Introduction
Cancer is a major cause of death worldwide. China is
the most populous country in the world and has been estimated to
present ~4.51 million cancer cases and 3.04 million cancer deaths
in 2020(1). Even as research into
cancer prevention and treatment increases, cancer is affecting an
increasing number of people (2).
Since cancer has common characteristics similar to numerous other
chronic diseases, including high incidence, long incubation periods
and a complex and diverse etiology, it is induced by a variety of
risk factors resulting in bodily dysfunction (3). As cancer treatments have improved,
cancer progression has been significantly delayed. Currently,
cancer has the highest global mortality rate of any chronic
disease, exceeding cardiovascular and cerebrovascular diseases and
accounting for a total mortality rate of 28.1% (4,5). CRC,
a digestive system tumor, accounts for 1/3 of all malignant tumors
(6,7). In China, with changes in the
environment and diet, as well as increasing stress and poor living
habits, the incidence of CRC is rising and the average age of
diagnosis is decreasing (8).
Depressive disorder is a common mental disorder
which mainly manifests as a persistent feeling of sadness, slow
thinking, psychomotor inhibition, difficulty concentrating and
mental and physical fatigue (9).
Causes of depression include biological, psychological and social
factors, as well as inputs from various life events, which are
closely associated to its onset (10). Clinical observations have found that
patients with malignant tumors are more likely to be affected by
depressive disorder (11). The
interaction between tumors and depression often aggravates physical
and psychological diseases in patients in a vicious circle
(12). Therefore, the role of
depressive disorder in tumor development is worth exploring.
It has been demonstrated that immune cells,
cytokines and other immune factors serve important roles in tumor
occurrence, development and metastasis (13). Immune-related pathogenesis has
become a focus of research for tumor etiology (14,15).
IL-22 serves key roles in autoimmune diseases and CRC tumor
pathogenesis and may be effective for analyzing tumor efficacy and
prognosis, as well as guiding therapies (16-18).
High expression levels of IL-22 in CRC tissue and serum may be
associated with the activation or inhibition of related signaling
pathways. Phosphorylation of ERK 1/2 by IL-22 has been observed in
CRC cells, resulting in the activation of the MAPK signaling
pathway, which regulates cell growth, development, differentiation
and migration. Following administration with ERK inhibitors, ERK1/2
phosphorylation is inhibited, showing that IL-22 activates MAPK
signaling and affects tumor occurrence and development (19,20).
Based on these previous studies, the present study
explored the therapeutic effect of antidepressants combined with
chemotherapy drugs on CRC and depression in a mouse model of CRC
with depressive disorder. The effects on cytokine IL-22 and the
MAPK signaling pathway are specifically discussed to provide a
reference for the clinical treatment of CRC tumors.
Materials and methods
Cell culture and preparation
CT-26, a strain of colonic cancer cells from BALB/C
mice, was purchased from the Shanghai Cell Bank of the Chinese
Academy of Sciences. Cells were seeded in a Petri dish at a density
of 1x106/ml and cultured with DMEM (Gibco; Thermo Fisher
Scientific, Inc.) supplemented with 10% FBS (Gibco; Thermo Fisher
Scientific, Inc.) and antibiotics (100 IU/ml penicillin and 100
µg/ml streptomycin) at 37˚C in a 5% CO2 incubator. At
the logarithmic growth phase, cells were digested with trypsin and
centrifuged at 300 x g for 5 min. Cells were collected, washed
twice with 0.9% NaCl solution and then counted and diluted into a
0.5x105/ml cell suspension.
Animals and grouping
All animal experiments were approved by the Animal
Ethics Committee of the Chinese Medical University (Shenyang,
China; approval no. CMU2018178). A total of 60 male BALB/c mice
(age, 6 weeks; weight, 25±1 g) were purchased from Beijing
Huafukang Bioscience Co., Inc. [animal production license, SCXK
(Jing) 2014-0004]. Mice were housed at 22±2˚C, 40-70% humidity and
12-h light/dark cycles with free access to food and water. Mice
were randomly divided into 6 groups of 10 mice: i) Cancer model
group (CA), mice grafted with CT-26 cells (5x105 in 200
µl DMEM) through subcutaneous injection to the right armpit; ii)
cancer depression model group (CD), in which following CT-26 cell
engraftment, depression was established by chronic mild stress from
day 1; iii) cancer depression with paroxetine group (CDP), CD model
mice were intraperitoneally injected with 10 mg/kg/day paroxetine
for 28 days from the 14th day of chronic mild stress; iv) cancer
depression with fluorouracil group (CDF), CD mice intraperitoneally
injected with 20 mg/kg/day fluorouracil for 10 days from the 14th
day of chronic stress; v) cancer depression with paroxetine and
fluorouracil group (CDP+F), CD mice intraperitoneally injected with
both 10 mg/kg/day paroxetine and 20 mg/kg/day fluorouracil; and vi)
control group (CON), mice given equal amounts of saline. Tumor
volumes were measured at 0, 7, 14, 17, 20, 23, 26, 29, 32, 35, 38
and 42 days and inhibition rate and behavioral changes were
calculated. A tumor diameter of 1.5 cm was set as the humane
endpoint. The following formula was used: Tumor inhibition rate =
(average tumor weight in CD mice-average tumor weight in treatment
group)/average tumor weight in CD x100%.
Chronic unpredictable mild stress
protocol
Chronic unpredictable mild stress was induced
according to previous literature (21,22).
Mice in the CON and CA groups were housed five per cage with normal
diet and mice in the other four groups were socially isolated.
Stress included 24 h food deprivation followed by 24 h water
deprivation, 12 h strobes, 24 h cage moisture, 24 h cage tilt at
45˚, 5 min cold swimming (at 4˚C), or overnight illumination. One
stressor was applied each day in a random order and not repeated
within 7 days, for 4 continuous weeks.
Sucrose preference experiment
Sucrose preference was tested before and after
chronic mild stress. After sucrose adaptation, all mice were
deprived of food and water for 12 h. They were then provided with a
bottle of 1% sucrose and a bottle of regular water. Following 12 h
free drinking, the locations of the sucrose bottle and the pure
water bottle were exchanged, and mice drank freely for another 12
h. To determine the baseline value of mouse sucrose preference, the
following formula was used: Sucrose preference percentage = sucrose
consumption/(sucrose consumption + water consumption) x100%. The
sucrose preference experiment was carried out every 2 weeks from
the establishment of each mouse model.
Forced swimming test
The experiment was conducted on two consecutive
days. The test was divided into two stages, an adaptation stage at
the first day and a test stage at the second day. Mice were placed
in a plexiglass cylinder containing 25˚C clean water. On the first
day, the adaptation stage, the mice were released into the water
container for 15 min and then removed, dried and returned to their
cages. On the second day, the test stage, the mice were released
into the water container and the first two min were not recorded.
