Introduction
Breast cancer has one of the highest incidence rates
among all types of cancer, with patients also displaying high
mortality worldwide (1). In the
majority of cases, estrogen receptor, progesterone receptor and
human EGF-like receptor 2 signaling are important drivers of the
development of breast tumors (2-4).
Despite advances in the diagnosis and treatment of breast cancer,
including radical surgery and adjuvant therapy, end-stage survival
rates remain low due to aggressive clinical behavior (5,6).
Therefore, identifying novel therapeutic targets for breast cancer
is important.
Increasing evidence has indicated the potential role
of long non-coding RNAs (lncRNAs) as biomarkers and therapeutic
targets for solid tumors (7-10).
lncRNAs have been reported to participate in various epigenetic
regulatory processes (11,12), serve important roles in numerous
biological functions and are also aberrantly expressed in a variety
of tumors (13-18).
However, the relevance of aberrant lncRNA expression in the
biological, prognosis and molecular classification of human breast
cancer is not completely understood. The TMPO antisense RNA 1
(TMPO-AS1) gene, located on human chromosome 12, is a recently
identified lncRNA consisting of 3,161 nucleotides that has rarely
been reported in human diseases (19). Moreover, the functional role and
potential regulatory mechanisms underlying TMPO-AS1 in breast
cancer are not completely understood. Therefore, the present study
aimed to investigate the functions and regulatory molecular
mechanisms underlying TMPO-AS1 in breast cancer.
In the present study, the functional roles of lncRNA
TMPO-AS1 in human breast cancer were investigated, with a focus on
its underlying regulatory mechanisms. Collectively, the present
study suggested a novel regulatory mechanism, by which TMPO-AS1
promoted breast cancer progression, providing a new perspective for
the study of the molecular mechanisms underlying breast
cancer-associated lncRNAs.
Materials and methods
Ethics statement
The present study was approved by the Ethics
Committee of the Third Affiliated Hospital of Harbin Medical
University (Harbin, China, approval number: 20170529003). All
patients provided written in formed consent.
Clinical specimens and cell lines
Breast cancer tissues were collected from patients
with breast cancer at The Third Affiliated Hospital of Harbin
Medical University. These patients included 40 females; the age
range was from 27-64 years, with a mean age of 45.4±5.6 years. At
the same time, a total of 15 healthy controls, which included 15
females with an age range between 31-67 years, with a mean age of
48.2±6.9 years serve as the control group. Immediately after
resection, the primary and matched adjacent non-cancerous tissues
were frozen in liquid nitrogen. Human breast cancer cells and
MCF-10A epithelial cells were purchased from ATCC. MCF7, T47D,
MDA-MB-231 and SKBR3 cells were cultured in DMEM (Gibco; Thermo
Fisher Scientific, Inc.), supplemented with 10% FBS (Gibco; Thermo
Fisher Scientific, Inc.), 100 U/ml penicillin, and 100 mg/ml
streptomycin at 37˚C with 5% CO2. BT20 cells were
cultured in RPMI-1640 (Gibco; Thermo Fisher Scientific, Inc.)
supplemented with 10% FBS, 100 U/ml penicillin, and 100 mg/ml
streptomycin at 37˚C with 5% CO2. MCF-10A cells were
cultured in M-171 medium (Gibco; Thermo Fisher Scientific, Inc.)
supplemented with mammary epithelial growth factors (Invitrogen;
Thermo Fisher Scientific, Inc.) at 37˚C with 5% CO2.
RNA interference
A specific small interfering (si)RNA targeted
against TMPO-AS1 and the corresponding negative control (NC) siRNA
were purchased from Shanghai GenePharma Co., Ltd. The following
siRNAs were used: siTMPO-AS1 forward, 5'-GAGCCGAACUACGAACCAATT-3'
and reverse, 5'-UUGGUUCGUAGUUCGGCUCTT-3'; scrambled siRNA NC
forward, 5'-UUCUCCGAACGUGUCACGUTT-3' and reverse,
5'-ACGUGACACGUUCGGAGAATT-3'. miR-140-5p mimic (miR10000431-1-5,
5'-UGAGAACUGAAUUCCAUGGGUU-3'), mimic NC (miR1N0000001-1-5,
5'-UUCUCCGAACGUGUCACGUTT-3'), miR-140-5p inhibitor
(miR20000431-1-5, 5'-AACCCAUGGAAUUCAGUUCUCA-3') and inhibitor NC
(miR2N0000001-1-5, 5'-UCUACUCUUUCUAGGAGGUUGUGA-3') were obtained
from Guangzhou RiboBio Co., Ltd. 1x106 cells were
transfected with 50 pg/µl siRNA, miRNA mimic, miRNA inhibitor or
corresponding NCs using Lipofectamine® 2000 (Thermo
Fisher Scientific, Inc.) and harvested 48-72 h after
transfection.
To establish stable TMPO-AS1-knockdown MCF-7 cells,
2x106 MCF-7 cells were transfected with 4 mg shTMPO-AS1
(5'-CCGGGAGCCGAACTACGAACCAACTCGAGTTGGTTCGTAGTTCGGCTCTTTTTG-3') or
sh-NC
(5'-CCGGTTCTCCGAACGTGTCACGTCTCGAGACGTGACACGTTCGGAGAATTTTTG-3')
plasmids (Hanheng Biotechnology Co., Ltd.) using HyFect™
DNA Transfection Reagent (Leadgene Biomedical, Inc.). At 24 h
post-transfection, stable transfectants were selected using 500
mg/ml puromycin (Sigma-Aldrich; Merck KGaA). The selection medium
was replaced every 3 days for 2 weeks, and clones of resistant
cells were isolated and allowed to proliferate in medium containing
puromycin (500 mg/ml).
RNA isolation, cDNA synthesis and
reverse transcription-quantitative PCR (RT-qPCR)
Total RNA was isolated from transfected cells using
TRIzol® (Invitrogen; Thermo Fisher Scientific, Inc.)
according to the manufacturer's protocol. RNA integrity was
evaluated by performing 1.5% agarose gel electrophoresis. Total RNA
(1 µg) was reverse transcribed into cDNA using the Superscript III
First-Strand Synthesis system (Toyobo Life Science). The following
temperature protocol was used for reverse transcription: 37˚C for
15 min, 50˚C for 5 min and 98˚C for 5 min. Subsequently, qPCR was
performed using Fast SYBR® Green Master Mix (Thermo
Fisher Scientific, Inc.). The following primers were used for qPCR:
TMPO-AS1 forward, 5'-GTGCTGCAGGACCGAGG-3' and reverse,
5'-GCTTTGTGTCCGCGAGTTTT-3'; and GAPDH forward,
5'-AACGGATTTGGTCGTATTGG-3' and reverse, 5'-TTGATTTTGGAGGGATCTCG-3'.
