Introduction
Osteosarcoma (OS) is the most common malignant tumor
in orthopedics and has been indicated to occur predominantly in
adolescents between 1-20 years of age (1,2). OS is
characterized by high malignancy, invasion and early lung
metastasis (3,4). Therefore, the prognosis of patients
with OS remains poor, and the 5-year survival rate following
amputation has been reported to be only 5-10% (5,6). At
present, the treatment of OS primarily includes amputation,
radiotherapy and chemotherapy, which have been indicated to improve
the survival rate of patients with OS (7,8).
Although considerable progress has been achieved in the treatment
and prognosis of OS, novel diagnostic biomarkers and treatments are
required.
MicroRNAs (miRNAs/miRs) are a class of small
endogenous non-coding RNA molecules with 15-25 nucleotides in
length (9,10). The first miRNA has been discovered
in nematodes (11). miRNAs bind to
the 3'-untranslated region (3'-UTR) of target mRNAs, and
subsequently inhibit or induce the degradation of mRNAs to achieve
their biological function (12,13).
Accumulating evidence has demonstrated that miRNAs may act as tumor
suppressors or oncogenes, thus serving an important role in the
occurrence and development of various types of cancers (14-16).
miR-384 has been reported to inhibit the proliferation and invasion
of OS via regulating insulin-like growth factor binding protein
3(17). Moreover, Liu et al
(18) reported that miR-200a
induced immunosuppression via activating phosphatase and tensin
homolog (PTEN).
miR-208a has been indicated to be associated with
the development of numerous types of cancer. Cui et al
(19) reported that compared with
miR-negative control (NC), miR-208a enhanced cell proliferation and
invasion in gastric cancer via targeting secreted frizzled-related
protein 1 (SFRP1) and negatively regulating maternally expressed
gene 3. Moreover, Tang et al (20) revealed that the level of miR-208a
increased in the serum of lung cancer patients following
radiotherapy, and indicated that miR-208a increased the
proliferation and radioresistance of human lung cancer cells via
targeting p21. In another study, Zou et al (21) indicated that circRAD18 interacted
with miR-208a and miR-3164 to promote triple-negative breast cancer
progression via regulating the expression of insulin-like growth
factor 1 and fibroblast growth factor 2. However, to the best of
our knowledge, the effect of miR-208a-3p in OS has not been
investigated. Therefore, the present study aimed to investigate
whether miR-208a-3p may affect the progression of OS.
Materials and methods
Clinical samples
OS tissue specimens (n=10) and adjacent normal
tissues were obtained from patients with OS, including 4 females
(aged between 19 and 23 years), 5 males (aged between 18 and 25
years) and a child (aged 12 years), who did not undergo
chemotherapy or radiotherapy prior to operation at The First
Affiliated Hospital of Harbin Medical University, Harbin, China,
between September 2017 and December 2018. All tissues were kept in
liquid nitrogen immediately after resection operation. All patients
or their legal guardians provided written informed consent and the
study was approved by the Medical Ethics Committee of The First
Affiliated Hospital of Harbin Medical University.
Cell culture and transfection
The human OS cell lines SaOS-2, U2OS and MG-63 were
purchased from Shanghai Zhongqiao Xinzhou Biotechnology Co., Ltd.
Cells were maintained in DMEM (Thermo Fisher Scientific, Inc.)
supplemented with 10% FBS (Biological Industries) and 1%
penicillin/streptomycin (Sigma-Aldrich; Merck KGaA) at 37˚C in a
humidified atmosphere with 5% CO2. The human
osteoblastic cell line (hFOB 1.19) was purchased from Shanghai Yubo
Biotechnology Co., Ltd. The cells were maintained in DMEM medium,
supplemented with 10% FBS, 1% penicillin/streptomycin and 0.3 mg/ml
neomycin G418 (Stemcell Technologies, Inc.) at 34˚C in a humidified
atmosphere with 5% CO2. 293T cells were maintained in
DMEM supplemented with 10% FBS and 1% penicillin/streptomycin at
37˚C in a humidified atmosphere with 5% CO2. miR-208a-3p
mimics, NC mimics and miR-208a-3p and NC inhibitors were
synthesized by Suzhou GenePharma Co., Ltd. The sequences used were
as follows: miR-208a-3p mimics, 5'-AUAAGACGAGCAAAAAGCU UGU-3' and
5'-AGCUUUUUGCUCGUCUUAUUU-3'; miR-208a-3p inhibitor,
5'-ACAAGCUUUUUGCUCGUCU UAU-3'; NC mimics,
5'-UUCUCCGAACGUGUCACGUTT-3' and 5'-ACGUGACACGUUCGGAGAATT-3' and NC
inhibitor, 5'-CAGUACUUUUGUGUAGUACAA-3'. When SaOS-2, U2OS and MG-63
cells reached 70-80% confluence, they were transfected with
miR-208a-3p or mimics NC (50 nM each) and miR-208a-3p or inhibitor
NC (100 nM each) using Lipofectamine® 2000 (Invitrogen;
Thermo Fisher Scientific, Inc.) according to the manufacturer's
instructions. After 24 h, the transfected cells were used for
subsequent experimentations.
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA from OS tissues, adjacent normal tissues
and cell lines (SaOS-2, U2OS and MG-63 cells) was extracted using
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.) according to the manufacturer's protocol. Subsequently,
complementary DNA (cDNA) was synthesized using the High-Capacity
cDNA Reverse Transcription kit (Applied Biosystems; Thermo Fisher
Scientific, Inc.) according to the manufacturer's instructions. The
thermocycling conditions were as follows: 42˚C for 15 min, 95˚C for
5 min and 4˚C for 10 min. PCR amplification was performed on an ABI
7500 Real-Time PCR system (Applied Biosystems; Thermo Fisher
Scientific, Inc.) using SYBR Green Master kit (Roche Diagnostics).
Relative expression levels were calculated using the
2-ΔΔCq method (22), U6
and GAPDH served as an internal reference. The sequences of the
primers used in the present study were as follows: miR-208a-3p
sense, 5'-CGGGGCATAAGACGAGCAA AAA-3' and antisense,
5'-ATCCAGTGCAGGGTCCGAGG-3'; PTEN sense,
5'-CCAGAGACAAAAAGGGAGTAACTA-3' and antisense,
5'-ACCTTTAGCTGGCAGACC-3'; U6 sense, 5'-CTCGCTTCGGCAGCACA-3' and
antisense, 5'-AACGCTT CACGAATTTGCGT-3'; and GAPDH sense, 5'-CATCAC
TGCCACCCAGAAGAC-3' and antisense, 5'-CCAGTGAGC TTCCCGTTCAG-3'. The
following thermocycling conditions were used for the qPCR: 95˚C for
30 sec, followed by 40 cycles of amplification at 95˚C for 5 sec,
59˚C for 30 sec and 72˚C for 30 sec.
Cell proliferation assay
Cell proliferation was assessed using an MTT assay.
