Introduction
Prostate cancer (PCa) is an epithelial malignancy
that occurs in the prostate (1).
PCa mainly occurs in men over the age of 50 and is the second
leading cause of cancer death worldwide (2,3).
Currently, endocrine therapy, including surgical or drug castration
and antiandrogen (bicalutamide or flutamide) therapy, is the main
treatment for hormone-sensitive advanced PCa patients (4). However, the vast majority of patients
are eventually treated with androgen-deprivation therapy and
progress to metastatic castration-resistant PCa (CRPC), which is
the leading cause of PCa-related mortality (5-7).
Novel treatments, such as docetaxel and abiraterone, were shown to
improve the survival of patients with metastatic CRPC; however,
most patients develop drug resistance (8,9).
While healthy cells rely on carbohydrate molecules
oxidized in mitochondria to acquire energy, most tumor cells obtain
their energy supply through relatively low-yield glycolysis, which
does not involve oxygen or mitochondria (10). Malignant, rapidly growing tumor
cells typically have a 200-fold higher rate of glycolysis compared
with normal tissues, even under oxygen-sufficient conditions
(11). Therefore, it is speculated
that this change in metabolism is the root cause of cancer
(12). It was reported that
glycolysis in cancer cells is characterized by high glucose
consumption and lactate production (13). Cancer cells often take up high
amounts of glucose and rely on glycolysis for ATP generation, more
efficiently converting glucose into macromolecules that are needed
for a variety of cellular processes (14-16).
Pyruvate kinase (PKM2), a key rate-limiting enzyme that
catalyzes the final step in glycolysis, was reported to be highly
expressed in multiple cancers (17,18)
and can promote glucose metabolism and cell growth (19). A number of studies showed that
upregulation of PKM2 can promote malignancy and
downregulation of PKM2 can inhibit cell growth, migration
and invasion in various types of cancer (20-24).
MicroRNAs (miRs/miRNAs) are small noncoding,
single-stranded RNAs which regulate gene expression by regulating
the stability or translation of target mRNAs by binding to their
3'-untranslated regions (UTRs) (25,26).
Studies showed that miRNAs have important applications in the field
of cancer diagnosis and treatment (27-32).
For example, exo-anti-miR-214 can reverse the resistance of gastric
cancer cells to cisplatin (33).
MiR-122, an abundant liver-specific miRNA, was shown to reverse
doxorubicin resistance in liver cancer cells by inhibiting
glycolysis in tumors via PKM2 inhibition (34,35).
In colon cancer, overexpression of miR-122 can increase the
sensitivity of fluorouracil (5-FU)-resistant colon cancer cells to
5-FU by PKM2 downregulation (36). Previous findings showed that miR-34a
and miR-21 play a role in the chemoresistance of PCa cells
(37-40).
The present study aimed to investigate the function of miRNA-122 in
the chemoresistance of PCa cells and the underlying mechanism.
Materials and methods
Cell culture
Prostate cancer docetaxel-resistant
(LNCaP/Docetaxel) and docetaxel-sensitive (LNCaP) LNCaP cells were
purchased from the Cell Bank of Type Culture Collection of the
Chinese Academy of Sciences. Cells were cultured with DMEM (cat.
no. SH30243.01; HyClone; GE Healthcare Life Sciences) supplemented
with 10% FBS (cat. no. 16000-044; Gibco; Thermo Fisher Scientific,
Inc.) and 1% penicillin and streptomycin (100X; cat. no. P1400;
Beijing Solarbio Science & Technology Co., Ltd.) in a 37˚C
incubator (Forma 3111; Thermo Fisher Scientific, Inc.) with 5%
CO2.
Isolation of primary PCa cells
PCa cells were isolated from 20 patients with PCa
who were treated in Shaoxing People's Hospital, Shaoxing, China
from February 2018 to February 2019 (age range, 60-80 years; mean
age, 70.12±8.43 years). The inclusion criteria were as follows: i)
patients did not receive any treatment and ii) clinical data of
patients were complete. All cases were confirmed by a review by the
Shaoxing People's Hospital Pathology Center. The exclusion criteria
were cases without complete clinical data. Fresh prostate cancer
tissues were washed three times with D-Hank's balanced salt
solution containing 500 IU/ml penicillin and streptomycin. Once
surrounding inactivated tissues (cloudy appearance, dull and loss
of normal tissue elasticity) were removed with ophthalmic scissors,
the tissues (normal color and elasticity) were cut into pieces
(~300 times) in a sterile 5 ml syringe and incubated with 5 ml
trypsin-EDTA (0.125% trypsin and 0.53 mol/l EDTA) for 5 min at
37˚C. A total of 10 ml RPMI-1640 medium (cat. no. 88365; Thermo
Fisher Scientific, Inc.) containing 10% FBS (Gibco; Thermo Fisher
Scientific, Inc.) was added to terminate digestion in a 15 ml
centrifuge tube. After centrifugation at 4˚C and 800 x g for 5 min,
the pellet was resuspended in 5 ml RPMI 1640 containing 5 ng/ml
epidermal growth factor (cat. no. PHG0311L; Gibco; Thermo Fisher
Scientific, Inc.), 50 µg/ml bovine pituitary extract (cat. no.
13028014; Gibco; Thermo Fisher Scientific, Inc.) and 10% FBS. A
total of 5 ml of the suspension was seeded in the T25 cell flasks
for incubation in a 37˚C and 5% CO2 incubator.
Fibroblasts can adherent growth in the T25 cell flasks after 4-6
days. Medium was replaced every two days, and the cells were
passaged when they were reached 80% confluence. Cells that were
sub-cultured 3 times were used for subsequent experiments.
Lentivirus construction
The coding sequence (CDS) of PKM2
(AY352517.1) was synthesized and validated by DNA sequencing.
Following the insertion of the PKM2 CDS into the pLVX-Puro
vector (Clontech Laboratories, Inc.) at the
EcoRI-BamHI site, the pLVX-Puro-PKM2 plasmid
was co-transfected into 293T cells (American Type Culture
Collection) cultured with DMEM containing 10% FBS and 1%
penicillin-streptomycin, with the viral packaging plasmids psPAX2
and pMD2.G (Addgene, Inc.) using Lipofectamine™ 2000 (Invitrogen;
Thermo Fisher Scientific, Inc). After 48 h of transfection, the
supernatant was collected by centrifugation for 5 min at 1,000 x g
and 4˚C.
Cell transfection
LNCaP and LNCaP/Docetaxel cells in the logarithmic
growth phase were suspended to 1x106 cells/ml after
trypsinization. Subsequently, 2 ml cell suspension was inoculated
into six-well plates for overnight culture at 37˚C with 5%
CO2. Once the cells grew to 60-70% confluence, LNCaP and
LNCaP/Docetaxel cells were transfected with 5 µl negative control
(NC)-miRNA (100 pmol; 5'-CAGUACUUUUGUGUAGUACAA-3'), 5 µl
hsa-miR-122-5p inhibitor (100 pmol; 5'-CAAACACCATTGTCACACTCCA-3')
or 5 µl of hsa-miR-122-5p mimic (100 pmol;
5'-UGGAGUGUGACAAUGGUGUUUG-3') using Lipofectamine™ 2000 (cat. no.
