIF-1α
(24). Xing et al (23) concluded that BA protects
chondrocytes by inhibiting NF-κB signaling pathways (23). Moreover, it was reported that BA
significantly inhibited the oxidative stress and decreased cell
apoptosis (25). The HIF-1α pathway
generally involves chondrogenesis and cartilage formation of
chondrocytes (16). BAY-87-2243 is
a potent and selective HIF-1α blocker and inhibitor (26,27),
it is suggested that BAY-87-2243 could block of HIF-1α expression
mediated IL-17 and CoCl2 in RAW 264.7 macrophage cells
(27). Although it is known that BA
has a protective effect on chondrocytes, the effect and mechanism
of BA on the HIF-1α pathway in chondrocytes has not yet been
elucidated.
Hence, the present study aimed to investigate the
expression of chondrogenic genes and ECM in chondrocytes treated
with or without BAY-87-2243 in the presence of BA. The regulatory
mechanism of BA involving the HIF-1α/SOX9 pathway was elucidated
using molecularbiology experiments and morphological analysis. The
present study provides a theoretical basis for clinical
applications of BA to treat patients with cartilage damage and
OA.
Materials and methods
Chemicals
BA was obtained from Bioruler Co., Ltd. Stock
solutions of BA were dissolved in dimethyl sulfoxide (DMSO;
Sigma-Aldrich; Merck KGaA) at room temperature and stored at -20˚C.
The final concentration of dimethyl sulfoxide used in the culture
was 0.01% (v/v), the concentration of BA working solution were 11,
22, 44, 88 and 176 µM. BAY-87-2243 was purchased from Selleck
Chemicals. Stock solutions of BAY-87-2243 were dissolved in
double-distilled water at room temperature and stored at -20˚C, the
working solution concentrations of BAY-87-2243 were 5, 10, 20 and
40 µM.
Animal experiments, cell culture and
cytotoxicity analysis
Three to four weeks-old C57BL/6 male mice (13-20 g)
were purchased from Medical Laboratory Animal Center (Guangzhou,
China). The mice were bred and housed freely in a specific
pathogen-free condition at a temperature of 18-22˚C with a relative
humidity of 50-60% on a 12 h light-dark cycle, with free access to
water and food. The mice were kept for five days prior to
chondrocytes isolation. Chondrocytes were isolated from the mice
and digested according to a previously described protocol (28-30).
Briefly, the mice (n=30) were sacrificed using carbon dioxide gas
(28% chamber volume per min) under general anesthesia; chondrocytes
were isolated immediately after the mice were euthanized. The knee
cartilage was digested with collagenase I (Cat. no. C0130;
Sigma-Aldrich; Merck KGaA) and collagenase D (cat. no. 11088858001,
Roche Diagnostics) for 30 min at 37˚C. The isolated chondrocytes
were seeded at a concentration of 5x105 cells/ml onto a
10-cm diameter Petri dishes containing complete Dulbecco's Modified
Eagle's Medium (DMEM; Gibco; Thermo Fisher Scientific, Inc.)
containing 10% (v/v) fetal bovine serum (FBS; Gibco; Thermo Fisher
Scientific, Inc.), 2 mM/l glutamine, 100 U/ml penicillin and 100
µg/ml streptomycin in a humidified atmosphere with 5%
CO2 at 37˚C, after three days of culture, the cell
confluence reached 80~90% and the harvested chondrocytes were used
for subsequent experiments. For the
[3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium]
(MTS) assay, ~4x103 chondrocytes were transferred to
96-well plates to make final volumes of 0.2 ml/well. For
cytotoxicity analysis of BA, the medium was removed and the new
medium containing gradient concentrations of BA (11, 22, 44, 88 and
176 µM) was added on the next day, the new medium of the control
groups contained no BA, the chondrocytes were treated with
different concentrations of BA respectively for 12, 24 and 48 h.
For cytotoxicity analysis of BAY-87-2243, the medium was removed
and the new medium containing gradient concentrations of
BAY-87-2243 (5, 10, 20 and 40 µM) was added on the next day, the
new medium of control groups contained no BAY-87-2243, the
chondrocytes were treated with different concentrations of BA
respectively for 8, 12 and 24 h. The chondrocytes in all groups
were incubated at 37˚C in a 5% CO2 container. Absorbance
was determined at 490 nm.
Alcian blue and Safranin O (SO)
staining of cultured cells
Chondrocytes were seeded in a 12-well plate at a
density of 1x105 cells per well in a final volume of 10
µl. After 4 h of incubation for adherence of chondrocytes, 2 ml of
complete DMEM medium containing various concentrations of BA (0,
11, 22, 44 and 176 µM) was added. After 14 days, cell masses were
washed and fixed for 20 min with 0.5 ml of 4.0% paraformaldehyde at
room temperature and then stained with 0.1% (w/v) Safranin O
(Sigma-Aldrich; Merck KGaA) for 30 min at room temperature and 1%
(w/v) Alcian blue (Sigma-Aldrich; Merck KGaA) for 1 h at room
temperature. Photographs of stained cells were captured using an
optical microscope (magnification, x4) (Microphot; Nikon
Corporation) and analyzed using Image-Pro Plus 6.0 software (Media
Cybernetics, Inc.).
Immunofluorescence
Chondrocytes at a concentration of
4x104/well were seeded on round glass coverslips placed
in 6-well plates and chondrocytes were exposed to 44 µM BA for 2 h
followed by 10 µM BAY-87-2243 for 24 h. The cells on the coverslips
were fixed for 15 min with 2 ml of 4.0% paraformaldehyde at room
temperature, the sections were blocked with 5% (v/v) bovine serum
albumin (BSA) for 1 h at room temperature and incubated with the
primary antibody against HIF-1α (1:50; cat. no. 36169; Cell
Signaling Technology, Inc.) at 4˚C overnight. Subsequently, the
secondary fluorescein-conjugated goat anti-rabbit antibody (1:200;
cat. no. ZF0311; OriGene Technologies, Inc.) was added and
incubated for 1 h at room temperature and the cover slips were
sealed. Images of stained cells were viewed and captured using
confocal laser scanning microscopy (CLSM; Zeiss LSM 510 META
System; magnification, x400) and compared to untreated cells.
Western blotting
Chondrocytes were seeded at a concentration of
2x105/well in 6-well plates, the chondrocytes were
pretreated with BA at 44 µM for 2 h and the chondrocytes were then
treated or not treated with 10 µM BAY-87-2243 for 24 h in the
presence of BA. The chondrocytes in the all groups were incubated
at 37˚C in an incubator with 5% CO2. The cells were
harvested using radioimmunoprecipitation assay buffer (cat. no.