The specific primer of miR-140-5p was: forward,
5'-GAGTGTCAGTGGTTACCGT-3', and reverse, 5'-GCATGGTCCGAGGTATTC-3'.
Primer of U6 was: forward 5'-CTCGCTTCGGCAGCACA-3' and reverse
5'-AACGCTTCACGAATTTGCGT-3' The following thermocycling conditions
were used for qPCR: Initial denaturation at 95˚C for 5 min;
followed by 40 cycles of denaturation at 95˚C for 15 sec, annealing
at 60˚C for 20 sec and elongation at 72˚C for 10 sec; and final
extension at 72˚C for 10 min. The relative expression level was
determined by the 2-ΔΔCq method
(20).
Plasmid construction and
transfection
A TMPO-AS1 expression vector was purchased from
Baizhi Biomedical Technology (Shanghai) Co., Ltd. to facilitate
TMPO-AS1 overexpression in breast cancer cells. The wild-type (WT)
and mutant (MUT) TMPO-AS1 sequences were cloned into the pmirGLO
vector (Promega Corporation). Cells were seeded (2x105
cells/well) into 6-well plates. At 60-70% confluence, cells were
transfected with 4 µg plasmid using Lipofectamine 2000 according to
the manufacturer's protocol. At 48 h post-transfection, cells were
collected and used for subsequent experiments.
Cell Counting Kit-8 (CCK-8) assay
Cell viability was assessed by performing a CCK-8
assay (Dojindo Molecular Technologies, Inc.) according to the
manufacturer's protocol. Cells were seeded (2x103
cells/well) in 96-well plates and cultured for 24 h. Following
transfection, cells were cultured for 24, 48 or 72 h. Subsequently,
the medium in each well was replaced with 100 µl complete medium
containing 10 µl CCK-8 solution, and the plate was incubated for 1
h at 37˚C. Absorbance was measured at wavelengths of 450 nm using a
Multiskan Spectrum spectrophotometer (Thermo Fisher Scientific,
Inc.).
Colony formation assay
Cells were seeded (1x103 cells/well) into
6-well plates. Following culture for 2 weeks, the colonies were
treated with 70% methanol at room temperature for 15 min, followed
by staining with 0.1% crystal violet (Sigma-Aldrich; Merck KGaA) at
room temperature for 20 min. Following extensive washing with
phosphate-buffered saline, the cells were observed under a light
microscope (DM1000; Leica Microsystems GmbH). Visible colonies of
≥50 cells were then counted (magnification, x100).
Wound healing and invasion assays
Cell migration was assessed by performing the wound
healing assay. At 80-90% confluence, a 10 µl sterile pipette tip
was used to create a scratch wound in the cell monolayer. Cells
were cultured in media containing 2% FBS for 24 h at room
temperature. The wounds were observed using a phase-contrast light
microscope (magnification, x100) (DM1000; Leica Microsystems GmbH).
The wound was imaged at 0 and 24 h and the percentage of migration
was calculated using ImageJ software (version 1.43; National
Institutes of Health). To assess cell invasion, an invasion assay
was performed using Transwell inserts (pore size, 8 µm). The upper
surface of the membrane was coated with Matrigel (BD Biosciences)
at 4˚C overnight according to the manufacturer's protocol.
Subsequently, cells (2x105) were added to the upper
chamber for 24 h at 37˚C. A total of 750 µl DMEM supplemented with
20% fetal bovine serum was added to the lower chamber. Non-invading
cells were removed using a cotton swab. Invading cells were fixed
with 70% methanol at room temperature for 15 min and stained with
leucocrystal violet for 20 min. To assess cell migration, the same
procedure was performed without the use of Matrigel. Stained cells
were visualized using a light microscope (magnification, x100)
(DM1000; Leica Microsystems GmbH).
In vivo lung-colonization assays
BALB/c nude mice (5-6 weeks old, 18-20 g) were
purchased from the Laboratory Animal Center of the Harbin Medical
University and housed in barrier facilities on a 12-h light/dark
cycle under specific pathogen-free conditions. Mice were maintained
at 20-25˚C with 40-70% humidity, 12-h light/dark cycles, and free
access to food and drinking water. Eating, feeding and operating
procedures strictly followed aseptic principles. The mice were
treated in accordance with protocols approved by The Third
Affiliated Hospital of Harbin Medical University. MCF7 cells
(2x106) or PBS were injected into the tail vein of each
mouse. At 5 weeks post-injection, the mice were euthanized
according to Institutional Animal Care and Use Committee protocols
(n=5 per group) (21). The lungs
were dissected and stored in liquid nitrogen or fixed in 4%
formalin for 30 min at room temperature for further analysis. Next,
5-µm-thick sections were stained with hematoxylin and eosin for 10
min at room temperature and scanned using a Scanscope XT digital
slide scanner (Aperio Technologies, Inc.). Digital images of lung
sections were used to analyse the metastatic burden. Lung tumour
lesions were digitally demarcated, and the number of lesions per
section and the individual lesion area were determined using
Spectrum software (version 11.2.0.3; Aperio Technologies, Inc.).
The metastatic burden was calculated as an average of the total
area of tumour lesions divided by the total lung area across
four-step sections. The following humane endpoints were used in the
present study: i) tumor size exceeded 10% body weight; ii) tumor
ulceration; and iii) extreme weight loss. None of the endpoints
were observed in the mice during the present study.
In vivo tumor growth model
BALB/c nude mice (5-6 weeks old, 18-20 g) were
purchased from the Laboratory Animal Center of the Harbin Medical
University and housed in barrier facilities on a 12-h light/dark
cycle under specific pathogen-free conditions. Breast cancer cells
were stably transfected with sh-TMPO-AS1 or sh-NC, washed with PBS
and resuspended in DMEM (1x108 cells/ml). Subsequently,
100 µl cell suspension was subcutaneously injected into the
posterior side of BALB/c mice (n=5 per group). Tumor size and
volume were recorded every 5 days. At 30 days post-injection, the
mice were euthanized, and the tumors separated for measurement and
weighing. The following humane endpoints were used in the present
study: i) tumor size exceeded 10% body weight; ii) tumor
ulceration; and iii) extreme weight loss. None of the endpoints
were observed in the mice during the present study.
