Introduction
Liver fibrosis is a tissue repair response to
various types of chronic liver injury and a common pathological
process among all chronic liver diseases (1). Without effective treatment, liver
fibrosis can develop into liver cirrhosis, portal hypertension and
even liver cancer (1), which can
seriously affect the quality of life of patients. Liver fibrosis is
characterized by the excessive generation and deposition of
extracellular matrix (ECM), including type I collagen (COL-I) and
fibronectin, in the liver parenchyma, resulting in damage to liver
structure and function (2,3). Basic and clinical studies have
reported that activated hepatic stellate cells (HSCs), namely
myofibroblasts, are the primary source of ECM in the process of
liver fibrosis (2,3).
Shikonin is a naphthoquinone compound derived from
the root of Lithospermum erythrorhizon (4). In addition to its previously reported
anti-inflammatory, antiviral and antitumor properties, shikonin
exerts an effect against tissue and organ fibrosis (5). For example, Nie et al (6) reported that shikonin inhibited lung
fibroblast proliferation and activation by regulating the Akt/P38
signaling pathway. Ding et al (7) demonstrated that shikonin inhibited the
activation of renal mesenchymal fibroblasts and renal fibrosis by
inhibiting aerobic glycolysis. Additionally, Fan et al
(8) observed that shikonin reduced
TGF-β1-induced collagen production and contraction in hypertrophic
scars derived from human skin fibroblasts. Yang et al
(9) reported that shikonin
alleviated isoproterenol-induced myocardial injury by inhibiting
fibrosis, apoptosis, inflammation and endoplasmic reticulum stress.
However, the action and mechanisms underlying shikonin in the
prevention of liver fibrosis are not completely understood.
Therefore, the present study aimed to investigate the effects of
shikonin on liver fibrosis and explore the possible underlying
mechanisms.
Materials and methods
Cell lines and cell culture
The classic HSC line LX-2 was obtained from the
American Type Culture Collection. Cells were cultured in DMEM
supplemented with 10% FBS, 100 mg/ml penicillin and 100 U/ml
streptomycin (all from Gibco; Thermo Fisher Scientific, Inc.) at
37˚C with 5% CO2. To induce cell activation, LX-2 cells
(1x105 cells/well) were incubated with TGF-β (5 ng/ml;
PeproTech, Inc.) for 24 h at 37˚C.
Cell viability assay
LX-2 cells (1x103 cells/well) were seeded
into 96-well plates and treated with shikonin (10 µmol; 99.8%;
MedChemExpress.) for 24 h at 37˚C. Subsequently, Cell Counting
Kit-8 (CCK-8; BioSharp Life Sciences) reagent was added for 4 h at
37˚C, according to the manufacturer's protocol. The absorbance of
each well was measured at a wavelength of 450 nm using a microplate
reader (Thermo Fisher Scientific, Inc.). DNA synthesis was measured
by performing an EdU incorporation assay (Shanghai Yeasen
Biotechnology Co., Ltd.), according to the manufacturer's protocol.
Cells were visualized using a fluorescence microscope
(magnification, x200; Nikon Corporation).
Western blotting
LX-2 cells (1x105 cells/well) were seeded
in 6-well plates. After culturing overnight, cells were stimulated
with TGF-β (5 ng/ml) for 24 h at 37˚C, followed by treatment with
shikonin (0.2 µmol/l) for 24 h at 37˚C. Total protein was extracted
using RIPA cell lysis buffer (Sigma-Aldrich; Merck KGaA) containing
a protease inhibitor mixture (Sigma-Aldrich; Merck KGaA) for 30
min. Subsequently, total protein was quantified using the BCA
method. Proteins (50 µg/lane) were separated via 12% SDS-PAGE and
transferred onto PVDF membranes (Merck KGaA). After blocking with
5% BSA (BD Biosciences) for 1 h at room temperature, the membranes
were incubated overnight at 4˚C with the following primary
antibodies: anti-α-smooth muscle actin (1:1,000; cat. no. ab7817),
anti-Bcl-2 (1:1,000; cat. no. ab59384), anti-Bax (1:1,000; cat. no.
ab32503), anti-light chain 3 (LC3)-II/I (1:1,000; cat. no.