P0013D, Beyotime Institute of Biotechnology). Subsequently, 100 µl
cell-lysate supernatants containing 1 mM PMSF (Sigma-Aldrich; Merck
KGaA) were analyzed using the bicinchoninic acid (BCA) protein
quantitation kit. In total, 30 µg of protein from each sample were
separated on a 10% SDS-PAGE gel. The separated proteins were
transferred onto polyvinylidene difluoride membranes (Bio-Rad
Laboratories, Inc.). The membranes were soaked overnight at 4˚C
with solutions of primary rabbit anti-GAPDH antibody (1:1,000; cat.
no. 5174), rabbit anti-HIF-1α (1:1,000; cat. no. 36169), and rabbit
anti-SOX9 antibodies (1:1,000; cat. no. 82630; all Cell Signaling
Technology, Inc.). The next day, the membranes were washed 3 times
with PBS containing 0.1% Tween-20 (TBS-T). Diluted HRP-labeled goat
anti-rabbit IgG (1:3,000; cat. no. ARG 65351; Arigo
Biolaboratories) was added and left for 1 h at room temperature.
GAPDH was used as the loading control. Images of the
stained-protein bands were recorded using an ECL Western Blotting
Substrate (Thermo Fisher Scientific, Inc.) kit and quantified using
Image lab system version 2.0. (Bio-Rad Laboratories, Inc.).
Reverse-transcription quantitative
PCR(RT-qPCR)
In total, ~2x105 chondrocytes were
transferred to 6-well plates and cultured at 37˚C with 5%
CO2. The chondrocytes in control and treatments were
pretreated with BA at 44 µM for 2 h and the chondrocytes were then
stimulated or not stimulated with 10 µM BAY-87-2243 for 24 h in the
presence of BA. Total RNA (Ribonucleic Acid) was extracted from
chondrocytes using TRIzol reagent (Thermo Fisher Scientific, Inc.)
and converted to cDNA using PrimeScript RT Master Mix (Takara Bio,
Inc.). RT-qPCR was performed with SYBR Premix ExTaq (Takara Bio,
Inc.) using the qTOWER version 3.0 PCR system (Jena Industries,
Inc.). The thermocycling conditions were as follows: At 95˚C for 2
min for pre-denaturation; 40 cycles of denaturation at 95˚C for 34
sec, and annealing and extension at 55˚C for 5 sec. The forward and
reverse primers for all the target genes are presented in Table I. The expression of all genes was
calculated by the 2-ΔΔCq method using β-actin as the
control (29).
 | Table ISequences of primers used for gene
amplification by reverse-transcription quantitative PCR. |
Table I
Sequences of primers used for gene
amplification by reverse-transcription quantitative PCR.
| Genes | Forward primer
sequence (5'-3') | Reverse primer
sequence (5'-3') |
|---|
| β-actin |
ATTGTGCACCGCAAATGCTT |
ACCACAGCACGATTGTCGAT |
| HIF-1α |
CCACCACAACTGCCACCACTG |
TGCCACTGTATGCTGATGCCTTAG |
| SOX9 |
TCAACGGCTCCAGCAAGAACAAG |
CTCCGCCTCCTCCACGAAGG |
| Col2α |
GGTCCTCCTGGTCCTGGCATC |
CGTGCTGTCTCAAGGTACTGTCTG |
| AGG |
CGTTGCAGACCAGGAGCAAT |
CTCGGTCATGAAAGTGGCGG |
| MMP9 |
AGAGACCACCACCACCACCAC |
TGCCTGCCTCCACTCCTTCC |
| MMP13 |
CTACCATCCTGCGACTCTTGCG |
CCACATCAGGCACTCCACATCTTG |
| ADAMTS5 |
AAGAGGAGGAGGAGGAGGAGGAG |
AATGGTTGTGAGCTGCCGTATGG |
| PHD1 |
AGGCTATGTCCGTCACGTTG |
TGGGCTTTGCCTTCTGGAAA |
| PHD2 |
ATATTGTGCCTTGCATGCGG |
TGGCTCACTAGTTGCCCATC |
| PHD3 |
AGGCAATGGTGGCTTGCTAT |
GACCCCTCCGTGTAACTTGG |
Statistical analysis
Results from 3 experimental repeats are presented as
the mean ± standard deviation. Comparisons were analyzed using
analysis of variance (one-way ANOVA) followed by the post hoc
Fisher's least significant difference or Bonferroni's tests using
SPSS version 22 software (IBM Corp.). *P<0.05 was
considered to indicate a significant difference and
**P<0.01 was considered to indicate a highly
significant difference.
Results
Potential cytotoxicity of BA and
BAY-87-2243 on chondrocytes was evaluated by MTS
BA is the most abundant and active component in
Scutellaria baicalensis Georgi, BA has a chemical formula
C21H18O11 (Fig. 1A) with molecular mass 446.361 g/mol
(30). The effect of BA on
chondrocytes was firstly determined by the MTS method. Chondrocytes
were subjected to the indicated BA concentration of 0, 11, 22, 44,
88, and 176 µM for 12, 24 and 48 h. As demonstrated in Fig. 1B, BA at 11 and 22 µM had no
cytotoxicity on chondrocytes; BA at 44 and 88 µM exhibited little
cytotoxicity on chondrocytes and BA at 176 µM exerted a significant
cytotoxicity effect on chondrocytes. This trend of the effect of BA
on chondrocytes remained almost consistent at 12, 24 and 48 h
(Fig. 1B). BAY-87-2243, a potent
and selective HIF-1α inhibitor (26,27),
BAY-87-2243 at higher concentrations (10, 20 and 40 µM) had no
effect on chondrocytes at the aforementioned 3 time points;
however, BAY-87-2243 at a concentration of 10 µM had no
cytotoxicity on chondrocytes cultured for 8, 6 and 24 h (Fig. 1C). The present study was designed to
investigate the effect of BA on the ECM synthesis of chondrocytes
instead of cell proliferation, hence, 44 µM BA and 10 µM
BAY-87-2243 were chosen for further experimentation.