Luciferase reporter assay
Luciferase reporter gene assay was implemented using
the Dual-Luciferase Reporter Assay System (Promega Corporation)
according to the manufacturer's instructions. Cells were seeded
into 24-well plates and transfected with a wild-type (WT)-TMPO-AS1
luciferase reporter gene vector, a mutant (Mut)-TMPO-AS1 vector
containing a 7-bp mutation on the predicted miR-140-5p binding site
within TMPO-AS1 (Hanheng Biotechnology Co., Ltd.), along with the
aforementioned miR-150-5p mimic or inhibitor using Lipofectamine
3000 (Life Technologies; Thermo Fisher Scientific, Inc.). Following
48 h, cells were lysed using passive lysis buffer (Promega
Corporation) and the luciferase activity was detected. Luciferase
activity was normalized against Renilla. All experiments
were performed at least three times.
Statistical analysis
Statistical analyses were performed using GraphPad
Prism software (version 6.0; GraphPad Software, Inc.). Data are
presented as the mean ± SEM. Comparisons between two groups were
analyzed using the paired Student's t-test. Comparisons among
multiple groups were analyzed using one-way or repeated measures
ANOVA followed by Bonferroni's post hoc test. P<0.05 was
considered to indicate a statistically significant difference.
Results
TMPO-AS1 expression is significantly
increased in breast cancer tissues and cell lines
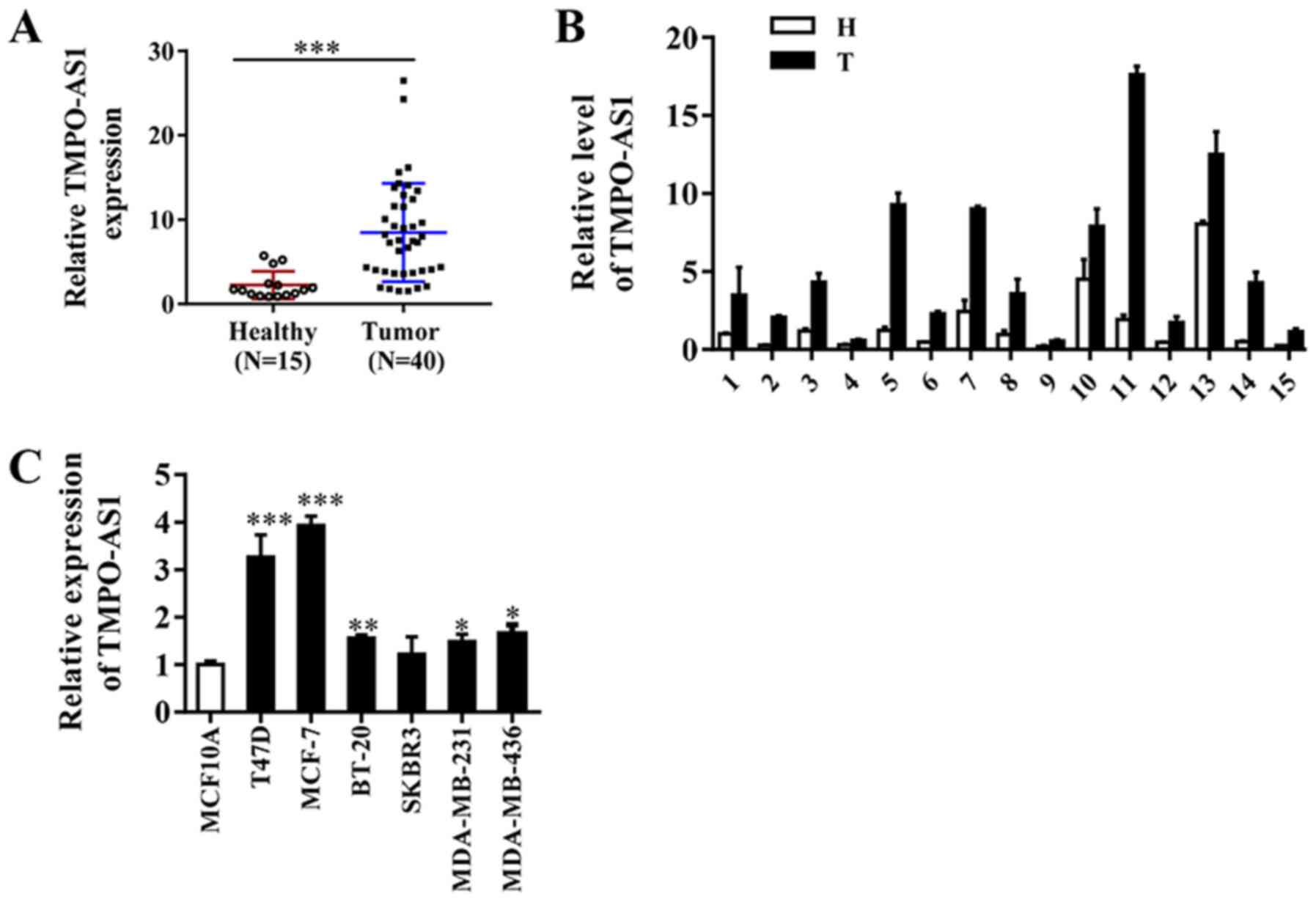
TMPO-AS1 expression was assessed in 40 breast cancer
and 15 adjacent non-cancerous tissue samples. The results indicated
that TMPO-AS1 expression was markedly upregulated in breast cancer
tissues compared with adjacent non-cancerous tissues (Fig. 1A and B). RT-qPCR was performed to further assess
differences in TMPO-AS1 expression in breast cancer and
immortalized breast epithelial cells. Compared with MCF-10A cells,
the expression levels of TMPO-AS1 were significantly increased in
breast cancer cell lines, except for the SKBR3 cell line (Fig. 1C). The results suggested that
increased expression of TMPO-AS1 may be associated with breast
cancer progression.