All movements of the mice were recorded for the next 5 min and
their immobility time was calculated by deducting their mobility
time from the total time (5 min). To avoid animal odor
interference, the water was replaced after each mouse test
(23,24).
Immunohistochemistry
Following completion of the experiment or when tumor
size reached the humane endpoint (diameters, 1.5 cm), mice were
euthanized by cervical dislocation and tumor tissue were fixed in
10% formalin neutral buffer solution (FUJIFILM Wako Pure Chemical
Corporation) at room temperature for 24 h and embedded in paraffin.
Paraffin-embedded sections were cut to 5 µm thickness and stained
with Ki-67 antibodies (1:300; cat. no. 9449; Cell Signaling
Technology, Inc.) for 20 min at room temperature, followed by
staining with horseradish peroxidase (HRP)-labeled anti-rabbit
secondary antibodies (1:1,000; cat. no. sc-2357; Santa Cruz
Biotechnology, Inc.) for 20 min at room temperature. Slices were
observed using a light microscope to detect Ki-67 expression in
tissue. The light density values were analyzed using Image pro-plus
software 6.0.(Media Cybernetics, Inc.)
Detection of serum cytokine expression
in mice using ELISAs
IL-22 plasma levels in mice were detected by ELISA
(cat. no. ARG80218; Arigo Biolaboratories Corp.), according to the
manufacturer's protocol. After incubation at 3˚C for 1 h, plate
450-nm absorbance values were read using a microplate reader
(EXL808; BioTek Instruments, Inc.).
Western blotting
RIPA lysate containing a 10% protease and
phosphatase cocktail (cat. no. ab201119; Abcam) was added to the
tumor tissue. Samples were lysed on ice for 30 min, centrifuged at
4,000 x g for 10 min and the supernatant was collected. Protein
concentration was measured using a BCA protein detection kit. After
diluting, 30 mg protein/well was separated by 10% SDS-PAGE
electrophoresis, transferred to PVDF membranes and blocked with 5%
skim milk in PBST at room temperature for 1 h. Membranes were
independently incubated with antibodies for IL-22 (1:1,000; cat.
no. ab133545), Bcl-2 (1:1,000; cat. no. ab32124), Bax (1:1,000;
cat. no. ab32503), caspase-3 (1:1,000; cat. no. ab13847), p38
(1:1,000; cat. no. ab170099), phosphorylated (p)-p38 (1:1,000; cat.
no. ab47363), ERK (1:1,000; cat. no. ab184699), p-ERK (1:1,000;
cat. no. ab201015), JNK (1:1,000; cat. no. ab179461) or p-JNK
(1:1,000; cat. no. ab124956) and GAPDH (1:1,000; cat. no. ab8245),
then incubated at 4˚C overnight. After rinsing with TBS-0.05%
Tween-20, Goat anti-rabbit HRP antibodies were added (1:1,000; cat.
no. ab6721) and incubated for 2 h at room temperature. All
antibodies were purchased from Abcam. Membranes were developed
using the ECL luminescence kit (cat. no. 32106; Thermo Fisher
Scientific, Inc.) and imaged using a gel imaging system. Gray
values were analyzed by ImageJ software (version no. 1.49; National
Institutes of Health).
Statistical analysis
SPSS 22.0 (IBM Corp.) was used for statistical
analysis of measurements and quantification. All data were analyzed
by one-way ANOVA with Tukey's multiple comparison post hoc test.
P<0.05 was considered to indicate a statistically significant
difference. The data are presented as the mean ± standard error.
Each experiment was repeated ≥3 times.
Results
Paroxetine may improve behavior in
mice with cancer and depression
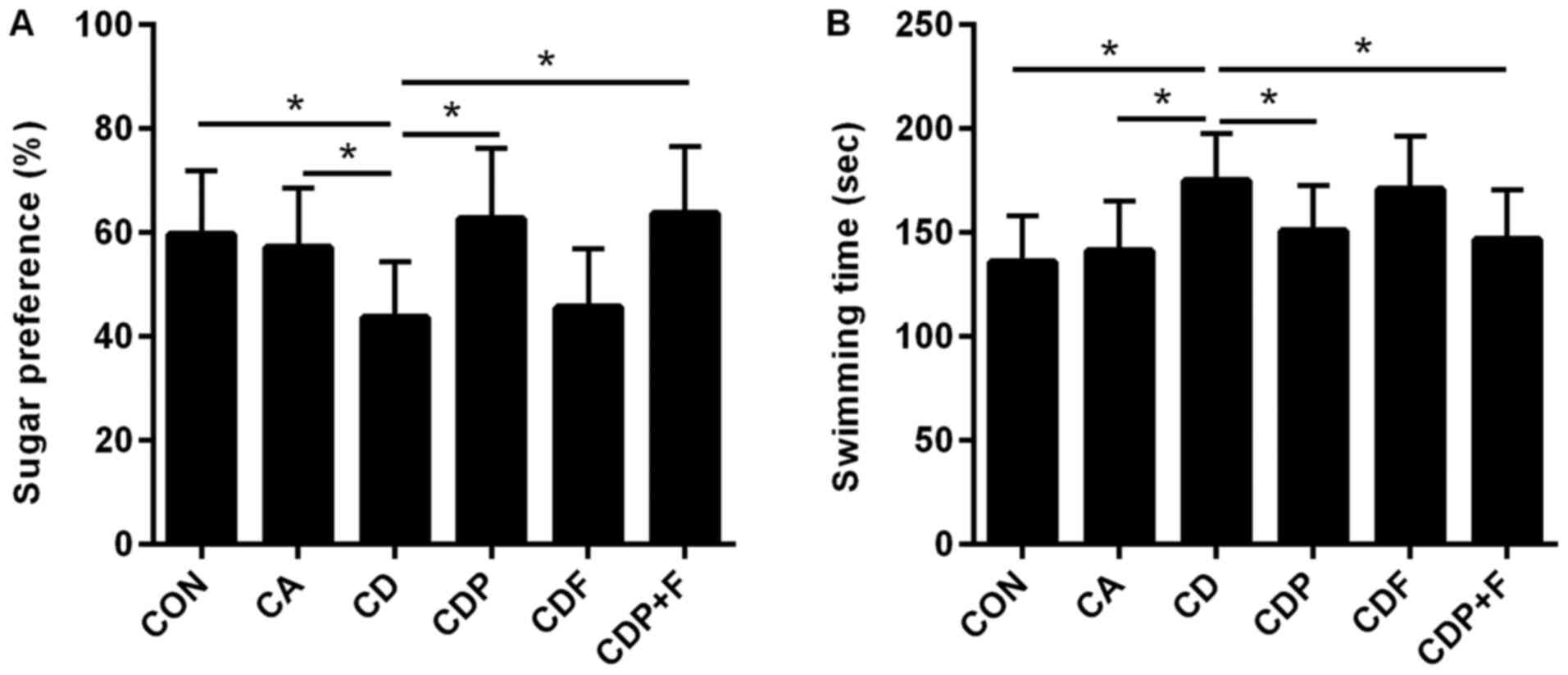
Sucrose preference tests showed a lower sucrose
preference percentage in the CD mice than in the CON or CA groups
(P<0.05). While chronic unpredictable mild stress reduced
sucrose preference values in the CD group, there was no difference
between the CA and CON groups (P>0.05). The percentage of
sucrose preference of mice in the CD group was smaller than the CDP
(P<0.05) and CDP+F groups (P<0.05). There was no significant
difference in sucrose preference between the CDF and CD groups
(P>0.05). These results suggested that paroxetine antidepressant
therapy may reverse changes in sucrose preference caused by chronic
stress and that fluorouracil has no effect on sucrose preference in
mice (Fig. 1A).