ab192890), anti-cleaved-caspase-3 (1:1,000; cat. no. ab49822),
anti-caspase-3 (1:1,000; cat. no. ab13847), anti-Beclin-1 (1:1,000;
cat. no. ab210498), anti-COL-I (1:1,000; cat. no. ab260043),
anti-P62 (1:1,000; cat. no. ab56416), anti-JNK (1:1,000; cat. no.
ab76125), anti-phosphorylated (p)-JNK (1:1,000; cat. no. ab76572),
anti-P38 (1:1,000; cat. no. ab31828), anti-p-P38 (1:1,000; cat. no.
ab47363) and anti-platelet-activating factor (PAF; 1:1,000; cat.
no. ab104162). Subsequently, the membranes were incubated with
secondary antibodies for horseradish peroxidase-conjugated goat
anti-rabbit immunoglobulin G (1:5,000; cat. no. ab6721) or goat
anti-mouse IgG (1:5,000; cat. no. ab6728) for 2 h at room
temperature. All primary and secondary antibodies were purchased
from Abcam. Protein bands were visualized using ECL
chemiluminescence (Merck KGaA) and analyzed using Image-Pro Plus
software (version 6.0; Media Cybernetics, Inc.). GAPDH (Abcam;
1:1,000; cat. no. ab9485) was used as the loading control.
Immunofluorescence
LX-2 cells (1x105 cells/well) were seeded
onto glass coverslips in 6-well plates. After culturing overnight,
cells were stimulated with TGF-β (5 ng/ml) for 24 h at 37˚C,
followed by treatment with shikonin (0.2 µmol/l) for 24 h at 37˚C.
Cells were fixed with 4% paraformaldehyde for 15 min at room
temperature. Cells were rinsed twice with PBS, permeabilized with
0.1% Triton X-100 in PBS for 30 min at 4˚C and blocked with 5% BSA
in PBS for 1 h at room temperature. Cells were incubated with
LC3-II/I primary antibodies (Abcam; 1:1,000; cat. no. ab192890)
overnight at 4˚C. After rinsing three times with PBS, cells were
incubated with an Alexa Fluor-568-conjugated secondary anti-rabbit
IgG antibodies (1:250; Invitrogen; Thermo Fisher Scientific, Inc.)
for 1 h at room temperature. Subsequently, cells were
counterstained with DAPI (Beijing Solarbio Science & Technology
Co., Ltd.) for 10 min at room temperature to detect nuclei. Stained
cells were observed using an LSM410 confocal laser scanning
microscope (magnification, x200; Carl Zeiss AG) and analyzed using
Image-Pro Plus software (version 6; Media Cybernetics, Inc.).
Flow cytometric analysis of cellular
apoptosis
LX-2 cell apoptosis was determined via flow
cytometry using an Annexin V-FITC/propidium iodide (PI) kit (BD
Biosciences), according to the manufacturer's protocol. Briefly,
LX-2 cells (1x105 cells/well) were collected and
resuspended in 200 µl binding buffer with Annexin V and PI.
Following treatment with shikonin for 24 h, cells were incubated at
room temperature for 20 min and analyzed by flow cytometry using a
FACSCalibur flow cytometry (BD Biosciences).
Transmission electron microscopy
(TEM)
LX-2 cells were maintained in 2.5% glutaraldehyde
for 12 h at 4˚C and fixed with 1% osmic acid for 3 h at 4˚C. Cells
were dehydrated and embedded in epoxy resin for 4 h at room
temperature. Subsequently, cells were stained with uranyl acetate
and lead citrate. Cells were observed using TEM (Leica
Microsystems, Inc.).
Statistical analysis
Comparisons among multiple groups were analyzed
using one-way ANOVA followed by Tukey's post-hoc test. Statistical
analyses were performed using SPSS software (version 16.0; SPSS
Inc.). P<0.05 was considered to indicate a statistically
significant difference. All experiments were repeated ≥3 times.
Data are presented as the mean ± SD.