SaOS-2, U2OS and MG-63 cells transfected with miR-208a-3p mimics,
inhibitor or respective NC were seeded into 96-well plates at a
density of 5x103 cells/well. Following cell culture for
12, 24, 36 and 48 h, the medium was removed and 100 µl MTT solution
(500 µg/ml; Biosharp Life Sciences) was added into each well.
Subsequently, the 96-well plates were incubated for 4 h at 37˚C,
and the purple formazan was dissolved in 150 µl DMSO. The
absorbance of each well was measured at a wavelength of 490 nm
using a microplate reader (Tecan Group, Ltd.). The optical density
value of each well indicated the OS cells proliferation rate.
Colony formation assay
SaOS-2, U2OS and MG-63 cells transfected with
miR-208a-3p and NC mimics and inhibitors were seeded into 6-well
plates at a concentration of 2x103 cells/well. The cells
were cultured at 37˚C for 2 weeks and the medium was replaced every
three days. The formed colonies were subsequently fixed with 4%
polyformaldehyde for 20 min at room temperature and stained with
0.5% crystal violet (Sigma-Aldrich; Merck KGaA) for 10 min at room
temperature. Images of the newly formed colonies were captured
using a light microscope (magnification, x1). The experiments were
repeated at least three times.
Wound healing assay
SaOS-2, U2OS and MG-63 cells were cultured in 6-well
plates (5x105 cells/well). Once the cells grew to ~90%
confluence, wounds were created by scraping the cellular monolayer
with a 200-µl pipette tip. The cells were subsequently washed with
PBS to remove the suspended cells, and the culture medium was
replaced with serum-free DMEM medium. Images were captured at 0,
12, 24, 36 and 48 h following wound formation using an optical
microscope (magnification, x40) (Eclipse TS100; Nikon Corporation).
The wound widths were analyzed using ImageJ v1.8.0 software
(National Institutes of Health).
Invasion assay
The invasion assay was performed in a Transwell
chamber coated with Matrigel (pore size, 8 µm; BD Biosciences).
Briefly, Matrigel was diluted in serum-free DMEM medium at a 1:5
ratio and added to the upper chamber of the Transwell inserts and
maintained at 37˚C for 5 h. A total of 5x104 cells were
suspended in 200 µl serum-free medium and seeded into the upper
chamber of 24-well inserts, while the lower chamber was filled with
500 µl 10% FBS-containing medium. Following cell culture for 24 h,
the invaded cells were stained with 0.1% crystal violet solution
(Sigma-Aldrich; Merck KGaA) at room temperature for 20 min. The
number of cells invaded through the pore was examined using an
optical microscope (magnification, x100).
Western blot analysis
Total protein extracts were isolated from MG-63
cells using ice-cold RIPA lysis buffer (Beyotime Institute of
Biotechnology) supplemented with protease inhibitors. Total protein
concentration was determined using a bicinchoninic acid protein
assay kit (Beyotime Institute of Biotechnology) according to the
manufacturer's instructions. Equal amounts (40 µg/lane) of protein
from each experimental group were denatured, separated using 10%
SDS-PAGE and then transferred onto a nitrocellulose membrane (EMD
Millipore). Following blocking with 5% skimmed milk for 2 h at room
temperature, the membranes were incubated overnight at 4˚C with the
following primary antibodies: Anti-PTEN (1:1,000; cat. no. ab32199;
Abcam) and anti-GAPDH (1:1,000; cat. no. ab9482; Abcam). After
rinsing, the membranes were incubated with an anti-mouse secondary
immunoglobulin G (IgG; 1:5,000; cat. no. sc-516102; Santa Cruz
Biotechnology, Inc) or anti-rabbit secondary IgG (1:5,000; cat. no.
sc-2357; Santa Cruz Biotechnology, Inc.) for 1 h at room
temperature. Finally, immunoreactive bands were visualized using an
Odyssey® CLx Imager and quantified with LI-COR Image
Studio v5.2.5 Software (LI-COR Biosciences). GAPDH served as the
internal control.
Dual-luciferase reporter assay
The 3'-UTR region of PTEN encompassing the putative
miR-208a-3p binding site was amplified using PCR and was
subsequently sub-cloned into the psiCHECK-2 dual luciferase vector
(Promega Corporation). 293T cells at a density of 2x104
cells/well were cultured in 24-well plates and co-transfected with
the luciferase vector encompassing the 3'-UTR of PTEN and
miR-208a-3p or NC mimics using Lipofectamine® 2000
(Invitrogen; Thermo Fisher Scientific, Inc.). Following 48 h,
luciferase activity was determined using a
Dual-Luciferase® Reporter assay kit (Promega
Corporation) according to the manufacturer's instructions. Firefly
luciferase activity was normalized to the corresponding
Renilla luciferase activity.
Bioinformatic analysis
The potential targets of miR-208a-3p were predicted
via TargetScan software v7.2 (http://www.targetscan.org/vert_72/) using the default
settings.
Statistical analysis
All quantitative experimental data are presented as
mean ± standard error of the mean. Statistical analyses were
performed using GraphPad Prism v7.0 (GraphPad Software, Inc.). The
statistical differences among experimental groups were evaluated
using unpaired Student's t-test or one-way ANOVA followed by
Tukey's post hoc test. P<0.05 was considered to indicate a
statistically significant difference. Each experiment was repeated
at least three times.
Results
miR-208a-3p is upregulated in OS
tissues and cell lines
It has been reported that miR-208a-3p was
dysregulated and promoted the development of several types of
cancer, including gastric and colon cancer (23,24).