11668-019; Invitrogen; Thermo Fisher Scientific, Inc.). Following
24 h of transfection, serum-free transfer solution was replaced
with complete medium to culture for a further 48 h.
Docetaxel treatment
LNCaP and LNCaP/Docetaxel cells were treated with
gradient concentrations of docetaxel (0.25, 0.5, 1, 2, 4, 8, 16 and
32 µg/ml; cat. no. 114977-28-5; Shanghai Aladdin Biochemical
Technology Co., Ltd.), followed by the detection of cell inhibition
rate. After treatment with docetaxel, the apoptosis levels of
primary PCa cells, treated LNCaP or treated LNCaP/Docetaxel cells
were detected.
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was isolated from PCa cells (LNCaP,
LNCaP/Docetaxel or primary PCa cells) with or without miR-122
inhibitor or mimic, and/or combined with PKM2 overexpression
(oePKM2) lentivirus using TRIzol® reagent (cat.
no. 15996026; Invitrogen; Thermo Fisher Scientific, Inc.) according
to the manufacturer's instructions. Following RNA quanrification
and integrity confirmation, ~1 µg of RNA was reversed transcribed
into cDNA using the RevertAid First Strand cDNA Synthesis kit (cat.
no. K1622; Thermo Fisher Scientific, Inc.) using the following heat
cycle: 37˚C for 5 min; 55˚C for 15 min and 85˚C for 5 min. qPCR was
performed on a 7300 Real-Time PCR system (Applied Biosystems;
Thermo Fisher Scientific, Inc.) with a miRNA RT-PCR Detection Kit
(cat. no. AOMD-Q020; GeneCopoeia, Inc.) or a Maxima SYBR Green/ROX
qPCR master mix (cat. no. K0223; Thermo Fisher Scientific, Inc.).
The following thermocycling conditions were used for the qPCR:
Initial denaturation at 95˚C for 10 min, 40 cycles of 95˚C for 15
sec and 60˚C for 45 sec and a final extension step at 95˚C for 15
sec. miR-122 expression were normalized to U6 levels and
PKM2 expression were normalized to GAPDH levels using the
2-ΔΔCq method (41). The primer pairs used for the qPCR
are listed in Table I.
 | Table IPrimer sequences used for reverse
transcription-quantitative PCR. |
Table I
Primer sequences used for reverse
transcription-quantitative PCR.
| Gene | Forward
sequence | Reverse
sequence |
|---|
| RT primer for
miR-122 | |
5'-GTCGTATCCAGTGCAGGGTCCGAGG |
| | |
TATTCGCACTGGATACGACGCCTAG-3' |
| miR-122 |
5'-CGCCATTATCACACTAAATAGCTACTG-3' |
5'-AGTGCAGGGTCCGAGGTATT-3' |
| PKM2 |
5'-TCCAGGTGAAGCAGAAAG-3' |
5'-CGGATGAATGACGCAAAC-3' |
| U6 |
5'-CTCGCTTCGGCAGCACA-3' |
5'-AACGCTTCACGAATTTGCGT-3' |
| GAPDH |
5'-AATCCCATCACCATCTTC-3' |
5'-AGGCTGTTGTCATACTTC-3' |
Western blot analysis
Total protein was isolated from PCa cells (LNCaP,
LNCaP/Docetaxel or primary PCa cells) with or without treatment of
miR-122 inhibitor or mimic, and/or combined with oePKM2
lentivirus using RIPA buffer supplemented with protease and
phosphatase inhibitors (cat. no. R0010; Beijing Solarbio Science
& Technology Co., Ltd.). Following protein quantification using
a BCA protein quantification kit (Thermo Fisher Scientific, Inc.),
25 µg of protein/lane was separated via 10% SDS PAGE followed by
semi-dry transfer onto PVDF membranes (cat. no. HATF00010; EMD
Millipore). After blocking with 5% skimmed milk for 1 h at room
temperature, membranes were incubated overnight with primary
antibodies against PKM2 (1:500; cat. no. ab137852; Abcam)
and GAPDH (1:2,000; cat. no. 5174; Cell Signaling Technology, Inc.)
at 4˚C with gentle agitationFollowing six washes with TBS-Tween-20,
membranes were incubated for 2 h at room temperature with goat
anti-rabbit horseradish peroxidase (HRP)-labeled secondary
antibodies (1:1.000; cat. no. A0208; Beyotime Institute of
Biotechnology). Protein bands were visualized using an Tanon 5200
chemiluminescent imaging system (Tanon Science and Technology Co.,
Ltd.) after 5 min of development with the Immobilon Western
Chemiluminescent HRP substrate (cat. no. WBKLS0100; EMD Millipore)
in the dark. PKM2 protein expression was quantified using
ImageJ (version 1.47; National Institutes of Health) with GAPDH as
the loading control.
Cell proliferation assay
PCa cells (LNCaP or LNCaP/Docetaxel) in the
logarithmic growth phase were trypsinized and a 3x104
cells/ml suspension was prepared by counting the cells under an
inverted microscope at x40 magnification (XDS-500C; Shanghai Caikon
Optical Instrument Co., Ltd.). Subsequently, in a 96-well culture
plate, 100 µl of the suspension was inoculated and cultured at 37˚C
overnight. A total of 100 µl DMEM was used as the blank control.
Cell Counting Kit-8 solution (CCK-8; cat. no. ab228554; Abcam) and
serum-free DMEM were mixed at a volume ratio of 1:10. Following 0,
24, 48 and 72 h of treatment with miR-122 inhibitor or miR-122
mimic and oePKM2 lentivirus, 100 µl of the above CCK-8
mixture was added to the cells and incubated in a 5% CO2
incubator at 37˚C for 1 h. The absorbance was read at a wavelength
of 450 nm using a microplate reader (cat. no. DNM-9602; Perlong
Medical Equipent Co., Ltd.).
Cell apoptosis assay
Following 48 h of treatment, each group of cells
(LNCaP, LNCaP/Docetaxel or primary PCa cells) was collected using
0.05% trypsin) to detect apoptosis using an Annexin V-FITC cell
apoptosis detection kit (cat. no. C1062L; Beyotime Institute of
Biotechnology). In brief, ~1x106 cells were centrifuged
at 1,000 x g for 5 min at 4˚C. After discarding the supernatant,
the cells were gently resuspended in 195 µl Annexin V-FITC binding
solution, followed by a 15-min incubation with 5 µl Annexin V-FITC
at 4˚C in the dark. Subsequently, 5 µl of propidium iodide (PI)
staining solution was added and the cells were incubated for 5 min
at 4˚C in the dark. A tube without Annexin V-FITC and PI was used
as a negative control. Flow cytometry was performed and apoptosis
percentages were assessed with BD Accuri C6 software (version
1.0.264.21; BD Biosciences).