The forced swimming test showed similar results. The
swimming time in the CD group was longer than in the CON and CA
groups (P<0.05), with no difference between the CA and CON
groups (P>0.05). After paroxetine treatment, the swimming time
of mice in the CDP group was shortened (P<0.05), while
fluorouracil had no effect on swimming time in mice (P>0.05)
compared with the CD group. These results showed that paroxetine
may improve the depressive behavior of mice with cancer and
depression (Fig. 1B).
Paroxetine combined with chemotherapy
inhibits tumor growth in mice
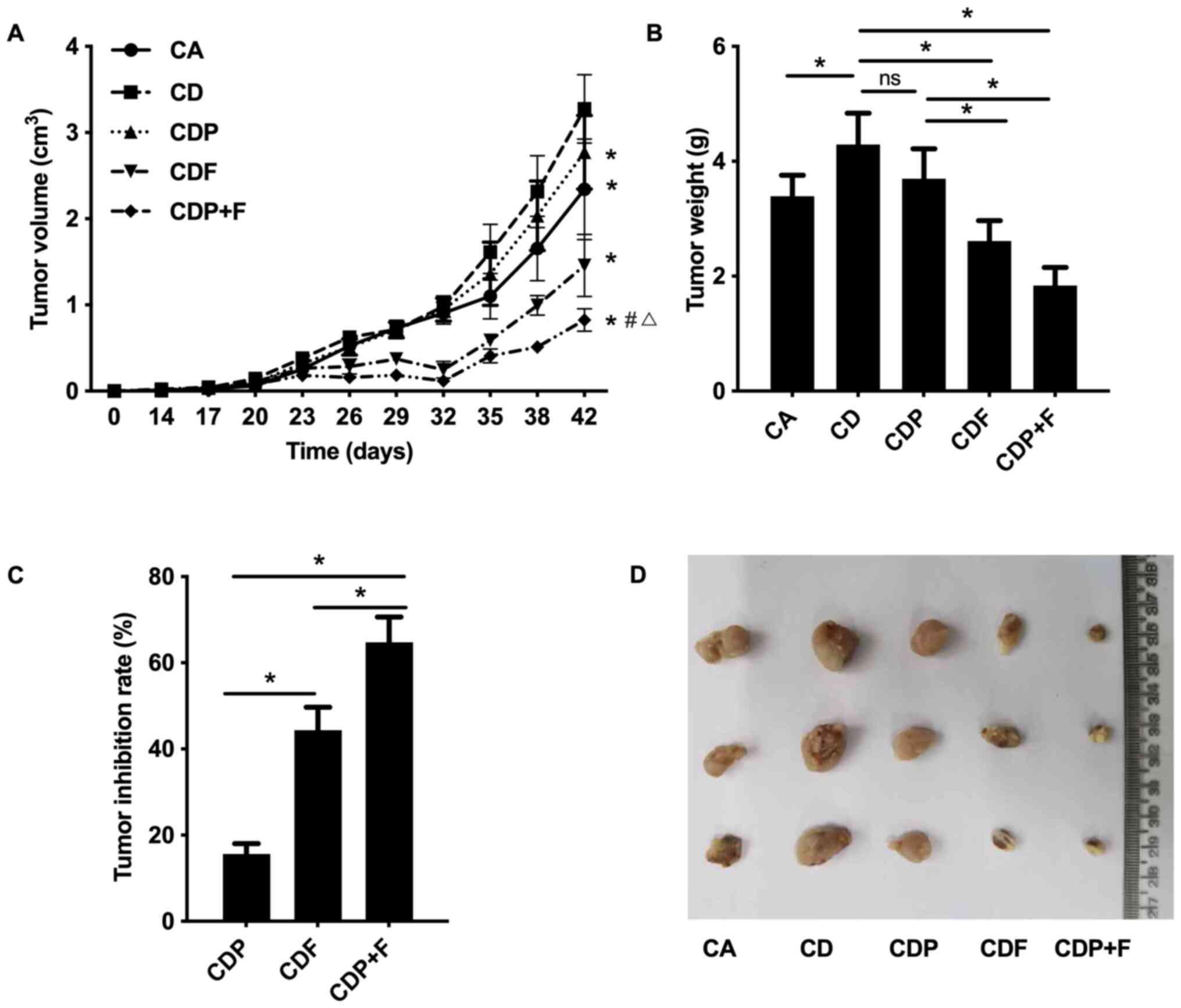
According to the dynamic tumor growth curve,
tumor-bearing mice in each group showed an overall growth trend.
Tumor volumes grew largest and fastest in the CD mice, followed by
the CDP group. In the CDF and CDP+F groups after 26 days of
inoculation (10-day drug intervention), tumors maintained slow or
even decreased growth until the 32nd day, when tumor growth rate
accelerated and tumor volumes increased in all tumor groups. Tumors
in CD mice grew faster than in CA mice (Fig. 2A and D; P<0.05), suggesting that the
depressive disorder promoted tumor growth. As compared to CD group,
tumor growth was slower in the CDF and CDP groups, and tumor growth
was the slowest in the CDP+F group (Fig. 2A and D; P<0.05). On the 42nd day, CD group
mice had the largest tumor volume and CDP+F mice had the smallest
tumor volume (Fig. 2A and D; P<0.05). As showed in Fig. 2B the difference of tumor weight in
different groups were similar to the difference of the tumor
volumes among different groups at day 42 in Fig. 2A. The CD Group had the largest tumor
weight and the CDP+F group had the smallest. Fluorouracil inhibited
tumor growth and reduced tumor weight. Furthermore, fluorouracil
combined with paroxetine was more effective; however, paroxetine
alone only demonstrated a trend to inhibit tumor growth which was
not found to be significant (P>0.05; Fig. 2A and B).