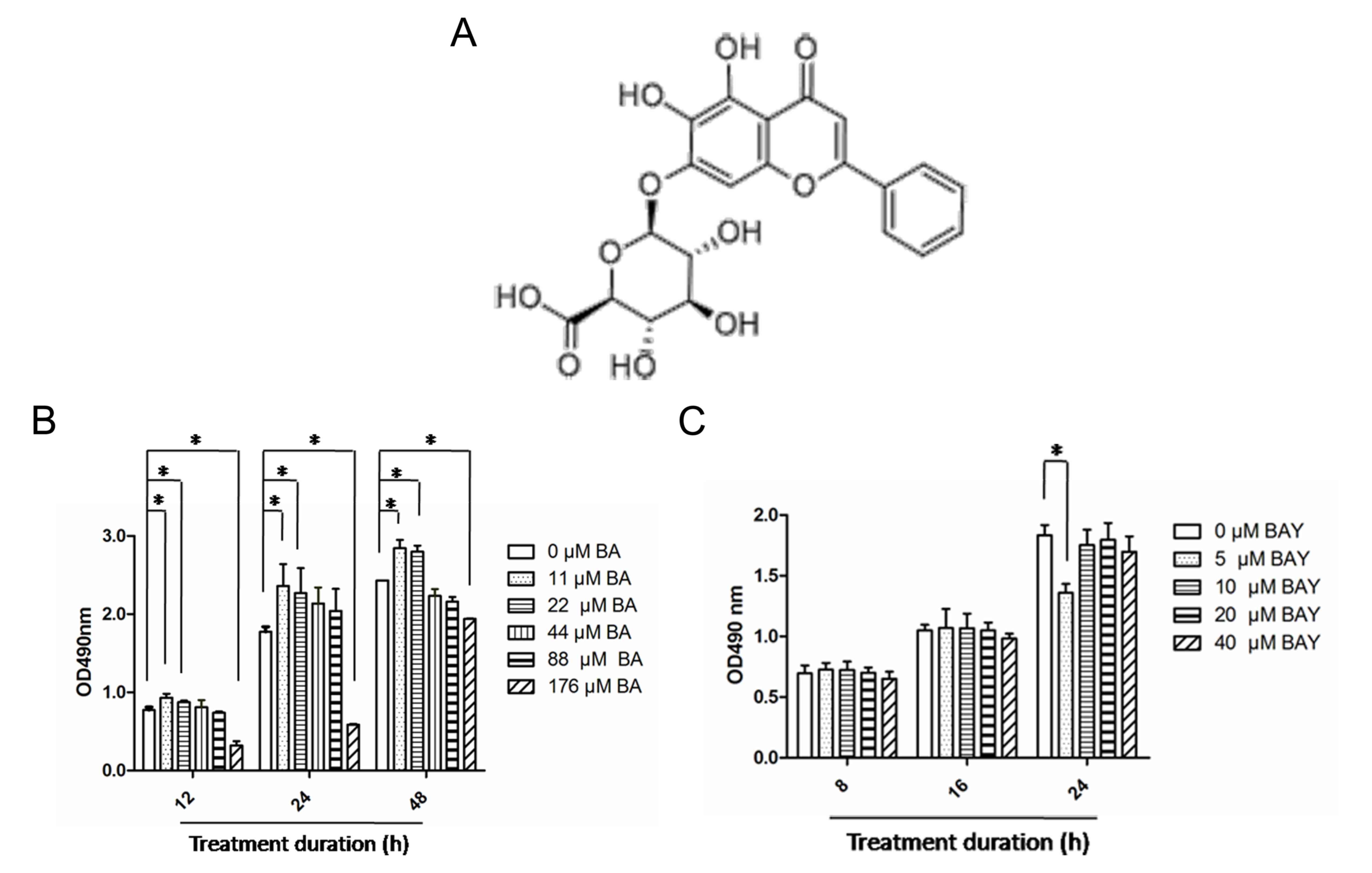 | Figure 1Effects of BA and BAY-87-2243 on
chondrocytes. (A) Molecular structure of BA. (B) Chondrocytes were
treated with BA (0, 11, 22, 44, 88 and 176 µM) for 12, 24 and 48 h
and cytotoxicity was assessed using the MTS assay. (C) Chondrocytes
were treated for 8, 16 and 24 h with BAY (5, 10, 20 and 40 µM) and
the cytotoxicity was determine by MTS assay. *P<0.05.
BA, baicalin; BAY, hypoxia inducible factor-1α inhibitor
(BAY-87-2243); MTS,
[3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium],
comparisons were analyzed using one-way ANOVA followed by
Bonferroni's tests. |
BA promotes the expression of SOX9,
AGG and Col2α and ECM synthesis
SO and Alcian blue staining are classic methods for
detecting the ECM component AGG (16), SO and Alcian blue staining indicated
that BA (at 22, 44 and 88 µM) promoted the secretion of AGG in
chondrocytes at varying degrees. Compared with untreated
chondrocytes, AGG secretion from chondrocytes in BA (44 µM) treated
groups was significantly increased (Fig. 2A). In the present study, the protein
level of SOX9 was significantly increased in the BA treatment
groups compared with that of the control group (P<0.01) and the
protein level of SOX9 was inhibited significantly by the
addition of 10 µM of BAY-87-2243 (P<0.01; Fig. 2B and C). The mRNA expression of SOX9, AGG
and Col2α was significantly upregulated following BA
treatment and the effect of BA on chondrocytes were obviously
eliminated by BAY-87-2243 (P<0.01; Fig. 2D-F). These results indicated that
ECM synthesis and chondrogenic expressionin chondrocytes were
upregulated by BA and downregulated by BAY-87-2243.