The present study aimed to investigate the role of miR-208a-3p in
OS. Therefore, RT-qPCR was performed to determine the expression
levels of miR-208a-3p in 10 pairs of tissues from patients with OS
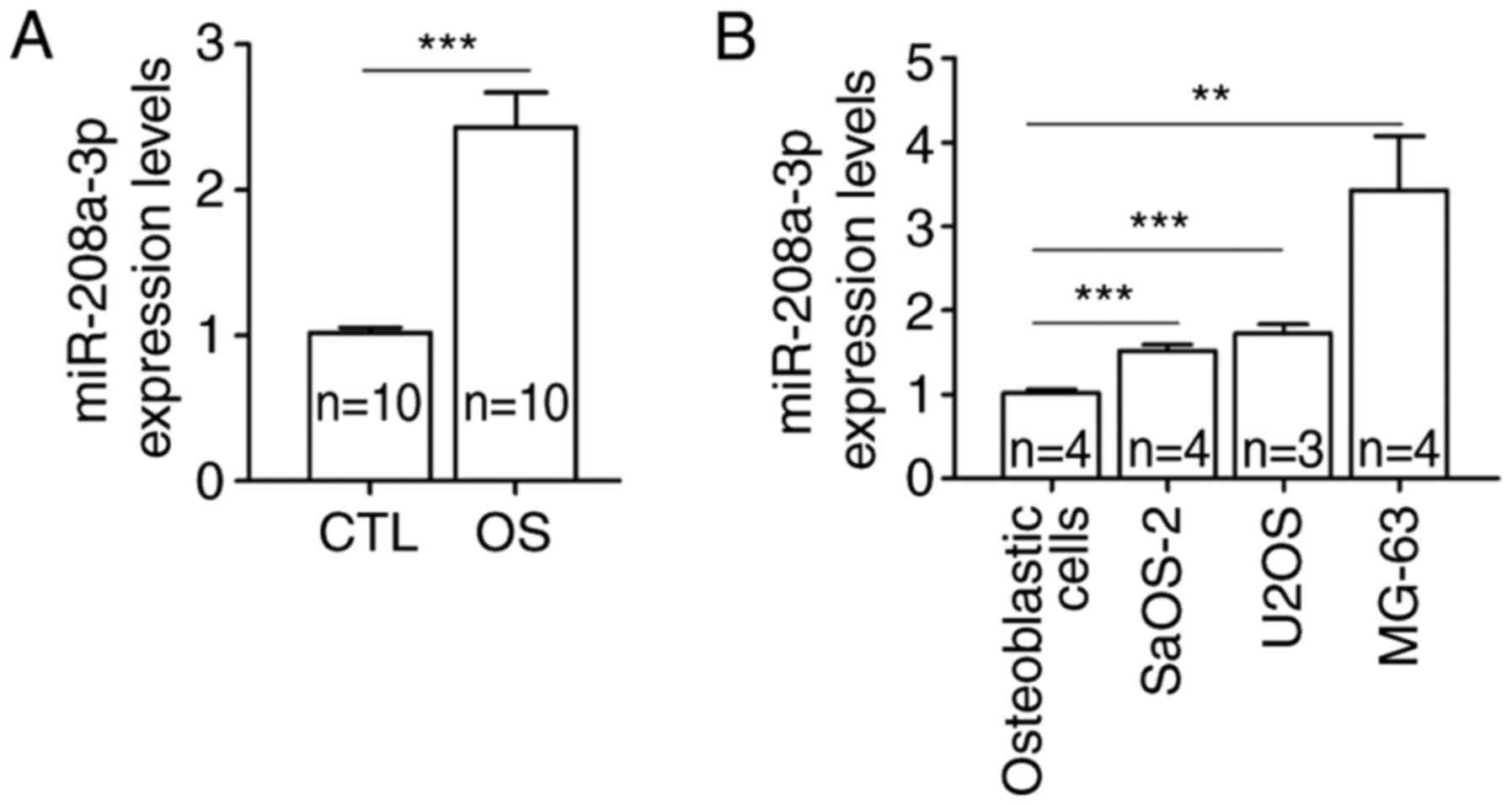
and adjacent normal tissues. The results showed that the expression
levels of miR-208a-3p were significantly higher in OS tissues
compared with adjacent normal tissues (Fig. 1A). Similarly, miR-208a-3p was
significantly upregulated in OS cell lines, namely SaOS-2, U2OS and
MG-63, compared with human osteoblastic cells (Fig. 1B).
Overexpression of miR-208a-3p promotes
the proliferation of OS cell lines
To verify the role of miR-208a-3p in OS cells,
SaOS-2, U2OS and MG-63 cells were transfected with miR-208a-3p
mimics to artificially alter the endogenous levels of miR-208a-3p
(Fig. S1). Following transfection,
the proliferation rate of OS cells was determined using MTT and
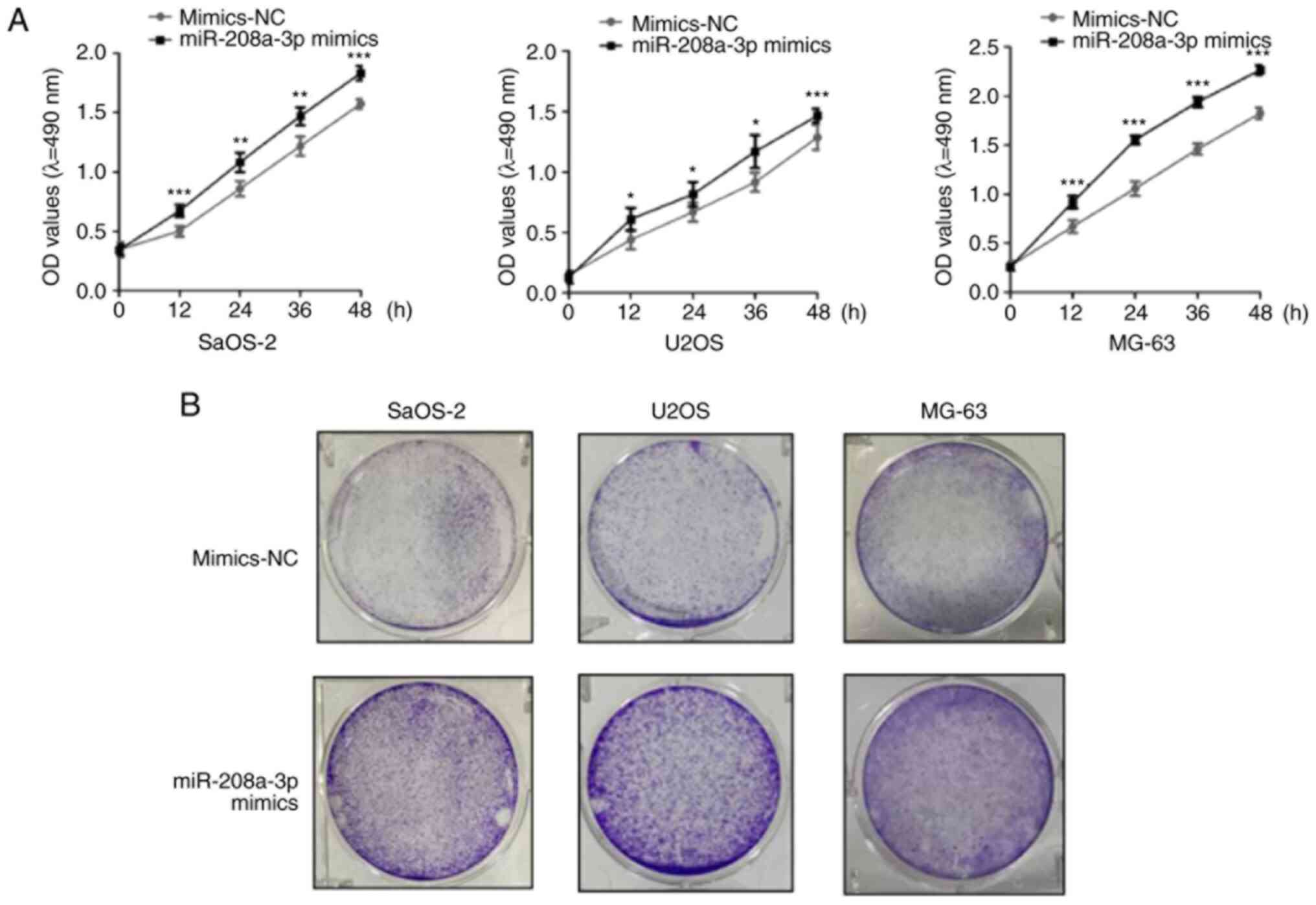
colony formation assays. More specifically, the MTT assays
demonstrated that compared with the mimics NC group, overexpression
of miR-208a-3p significantly promoted the proliferation of SaOS-2,
U2OS and MG-63 cells (Fig. 2A).