Comparing the tumor inhibition rate of mice in each
drug treatment group, the CDP+F group had the highest inhibitory
rate (P<0.05) followed by the CDF group, with the CDP group
having the lowest inhibitory rate. This demonstrated that combined
treatment with paroxetine and fluorouracil had a better inhibitory
effect on tumor growth in mice with CRC and depression (Fig. 2C).
Tumor cell proliferation is inhibited
by paroxetine combined with fluorouracil
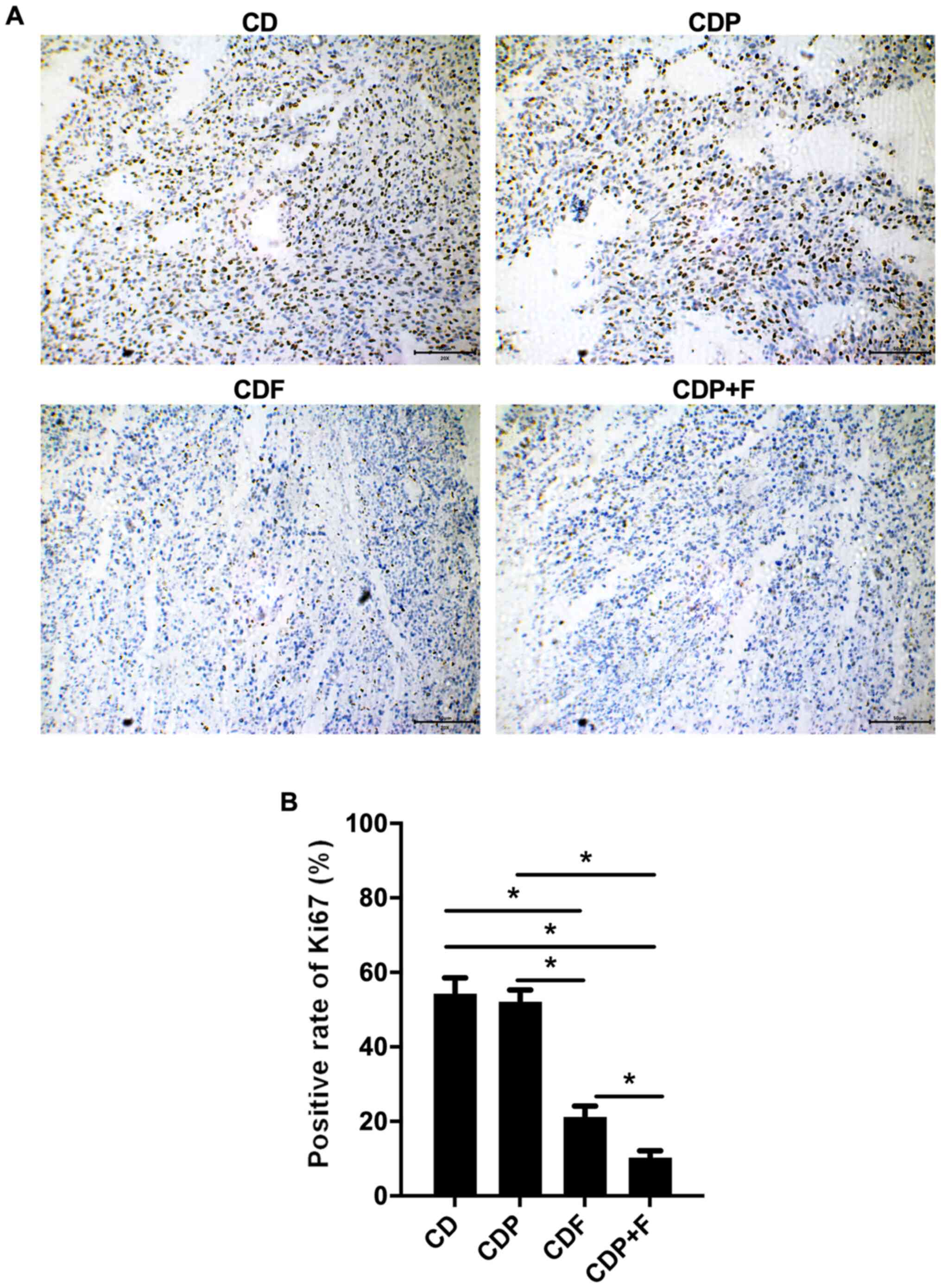
To detect the proliferation of tumor cells in
various groups, Ki-67 staining was used. Ki-67 expression in the
CDF and CDP+F groups was significantly lower than in the CD group
(P<0.05), with the lowest Ki-67 rate in the CDP+F group.
Although paroxetine alone in the CDP group was not indicated to
inhibit Ki67 expression significantly compared with the CD group
(P>0.05), the combination of paroxetine and fluorouracil
inhibited tumor cell proliferation in mice with CRC and depression
(Fig. 3).
Paroxetine combined with fluorouracil
promotes tumor cell apoptosis in mice
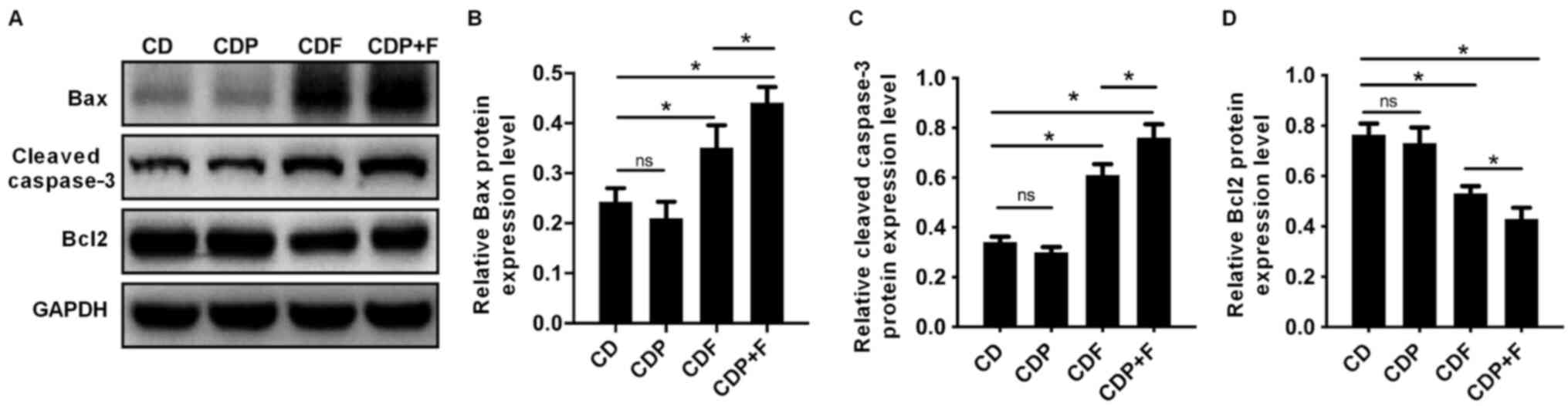
The expression levels apoptosis-related proteins
Bcl-2, Bax and cleaved-caspase-3 were detected by western blotting.