Detection of glucose uptake and
lactate production
PCa cells (LNCaP or LNCaP/Docetaxel) were seeded in
24-well plates and cultured overnight and treated with miR-122
inhibitor or miR-122 mimic and oePKM2 lentivirus. A 2-NBDG
Glucose Uptake Assay kit (cat. no. K682-50; BioVision, Inc.) was
used for glucose uptake detection. Following 72 h of treatment, the
cells were incubated with 100 µM 2-NBDG for 1 h. After two washes
with PBS, cells were trypsinized and resuspended in DMEM containing
10% FBS, followed by incubation with 5 µg/ml PI for staining.
Subsequently, flow cytomtery was performed to measure the
proportion of PI-negative and 2-NBDG-positive cells, and glucose
uptake was calculated. Lactate production was measured using a
lactate test kit (cat. no. A019-2; Nanjing Jiancheng Bioengineering
Institute) according to the manufacturer's instructions. The
absorbance was measured at a wavelength of 530 nm using a
spectrophotometer and lactate production was calculated.
Luciferase reporter assay
TargetScanHuman 7.2 (http://www.targetscan.org/vert_72/) was used to
predict miR-122 target sites on PKM2. PCa cells (LNCaP or
LNCaP/Docetaxel) in the logarithmic growth phasewere trypsinized
and centrifuged at 800 x g for 5 min at room temperature. After
discarding the supernatant, cells were gently resuspended in 1 ml
DMEM and counted under an inverted microscope at x40 magnification
(XDS-500C; Shanghai Caikon Optical Instrument Co., Ltd.). The cell
suspension was inoculated into a six-well plate at a density of
5x105 cells/well and cultured in an incubator at 37˚C.
Following 24 h of culture, the cells were co-transfected with 1.5
µg luciferase plasmid (pGL3-Promoter-PKM; Promega Corporation) and
miR-122-5p inhibitor or mimic using Lipofectamine™ 2000 (cat. no.
11668-019; Invitrogen; Thermo Fisher Scientific, Inc.). Following
transfection, the cells were washed with PBS, and then incubated in
500 µl PLB for 15 mins with gentle agitation at room temperature.
The luciferase activity of the PKM2 reporter was determined
using a Dual-Luciferase Reporter Assay system (cat. no. E1910;
Promega Corporation) on a GloMax®-Multi+ Microplate
Multimode reader (Promega Corporation). Firefly luciferase activity
was detected following addition of 100 µl LAR II and 20 µl sample
lysate in 96-well plates. Renilla luciferase activity was
detected following addition of 100 µl Stop & Glo reagent.
Statistical analysis
GraphPad Prism 7.0 (GraphPad Software, Inc.) was
used for statistical analysis. Data are presented as the mean ± SD
from triplicate experiments. Unpaired Student's t-test was used to
determine the significance between two groups, while multiple
groups were compared by one-way ANOVA and Tukey's post hoc test.
The Pearson correlation coefficient was used to analyze the
correlation between miR-122 and PKM2. P<0.05 was
considered to indicate a statistically significant difference.
Results
Expression of miR-122 significantly
decreased in LNCaP/Docetaxel cells, and inhibition of miR-122 in
LNCaP cells significantly promotes proliferation and glycolysis and
inhibits apoptosis
Following treatment with gradient concentrations of
docetaxel, cell proliferation was detected to determine the drug
resistance of LNCaP/Docetaxel cells to docetaxel. As shown in
Fig. 1A, the half-maximal
inhibitory concentration (IC50) in LNCaP/docetaxel cells
was significantly higher compared with LNCaP cells, indicating that
LNCaP/Docetaxel cells were docetaxel-resistant. Consistent with a
previous report (42), 10 µg/ml of
docetaxel was used for subsequent experiments. Baseline
proliferation levels of LNCaP/Docetaxel cells were significantly
higher compared with LNCaP cells (Fig.
1B). RT-qPCR was performed to detect the expression of miR-122
in human PCa docetaxel-resistant (LNCaP/Docetaxel) and -sensitive
(LNCaP) cell strains. The results in Fig. 1C show that compared with LNCaP
cells, miR-122 levels significantly decreased in LNCaP/Docetaxel
cells. Furthermore, LNCaP cells were treated with miR-122 inhibitor
(Fig. 1D). Following miR-122
inhibition, the proliferation of LNCaP cells significantly
increased (Fig. 1E), and apoptosis
was significantly decreased (Fig.
1F), which was accompanied by significantly increased glucose
uptake (Fig. 1G) and lactate
production (Fig. 1H) compared with
the NC group. The findings suggested that miR-122 expression may be
associated with docetaxel resistance in PCa.
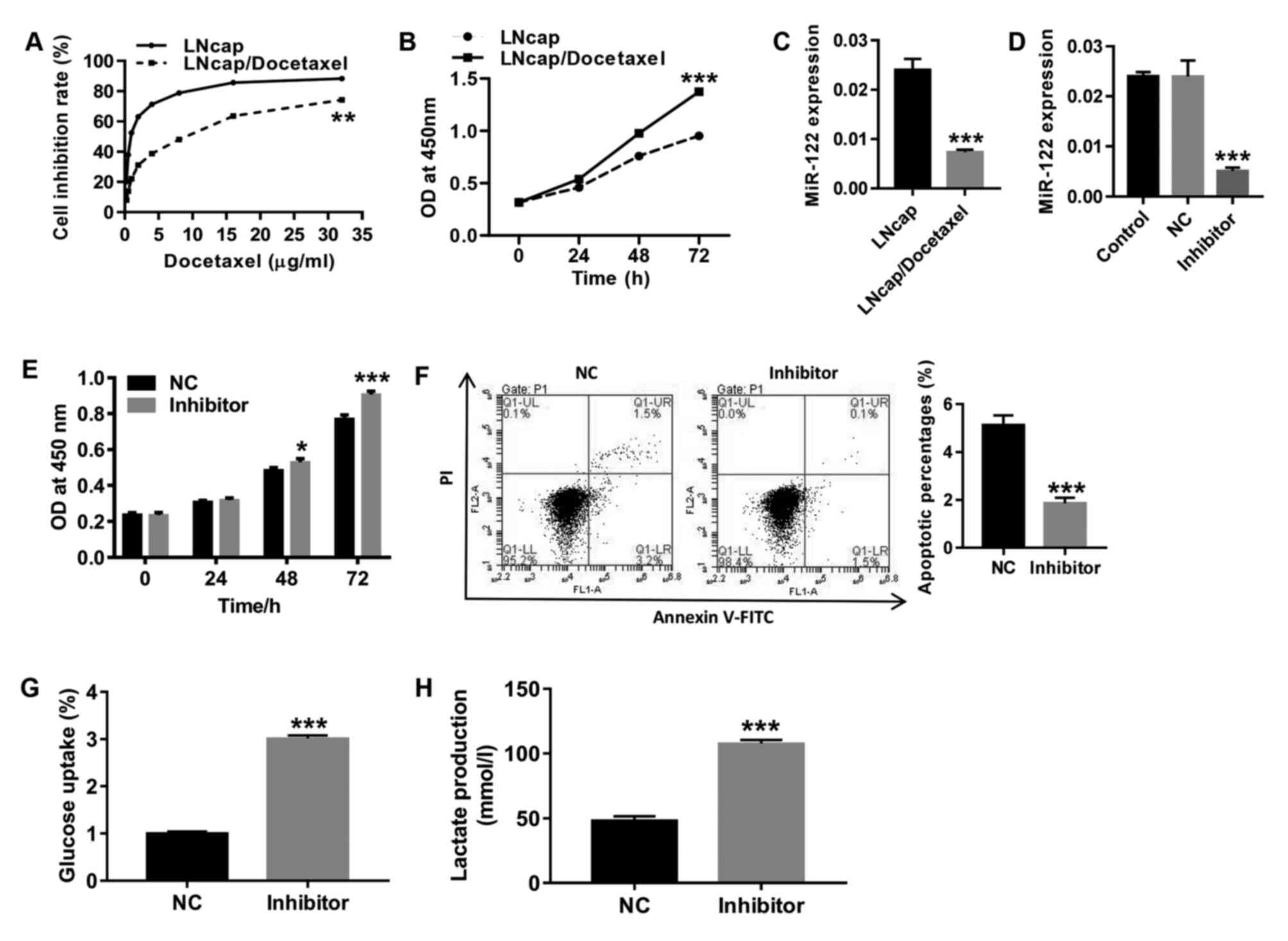 | Figure 1Expression of miR-122 significantly
decreased in LNCaP/Docetaxel cells and inhibition of miR-122 in
LNCaP cells significantly promotes proliferation and glycolysis and
inhibits apoptosis. (A) Following treatment with gradient
concentrations of docetaxel (0.25, 0.5, 1, 2, 4, 8, 16 and 32
µg/ml), cell proliferation was assessed to determine resistance to
docetaxel in LNCaP and LNCaP/Docetaxel cells.