Moreover, the results of the colony formation assays were
consistent with the MTT assay findings, as miR-208a-3p
overexpression was indicated to increase colony formation by
SaOS-2, U2OS and MG-63 cells compared with the mimics NC group
(Fig. 2B). Overall, these data
suggested that overexpression of miR-208a-3p increased the
proliferation rate of OS cells.
Overexpression of miR-208a-3p promotes
the invasion and migration of OS cells
To determine the association between miR-208a-3p
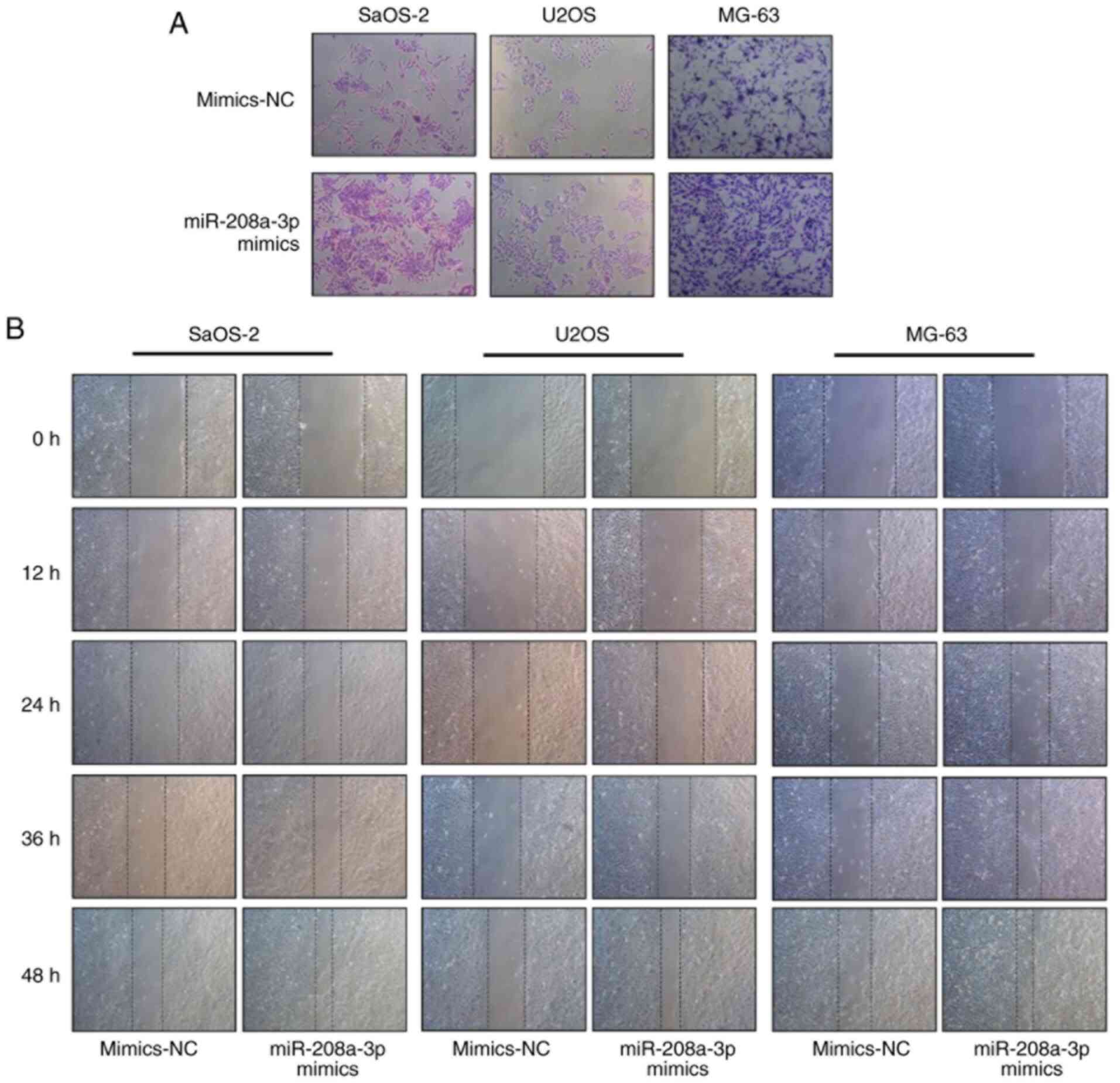
with the invasion of OS cells, Transwell assays were performed. The
results revealed that the number of invading OS cells was higher in
the miR-208a-3p mimics group compared with the mimics NC group
(Fig. 3A). Furthermore, wound
healing assays demonstrated that miR-208a-3p overexpression
improved the migratory ability of SaOS-2, U2OS and MG-63 cells at
12, 24, 36 and 48 h following wound formation (Fig. 3B). These findings indicated that
overexpression of miR-208a-3p positively regulated the migratory
and invasive abilities of SaOS-2, U2OS and MG-63 cells.
Knockdown of miR-208a-3p inhibits the
proliferation of OS cells
The aforementioned results indicated that the
overexpression of miR-208a-3p promoted the proliferation, invasion
and migration of OS cells; however, whether miR-208a-3p knockdown
can induce an opposite phenotype in OS cells remains unclear.
Therefore, additional studies were performed. OS cells were
initially transfected with a miR-208a-3p inhibitor to silence
miR-208a-3p expression (Fig. S1).