Bax and cleaved-caspase-3 expression levels significantly increased
and Bcl-2 expression significantly decreased (P<0.05) in the CDF
and CDP+F groups compared with the CD group. Paroxetine treatment
alone had no effect on Bax, Bcl-2 or cleaved-caspase-3 expression
compared with the CD group (P>0.05). Expression levels of Bax
and cleaved-caspase-3 levels were significantly increased, and
Bcl-2 levels significantly decreased in the CDP+F group compared
with the CDF group (P<0.05). Thus, combining paroxetine and
fluorouracil in CDP+F group promoted tumor cell apoptosis in mice
with CRC and depression compared to CDF (P>0.05), indicating
that paroxetine and fluorouracil exhibited synergistic effects
(Fig. 4).
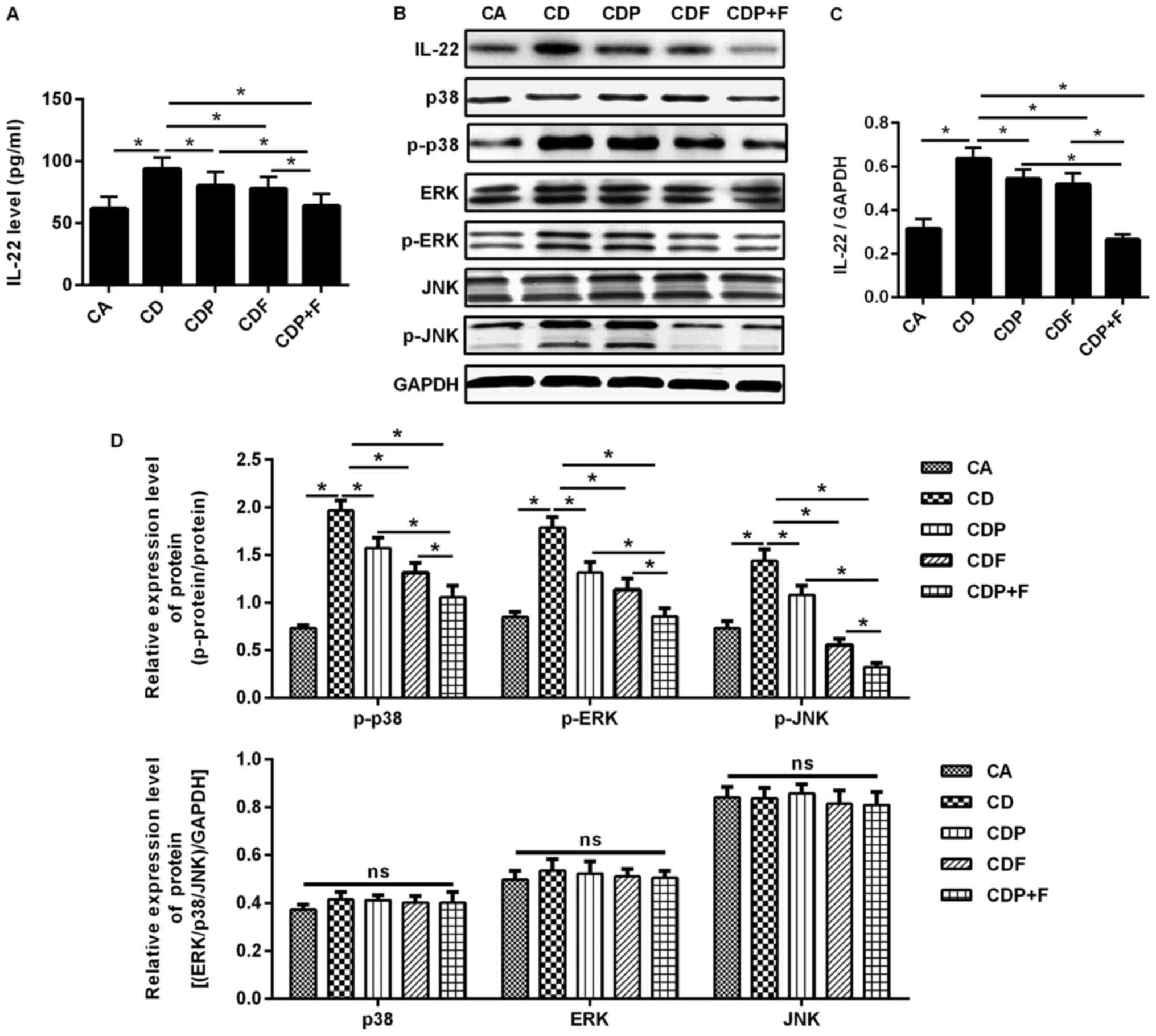
Paroxetine combined with fluorouracil
may reduce IL-22 levels
ELISAs and western blotting were used to detect
IL-22 expression levels in mouse blood and tumor tissues. The
protein levels of IL-22 in the serum and tissues of CD mice was
significantly increased compared to CA mice (P<0.05); however,
IL-22 was reduced by both paroxetine or fluorouracil, with
paroxetine combined with fluorouracil reducing IL-22 level the most
when compared to all cancer and depression groups (P<0.05;
Fig. 5A-C).
Paroxetine combined with fluorouracil
inhibits activation of the MAPK signaling pathway
To evaluate whether paroxetine combined with
fluorouracil inhibits the growth of CRC cells by inhibiting IL-22
expression, thus blocking the MAPK signaling pathway, western
blotting was used to detect the expression of MAPK signaling
pathway-related proteins p38, p-p38, ERK, p-ERK, JNK and p-JNK.
Significantly increased levels of p-p38, p-ERK and p-JNK were found
in the CD group compared to the CA group (P<0.05) and p38, ERK
and JNK were not significantly changed, indicating that the MAPK
signaling pathway was activated. Compared with the CD, CDP and CDF
group, in CDP+F group the expression of p-p38, p-ERK and p-JNK was
significantly reduced, while the expression of unphosphorylated
p38, ERK and JNK was not significantly different between groups;
furthermore, the CDP+F group was demonstrated to induce the most
pronounced reduction of p-p38, p-ERK and p-JNK expression
(P<0.05), indicating that MAPK signaling pathway was
significantly inhibited (Fig. 5B
and D).
Discussion
The incidence of psychological depression is
increasing annually. Globally, >300 million people suffer from
depression, which is listed by the World Health Organization (WHO)
as the single largest factor contributing to global disability
(25). Depressive disorder has
adverse effects on the occurrence, development and prognosis of
malignant tumors (26). In patients
with malignant tumors, depression affects immune function, thus
altering the occurrence, development and outcomes of malignant
tumors (12,27,28).