**P<0.01. (B) Baseline proliferation levels of LNCaP
and LNCaP/Docetaxel cells were detected. ***P<0.001.
(C) Expression of miR-122 in LNCaP and LNCaP/Docetaxel cells was
detected. ***P<0.001. (D) The expression of miR-122
in miR-122-treated LNCaP cells was detected.
***P<0.001 vs. NC. (E) Cell proliferation was
detected at 0, 24, 48 and 72 h. *P<0.05 and
***P<0.001. (F) Cell apoptosis was detected at 48 h.
The ordinate of the histogram is the sum of early and late
apoptosis. ***P<0.001. (G) Glucose uptake and (H)
lactate production were detected. ***P<0.001.
miR-122, microRNA-122; OD, optical density; NC, negative control;
PI, propidium iodide. |
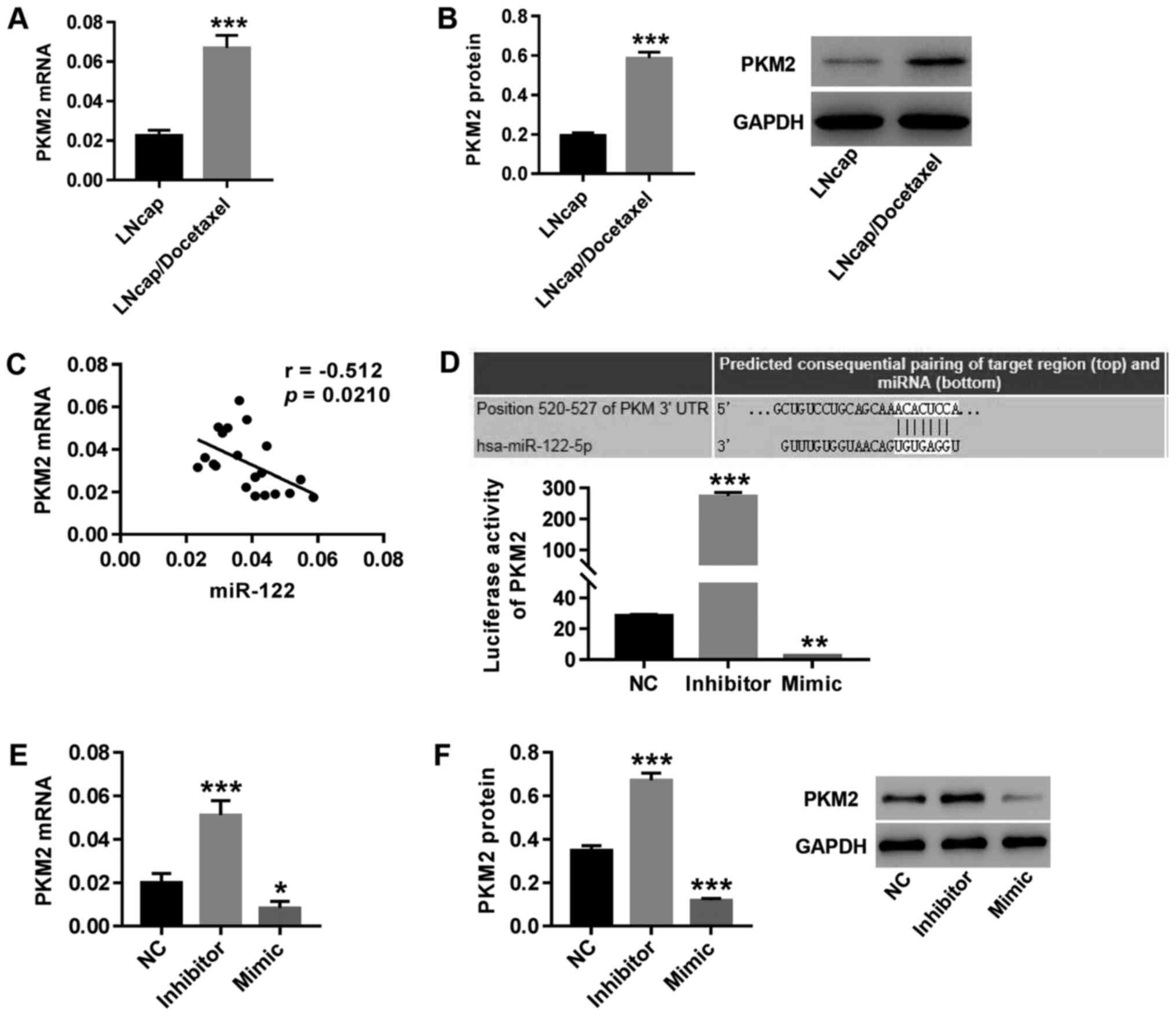
PKM2 may be a target gene of miR-122
in regulating PCa
Both mRNA (Fig. 2A)
and protein (Fig. 2B) expression of
PKM2 significantly increased in LNCaP/Docetaxel cells
compared with LNCaP cells. PKM2 expression was negatively
correlated with miR-122 expression in primary PCa cells isolated
from tumor tissues of 20 patients with PCa (Fig. 2C). Furthermore, TargetScan was used
to predict the binding site between the PKM2 3'-UTR and
miR-122. Compared with the NC group, theluciferase reporter assay
showed that following inhibition of miR-122, the luciferase
activity of the PKM2 reporter was significantly increased
(Fig. 2D), accompanied by increased
expression of PKM2 (Fig. 2E
and F), whilemiR-122 mimic
transfection had the opposite effect. The results indicated that
PKM2 might be a target gene of miR-122 in regulating
PCa.