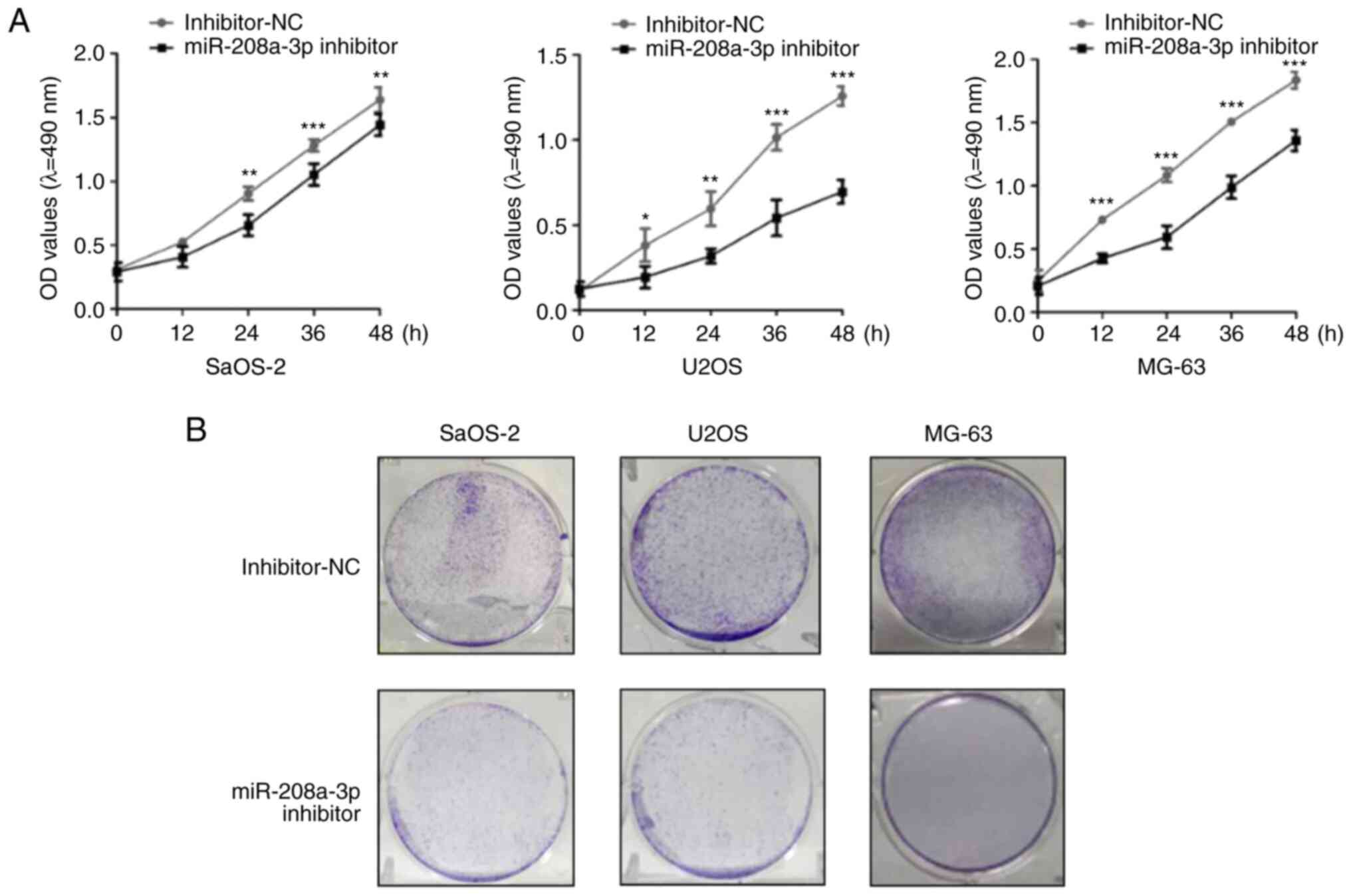
Following miR-208a-3p knockdown, MTT assays revealed that compared
with the inhibitor NC group, the proliferative ability of SaOS-2,
U2OS and MG-63 cells significantly decreased in the miR-208a-3p
inhibitor group (Fig. 4A).
Similarly, colony formation assays demonstrated that the colony
formation ability of OS cell lines in the miR-208a-3p inhibitor
group was reduced compared with the inhibitor NC group (Fig. 4B). These results indicated that
miR-208a-3p knockdown decreased the proliferative ability of
SaOS-2, U2OS and MG-63 cells.
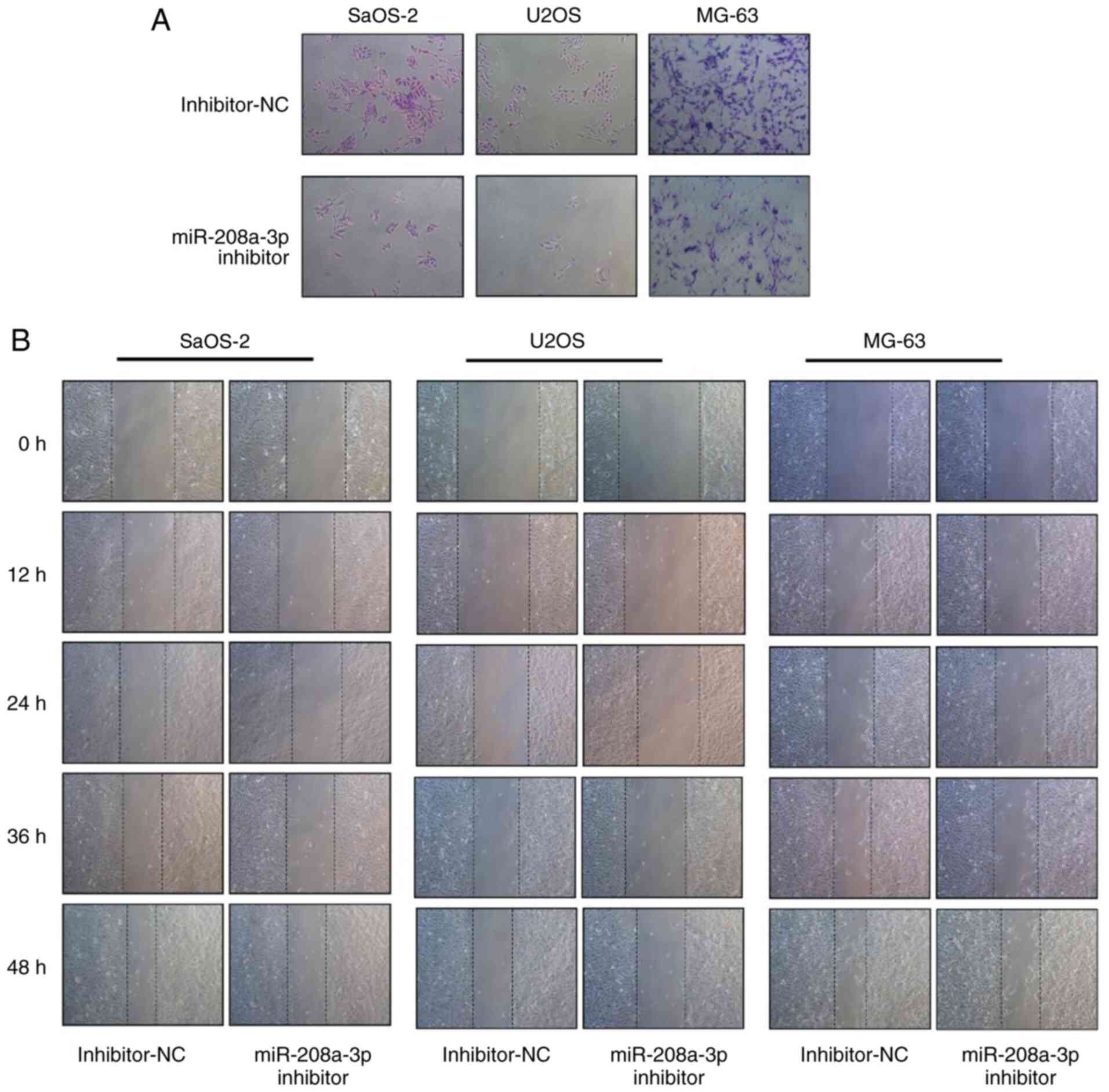
Knockdown of miR-208a-3p suppresses
the invasion and migration of OS cells
The effect of miR-208a-3p knockdown in the migratory
and invasive ability of OS cells was investigated using wound
healing and Transwell assays, respectively. The results of the
invasion assays indicated that miR-208a-3p knockdown decreased the
number of SaOS-2, U2OS and MG-63 cells that invaded through
Matrigel compared with the inhibitor NC group (Fig. 5A). This finding suggested that
miR-208a-3p knockdown reduced the invasive ability of SaOS-2, U2OS
and MG-63 cells. Additionally, the results of the wound healing
assays indicated that miR-208a-3p knockdown inhibited the migratory
ability of OS cells at 12, 24, 36 and 48 h following wound
formation (Fig. 5B).