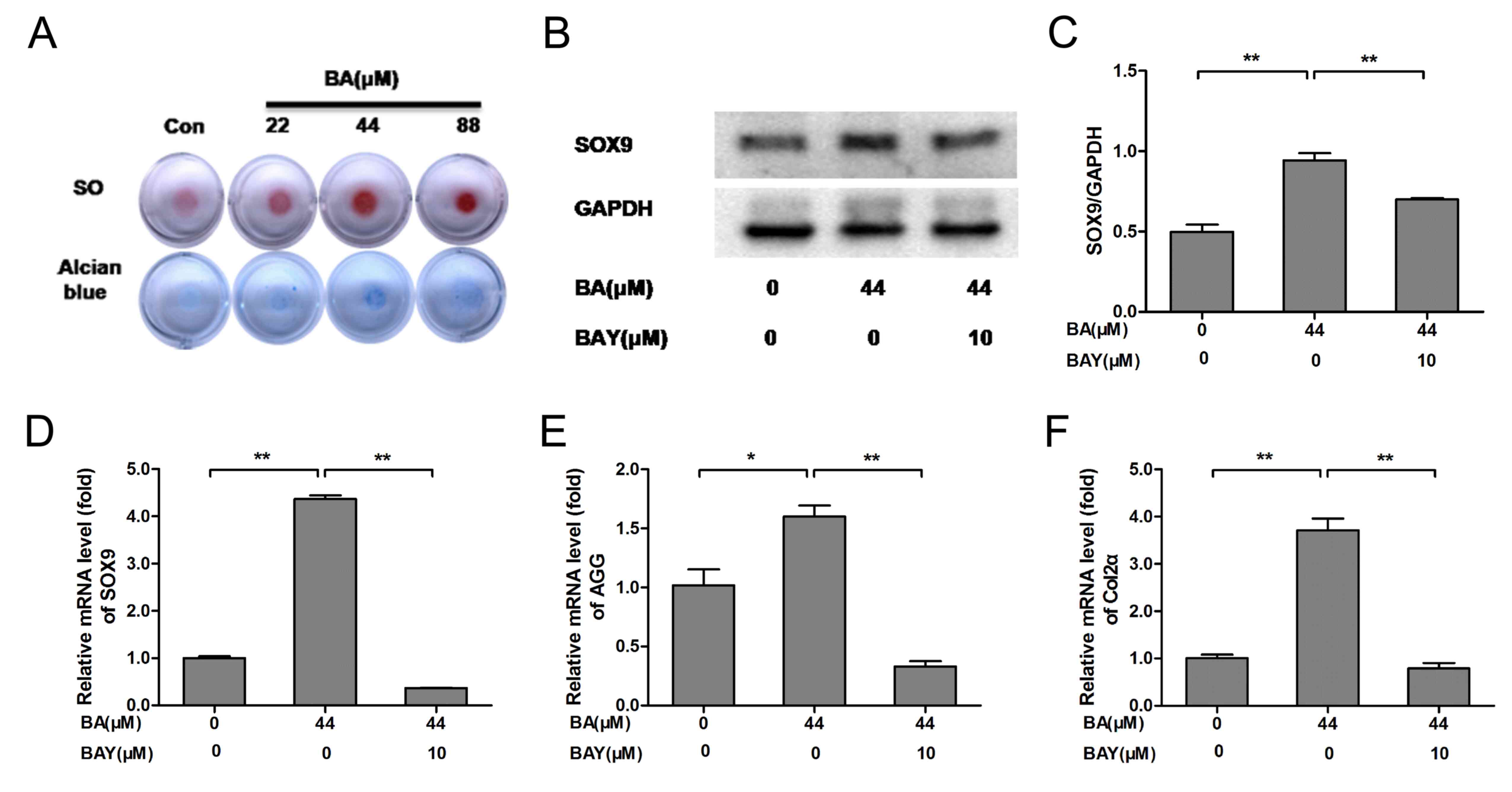 | Figure 2BA enhances extracellular matrix
synthesis and chondrogenic marker expression, while the effect was
decreased by the HIF-1α inhibitor BAY(BAY-87-2243). (A) The
chondrocyte masses treated by gradient concentration of BA (0, 22,
44 and 88 µM) were stained by SO and Alcian blue staining (4x).
(B-D) Protein and mRNA expression of SOX9 was analyzed by western
blotting and RT-qPCR, respectively and quantified using
histograms). (E) AGG and (F) Col2α mRNA expression was analyzed by
RT-qPCR and β-actin was used as an internal control for RT-qPCR.
*P<0.05, **P<0.01. HIF-1α,
hypoxia-inducible factor-1α; BA, baicalin; BAY, hypoxia inducible
factor-1α inhibitor (BAY-87-2243); SO, Safranin O; RT-qPCR,
reverse-transcription quantitative; AGG, aggrecan; Col2α, collagen
type 2; con, control. |
BA activates HIF-1α expression in
chondrocytes compared with the untreated chondrocytes
HIF-1α (green) nuclei expression was increased in
chondrocytes treated with BA for 24 h, compared with the BA
(only)-treated chondrocytes. HIF-1α (green) nuclei expression was
decreased significantly in chondrocytes treated by BAY-87-2243 in
the presence of BA (Fig. 2A).
Simultaneously, BA stimulated an upregulation in HIF-1α mRNA
(P<0.01; Fig. 3B) and HIF-1α
protein expression (P<0.01; Fig.
3C and D) in the chondrocytes
in accordance with the observed upregulation of HIF-1α
stabilization and nuclear translocation in immunofluorescence
experiments. The expression level of HIF-1α in chondrocytes treated
with BA was decreased significantly by BAY (Fig. 3). These results indicated that BA
could activate the nuclear expression of HIF-1α.