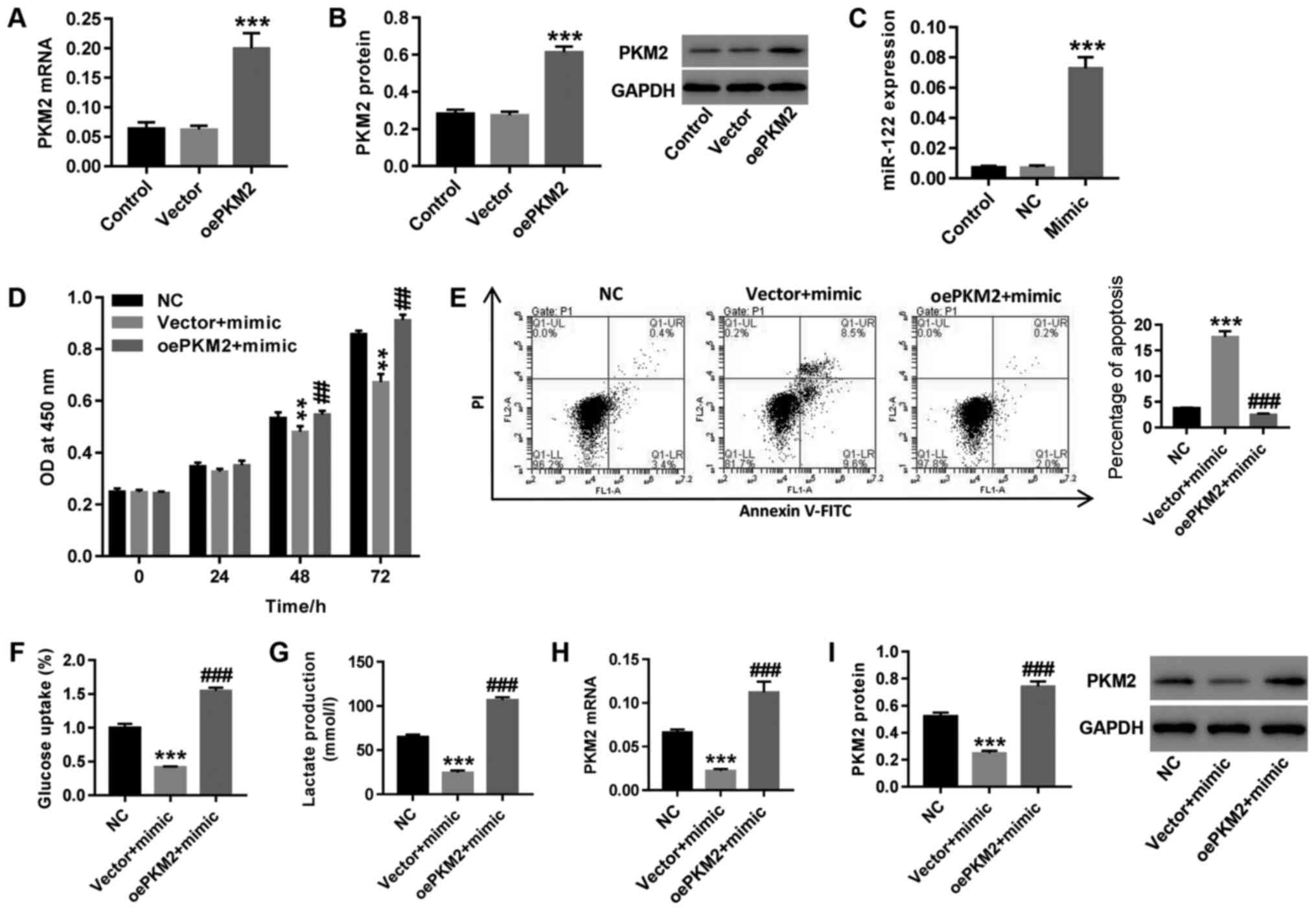
miR-122 possibly regulates the
docetaxel resistance of PCa cells via PKM2 regulation
The human PCa docetaxel-resistant cell strain
LNCaP/Docetaxel was treated with both miR-122 mimic and
oePKM2 lentivirus. As shown in Fig. 3A-C, oePKM2 and miR-122 mimic
treatment in LNCaP/Docetaxel cells significantly increased
PKM2 and mir-122 expression compared with the vector and NC
groups, respectively. Upregulation of miR-122 following miR-122
mimic transfection resulted in significantly increased cell
proliferation (Fig. 3D), glucose
uptake (Fig. 3F) and lactate
production (Fig. 3G) in
LNCaP/Docetaxel, cells, whereas cell apoptosis (Fig. 3E) increased, concurrent with a
decrease inthe expression of PKM2 (Fig. 3H and I). Overexpression of PKM2
counteracted the effects of miR-122 mimic on LNCaP/Docetaxel cells.
The results demonstrated that miR-122 regulated docetaxel
resistance in PCa cells, possibly via regulating PKM2.