TMPO-AS1 knockdown inhibits breast
cancer cell viability in vitro and in vivo
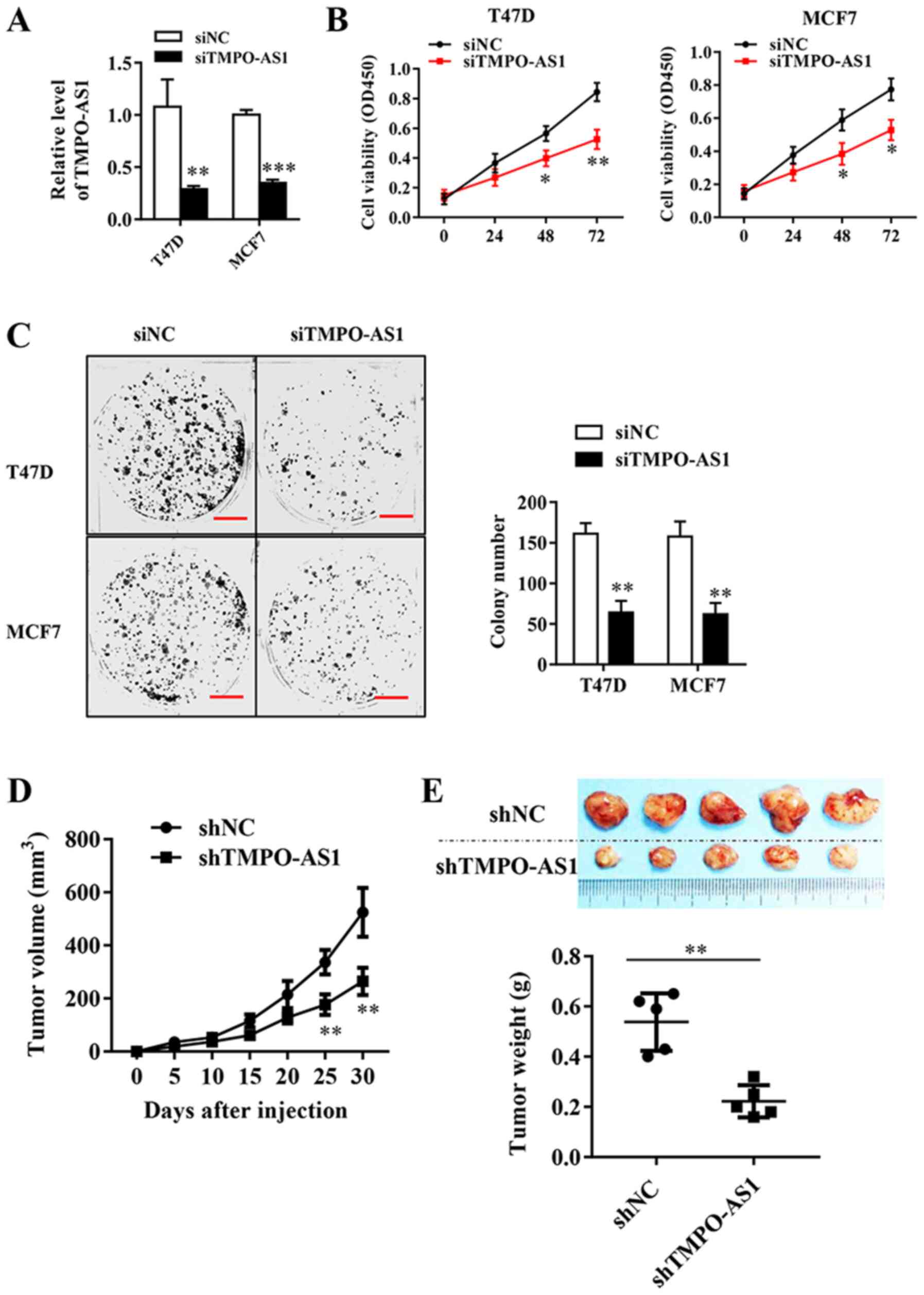
To directly evaluate the role of TMPO-AS1 in breast
cancer, T47D and MCF7 cells were transfected with TMPO-AS1 siRNA,
which significantly inhibited the expression of endogenous TMPO-AS1
compared with the siNC group (Fig.
2A). The effects of TMPO-AS1 knockdown on breast cancer cell
viability were analyzed by performing a CCK-8 assay. The results
suggested that TMPO-AS1 knockdown significantly inhibited breast
cancer cell viability at 48 and 72 h compared with the siNC group
(Fig. 2B). Similarly, TMPO-AS1
knockdown significantly reduced the colony forming abilities of
MCF-7 and T47D cells compared with siNC (Fig. 2C). To determine the effects of
TMPO-AS1 on tumor growth in vivo, tumor xenograft
experiments were performed using nude mice and established stable
TMPO-AS1-knockdown MCF cell lines (Fig.
2D). The tumor volumes were recorded every 5 days, and
the results indicated that TMPO-AS1 knockdown significantly
inhibited tumor growth in vivo from 25 days post-injection
compared with the shNC group (Fig.
2E). At the end of the experiment, the weight of each tumor was
measured, which indicated that TMPO-AS1 knockdown significantly
reduced tumor size compared with the shNC group (Fig. 2F). In summary, TMPO-AS1 knockdown
significantly inhibited cellular viability both in vivo and
in vitro.
TMPO-AS knockdown inhibits breast
cancer cell migration and invasion in vitro and in vivo
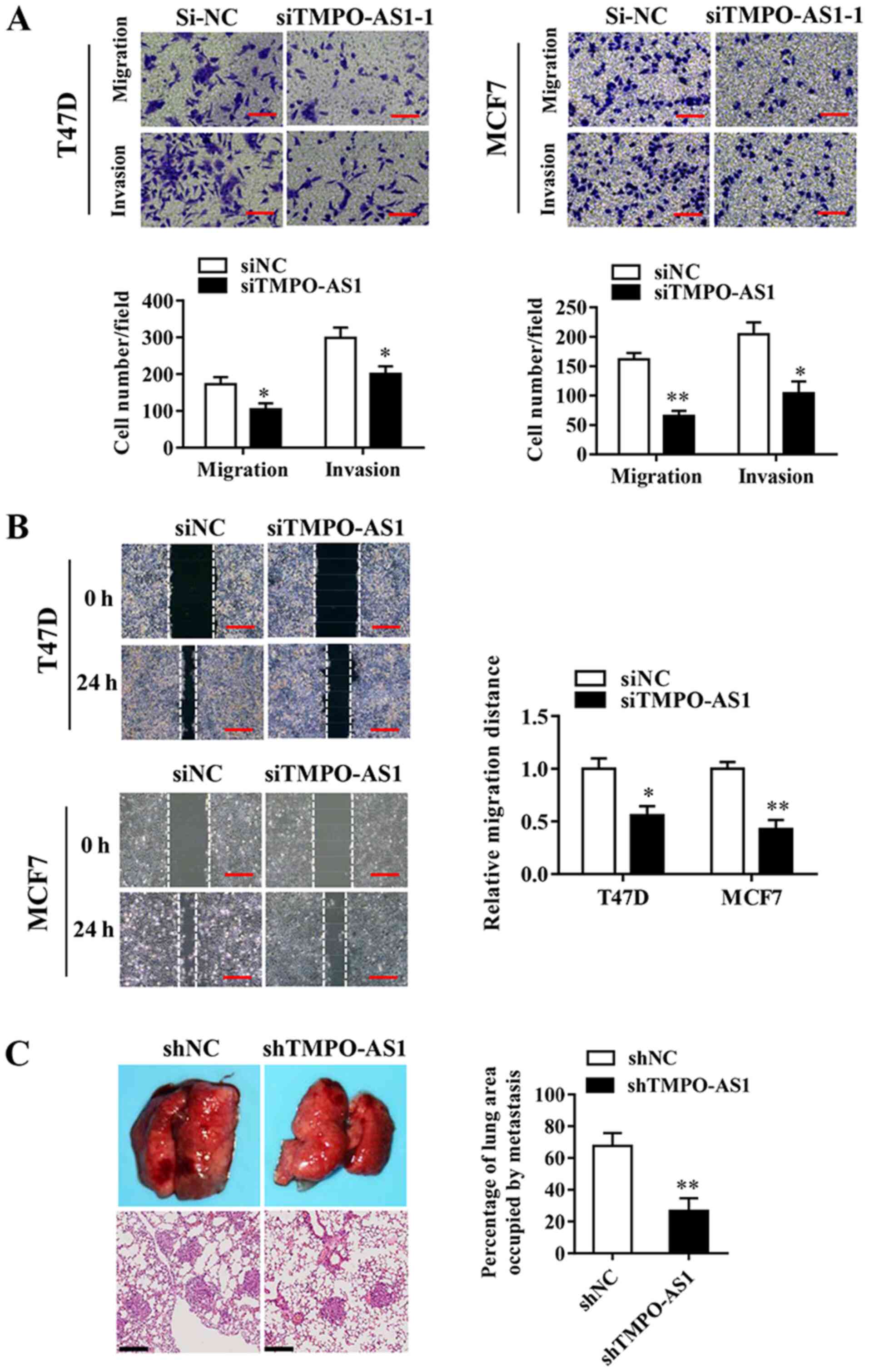
The effects of TMPO-AS1 knockdown on the invasive
abilities of T47D and MCF7 cells were examined, and the association
between TMPO-AS1 and breast cancer progression was investigated.