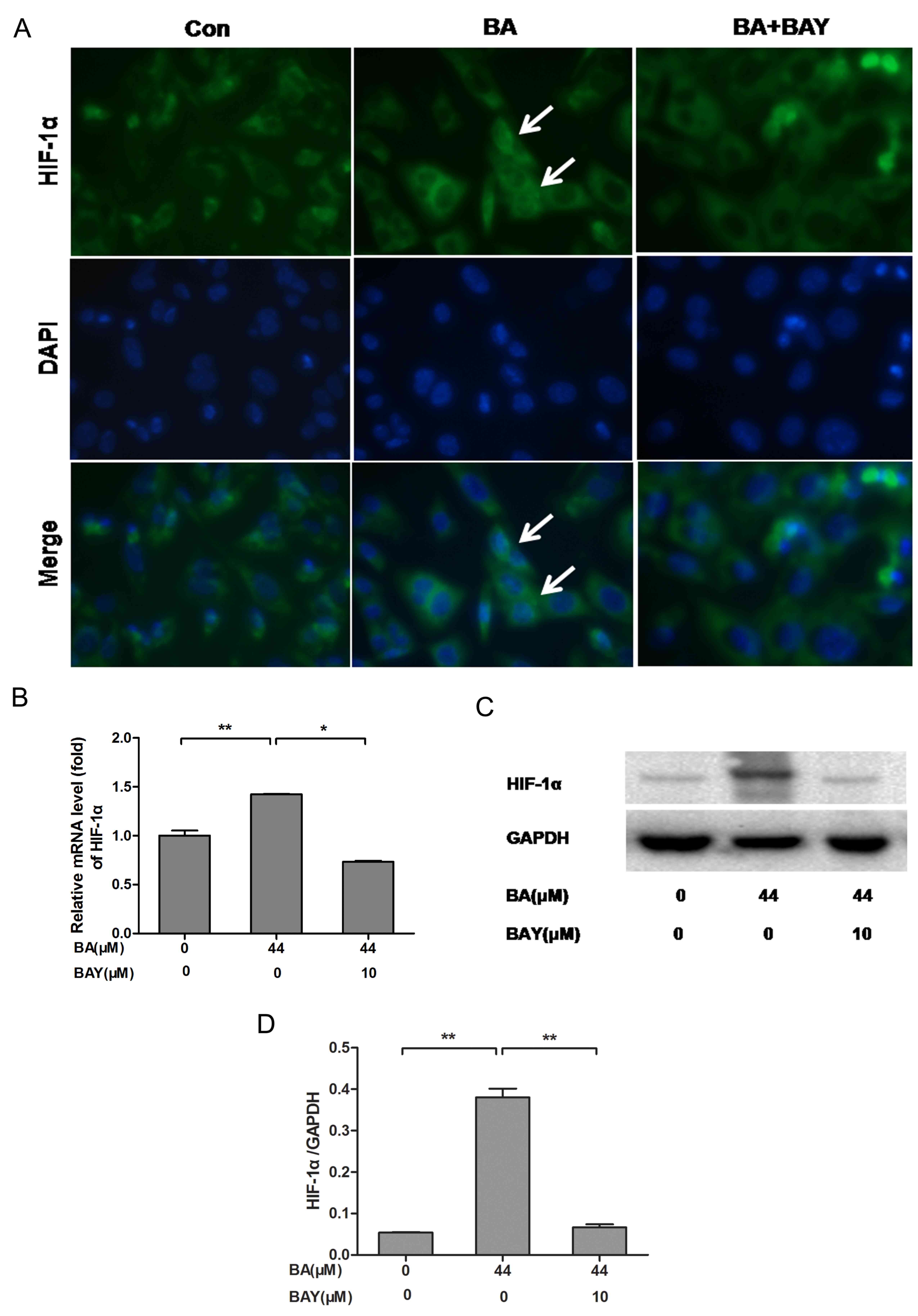 | Figure 3BA promotes the expression of HIF-1α
in newborn-mouse cartilage chondrocytes and this effect was
reversed by BAY. (A) HIF-1α nuclear localization in BA (44
µM)-treated chondrocytes with or without BAY (10 µM) was observed
under a fluorescence microscope (magnification, 400x), the white
arrows indicate chondrocytes with strong nuclear HIF-1α expression.
(B-D) mRNA and protein levels of HIF-1α measured by RT-qPCR and
western blotting, respectively. β-actin was the control for RT-qPCR
and GAPDH was the control for western blotting.
*P<0.05, **P<0.01. HIF-1α,
hypoxia-inducible factor-1α; BA, baicalin; BAY, hypoxia inducible
factor-1α inhibitor (BAY-87-2243); RT-qPCR, reverse-transcription
quantitative; con, control; BAY, hypoxia inducible factor-1α
inhibitor. |
BA downregulates MMP9, MMP13, ADAMTS5
and prolyl hydroxylases (PHD) genes expression
MMPs (especially MMP9 and MMP13) and ADAMTS5 are
regarded as the key enzymes in the progression of OA that involves
the breakdown of the ECM (31).
Compared with respective control groups, the mRNA expression of
ADAMTS5, MMP9, MMP13, PHD1, PHD2 and PHD3 in chondrocytes of
BA-treated groups were significantly downregulated (P<0.01;
Fig. 4A-F). Compared with the
BA-treated chondrocytes, the mRNA expression of ADAMTS5, MMP9,
MMP13, PHD1, PHD2 and PHD3 were increased significantly
in chondrocytes treated by BAY-87-2243 in the presence of BA
(Fig. 4A-F). These results
indicated that BA exerted a protective effect on chondrocytes by
inhibiting the expression of catabolic genes.
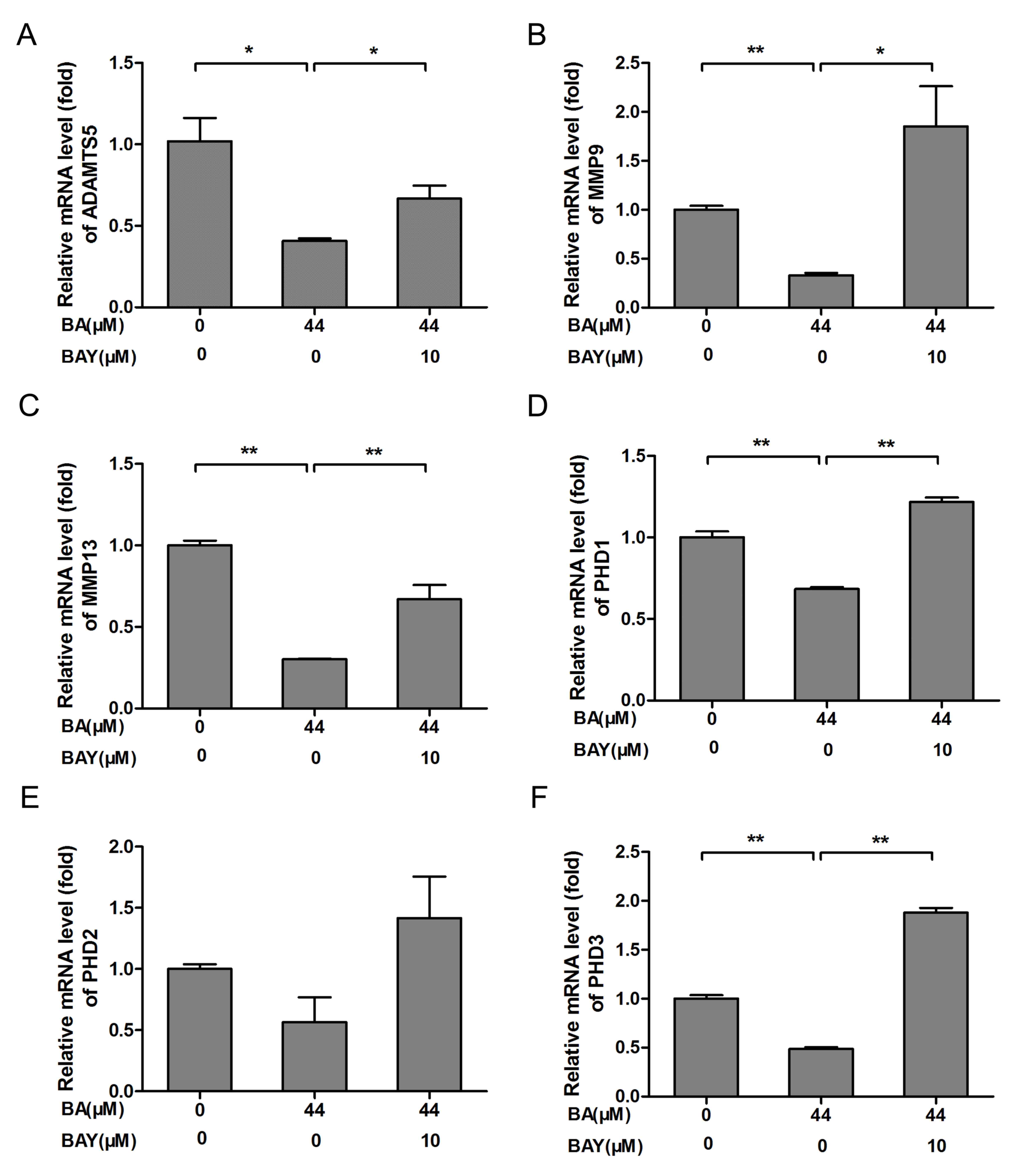 | Figure 4BA treatment decreased chondrogenic
catabolic genes and PHD expression, the effect was blocked
by the HIF-1α inhibitor BAY. Chondrocytes originating from newborn
mice were treated with or without 10 µM BAY in the presence of BA
(44 µM) for 24 h. (A) ADAMTS5, (B) MMP9 and (C)
MMP13 mRNA expression was detected by RT-qPCR. (D)
PHD1, (E) PHD2 and (F) PHD3 mRNA expression
was detected by RT-qPCR, β-actin was the control for RT-qPCR.