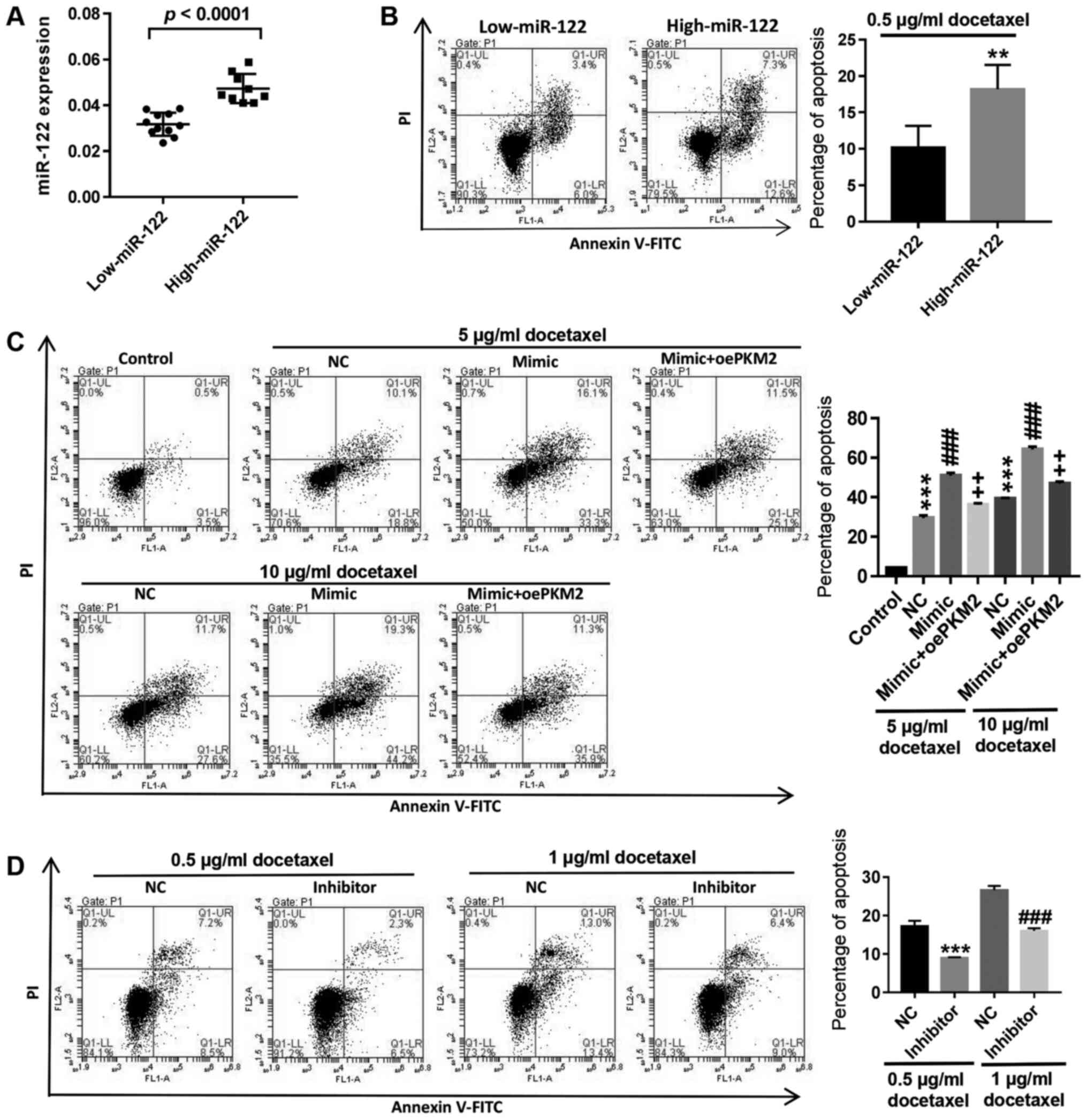
Upregulation of miR-122 could reverse
the resistance of LNCaP/Docetaxel cells to docetaxel
The primary cells isolated from 20 patients with PCa
were divided into two groups: Low expression and high expression of
miR-122 (Fig. 4A). Flow cytometry
analysis showed that following treatment with 0.5 µg/ml docetaxel,
apoptosis in primary PCa cells with high miR-122 expression
significantly increased compared with cells with low miR-122
expression (Fig. 4B). In
LNCaP/Docetaxel cells, upregulation of miR-122 expression by
miR-122 mimic transfection significantly increased
docetaxel-induced apoptosis, while overexpression of PKM2
counteracted the effect of miR-122 mimic transfection, and the
effect of 10 µg/ml docetaxel was more significant compared with 5
µg/ml docetaxel (Fig. 4C). By
contrast, inhibition of miR-122 in LNCaP cells significantly
decreased docetaxel-induced apoptosis (Fig. 4D). The results demonstrated that
high expression of miR-122 could promote docetaxel-induced
apoptosis in PCa cells and upregulation of miR-122 could reverse
the resistance of LNCaP/Docetaxel cells to docetaxel.
Discussion
An increasing number of studies have reported that
the sensitivity of tumor cells to anticancer drugs can be altered
by miRNAs (43-45).
miR-34a was reported to enhance the chemosensitivity of PC3 cells
to paclitaxel and camptothecin and PCa cells treated with miR-143
showed higher chemosensitivity to docetaxel (37,46).
Previous research also showed that miR-122 can reverse drug
resistance in several cancers (34,36).
In the present study, significantly decreased miR-122 levels were
observed in human PCa LNCaP/Docetaxel cells compared with LNCaP
cells. miR-122 mimic transfection in LNCaP/Docetaxel PCa cells
significantly decreased cell proliferation, increased apoptosis and
inhibited glycolysis, whilemiR-122 inhibitor transfection in LNCaP
showed the opposite effect, suggesting that miR-122 expression may
be associated with docetaxel resistance in PCa by regulating cell
proliferation, apoptosis and glycolysis. In primary PCa cells, high
expression of miR-122 could promote docetaxel-induced apoptosis in
PCa cells, and miR-122 mimic transfection significantly increased
docetaxel-induced apoptosis in LNCaP/Docetaxel cells, while
inhibition of miR-122 showed the opposite effect. Thus, it was
speculated that upregulation of miR-122 could reverse the
resistance of LNCaP/Docetaxel cells to docetaxel, which may
contribute to the treatment of PCa chemoresistance.
Furthermore, the underlying mechanism of miR-122 in
regulating docetaxel resistance in PCa was investigated. In tumors,
miRNAs primarily function via regulation of their target genes by
targeting specific mRNAs for degradation or translation inhibition
(47). A study reported that
upregulation of miR-328 can enhance docetaxel sensitivity, decrease
cell proliferation and increase apoptosis in PCa cells by directly
targeting p21-activated protein kinase 6(48). Additionally, the ectopic expression
of miR-21 can increase the resistance of PC3 cells to docetaxel by
targeting the tumor suppressor programmed cell death protein
4(40). In addition, a previous
study revealed that in human lung cancer xenografts in mice,
inhibition of PKM2 could enhance the efficacy of docetaxel
(49). Results of the present study
showed that the expression of PKM2 in PCa cells negatively
correlated with miR-122 expression, and the luciferase reporter
assay showed that miR-122 regulated PKM2 expression by
binding to the 3'-UTR of PKM2. In LNCaP/Docetaxel PCa cells,
miR-122 mimic-induced cell proliferation decreased, apoptosis
increased and glycolysis inhibition was counteracted by PKM2
overexpression. Consistent with previous reports on miR-122 in
cancer chemoresistance (34-36),
it can be inferred that the upregulation of miR-122 expression may
reverse the resistance of PCa LNCaP/Docetaxel cells to docetaxel
via downregulation of its target gene PKM2. However, the
present study also had limitations, such as the lack of sequencing
data and validation, as well as the lack of studies on other
prostate cancer cell types. Mechanisms of miR-122 involved in
docetaxel resistance and the function of miR-122 in other prostate
cancer cell types can be investigated in future studies to further
confirm the current results.
In conclusion, the results demonstrated that high
expression of miR-122 could promote docetaxel-induced apoptosis in
PCa cells and that the upregulation of miR-122 could reverse the
resistance of LNCaP/Docetaxel cells to docetaxel, possibly via the
regulation of its target protein PKM2 by binding to the
3'-UTR. These findings may provide a link between PCa
chemoresistance and miRNAs, and targeting miRNA-122 may offer a
novel therapy for the chemoresistance of PCa.