Cell migration/invasion assays were performed using 24-well
Transwells, coated without (migration) or with (invasion) Matrigel.
The Transwell invasion assay results indicated that TMPO-AS1
knockdown significantly inhibited T47D and MCF7 breast cancer cell
invasion and migration compared with the siNC group (Fig. 3A and B). In addition, the effects of TMPO-AS1 on
lung metastasis were observed by injecting TMPO-AS1-knockdown
breast cancer cells into nude mice via the tail vein. At 5 weeks
post-injection, TMPO-AS1 knockdown significantly reduced the number
of pulmonary nodules compared with the shNC group (Fig. 3C). The results indicated that
TMPO-AS1 knockdown impaired breast cancer cell invasion both in
vitro and in vivo.
TMPO-AS1 is a molecular sponge for
miR-140-5p
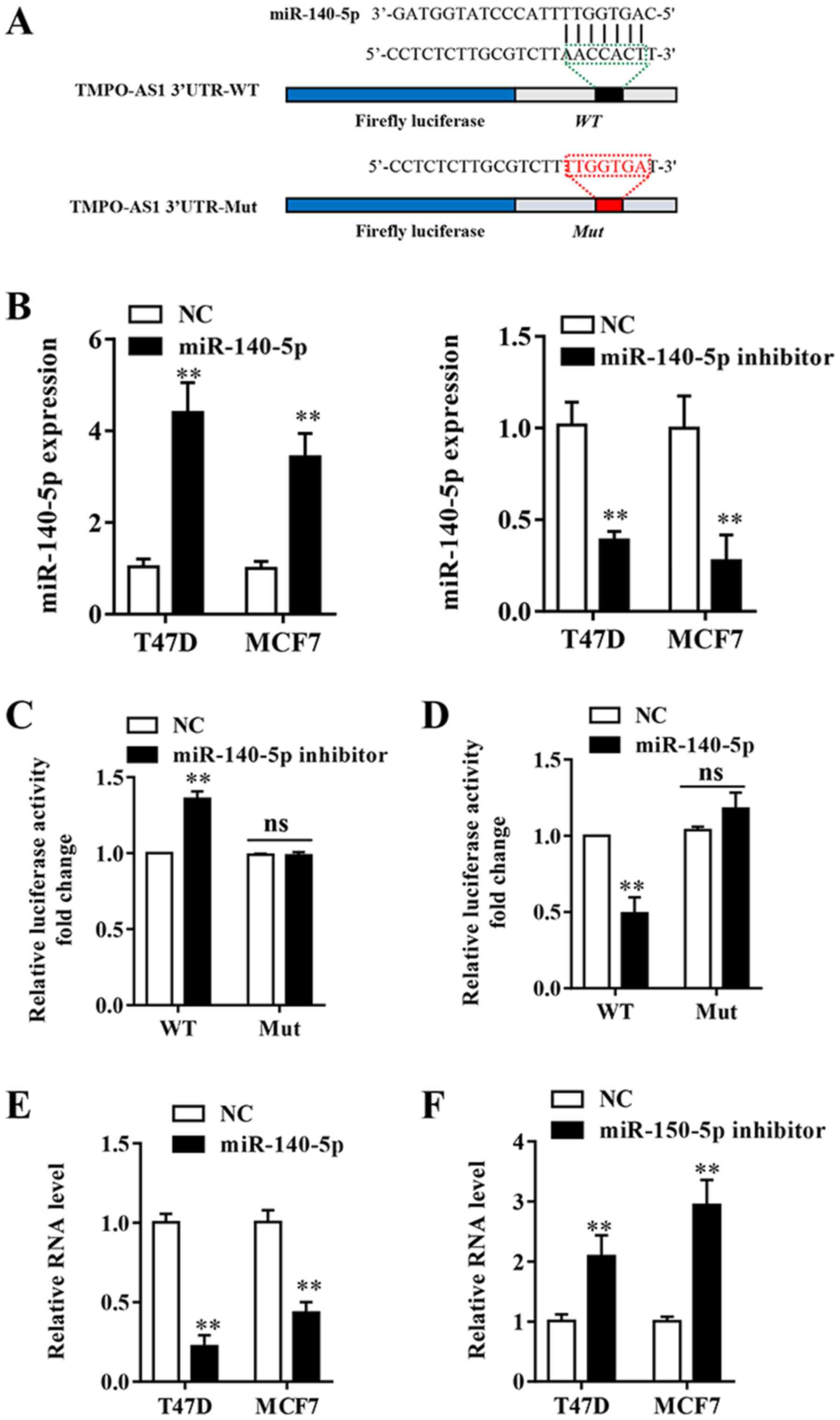
lncRNAs can exert their regulatory functions as
competitive endogenous RNAs (ceRNAs) (22). To further determine the potential
molecular mechanisms underlying lncRNAs in breast cancer, the
StarBase v2.0 (http://starbase.sysu.edu.cn/) and miRcode (version 11;
http://mircode.org/) online tools were used to predict
the potential binding partners of TMPO-AS1. miR-140-5p was
predicted to be a potential target for TMPO-AS1 (Fig. 4A). RT-qPCR was performed to assess
the transfection efficiency of miR-140-5p mimic and miR-140-5p
inhibitor. The results indicated that miR-140-5p expression was
significantly decreased in the miR-140-5p inhibitor group and
significantly increased in the miR-140-5p mimic group compared with
the NC group (Fig. 4B). A
Dual-Luciferase reporter assay was conducted and the results
indicated that miR-140-5p mimic significantly inhibited the
luciferase activity of TPSO-AS1-WT compared with the NC group,
whereas miR-140-5p inhibitor significantly increased the luciferase
activity of TPSO-AS1-WT compared with the NC group in MCF-7 cells
(Fig. 4C and D). Following mutation of the predicted
binding sites within TMPO-AS1, the effects of miR-140-5p mimic and
miR-140-5p inhibitor on luciferase activity were abolished
(Fig. 4C and D). The results indicated that TMPO-AS1
directly bound to miR-140-5p. Furthermore, miR-140-5p inhibitor
significantly upregulated TMPO-AS1 expression compared with the NC
group, whereas miR-140-5p mimic significantly decreased the
expression of TMPO-AS1 compared with the NC group in both cell
lines (Fig. 4E and F). The results suggested that miR-140-5p
directly and negatively regulated the expression of lncRNA
TMPO-AS1.