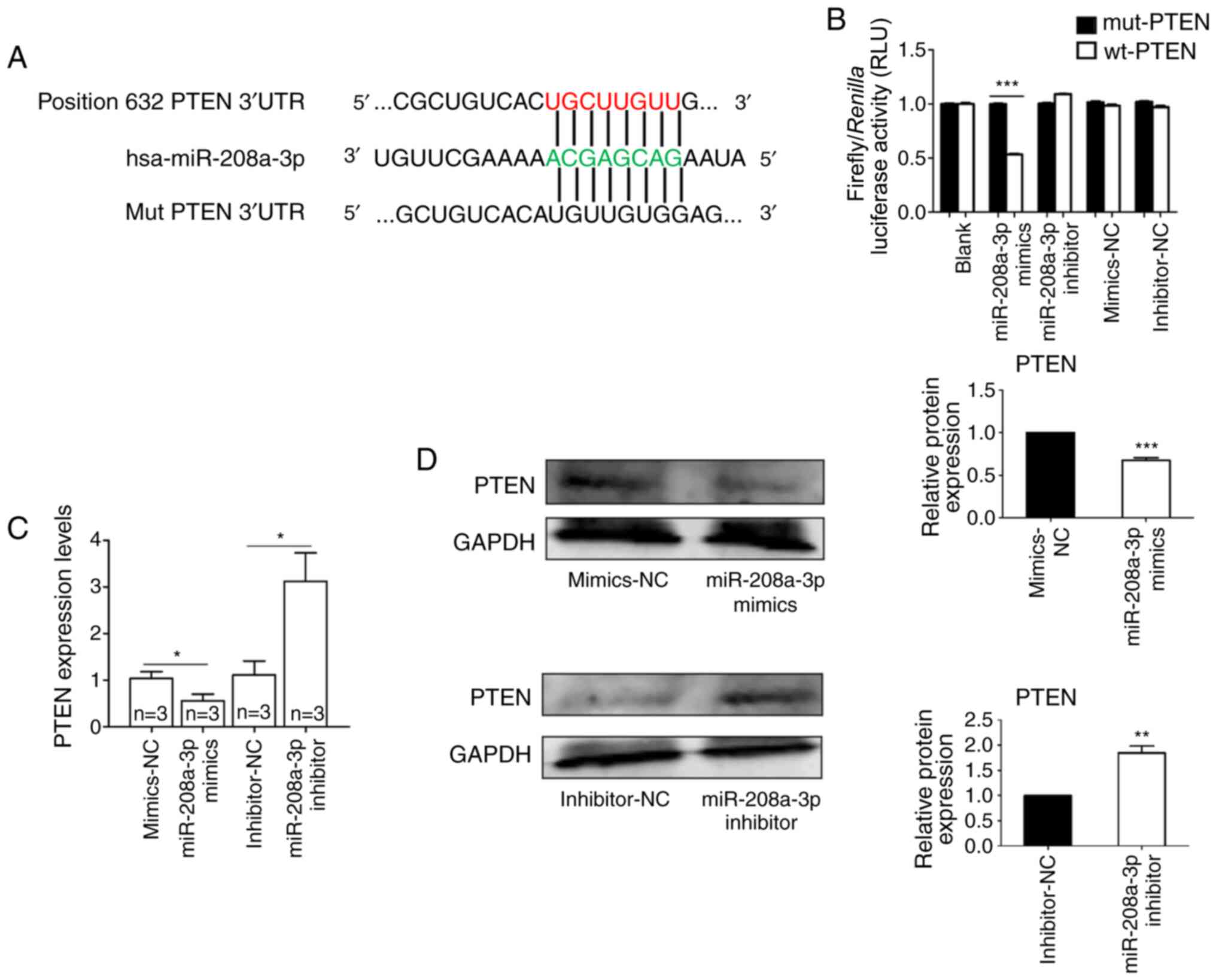
Direct targets of miR-208a-3p
To better characterize the underlying molecular
mechanisms associated with miR-208a-3p-mediated progression of OS,
bioinformatics analysis was performed. Therefore, TargetScan
software was used to predict the potential targets of miR-208a-3p.
The analysis revealed that the PTEN 3'-UTR encompassed a
complementary binding site for miR-208a-3p (Fig. 6A). Furthermore, a luciferase
reporter assay was employed to examine whether the 3'-UTR of PTEN
was a direct target of miR-208a-3p. The results demonstrated that
miR-208a-3p mimics significantly suppressed the luciferase activity
of the wild-type plasmid, but not that of the mutant plasmid
(Fig. 6B). In addition, RT-qPCR
analysis indicated that compared with their respective NC groups,
miR-208a-3p overexpression significantly suppressed the mRNA
expression of PTEN, and miR-208a-3p knockdown exhibited the
opposite effect (Fig. 6C).
Similarly, western blot analysis revealed that compared with their
respective NC groups, overexpression of miR-208a-3p significantly
downregulated the protein expression of PTEN, while miR-208a-3p
knockdown significantly increased PTEN protein levels (Fig. 6D). Taken together, these results
suggested that miR-208a-3p may regulate the progression of OS via
directly targeting PTEN.
Discussion
OS is a malignant bone tumor with a high incidence
in adolescents, while its prognosis is poor due to pulmonary
metastasis (25). Accumulating
evidence has indicated that miRNAs act as tumor suppressor genes or
oncogenes associated with the progression of OS (26,27).
For example, miR-138 has been reported to inhibit cell
proliferation and invasion, and promote cell apoptosis of human OS
cells via targeting differentiated embryonic chondrocyte gene
2(28). In addition, miR-33a has
been revealed to be upregulated in chemoresistant OS and promote OS
cell resistance to cisplatin via downregulating Twist-related
protein 1(29). Furthermore, it has
been demonstrated that exosomal miR-1228 from cancer-associated
fibroblasts promoted OS cell migration and invasion via directly
targeting suppressor of cancer cell invasion (30). The present study aimed to
investigate the role of miR-208a-3p in the regulation of OS
progression.
It has been identified that miR-208a-3p is a highly
conserved miRNA that serves a novel important regulator in heart
diseases by affecting autophagy and other processes (31-33).
miR-208a-3p has also been associated with the progression of
several types of cancer. For example, miR-208a-3p, as an oncogenic
miRNA, has been indicated to suppress apoptosis in gastric cancer
cells (23), and promote cell
proliferation, invasion and migration in colon cancer via targeting
programmed cell death protein 4(24). However, to the best of our
knowledge, the role of miR-208a-3p in OS has not yet been reported.