Acknowledgements
Not applicable.
Funding
This study was funded by the Shaoxing Municipal
Bureau of Science and Technology in China (grant no.
2017B70032).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
ZZ and JY conceived and designed the study. ZZ, JY
and GT performed the experiments. ZZ and JY wrote the manuscript.
All authors read and approved the final manuscript.
Ethics approval and consent to
participate
All experiments conducted in this study were
approved by the Ethics Committee of Shaoxing People's Hospital.
Written informed consent was obtained.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Han W and Li J: Structure-activity
relationship analysis of 3-phenylpyrazole derivatives as androgen
receptor antagonists. J Biomol Struct Dyn: 1-10, Jul 5, 2019 (Epub
ahead of print).
|
|
2
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2019. CA Cancer J Clin. 69:7–34. 2019.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Fendler A, Jung M, Stephan C, Honey RJ,
Stewart RJ, Pace KT, Erbersdobler A, Samaan S, Jung K and Yousef
GM: miRNAs can predict prostate cancer biochemical relapse and are
involved in tumor progression. Int J Oncol. 39:1183–1192.
2011.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Pereira-Lourenço M, Vieira E, Brito D,
Peralta JP, Godinho R, Conceiçao P, Reis M, Rabaça C and Sismeiro
A: Influence of sociodemographic factors on treatment's choice for
localized prostate cancer in Portugal. Arch Ital Urol Androl.
92:45–49. 2020.
|
|
5
|
van Brussel JP and Mickisch GH: Multidrug
resistance in prostate cancer. Onkologie. 26:175–181.
2003.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Sturge J, Caley MP and Waxman J: Bone
metastasis in prostate cancer: Emerging therapeutic strategies. Nat
Rev Clin Oncol. 8:357–368. 2011.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Chi KN, Bjartell A, Dearnaley D, Saad F,
Schröder FH, Sternberg C, Tombal B and Visakorpi T:
Castration-resistant prostate cancer: From new pathophysiology to
new treatment targets. Eur Urol. 56:594–605. 2009.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Cookson MS, Lowrance WT, Murad MH and
Kibel AS: Castration-resistant prostate cancer: AUA guideline
amendment. J Urol. 193:491–499. 2015.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Tannock IF, de Wit R, Berry WR, Horti J,
Pluzanska A, Chi KN, Oudard S, Théodore C, James ND, Turesson I, et
al: Docetaxel plus prednisone or mitoxantrone plus prednisone for
advanced prostate cancer. N Engl J Med. 351:1502–1512.
2004.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Bolten CJ, Heinzle E, Müller R and
Wittmann C: Investigation of the central carbon metabolism of
Sorangium cellulosum: Metabolic network reconstruction and
quantification of pathway fluxes. J Microbiol Biotechnol. 19:23–36.
2009.PubMed/NCBI
|
|
11
|
Tyszka-Czochara M, Konieczny P and Majka
M: Recent advances in the role of AMP-activated protein kinase in
metabolic reprogramming of metastatic cancer cells: Targeting
cellular bioenergetics and biosynthetic pathways for anti-tumor
treatment. J Physiol Pharmacol. 69:2018.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Liberti MV and Locasale JW: The warburg
effect: How does it benefit cancer cells? Trends Biochem Sci.
41:211–218. 2016.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Chaneton B and Gottlieb E: Rocking cell
metabolism: Revised functions of the key glycolytic regulator PKM2
in cancer. Trends Biochem Sci. 37:309–316. 2012.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Wang J, Wang H, Liu A, Fang C, Hao J and
Wang Z: Lactate dehydrogenase A negatively regulated by miRNAs
promotes aerobic glycolysis and is increased in colorectal cancer.
Oncotarget. 6:19456–19468. 2015.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Ha TK, Her NG, Lee MG, Ryu BK, Lee JH, Han
J, Jeong SI, Kang MJ, Kim NH, Kim HJ and Chi SG: Caveolin-1
increases aerobic glycolysis in colorectal cancers by stimulating
HMGA1-mediated GLUT3 transcription. Cancer Res. 72:4097–4109.
2012.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Xu X, Li J, Sun X, Guo Y, Chu D, Wei L, Li
X, Yang G, Liu X, Yao L, et al: Tumor suppressor NDRG2 inhibits
glycolysis and glutaminolysis in colorectal cancer cells by
repressing c-Myc expression. Oncotarget. 6:26161–26176.
2015.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Dong G, Mao Q, Xia W, Xu Y, Wang J, Xu L
and Jiang F: PKM2 and cancer: The function of PKM2 beyond
glycolysis. Oncol Lett. 11:1980–1986. 2016.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Liang J, Cao R, Zhang Y, Xia Y, Zheng Y,
Li X, Wang L, Yang W and Lu Z: PKM2 dephosphorylation by Cdc25A
promotes the Warburg effect and tumorigenesis. Nat Commun.
7(12431)2016.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Luan W, Wang Y, Chen X, Shi Y, Wang J,
Zhang J, Qian J, Li R, Tao T, Wei W, et al: PKM2 promotes glucose
metabolism and cell growth in gliomas through a mechanism involving
a let-7a/c-Myc/hnRNPA1 feedback loop. Oncotarget. 6:13006–130018.
2015.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Lu W, Cao Y, Zhang Y, Li S, Gao J, Wang
XA, Mu J, Hu YP, Jiang L, Dong P, et al: Up-regulation of PKM2
promote malignancy and related to adverse prognostic risk factor in
human gallbladder cancer. Sci Rep. 6(26351)2016.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Zhang HS, Zhang FJ, Li H, Liu Y, Du GY and
Huang YH: Tanshinone ⅡA inhibits human esophageal cancer cell
growth through miR-122-mediated PKM2 down-regulation. Arch Biochem
Biophys. 598:50–56. 2016.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Xu Q, Zhang M, Tu J, Pang L, Cai W and Liu
X: MicroRNA-122 affects cell aggressiveness and apoptosis by
targeting PKM2 in human hepatocellular carcinoma. Oncol Rep.
34:2054–2064. 2015.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Guo M, Zhao X, Yuan X, Jiang J and Li P:
MiR-let-7a inhibits cell proliferation, migration, and invasion by
down-regulating PKM2 in cervical cancer. Oncotarget. 8:28226–28236.
2017.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Taniguchi K, Sugito N, Kumazaki M,
Shinohara H, Yamada N, Nakagawa Y, Ito Y, Otsuki Y, Uno B, Uchiyama
K and Akao Y: MicroRNA-124 inhibits cancer cell growth through
PTB1/PKM1/PKM2 feedback cascade in colorectal cancer. Cancer Lett.
363:17–27. 2015.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297.
2004.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Rigoutsos I: New tricks for animal
microRNAS: Targeting of amino acid coding regions at conserved and
nonconserved sites. Cancer Res. 69:3245–3248. 2009.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Gandellini P, Profumo V, Casamichele A,
Fenderico N, Borrelli S, Petrovich G, Santilli G, Callari M,
Colecchia M, Pozzi S, et al: miR-205 regulates basement membrane
deposition in human prostate: Implications for cancer development.