*P<0.05, **P<0.01. HIF-1α,
hypoxia-inducible factor-1α; BA, baicalin; BAY, hypoxia inducible
factor-1α inhibitor (BAY-87-2243); RT-qPCR, reverse-transcription
quantitative; con, control; MMP, matrix metalloproteinase; ADAMTS5,
a disintegrin and metalloproteinase with thrombospondin motifs 5;
PHD, prolyl hydroxylase. |
Discussion
In the present study, it was found that BA promoted
the synthesis of ECM in chondrocytes. In addition, it was found
that the effect of BA on chondrocytes was dependent on the
activation and expression of HIF-1α by BA. The addition of the
HIF-1α inhibitor BAY-87-2243 significantly inhibited the expression
and activation of HIF-1α. The findings of the present study
indicated that BA may be a candidate drug to treat patients with
cartilage damage and OA.
As shown in Fig. 1B,
Low concentrations of BA (11 and 22 µM) had no cytotoxic effect on
chondrocyte, however, high concentrations of BA (176 µM) had
significant cytotoxic effect on chondrocytes. Notably, it has been
reported that BA at 1.25 µM increased rabbit primary chondrocyte
proliferation (32). The present
study found that primary mouse chondrocytes were less sensitive to
BA compared with rabbit chondrocytes used in the aforementioned
study. It also indicated that low concentration BA (11 and 22 µM)
promoted cell proliferation, suitable concentration of BA (11 and
22 µM) had no toxic side effects on chondrocytes, high
concentration BA (176 µM) had cytotoxic effect toxic side effects
on chondrocytes. In the present study, the effect of BA on the ECM
synthesis and the expression of HIF-1α in chondrocytes was
explored. In addition, 44 µM BA was demonstrated to have no
proliferative effect or side effects on chondrocytes and was chosen
for subsequent experiments.
In the present research, 10 µM BAY-87-2243 had no
cytotoxicity effect on chondrocytes, which was similar with the the
conclusion reported in literature (33). Therefore, 10 uM BAY-87-2243 was used
for western blotting, RT-qPCR and immunofluorescence experiments.
In the present study, the effects of BA on chondrocyte ECM
production and function were examined by SO and Alcian blue
staining and it was demonstrated that BA promoted AGG synthesis in
chondrocytes. The results of the present study demonstrated that BA
treatment promoted AGG synthesis by chondrocytes at concentrations
ranging from 22-88 µM. Specifically, BA at 44 µM had the strongest
effect on AGG synthesis and secretion. Further, the upregulation of
AGG and Col2α mRNA supported the conclusion that the
BA exerted an effect on ECM synthesis. Key catabolic genes
including ADAMTS5, MMP9, and MMP13 mRNA levels were
downregulated following BA treatment. BAY-87-2243 significantly
reversed the above effects of BA. The findings of the present study
indicated that BA inhibited the expressions of catabolic genes
including MMP9, MMP13, PHD1, PHD2 and PHD3.
In the present study, in order to explore the
mechanism of action of BA on chondrocytes, HIF-1α expression was
examined after treatment with BA and HIF-1α inhibitor BAY-87-2243.
The expression of HIF-1α and SOX9 was significantly increased as
was the mRNA expression of AGG and Col2α in chondrocytes compared
to the control cells. However, BAY-87-2243 inhibited the expression
of HIF-1α and SOX9 in the present study. Previous studies have
reported that HIF-1α exhibited cartilage-formation ability and can
also maintain chondrocyte phenotype by regulating SOX9 expression
(34-36),
Using a HIF-1α deletion model, our previous study indicated that
icariin (ICA) increased ECM expression by activating HIF-1α
(37), the conclusion of the
present study was consistent with those studies reported
previously.
In the present study, PHD1, PHD2 and
PHD3 mRNA expression significantly decreased following BA
treatment. HIF-1α is usually induced by hypoxia, in addition, it
has also been suggested that certain flavonoids sequester iron ions
that are cofactors required for PHD activity, hence leading to the
inactivation of PHDs (38). Once
PHDs are inactivated, HIF-1α enters the nucleus and then activates
the expression of the downstream genes including SOX9 and
AGG (39). It was documented
that some flavonoids (such as quercetin, galangin and icariin)
chelated cellular iron ions, a required cofactor for PHD activity,
leading to the inhibition of PHD-catalyzed HIF prolyl hydroxylation
and subsequently HIF-1α/2α accumulation (38-40).
Thus, it was determined whether BA induced the accumulation of
HIF-1α in chondrocytes through the similar mechanism as other
flavonoids. It was revealed that the protein expression of PHD1,
PHD2 and PHD3 in chondrocytes was also reduced by BA treatment. It
is purported that protein levels and activity of the
hypoxia-inducible transcription factors HIF-1α is controlled by
hydroxylation of the regulatory α chains. Proline hydroxylases
(PHDs) target the protein for degradation via the
von-Hippel-Lindau-ubiquitin-ligase complex, and asparagine
hydroxylation by Factor Inhibiting HIF leads to transcriptional
inactivation (41-43).