The results of the present study demonstrated that miR-208a-3p was
significantly increased in OS tissues (compared with adjacent
normal tissues) and three OS cell lines (compared with osteoblastic
cells). Furthermore, miR-208a-3p overexpression promoted cell
growth and proliferation and facilitated the invasion and migration
of SaOS-2, U2OS and MG-63 cells. By contrast, knockdown of
miR-208a-3p exhibited the opposite the effects compared with
miR-208a-3p mimics. To the best of our knowledge, the present study
was the first to demonstrate that miR-208a-3p may serve as an
oncogene via regulating cell growth, proliferation and metastasis
of OS cells, which was consistent with a previous report indicating
that miR-208a-3p as an oncogene promoted colon cancer progression
(24).
To further investigate the underlying mechanisms of
miR-208a-3p-mediated regulation of OS progression, the downstream
targets of miR-208a-3p were examined. Several genes have been
identified to be directly targeted by miR-208a-3p, including SFRP1,
cadherin 9 and thyroid hormone receptor associated protein 1
(19,34,35).
The present study verified that PTEN was a direct target of
miR-208a-3p in OS cells. Downregulation of PTEN, a well-known tumor
suppressor protein, may result in tumor progression (36,37).
As a tumor suppressor, PTEN has been associated with several
biological processes, including maintenance of genomic stability,
cell survival, migration, proliferation and metabolism (38). Recently, a number of studies
revealed that PTEN downregulation was associated with poor
prognosis in several types of cancers, including breast, prostate
and non-small cell lung cancers (39-41).
Moreover, loss of PTEN expression has been implicated in the
promotion of OS cell proliferation and metastasis (42). In the present study, bioinformatics
analysis and luciferase assays revealed that PTEN was a direct
target of miR-208a-3p. Subsequently, the detection of PTEN mRNA and
protein expression levels in OS cells following transfection with
miR-208a-3p mimics or inhibitors demonstrated that miR-208a-3p
mimics downregulated while miR-208a-3p inhibitor upregulated PTEN
expression.
In summary, the present study revealed that
miR-208a-3p was upregulated in OS tissues and cell lines, and
promoted cell proliferation, migration and invasion of OS cells via
targeting PTEN. Overall, these findings indicate that miR-208a-3p,
acting as an oncogene, promotes OS progression and may be
considered a diagnostic biomarker for OS.
Supplementary Material
Transfection efficiency of miR-208a-3p
mimics and inhibitor. The expression levels of miR-208a-3p in U2OS,
SaOS-2 and MG-63 cells following transfection with miR-208a-3p
mimics or inhibitor was detected via reverse
transcription-quantitative PCR. ***P<0.001 vs. NC
groups. NC, negative control; miR, microRNA.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
All datasets used and/or generated during the
present study are available from the corresponding author on
reasonable request.
Authors' contributions
YF and ZB contributed to study design, data
collection, statistical analysis, data interpretation and
manuscript preparation. YW, LY, KB, YS, YL and CF contributed to
data collection and statistical analysis. BL, ZL and FZ contributed
to experimental design, manuscript revision and data proofreading.
All authors read and approved the final manuscript.
Ethics approval and consent to
participate
The research was approved by the Medical Ethics
Committee of The First Affiliated Hospital of Harbin Medical
University (Harbin, China; approval no. 201516). Written informed
consent was obtained from all patients.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Bielack S, Carrle D and Casali PG: ESMO
Guidelines Working Group. Osteosarcoma: ESMO clinical
recommendations for diagnosis, treatment and follow-up. Ann Oncol.
20 (Suppl 4):137–139. 2009.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Camuzard O, Santucci-Darmanin S, Carle GF
and Pierrefite-Carle V: Role of autophagy in osteosarcoma. J Bone
Oncol. 16(100235)2019.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Yao Z, Han L, Chen Y, He F, Sun B, Kamar
S, Zhang Y, Yang Y, Wang C and Yang Z: Hedgehog signalling in the
tumourigenesis and metastasis of osteosarcoma, and its potential
value in the clinical therapy of osteosarcoma. Cell Death Dis.
9(701)2018.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Sasaki R, Osaki M and Okada F:
MicroRNA-Based Diagnosis and Treatment of Metastatic Human
Osteosarcoma. Cancers (Basel). 11(11)2019.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Anninga JK, Gelderblom H, Fiocco M, Kroep
JR, Taminiau AH, Hogendoorn PC and Egeler RM: Chemotherapeutic
adjuvant treatment for osteosarcoma: Where do we stand? Eur J
Cancer. 47:2431–2445. 2011.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Tang QX, Wang LC, Wang Y, Gao HD and Hou
ZL: Efficacy of methotrexate, doxorubicin, and cisplatin for
osteosarcoma: Study protocol for a systematic review of randomized
controlled trial. Medicine (Baltimore). 98(e14442)2019.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Pang KL and Chin KY: Emerging Anticancer
Potentials of Selenium on Osteosarcoma. Int J Mol Sci.
20(20)2019.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Carina V, Costa V, Sartori M, Bellavia D,
De Luca A, Raimondi L, Fini M and Giavaresi G: Adjuvant Biophysical
Therapies in Osteosarcoma. Cancers (Basel). 11(11)2019.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Remsburg C, Konrad K, Sampilo NF and Song
JL: Analysis of microRNA functions. Methods Cell Biol. 151:323–334.
2019.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Ichiyama K and Dong C: The role of miR-183
cluster in immunity. Cancer Lett. 443:108–114. 2019.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Akhtar MM, Micolucci L, Islam MS, Olivieri
F and Procopio AD: Bioinformatic tools for microRNA dissection.
Nucleic Acids Res. 44:24–44. 2016.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297.
2004.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Fabian MR and Sonenberg N: The mechanics
of miRNA-mediated gene silencing: A look under the hood of miRISC.
Nat Struct Mol Biol. 19:586–593. 2012.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Cheng WC, Liao TT, Lin CC, Yuan LE, Lan
HY, Lin HH, Teng HW, Chang HC, Lin CH, Yang CY, et al:
RAB27B-activated secretion of stem-like tumor exosomes delivers the
biomarker microRNA-146a-5p, which promotes tumorigenesis and
associates with an immunosuppressive tumor microenvironment in
colorectal cancer. Int J Cancer. 145:2209–2224. 2019.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Di Cosimo S, Appierto V, Pizzamiglio S,
Tiberio P, Iorio MV, Hilbers F, de Azambuja E, de la Peña L,
Izquierdo M, Huober J, et al: Plasma miRNA Levels for Predicting
Therapeutic Response to Neoadjuvant Treatment in HER2-positive
Breast Cancer: Results from the NeoALTTO Trial. Clin Cancer Res.
25:3887–3895. 2019.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Yang Y, Ishak Gabra MB, Hanse EA, Lowman
XH, Tran TQ, Li H, Milman N, Liu J, Reid MA, Locasale JW, et al:
miR-135 suppresses glycolysis and promotes pancreatic cancer cell
adaptation to metabolic stress by targeting phosphofructokinase-1.