Cell Death Differ. 19:1750–1760. 2012.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Hudson RS, Yi M, Esposito D, Glynn SA,
Starks AM, Yang Y, Schetter AJ, Watkins SK, Hurwitz AA, Dorsey TH,
et al: MicroRNA-106b-25 cluster expression is associated with early
disease recurrence and targets caspase-7 and focal adhesion in
human prostate cancer. Oncogene. 32:4139–4147. 2013.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Boll K, Reiche K, Kasack K, Mörbt N,
Kretzschmar AK, Tomm JM, Verhaegh G, Schalken J, von Bergen M, Horn
F and Hackermüller J: MiR-130a, miR-203 and miR-205 jointly repress
key oncogenic pathways and are downregulated in prostate carcinoma.
Oncogene. 32:277–285. 2013.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Martens-Uzunova ES, Jalava SE, Dits NF,
van Leenders GJ, Møller S, Trapman J, Bangma CH, Litman T,
Visakorpi T and Jenster G: Diagnostic and prognostic signatures
from the small non-coding RNA transcriptome in prostate cancer.
Oncogene. 31:978–991. 2011.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Musumeci M, Coppola V, Addario A, Patrizii
M, Maugeri-Saccà M, Memeo L, Colarossi C, Francescangeli F, Biffoni
M, Collura D, et al: Control of tumor and microenvironment
cross-talk by miR-15a and miR-16 in prostate cancer. Oncogene.
30:4231–4242. 2011.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Takayama K, Tsutsumi S, Katayama S,
Okayama T, Horie-Inoue K, Ikeda K, Urano T, Kawazu C, Hasegawa A,
Ikeo K, et al: Integration of cap analysis of gene expression and
chromatin immunoprecipitation analysis on array reveals genome-wide
androgen receptor signaling in prostate cancer cells. Oncogene.
30:619–630. 2011.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Wang X, Zhang H, Bai M, Ning T, Ge S, Deng
T, Liu R, Zhang L, Ying G and Ba Y: Exosomes serve as nanoparticles
to deliver anti-miR-214 to reverse chemoresistance to cisplatin in
gastric cancer. Mol Ther. 26:774–783. 2018.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Pan C, Wang X, Shi K, Zheng Y, Li J, Chen
Y, Jin L and Pan Z: MiR-122 reverses the doxorubicin-resistance in
hepatocellular carcinoma cells through regulating the tumor
metabolism. PLoS One. 11(e0152090)2016.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Kishikawa T, Otsuka M, Tan PS Ohno M, Sun
X, Yoshikawa T, Shibata C, Takata A, Kojima K, Takehana K, et al:
Decreased miR122 in hepatocellular carcinoma leads to
chemoresistance with increased arginine. Oncotarget. 6:8339–8252.
2015.PubMed/NCBI View Article : Google Scholar
|
|
36
|
He J, Xie G, Tong J, Peng Y, Huang H, Li
J, Wang N and Liang H: Overexpression of microRNA-122 re-sensitizes
5-FU-resistant colon cancer cells to 5-FU through the inhibition of
PKM2 in vitro and in vivo. Cell Biochem Biophys. 70:1343–1350.
2014.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Fujita Y, Kojima K, Hamada N, Ohhashi R,
Akao Y, Nozawa Y, Deguchi T and Ito M: Effects of miR-34a on cell
growth and chemoresistance in prostate cancer PC3 cells. Biochem
Biophys Res Commun. 377:114–119. 2008.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Singh S, Chitkara D, Mehrazin R, Behrman
SW, Wake RW and Mahato RI: Chemoresistance in prostate cancer cells
is regulated by miRNAs and Hedgehog pathway. PLoS One.
7(e40021)2012.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Li J, Yang X, Guan H, Mizokami A, Keller
ET, Xu X, Liu X, Tan J, Hu L, Lu Y and Zhang J: Exosome-derived
microRNAs contribute to prostate cancer chemoresistance. Int J
Oncol. 49:838–846. 2016.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Shi G, Ye D, Yao X, Zhang SL, Dai B, Zhang
HL, Shen YJ, Zhu Y, Zhu YP, Xiao WJ and Ma CG: Involvement of
microRNA-21 in mediating chemo-resistance to docetaxel in
androgen-independent prostate cancer PC3 cells. Acta Pharmacol Sin.
31:867–873. 2010.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2−ΔΔCT method. Methods.
25:402–408. 2001.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Egawa T, Kubota T, Suto A, Otani Y,
Furukawa T, Watanabe M, Kumai K and Kitajima M: Docetaxel enhances
the cytotoxicity of anthracyclines by increasing intracellular drug
accumulation. Oncol Rep. 9:777–781. 2002.PubMed/NCBI
|
|
43
|
Lin W, Miao Y, Meng X, Huang Y, Zhao W and
Ruan J: miRNA-765 mediates multidrug resistance via targeting BATF2
in gastric cancer cells. FEBS Open Bio: Mar 12, 2020 (Epub ahead of
print).
|
|
44
|
Hong ST, Lin H, Wang CS, Chang CH, Lin
AMY, Yang JCH and Lo YL: Improving the anticancer effect of
afatinib and microRNA by using lipid polymeric nanoparticles
conjugated with dual pH-responsive and targeting peptides. J
Nanobiotechnology. 17(89)2019.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Hong BS, Ryu HS, Kim N, Kim J, Lee E, Moon
H, Kim KH, Jin MS, Kwon NH, Kim S, et al: Tumor Suppressor
miRNA-204-5p regulates growth, metastasis, and immune
microenvironment remodeling in breast cancer. Cancer Res.
79:1520–1534. 2019.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Xu B, Niu X, Zhang X, Tao J, Wu D, Wang Z,
Li P, Zhang W, Wu H, Feng N, et al: miR-143 decreases prostate
cancer cells proliferation and migration and enhances their
sensitivity to docetaxel through suppression of KRAS. Mol Cell
Biochem. 350:207–213. 2011.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Bian Z, Li L, Tang R, Hou DX, Chen X,
Zhang CY and Zen K: Identification of mouse liver
mitochondria-associated miRNAs and their potential biological
functions. Cell Res. 20:1076–1078. 2010.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Liu C, Zhang L, Huang Y, Lu K, Tao T, Chen
S, Zhang X, Guan H, Chen M and Xu B: MicroRNA-328 directly targets
p21-activated protein kinase 6 inhibiting prostate cancer
proliferation and enhancing docetaxel sensitivity. Mol Med Rep.
12:7389–7395. 2015.PubMed/NCBI View Article : Google Scholar
|
|
49
|
Shi HS, Li D, Zhang J, Wang YS, Yang L,
Zhang HL, Wang XH, Mu B, Wang W, Ma Y, et al: Silencing of pkm2
increases the efficacy of docetaxel in human lung cancer xenografts
in mice. Cancer Sci. 101:1447–1453. 2010.PubMed/NCBI View Article : Google Scholar
|