The present study only evaluated inhibition and accumulation of
HIF-1α in chondrocytes treated by BA; however, the method by which
BA regulates PHD expression and whether the reduction in PHD
expression also contributes to the HIF-1α accumulation and has
other biological significance in chondrocytes, need to be further
studied.
The present study had some limitations. Firstly, the
effectof BA on chondrocytes was not studied by performing chelation
experiments. This can be the focus of research for further studies.
Secondly, there was no in vivo experiments performed in the
present study. Future studies are required for in vivo
verification of the findings of the present study. Further studies
are also required to investigatethe pharmacological mechanisms of
BA effects on chondrocytes.
In summary, it was confirmed that BA exerted a
protective effect on mouse chondrocytes in vitro. In
addition, it was concluded that BA promoted ECM synthesis at least
partly by activating the HIF-1α/SOX9 pathway. The findings of the
present study may lead to BA being developed as a potential drug to
treat patients with OA.
Acknowledgements
Not applicable.
Funding
This study was funded by grants from the Guangzhou
Municipal Health and Family Planning Commission (grant nos.
20191A011018 and 20192A010009 to PW) and Guangzhou Science and
Technology Project (grant nos. 201607010009 to SL and 201607010010
to QM).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
SL, QM and PW conceived and designed the
experiments; PW performed the majority of the experiments; QM and
RL performed the MTS assay and parts of the western blotting
experiments; PZ performed the immunofluorescence experiments and
analyzed the data; SL, QM and PW analyzed data; SL, PW and QM wrote
the manuscript. All authors have read and approved the
manuscript.
Ethics approval and consent to
participate
Animal experiments were approved (approval no.
2018-002-01) by the Ethics Committee of Guangzhou Red Cross
Hospital (Guangzhou, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Thomas AC, Hubbard-Turner T, Wikstrom EA
and Palmieri-Smith RM: Epidemiology of posttraumatic
osteoarthritis. J Athl Train. 52:491–496. 2017.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Centers for Disease Control and Prevention
(CDC): Public health and aging: projected prevalence of
self-reported arthritis or chronic joint symptoms among persons
aged >65 years--United States, 2005-2030. MMWR Morb Mortal Wkly
Rep. 52(21):489-491, 2003.
|
|
3
|
Zhou X, Tenaglio S, Esworthy T, Hann SY,
Cui H, Webster TJ, Fenniri H and Zhang LG: Three-Dimensional
printing biologically inspired DNA-based gradient scaffolds for
cartilage tissue regeneration. ACS Appl Mater Interfaces.
12:33219–33228. 2020.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Frenkel SR, Clancy RM, Ricci JL, Di Cesare
PE, Rediske JJ and Abramson SB: Effects of nitric oxide on
chondrocyte migration E, adhesion, and cytoskeletal assembly.
Arthritis Rheum. 39:1905–1912. 2015.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Heinemeier KM, Schjerling P, Heinemeier J,
Møller MB, Krogsgaard MR, Grum-Schwensen T, Petersen MM and Kjaer
M: Radiocarbon dating reveals minimal collagen turnover in both
healthy and osteoarthritic human cartilage. Sci Transl Med.
8(346ra90)2016.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Oda K and Minata M: Drug free remission
after steroid-dependent disappearance of lymphoproliferative
disorder in rheumatoid arthritis patient treated with TNF-alpha
blockade: Case study. Springerplus. 4(41)2015.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Yammani RR, Carlson CS, Bresnick AR and
Loeser RF: Increase in production of matrix metalloproteinase 13 by
human articular chondrocytes due to stimulation with S100A4: Role
of the receptor for advanced glycation end products. Arthritis
Rheum. 54:2901–2911. 2006.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Thomas CM, Fuller CJ, Whittles CE and
Sharif M: Chondrocyte death by apoptosis is associated with
cartilage matrix degradation. Osteoarthritis Cartilage. 15:27–34.
2007.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Goldring MB, Fukuo K, Birkhead JR, Dudek E
and Sandell LJ: Transcriptional suppression by interleukin-1 and
interferon-gamma of type II collagen gene expression in human
chondrocytes. J Cell Biochem. 54:85–99. 1994.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Zheng W, Tao Z, Chen C, Zhang C, Zhang H,
Ying X and Chen H: Plumbagin prevents IL-1β-induced inflammatory
response in human osteoarthritis chondrocytes and prevents the
progression of osteoarthritis in mice. Inflammation. 40:849–860.
2017.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Xue M, McKelvey K, Shen K, Minhas N, March
L, Park SY and Jackson CJ: Endogenous MMP-9 and not MMP-2 promotes
rheumatoid synovial fibroblast survival, inflammation and cartilage
degradation. Rheumatology (Oxford). 53:2270–2279. 2014.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Alvarez J, Horton J, Sohn P and Serra R:
The perichondrium plays an important role in mediating the effects
of TGF-beta1 on endochondral bone formation. Dev Dyn. 221:311–321.
2001.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Kim BS, Kang KS and Kang SK: Soluble
factors from ASCs effectively direct control of chondrogenic fate.
Cell Prolif. 43:249–261. 2010.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Rodenas-Rochina J, Kelly DJ, Gómez
Ribelles JL and Lebourg M: Influence of oxygen levels on
chondrogenesis of porcine mesenchymal stem cells cultured in
polycaprolactone scaffolds. J Biomed Mater Res A. 105:1684–1691.
2017.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Zhang FJ, Luo W and Lei GH: Role of HIF-1α
and HIF-2α in osteoarthritis. Joint Bone Spine. 82:144–147.
2015.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Hu S, Zhang C, Ni L, Huang C, Chen D, Shi
K, Jin H, Zhang K, Li Y, Xie L, et al: Stabilization of HIF-1α
alleviates osteoarthritis via enhancing mitophagy. Cell Death Dis.
11(481)2020.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Hirota K and Semenza GL: Regulation of
hypoxia-inducible factor 1 by prolyl and asparaginyl hydroxylases.
Biochem Biophys Res Commun. 338:610–616. 2005.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Araldi E and Schipani E: Hypoxia, HIFs and
bone development. Bone. 47:190–196. 2010.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Denko NC: Hypoxia, HIF1 and glucose
metabolism in the solid tumour. Nat Rev Cancer. 8:705–713.