Nat Commun. 10(809)2019.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Tan Y, Chen L, Li S, Hao H and Zhang D:
miR-384 Inhibits Malignant Biological Behavior Such as
Proliferation and Invasion of Osteosarcoma by Regulating IGFBP3.
Technol Cancer Res Treat. 19(1533033820909125)2020.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Liu Z, Wen J, Wu C, Hu C, Wang J, Bao Q,
Wang H, Wang J, Zhou Q, Wei L, et al: MicroRNA-200a induces
immunosuppression by promoting PTEN-mediated PD-L1 upregulation in
osteosarcoma. Aging (Albany NY). 12:1213–1236. 2020.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Cui HB, Ge HE, Wang YS and Bai XY:
miR-208a enhances cell proliferation and invasion of gastric cancer
by targeting SFRP1 and negatively regulating MEG3. Int J Biochem
Cell Biol. 102:31–39. 2018.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Tang Y, Cui Y, Li Z, Jiao Z, Zhang Y, He
Y, Chen G, Zhou Q, Wang W, Zhou X, et al: Erratum to:
Radiation-induced miR-208a increases the proliferation and
radioresistance by targeting p21 in human lung cancer cells. J Exp
Clin Cancer Res. 35(20)2016.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Zou Y, Zheng S, Xiao W and Xie X, Yang A,
Gao G, Xiong Z, Xue Z, Tang H and Xie X: circRAD18 sponges
miR-208a/3164 to promote triple-negative breast cancer progression
through regulating IGF1 and FGF2 expression. Carcinogenesis.
40:1469–1479. 2019.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) Method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Yin K, Liu M, Zhang M, Wang F, Fen M, Liu
Z, Yuan Y, Gao S, Yang L, Zhang W, et al: miR-208a-3p suppresses
cell apoptosis by targeting PDCD4 in gastric cancer. Oncotarget.
7:67321–67332. 2016.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Wu H, Xu L, Chen Y and Xu C: miR-208a-3p
functions as an oncogene in colorectal cancer by targeting PDCD4.
Biosci Rep. 39(39)2019.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Ahmed G, Zamzam M, Kamel A, Ahmed S,
Salama A, Zaki I, Kamal N and Elshafiey M: Effect of timing of
pulmonary metastasis occurrence on the outcome of metastasectomy in
osteosarcoma patients. J Pediatr Surg. 54:775–779. 2019.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Huang Y, Zhang J, Shao H, Liu J, Jin M,
Chen J and Zhao H: miR-33a Mediates the Anti-Tumor Effect of
Lovastatin in Osteosarcoma by Targeting CYR61. Cell Physiol
Biochem. 51:938–948. 2018.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Wang X, Peng L, Gong X, Zhang X, Sun R and
Du J: miR-423-5p Inhibits Osteosarcoma Proliferation and Invasion
Through Directly Targeting STMN1. Cell Physiol Biochem.
50:2249–2259. 2018.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Jiang B, Mu W, Wang J, Lu J, Jiang S, Li
L, Xu H and Tian H: MicroRNA-138 functions as a tumor suppressor in
osteosarcoma by targeting differentiated embryonic chondrocyte gene
2. J Exp Clin Cancer Res. 35(69)2016.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Zhou Y, Huang Z, Wu S, Zang X, Liu M and
Shi J: miR-33a is up-regulated in chemoresistant osteosarcoma and
promotes osteosarcoma cell resistance to cisplatin by
down-regulating TWIST. J Exp Clin Cancer Res. 33(12)2014.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Wang JW, Wu XF, Gu XJ and Jiang XH:
Exosomal miR-1228 From Cancer-Associated Fibroblasts Promotes Cell
Migration and Invasion of Osteosarcoma by Directly Targeting SCAI.
Oncol Res. 27:979–986. 2019.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Wang L, Ye N, Lian X, Peng F, Zhang H and
Gong H: miR-208a-3p aggravates autophagy through the PDCD4-ATG5
pathway in Ang II-induced H9c2 cardiomyoblasts. Biomed
Pharmacother. 98:1–8. 2018.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Li S, Jiang Z, Wen L, Feng G and Zhong G:
MicroRNA-208a-3p contributes to connexin40 remolding in human
chronic atrial fibrillation. Exp Ther Med. 14:5355–5362.
2017.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Cai B, Pan Z and Lu Y: The roles of
microRNAs in heart diseases: A novel important regulator. Curr Med
Chem. 17:407–411. 2010.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Zhang S, Zhang R, Wu F and Li X:
MicroRNA-208a Regulates H9c2 Cells Simulated Ischemia-Reperfusion
Myocardial Injury via Targeting CHD9 through Notch/NF-kappa B
Signal Pathways. Int Heart J. 59:580–588. 2018.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Callis TE, Pandya K, Seok HY, Tang RH,
Tatsuguchi M, Huang ZP, Chen JF, Deng Z, Gunn B, Shumate J, et al:
MicroRNA-208a is a regulator of cardiac hypertrophy and conduction
in mice. J Clin Invest. 119:2772–2786. 2009.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Moses C, Nugent F, Waryah CB, Garcia-Bloj
B, Harvey AR and Blancafort P: Activating PTEN Tumor Suppressor
Expression with the CRISPR/dCas9 System. Mol Ther Nucleic Acids.
14:287–300. 2019.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Tu J, Cheung HH, Lu G, Chen Z and Chan WY:
MicroRNA-10a promotes granulosa cells tumor development via
PTEN-AKT/Wnt regulatory axis. Cell Death Dis.
9(1076)2018.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Lee YR, Chen M and Pandolfi PP: The
functions and regulation of the PTEN tumour suppressor: New modes
and prospects. Nat Rev Mol Cell Biol. 19:547–562. 2018.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Patsouris A, Augereau P, Frenel JS, Robert
M, Gourmelon C, Bourbouloux E, Berton-Rigaud D, Chevalier LM and
Campone M: Benefits versus risk profile of buparlisib for the
treatment of breast cancer. Expert Opin Drug Saf. 18:553–562.
2019.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Abou-Ouf H, Ghosh S, Box A, Palanisamy N
and Bismar TA: Combined loss of TFF3 and PTEN is associated with
lethal outcome and overall survival in men with prostate cancer. J
Cancer Res Clin Oncol. 145:1751–1759. 2019.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Ling C, Wang X, Zhu J, Tang H, Du W, Zeng
Y, Sun L, Huang JA and Liu Z: MicroRNA-4286 promotes cell
proliferation, migration, and invasion via PTEN regulation of the
PI3K/Akt pathway in non-small cell lung cancer. Cancer Med.
8:3520–3531. 2019.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Yu C, Zhang B, Li YL and Yu XR: SIX1
reduces the expression of PTEN via activating PI3K/AKT signal to
promote cell proliferation and tumorigenesis in osteosarcoma.
Biomed Pharmacother. 105:10–17. 2018.PubMed/NCBI View Article : Google Scholar
|