CRC has become one of the most common malignant tumors of the
digestive system (29). Of all
clinically hospitalized patients with CRC, 28.13-31.04% of patients
with breast cancer experience symptoms of depression during the
perioperative period, which a significantly higher incidence than
in the general population (30-32).
In the present study, tumors grew faster in transplanted tumor mice
with depressive disorder, with a higher tumor volume than in
transplanted tumor mice without chronic mild stress. The dynamic
curves of tumor growth demonstrated that CRC tumor volume was
associated with depression following prolonged chronic mild stress.
Fluorouracil treatment effectively inhibited CRC growth and reduced
Ki-67 expression, causing inhibited tumor cell proliferation and
promoted tumor cell apoptosis. The combination of antidepressants
and chemotherapy drugs may improve depressive behaviors and serve a
role in tumor inhibition in tumor-bearing mice with depression. All
anti-tumor effects were more obvious with combined treatments.
These results demonstrated that depressive disorder may promote
tumor development in grafted tumor mice and that combined therapy
with antidepressants and chemotherapy drugs may have synergistic
roles in improving the depressive behavior of mice, while also
inhibiting tumor progression.
Previous research has reported that mental and
social dysfunction, including depression, changes cytokine levels
in vivo, leading to decreased immune function, which may
weaken the inhibitory effects of the immune system on malignant
tumor growth (33). In a state of
depression, the hypothalamus-pituitary-adrenal axis is activated
and the secretion of cortisol hormones by the adrenal gland is
increased (34). Cortisol hormones
inhibit IL-2, IL-6, TNF and other cytokines, as well as antibodies
used by the immune system (35).
Antidepressant therapy improves depressive symptoms in patients
with malignant tumors and has positive effects on their immune
function (36,37). IL-22 is produced following IL-9
stimulation of BW5147 lymphoma cells in mice (38). IL-22 has functions similar to IL-10,
inhibiting immunity and inflammation, and exhibits 22% amino acid
homology with IL-10(39). IL-22 is
mainly produced by immune cells such as TH1, TH17 and TH22 cells
(40). Numerous chronic diseases,
including enteritis, mesenchymal lung disease and rheumatoid
arthritis, exhibit increased IL-22 expression (41-43).
Previous studies have confirmed that IL-22 can raise acute phase
reaction products in hepatocytes (20) and activate pancreatic-related
proteins in pancreatic acinar cells (44), indicating that IL-22 serves
important roles in the inflammatory response phase. Similarly,
IL-22 induces the activation of STAT in various cell lines,
including mesangial cells (45),
colon epithelial cells (46),
nonmelanoma skin cancer (47) and
hepatoma cells (20).
A previous study revealed higher IL-22 levels in
tumor tissues than in adjacent tissues in patients with
hepatocellular carcinoma, with raised IL-22 levels at more
developed tumor stages (48).
However, the changes and effects of IL-22 expression in patients
with tumors and depression are unclear. Significantly increased
IL-22 was found in CD group mice, suggesting that IL-22 is
associated with the pathogenesis of tumor depression.
Antidepressant-selective serotonin reuptake inhibitors have been
demonstrated to reduce depressive symptoms and regulate cytokine
expression. Paroxetine has been used as the antidepressant
treatment for tumor-depressive mice (49). Paroxetine may serve a therapeutic
role by reducing the levels of IL-22 in these mice, with a more
significant reduction seen by combination with fluorouracil
therapy. This indicates that paroxetine may improve immune status
by reducing IL-22 levels and enhancing the therapeutic effect of
chemotherapy drugs.
By combining with specific cell surface receptors,
cytokines have biological functions in activating cell signal
transduction pathways. MAPK is a serine threonine protein kinase in
eukaryotic cells and has signal transduction functions in multiple
pathways, mediating extracellular signaling into cells (50,51),
regulating cell growth, differentiation and migration (52). MAPK is one of the major signal
transduction pathways. MAPK family members ERK, p38 and JNK promote
cell proliferation and suppress apoptosis to promote tumor
development (53,54). Studies have demonstrated that ERK1/2
inhibiters can block the proliferation of colon cancer cells by
inducing cell cycle arrest in the S-phase (55,56).
IL-22 has been shown to induce ERK1/2 and JNK protein
phosphorylation, activate the MEK-ERK/JNK pathway, stimulate the
growth of tumor cells and promote tumor cell proliferation while
inhibiting tumor cell apoptosis, thus promoting CRC development
(57,58). In the present study, it was found
that paroxetine alone, fluorouracil alone and paroxetine combined
with fluorouracil reduced p-p38, p-ERK and p-JNK protein expression
levels. Combining the two drugs has the largest regulatory effect
on the MAPK signaling pathway in tumor depression mice.
The aforementioned results indicated that the
combination of paroxetine and fluorouracil may inhibit tumor cell
growth in mice with CRC and depression by inhibiting the expression
of IL-22. Combined treatment regulated the activation of the MAPK
signaling pathway, which provided novel ideas for the treatment of
patients with CRC and depression.
Acknowledgements
Not applicable.
Funding
The current study was supported by the Natural
Science Foundation of China (grant no. 81472853).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
HZ designed the research. MC and YL performed the
mice experiments. XD and CZ conducted histological examinations and
western blotting. HJ and XC performed cell culture experiments and
ELISA. HZ and CZ analyzed the results and wrote the manuscript. All
authors read and approved the final manuscript.
Ethics approval and consent to
participate
All animal experiments were approved by the Animal
Ethics Committee of the Chinese Medical University (Shenyang,
China; approval no. CMU2018178).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Li Q, Liu Y, Su B, Zhao H, Lin Q, Zhu Y,
Zhang L, Weng D, Gong X, Sun X and Xu Y: The CT appearance pattern
of radiation-induced lung injury and tumor recurrence after
stereotactic body radiation therapy in early stage non-small cell
lung cancer. Transl Lung Cancer Res. 9:713–721. 2020.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Feng RM, Zong YN, Cao SM and Xu RH:
Current cancer situation in China: Good or bad news from the 2018
global cancer statistics? Cancer Commun (Lond).
39(22)2019.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Kenzik KM, Kent EE, Martin MY, Bhatia S
and Pisu M: Chronic condition clusters and functional impairment in
older cancer survivors: A population-based study. J Cancer Surviv.
10:1096–1103. 2016.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Miller KD, Sauer AG, Ortiz AP, Fedewa SA,
Pinheiro PS, Tortolero-Luna G, Martinez-Tyson D, Jemal A and Siegel
RL: Cancer statistics for hispanics/latinos, 2018. CA Cancer J
Clin. 68:425–445. 2018.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Chen W, Zheng R, Baade PD, Zhang S, Zeng
H, Bray F, Jemal A, Yu XQ and He J: Cancer statistics in China,
2015. CA Cancer J Clin. 66:115–132. 2016.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Zamora-Ros R, Shivappa N, Steck SE,
Canzian F, Landi S, Alonso MH, Hébert JR and Moreno V: Dietary
inflammatory index and inflammatory gene interactions in relation
to colorectal cancer risk in the bellvitge colorectal cancer
case-control study. Genes Nutr. 10(447)2015.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Raskov H, Pommergaard HC, Burcharth J and
Rosenberg J: Colorectal carcinogenesis-update and perspectives.