TMPO-AS1 promotes breast cancer
progression by competitively binding to miR-140-5p
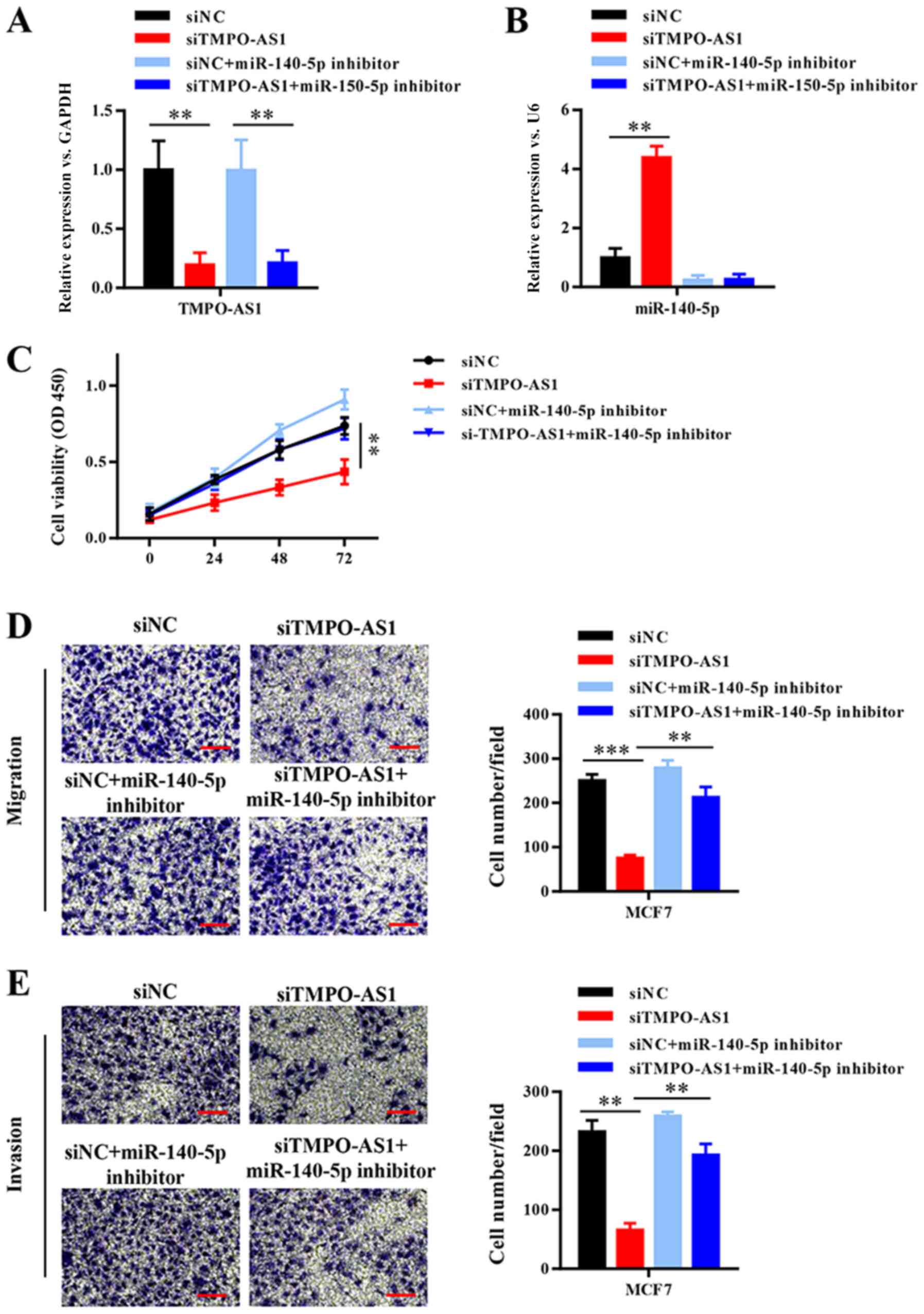
To further investigate whether TMPO-AS1-mediated
promotion of breast cancer cell malignancy was dependent on
negative regulation by miR-140-5p, TMPO-AS1 was knocked down in
MCF-7 cells, which were then transfected with miR-140-5p inhibitor
(Fig. 5A and B). Compared with the siNC group, TMPO-AS1
knockdown significantly inhibited cell viability, migration and
invasion, whereas miR-140-5p inhibition reversed TMPO-AS1
knockdown-mediated effects (Fig.
5C-E). The results suggested that TMPO-AS1 promoted breast
cancer progression by competitively binding to miR-140-5p.