2008.PubMed/NCBI View
Article : Google Scholar
|
|
20
|
Chen H, Xu Y, Wang J, Zhao W and Ruan H:
Baicalin ameliorates isoproterenolinduced myocardial infarction
through iNOS, inflammation and oxidative stress in rat. Int J Clin
Exp Pathol. 8:10139–10147. 2015.PubMed/NCBI
|
|
21
|
Chen C, Zhang C, Cai L, Xie H, Hu W, Wang
T, Lu D and Chen H: Baicalin suppresses IL-1β-induced expression of
inflammatory cytokines via blocking NF-κB in human osteoarthritis
chondrocytes and shows protective effect in mice osteoarthritis
models. Int Immunopharmacol. 52:218–226. 2017.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Zhu M, Ying J, Lin C, Wang Y, Huang K,
Zhou Y and Teng H: Baicalin induces apoptotic death of human
chondrosarcoma cells through mitochondrial dysfunction and
downregulation of the PI3K/Akt/mTOR pathway. Planta Med.
85:360–369. 2019.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Xing D, Gao H, Liu Z, Zhao Y and Gong M:
Baicalin inhibits inflammatory responses to interleukin-1β
stimulation in human chondrocytes. J Interferon Cytokine Res.
37:398–405. 2017.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Idelevich A, Rais Y and Monsonego-Ornan E:
Bone Gla protein increases HIF-1alpha-dependent glucose metabolism
and induces cartilage and vascular calcification. Arterioscler
Thromb Vasc Biol. 31:e55–e71. 2011.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Pan Y, Chen D, Lu Q, Liu L, Li X and Li Z:
Baicalin prevents the apoptosis of endplate chondrocytes by
inhibiting the oxidative stress induced by H2O2. Mol Med Rep.
16:2985–2991. 2017.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Griggio V, Vitale C, Todaro M, Riganti C,
Kopecka J, Salvetti C, Bomben R, Bo MD, Magliulo D, Rossi D, et al:
HIF-1α is over-expressed in leukemic cells from TP53-disrupted
patients and is a promising therapeutic target in chronic
lymphocytic leukemia. Haematologica. 105:1042–1054. 2020.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Samarpita S, Doss HM, Ganesan R and Rasool
M: Interleukin 17 under hypoxia mimetic condition augments
osteoclast mediated bone erosion and expression of HIF-1α and
MMP-9. Cell Immunol. 332:39–50. 2018.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Gosset M, Berenbaum F, Thirion S and
Jacques C: Primary culture and phenotyping of murine chondrocytes.
Nat Protoc. 3:1253–1260. 2008.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Chen C, Zhang C, Cai L, Xie H, Hu W, Wang
T, Lu D and Chen H: Baicalin suppresses IL-1β-induced expression of
inflammatory cytokines via blocking NF-κB in human osteoarthritis
chondrocytes and shows protective effect in mice osteoarthritis
models. Int Immunopharmacol. 52:218–226. 2017.PubMed/NCBI View Article : Google Scholar
|
|
31
|
He Y, Moqbel SAA, Xu L, Ran J, Ma C, Xu K,
Bao J, Jiang L, Chen W, Xiong Y and Wu L: Costunolide inhibits
matrix metalloproteinases expression and osteoarthritis via the
NF-κB and Wnt/β-catenin signaling pathways. Mol Med Rep.
20:312–322. 2019.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Huang X, Wu H, Wang L, Zheng L and Zhao J:
Protective effects of baicalin on rabbit articular chondrocytes
in vitro. Exp Ther Med. 13:1267–1274. 2017.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Ellinghaus P, Heisler I, Unterschemmann K,
Haerter M, Beck H, Greschat S, Ehrmann A, Summer H, Flamme I, Oehme
F, et al: BAY 87-2243, a highly potent and selective inhibitor of
hypoxia-induced gene activation has antitumor activities by
inhibition of mitochondrial complex I. Cancer Med. 2:611–624.
2013.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Robins JC, Akeno N, Mukherjee A, Dalal RR,
Aronow BJ, Koopman P and Clemens TL: Hypoxia induces
chondrocyte-specific gene expression in mesenchymal cells in
association with transcriptional activation of Sox9. Bone.
37:313–322. 2005.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Lefebvre V and Dvir-Ginzberg M: SOX9 and
the many facets of its regulation in the chondrocyte lineage.
Connect Tissue Res. 58:2–14. 2017.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Duval E, Baugé C, Andriamanalijaona R,
Bénateau H, Leclercq S, Dutoit S, Poulain L, Galéra P and
Boumédiene K: Molecular mechanism of hypoxia-induced chondrogenesis
and its application in in vivo cartilage tissue engineering.
Biomaterials. 33:6042–6051. 2012.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Wang P, Zhang F, He Q, Wang J, Shiu HT,
Shu Y, Tsang WP, Liang S, Zhao K and Wan C: Flavonoid compound
icariin activates hypoxia inducible factor-1α in chondrocytes and
promotes articular cartilage repair. PLoS One.
11(e0148372)2016.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Park SS, Bae I and Lee YJ:
Flavonoids-induced accumulation of hypoxia-inducible factor
(HIF)-1alpha/2alpha is mediated through chelation of iron. J Cell
Biochem. 103:1989–1998. 2008.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Leopoldini M, Russo N, Chiodo S and
Toscano M: Iron chelation by the powerful antioxidant flavonoid
quercetin. J Agric Food Chem. 54:6343–6351. 2006.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Davis CK, Jain SA, Bae ON, Majid A and
Rajanikant GK: Hypoxia mimetic agents for ischemic stroke. Front
Cell Dev Biol. 6(175)2019.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Terkhorn SP, Bohensky J, Shapiro IM,
Koyama E and Srinivas V: Expression of HIF prolyl hydroxylase
isozymes in growth plate chondrocytes: Relationship between
maturation and apoptotic sensitivity. J Cell Physiol. 210:257–265.
2007.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Wang W, Xu B, Xuan H, Ge Y, Wang Y, Wang
L, Huang J, Fu W, Michie SA and Dalman RL: Hypoxia-inducible factor
1 in clinical and experimental aortic aneurysm disease. J Vasc
Surg. 68:1538–1550.e2. 2018.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Watts ER and Walmsley SR: Inflammation and
hypoxia: HIF and PHD isoform selectivity. Trends Mol Med. 25:33–46.
2019.PubMed/NCBI View Article : Google Scholar
|