World J Gastroenterol. 20:18151–18164. 2014.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Gu MJ, Huang QC, Bao CZ, Li YJ, Li XQ, Ye
D, Ye ZH, Chen K and Wang JB: Attributable causes of colorectal
cancer in China. BMC Cancer. 18(38)2018.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Wang X, Wang B, Zhao J, Liu C, Qu X and Li
Y: MiR-155 is involved in major depression disorder and
antidepressant treatment via targeting SIRT1. Biosci Rep.
38(BSR20181139)2018.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Kosari-Nasab M, Shokouhi G, Ghorbanihaghjo
A, Abbasi MM and Salari AA: Hesperidin attenuates
depression-related symptoms in mice with mild traumatic brain
injury. Life Sci. 213:198–205. 2018.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Rooney AG, McNamara S, Mackinnon M, Fraser
M, Rampling R, Carson A and Grant R: The frequency, longitudinal
course, clinical associations, and causes of emotional distress
during primary treatment of cerebral glioma. Neuro oncol.
15:635–643. 2013.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Hong JS and Tian J: Prevalence of anxiety
and depression and their risk factors in Chinese cancer patients.
Support Care Cancer. 22:453–459. 2014.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Van Gorp H and Lamkanfi M: The emerging
roles of inflammasome-dependent cytokines in cancer development.
EMBO Rep. 20(e47575)2019.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Nocera NF, Lee MC, De La Cruz LM,
Rosemblit C and Czerniecki BJ: Restoring lost anti-HER-2 Th1
immunity in breast cancer: A crucial role for Th1 cytokines in
therapy and prevention. Front Pharmacol. 7(356)2016.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Sconocchia G, Eppenberger S, Spagnoli GC,
Tornillo L, Droeser R, Caratelli S, Ferrelli F, Coppola A, Arriga
R, Lauro D, et al: NK cells and T cells cooperate during the
clinical course of colorectal cancer. Oncoimmunology.
3(e952197)2014.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Kryczek I, Lin Y, Nagarsheth N, Peng D,
Zhao L, Zhao E, Vatan L, Szeliga W, Dou Y, Owens S, et al:
IL-22+ CD4+ T cells promote colorectal cancer
stemness via STAT3 transcription factor activation and induction of
the methyltransferase DOT1L. Immunity. 40:772–784. 2014.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Huang YH, Cao YF, Jiang ZY, Zhang S and
Gao F: Th22 cell accumulation is associated with colorectal cancer
development. World J Gastroenterol. 21:4216–4224. 2015.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Trifari S, Kaplan CD, Tran EH, Crellin NK
and Spits H: Identification of a human helper T cell population
that has abundant production of interleukin 22 and is distinct from
TH-17, TH 1 and TH 2 cells. Nat Immunol. 10:864–871.
2009.PubMed/NCBI View
Article : Google Scholar
|
|
19
|
Lejeune D, Dumoutier L, Constantinescu S,
Kruijer W, Schuringa JJ and Renauld JC: Interleukin-22 (IL-22)
activates the JAK/STAT, ERK, JNK, and p38 MAP kinase pathways in a
rat hepatoma cell line pathways that are shared with and distinct
from IL-10. J Biol Chem. 277:33676–33682. 2002.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Liu Y, Pan W, Yang S, Wu X, Wu J, Ma J,
Yuan Z and Meng S: Interleukin-22 protects rat PC12
pheochromocytoma cells from serum deprivation-induced cell death.
Mol Cell Biochem. 371:137–146. 2012.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Yu H, Fan C, Yang L, Yu S, Song Q, Wang P
and Mao X: Ginsenoside Rg1 prevents chronic stress-induced
depression-like behaviors and neuronal structural plasticity in
rats. Cell Physiol Biochem. 48:2470–2482. 2018.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Andrus BM, Blizinsky K, Vedell PT, Dennis
K, Shukla PK, Schaffer DJ, Radulovic J, Churchill GA and Redei EE:
Gene expression patterns in the hippocampus and amygdala of
endogenous depression and chronic stress models. Mol Psychiatry.
17:49–61. 2012.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Porsolt RD, Le Pichon M and Jalfre M:
Depression: A new animal model sensitive to antidepressant
treatments. Nature. 266:730–732. 1977.PubMed/NCBI View
Article : Google Scholar
|
|
24
|
Osanloo N, Najafi-Abedi A, Jafari F, Javid
F, Pirpiran M, Memar Jafari MR, Khosravi SAM, Behzadi MR, Ranjbaran
M and Sahraei H: Papaver rhoeas L. hydroalcoholic extract
exacerbates forced swimming test-induced depression in mice. Basic
Clin Neurosci. 7:195–202. 2016.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Smith K: Mental health: A world of
depression. Nature. 515(181)2014.PubMed/NCBI View
Article : Google Scholar
|
|
26
|
Mugge L, Mansour TR, Crippen M, Alam Y and
Schroeder J: Depression and glioblastoma, complicated concomitant
diseases: A systemic review of published literature. Neurosurg Rev.
43:497–511. 2020.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Dowlati Y, Herrmann N, Swardfager W, Liu
H, Sham L, Reim EK and Lanctôt KL: A meta-analysis of cytokines in
major depression. Biol Psychiatry. 67:446–457. 2010.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Thaker PH, Han LY, Kamat AA, Arevalo JM,
Takahashi R, Lu C, Jennings NB, Armaiz-Pena G, Bankson JA, Ravoori
M, et al: Chronic stress promotes tumor growth and angiogenesis in
a mouse model of ovarian carcinoma. Nat Med. 12:939–944.
2006.PubMed/NCBI View
Article : Google Scholar
|
|
29
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Quach C, Sanoff HK, Williams GR, Lyons JC
and Reeve BB: Impact of colorectal cancer diagnosis and treatment
on health-related quality of life among older Americans: A
population-based, case-control study. Cancer. 121:943–950.
2015.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Shaheen Al Ahwal M, Al Zaben F, Khalifa
DA, Sehlo MG, Ahmad RG and Koenig HG: Depression in patients with
colorectal cancer in Saudi Arabia. Psychooncology. 24:1043–1050.
2015.PubMed/NCBI View
Article : Google Scholar
|
|
32
|
White AJ, Reeve BB, Chen RC, Stover AM and
Irwin DE: Coexistence of urinary incontinence and major depressive
disorder with health-related quality of life in older Americans
with and without cancer. J Cancer Surviv. 8:497–507.