Discussion
Despite the variety of available treatments, the
mortality rate of breast cancer remains one of the highest among
all cancer types worldwide (23),
and the incidence of breast cancer has increased rapidly (24). At present, the efficacy and
prognosis of patients with breast cancer are largely dependent on
clinical and pathological parameters (25). However, the underlying mechanisms
regulating the development of breast cancer are not completely
understood. Therefore, the identification of novel molecular
markers associated with breast cancer malignancy is important. In
the present study, TMPO-AS1 was significantly upregulated in breast
cancer tissues and cell lines compared with adjacent non-cancerous
tissues and MCF-10A cells, respectively.
lncRNAs are RNA molecules >200 nucleotides in
length that do not encode protein (26). In previous years, lncRNAs have been
reported to be involved in a number of biological and pathological
processes, and numerous lncRNAs have been associated with the
development and progression of malignancies, including breast
cancer (27). For instance, HOX
transcriptional antisense RNA is involved in breast cancer
metastasis by reprogramming the chromatin state (28), and kinase-activated long intergenic
non-coding RNAs promote tumor growth by activating the
hypoxia-inducible factor 1 signaling pathway (29). In the present study, the results
indicated that TMPO-AS1 was functionally associated with the
tumorigenicity and metastasis of breast cancer. Previous studies
have suggested that highly expressed lncRNAs may serve oncogenic
roles in tumorigenesis (30-32).
The RT-qPCR results in the present study indicated that TMPO-AS1
expression was significantly higher in breast cancer tissues
compared with adjacent non-cancerous tissues. Compared with MCF-10A
immortalized mammary epithelial cells, the expression of TMPO-AS1
was significantly increased in the breast cancer cell lines, except
for the SKBR3 cell line. In addition, compared with the siNC and
shNC groups, TMPO-AS1 knockdown inhibited breast cancer cell
viability and invasion in vivo and in vitro. The
results suggested that TMPO-AS1 may serve a tumorigenic role in
breast cancer. Besides, use of 2% FBS in the wound healing assay
was a limitation of the present study.
Although a large number of lncRNAs are reportedly
involved in human diseases (33),
the underlying regulatory mechanisms are not completely understood.
miRs are ~22 nucleotides in length and serve an essential role in
regulating target gene expression by base-pairing with
complementary sites in the 3'-untranslated (3'UTRs), 5'UTRs, and
coding regions of target mRNAs (34-36).
Based on the previously proposed ceRNA hypothesis, multiple lncRNA
transcripts have been confirmed to possess binding sites for
endogenous miRNAs, the binding of which allows for the regulation
of both lncRNA and miRNA activity (37-39).
In the present study, bioinformatics analysis indicated that
TMPO-AS1 contained miR-140-5p binding sites, which was further
verified by performing a dual luciferase reporter assay. The CCK-8
and Transwell assays were also performed to further determine
whether TMPO-AS1 promoted cell viability, migration and
invasiveness by inhibiting miR-140-5p. The results suggested that
miR-140-5p knockdown reversed TMPO-AS1 knockdown-mediated effects
on cell viability, migration and invasion. In summary, the results
suggested that TMPO-AS1 may serve as an oncogene that promotes
breast cancer progression by negatively regulating the tumor
suppressor miR-140-5p. Therefore, the TMPO-AS1/miR-140-5p complex
may serve as a novel target for the treatment of breast cancer.
In conclusion, the results of the present study
indicated that lncRNA TMPO-AS1 expression was increased in breast
cancer. The function of TMPO-AS1 in breast cancer cells also
suggested that it displayed carcinogenic characteristics during the
development of breast cancer. In addition, the results suggested
that TMPO-AS1 promoted breast cancer cell viability and migration
by sponging miR-140-5p. The results may facilitate the detection of
lncRNAs to guide the development of improved diagnostic and
therapeutic strategies for breast cancer.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
DZ and QZ conceived and designed the study, drafted
and revised the manuscript and supervised the project. DZ, QZ and
WL performed the experiments. DZ, QZ, XZ, YH, HY, YY and GZ
analyzed and interpreted the data. All authors read and approved
the final manuscript.
Ethics approval and consent to
participate
The present study was approved by The Animal Care
and Use Committee of the Third Affiliated Hospital of Harbin
Medical University (Harbin, China; approval number:
20170529003).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
DeSantis CE, Fedewa SA, Goding Sauer A,
Kramer JL, Smith RA and Jemal A: Breast cancer statistics, 2015:
Convergence of incidence rates between black and white women. CA
Cancer J Clin. 66:31–42. 2016.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Graveel CR, Calderone HM, Westerhuis JJ,
Winn ME and Sempere LF: Critical analysis of the potential for
microRNA biomarkers in breast cancer management. Breast Cancer
(Dove Med Press). 7:59–79. 2015.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Antonarakis ES: AR signaling in human
malignancies: Prostate cancer and beyond. Cancers (Basel).
10(22)2018.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Rahim B and O'Regan R: AR signaling in
breast cancer. Cancers (Basel). 9(21)2017.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Anastasiadi Z, Lianos GD, Ignatiadou E,
Harissis HV and Mitsis M: Breast cancer in young women: An
overview. Updates Surg. 69:313–317. 2017.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Torre LA, Siegel RL, Ward EM and Jemal A:
Global cancer incidence and mortality rates and trends-an update.
Cancer Epidemiol Biomarkers Prev. 25:16–27. 2016.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Djebali S, Davis CA, Merkel A, Dobin A,
Lassmann T, Mortazavi A, Tanzer A, Lagarde J, Lin W, Schlesinger F,
et al: Landscape of transcription in human cells. Nature.
489:101–108. 2012.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Santosh B, Varshney A and Yadava PK:
Non-coding RNAs: Biological functions and applications. Cell
Biochem Funct. 33:14–22. 2015.PubMed/NCBI View
Article : Google Scholar
|
|
9
|
ENCODE Project Consortium; Birney E,
Stamatoyannopoulos JA, Dutta A, Guigó R, Gingeras TR, Margulies EH,
Weng Z, Snyder M, Dermitzakis ET, et al: Identification and
analysis of functional elements in 1% of the human genome by the
ENCODE pilot project. Nature 447: 799-816, 2007.
|
|
10
|
Yuan JH, Yang F, Wang F, Ma JZ, Guo YJ,
Tao QF, Liu F, Pan W, Wang TT, Zhou CC, et al: A long noncoding RNA
activated by TGF-β promotes the invasion-metastasis cascade in
hepatocellular carcinoma. Cancer Cell. 25:666–681. 2014.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Han P and Chang CP: Long non-coding RNA
and chromatin remodeling. RNA Biol. 12:1094–1098. 2015.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Ponting CP, Oliver PL and Reik W:
Evolution and functions of long noncoding RNAs. Cell. 136:629–641.
2009.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Li Y, Zhu G, Ma Y and Qu H: lncRNA CCAT1
contributes to the growth and invasion of gastric cancer via
targeting miR-219-1. J Cell Biochem. 120:19457–19468.
2019.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Han M, Wang Y, Gu Y, Ge X, Seng J, Guo G,
Zhang X, Zhao Y and Dou D: lncRNA GHET1 knockdown suppresses breast
cancer activity in vitro and in vivo. Am J Transl Res. 11:31–44.
2019.PubMed/NCBI
|
|
15
|
He P, Zhang Z, Huang G, Wang H, Xu D, Liao
W and Kang Y: miR-141 modulates osteoblastic cell proliferation by
regulating the target gene of lncRNA H19 and lncRNA H19-derived
miR-675. Am J Transl Res. 8:1780–1788. 2016.PubMed/NCBI
|
|
16
|
Yang Q, Tang Y, Tang C, Cong H, Wang X,
Shen X and Ju S: Diminished LINC00173 expression induced miR-182-5p
accumulation promotes cell proliferation, migration and apoptosis
inhibition via AGER/NF-kappaB pathway in non-small-cell lung
cancer. Am J Transl Res. 11:4248–4262. 2019.PubMed/NCBI
|
|
17
|
Zhang L, Li S, Choi YL, Lee J, Gong Z, Liu
X, Pei Y, Jiang A, Ye M, Mao M, et al: Systematic identification of
cancer-related long noncoding RNAs and aberrant alternative
splicing of quintuple-negative lung adenocarcinoma through RNA-Seq.
Lung Cancer. 109:21–27. 2017.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Yin YZ, Zheng WH, Zhang X, Chen YH and Tuo
YH: LINC00346 promotes hepatocellular carcinoma progression via
activating the JAK-STAT3 signaling pathway. J Cell Biochem.
121:735–742. 2019.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Mitobe Y, Ikeda K, Sato W, Kodama Y, Naito
M, Gotoh N, Miyata K, Kataoka K, Sasaki H, Horie-Inoue K and Inoue
S: Proliferation-associated long noncoding RNA, TMPO-AS1, is a
potential therapeutic target for triple-negative breast cancer.
Cancer Sci. 111:2440–2450. 2020.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Ngan E, Stoletov K, Smith HW, Common J,
Muller WJ, Lewis JD and Siegel PM: LPP is a Src substrate required
for invadopodia formation and efficient breast cancer lung
metastasis. Nat Commun. 8(15059)2017.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Sumazin P, Yang X, Chiu HS, Chung WJ, Iyer
A, Llobet-Navas D, Rajbhandari P, Bansal M, Guarnieri P, Silva J
and Califano A: An extensive microRNA-mediated network of RNA-RNA
interactions regulates established oncogenic pathways in
glioblastoma. Cell. 147:370–381. 2011.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Ogretmen B and Hannun YA: Biologically
active sphingolipids in cancer pathogenesis and treatment. Nat Rev
Cancer. 4:604–616. 2004.PubMed/NCBI View
Article : Google Scholar
|
|
24
|
Wang Y, Zhou J, Wang Z, Wang P and Li S:
Upregulation of SOX2 activated LncRNA PVT1 expression promotes
breast cancer cell growth and invasion. Biochem Biophys Res Commun.
493:429–436. 2017.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Patani N, Martin LA and Dowsett M:
Biomarkers for the clinical management of breast cancer:
International perspective. Int J Cancer. 133:1–13. 2013.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Wang KC and Chang HY: Molecular mechanisms
of long noncoding RNAs. Mol Cell. 43:904–914. 2011.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Mitobe Y, Takayama KI, Horie-Inoue K and
Inoue S: Prostate cancer-associated lncRNAs. Cancer Lett.
418:159–166. 2018.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Gupta RA, Shah N, Wang KC, Kim J, Horlings
HM, Wong DJ, Tsai MC, Hung T, Argani P, Rinn JL, et al: Long
non-coding RNA HOTAIR reprograms chromatin state to promote cancer
metastasis. Nature. 464:1071–1076. 2010.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Lin A, Li C, Xing Z, Hu Q, Liang K, Han L,
Wang C, Hawke DH, Wang S, Zhang Y, et al: The LINK-A lncRNA
activates normoxic HIF1α signalling in triple-negative breast
cancer. Nat Cell Biol. 18:213–224. 2016.PubMed/NCBI View
Article : Google Scholar
|
|
30
|
Dong HT, Liu Q, Zhao T, Yao F, Xu Y, Chen
B, Wu Y, Zheng X, Jin F, Li J and Xing P: Long non-coding RNA
LOXL1-AS1 drives breast cancer invasion and metastasis by
antagonizing miR-708-5p expression and activity. Mol Ther Nucleic
Acids. 19:696–705. 2020.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Xiu B, Chi Y, Liu L, Chi W, Zhang Q, Chen
J, Guo R, Si J, Li L, Xue J, et al: LINC02273 drives breast cancer
metastasis by epigenetically increasing AGR2 transcription. Mol
Cancer. 18(187)2019.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Lv L, He L, Chen S, Yu Y, Che G, Tao X,
Wang S, Jian Z and Zhang X: Long non-coding RNA LINC00114
facilitates colorectal cancer development through
EZH2/DNMT1-induced miR-133b suppression. Front Oncol.
9(1383)2019.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Abe M and Bonini NM: MicroRNAs and
neurodegeneration: Role and impact. Trends Cell Biol. 23:30–36.
2013.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Bartel DP: MicroRNAs: Target recognition
and regulatory functions. Cell. 136:215–233. 2009.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Da Sacco L and Masotti A: Recent insights
and novel bioinformatics tools to understand the role of microRNAs
binding to 5' untranslated region. Int J Mol Sci. 14:480–495.
2012.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Brummer A and Hausser J: MicroRNA binding
sites in the coding region of mRNAs: Extending the repertoire of
post-transcriptional gene regulation. Bioessays. 36:617–626.
2015.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Cesana M, Cacchiarelli D, Legnini I,
Santini T, Sthandier O, Chinappi M, Tramontano A and Bozzoni I: A
long noncoding RNA controls muscle differentiation by functioning
as a competing endogenous RNA. Cell. 147:358–369. 2011.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Karreth FA, Tay Y, Perna D, Ala U, Tan SM
Rust AG, DeNicola G, Webster KA, Weiss D, Perez-Mancera PA, et al:
In vivo identification of tumor-suppressive PTEN ceRNAs in an
oncogenic BRAF-induced mouse model of melanoma. Cell. 147:382–395.
2011.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Salmena L, Poliseno L, Tay Y, Kats L and
Pandolfi PP: A ceRNA hypothesis: The Rosetta Stone of a hidden RNA
language? Cell. 146:353–358. 2011.PubMed/NCBI View Article : Google Scholar
|