2014.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Santos JC and Pyter LM: Neuroimmunology of
behavioral comorbidities associated with cancer and cancer
treatments. Front Immunol. 9(1195)2018.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Ehlert U, Gaab J and Heinrichs M:
Psychoneuroendocrinological contributions to the etiology of
depression, posttraumatic stress disorder, and stress-related
bodily disorders: The role of the hypothalamus-pituitary-adrenal
axis. Biol Psychol. 57:141–152. 2001.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Wang J, Chen Y, Dai C, Shang Y and Xie J:
Ginsenoside Rh2 alleviates tumor-associated depression in a mouse
model of colorectal carcinoma. Am J Transl Res. 8:2189–2195.
2016.PubMed/NCBI
|
|
36
|
Hawkins NA, Soman A, Buchanan Lunsford N,
Leadbetter S and Rodriguez JL: Use of medications for treating
anxiety and depression in cancer survivors in the United States. J
Clin Oncol. 35:78–85. 2017.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Kulkarni SK and Dhir A: Current
investigational drugs for major depression. Expert Opin Investig
Drugs. 18:767–788. 2009.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Dumoutier L, Louahed J and Renauld JC:
Cloning and characterization of IL-10-related T cell-derived
inducible factor (IL-TIF), a novel cytokine structurally related to
IL-10 and inducible by IL-9. J Immunol. 164:1814–1819.
2000.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Logsdon NJ, Jones BC, Josephson K, Cook J
and Walter MR: Comparison of interleukin-22 and interleukin-10
soluble receptor complexes. J Interferon Cytokine Res.
22:1099–1112. 2002.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Rutz S, Eidenschenk C and Ouyang W: IL-22,
not simply a Th17 cytokine. Immunol Rev. 252:116–132.
2013.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Brand S, Beigel F, Olszak T, Zitzmann K,
Eichhorst ST, Otte JM, Diepolder H, Marquardt A, Jagla W, Popp A,
et al: IL-22 is increased in active Crohn's disease and promotes
proinflammatory gene expression and intestinal epithelial cell
migration. Am J Physiol Gastrointest Liver Physiol. 290:G827–G838.
2006.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Ikeuchi H, Kuroiwa T, Hiramatsu N, Kaneko
Y, Hiromura K, Ueki K and Nojima Y: Expression of interleukin-22 in
rheumatoid arthritis: Potential role as a proinflammatory cytokine.
Arthritis Rheum. 52:1037–1046. 2005.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Wolk K, Kunz S, Witte E, Friedrich M,
Asadullah K and Sabat R: IL-22 increases the innate immunity of
tissues. Immunity. 21:241–254. 2004.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Gurney AL: IL-22, a Th1 cytokine that
targets the pancreas and select other peripheral tissues. Int
Immunopharmacol. 4:669–677. 2004.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Aujla SJ and Kolls JK: IL-22: A critical
mediator in mucosal host defense. J Mol Med (Berl). 87:451–454.
2009.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Nagalakshmi ML, Rascle A, Zurawski S,
Menon S and de Waal Malefyt R: Interleukin-22 activates STAT3 and
induces IL-10 by colon epithelial cells. Int Immunopharmacol.
4:679–691. 2004.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Nardinocchi L, Sonego G, Passarelli F,
Avitabile S, Scarponi C, Failla CM, Simoni S, Albanesi C and Cavani
A: Interleukin-17 and interleukin-22 promote tumor progression in
human nonmelanoma skin cancer. Eur J Immunol. 45:922–931.
2015.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Park O, Wang H, Weng H, Feigenbaum L, Li
H, Yin S, Ki SH, Yoo SH, Dooley S, Wang FS, et al: In vivo
consequences of liver-specific interleukin-22 expression in mice:
Implications for human liver disease progression. Hepatology.
54:252–261. 2011.PubMed/NCBI View Article : Google Scholar
|
|
49
|
Herzallah MM, Moustafa AA, Natsheh JY,
Danoun OA, Simon JR, Tayem TI, Sehwail MA, Amleh I, Bannoura I,
Petrides G, et al: Depression impairs learning, whereas the
selective serotonin reuptake inhibitor, paroxetine, impairs
generalization in patients with major depressive disorder. J Affect
Disord. 151:484–492. 2013.PubMed/NCBI View Article : Google Scholar
|
|
50
|
Chang L and Karin M: Mammalian MAP kinase
signalling cascades. Nature. 410:37–40. 2001.PubMed/NCBI View Article : Google Scholar
|
|
51
|
Devendrakumar KT, Li X and Zhang Y: MAP
kinase signalling: Interplays between plant PAMP-and
effector-triggered immunity. Cell Mol Life Sci. 75:2981–2989.
2018.PubMed/NCBI View Article : Google Scholar
|
|
52
|
Zhang X, Liu W, Li Y, Li G and Xu JR:
Expression of HopAI interferes with MAP kinase signalling in
magnaporthe oryzae. Environ Microbiol. 19:4190–4204.
2017.PubMed/NCBI View Article : Google Scholar
|
|
53
|
Guo C, Hou J, Ao S, Deng X and Lyu G:
HOXC10 up-regulation promotes gastric cancer cell proliferation and
metastasis through MAPK pathway. Chin J Cancer Res. 29:572–580.
2017.PubMed/NCBI View Article : Google Scholar
|
|
54
|
Zhang G, Cheng Y, Zhang Q, Li X, Zhou J,
Wang J and Wei L: ATX-LPA axis facilitates estrogen-induced
endometrial cancer cell proliferation via MAPK/ERK signaling
pathway. Mol Med Rep. 17:4245–4252. 2018.PubMed/NCBI View Article : Google Scholar
|
|
55
|
Park SS, Park SK, Lim JH, Choi YH, Kim WJ
and Moon SK: Esculetin inhibits cell proliferation through the
Ras/ERK1/2 pathway in human colon cancer cells. Oncol Rep.
25:223–230. 2011.PubMed/NCBI
|
|
56
|
Meloche S and Pouysségur J: The ERK1/2
mitogen-activated protein kinase pathway as a master regulator of
the G1-to S-phase transition. Oncogene. 26:3227–3239.
2007.PubMed/NCBI View Article : Google Scholar
|
|
57
|
Kim K, Kim G, Kim JY, Yun HJ, Lim SC and
Choi HS: Interleukin-22 promotes epithelial cell transformation and
breast tumorigenesis via MAP3K8 activation. Carcinogenesis.
35:1352–1361. 2014.PubMed/NCBI View Article : Google Scholar
|
|
58
|
Tan AHM and Lam KP: Pharmacologic
inhibition of MEK-ERK signaling enhances Th17 differentiation. J
Immunol. 184:1849–1857. 2010.PubMed/NCBI View Article : Google Scholar
|