Results
Shikonin reduces TGF-β-induced
fibrosis in LX-2 cells
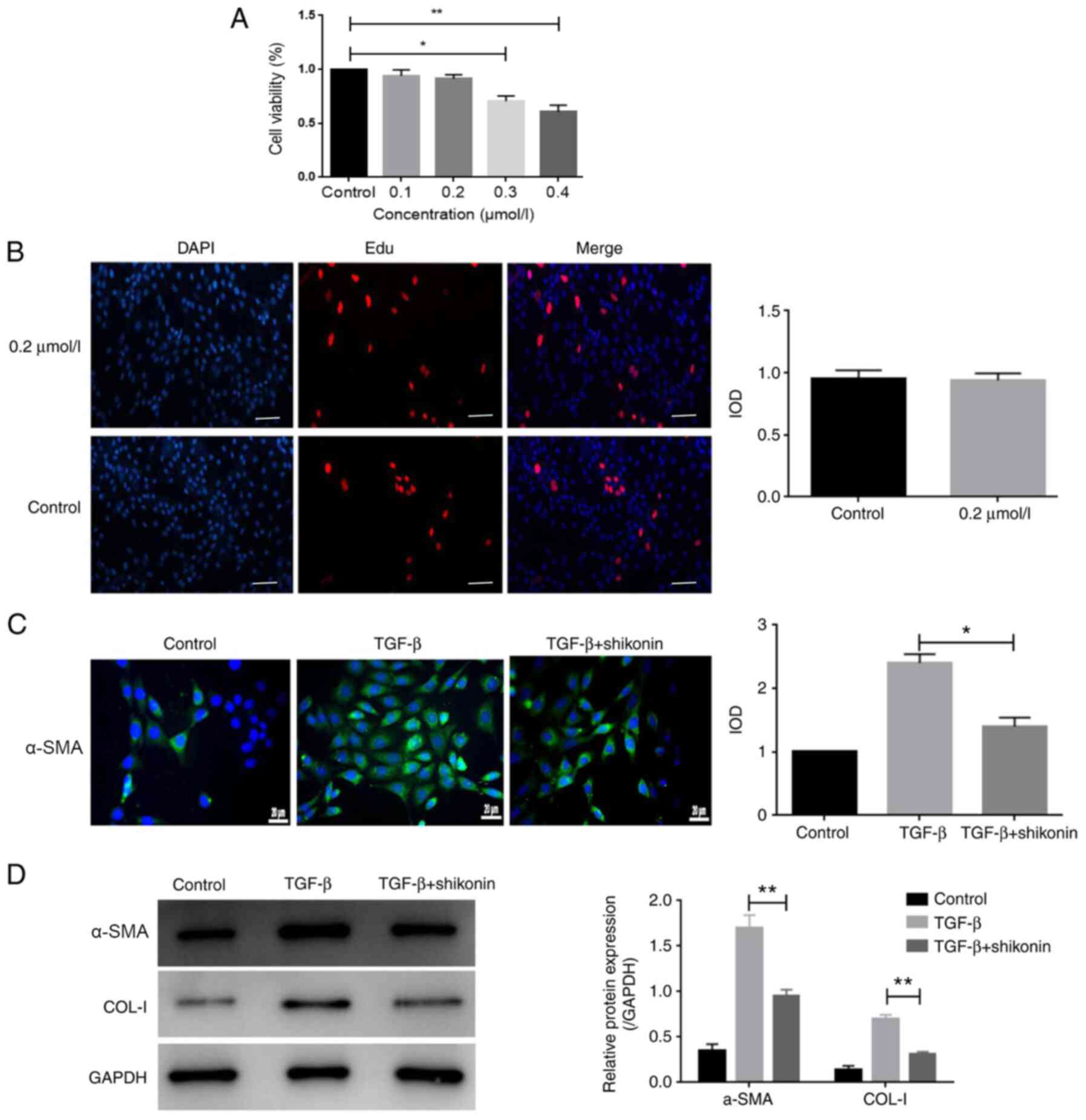
Cell viability was evaluated using the CCK-8 assay.
Cells were treated with 0.1-0.4 µmol/l shikonin for 24 h. The
results indicated that 0.1 and 0.2 µmol/l shikonin did not
significantly alter LX-2 cell viability compared with the control
group (Fig. 1A). By contrast, 0.3
and 0.4 µmol/l shikonin significantly reduced LX-2 cell viability
compared with the control group (Fig.
1A). Additionally, the results of the EdU incorporation assay
indicated that 0.2 µmol/l shikonin had no significant effect on
LX-2 cell viability compared with the control group (Fig. 1B). The cytotoxicity of shikonin is a
major index of its safety, which is a prerequisite for clinical
application; therefore, a non-toxic concentration (0.2 µmol/l) of
shikonin was used for subsequent experiments. To observe the effect
of shikonin on fibrosis in LX-2 cells, the expression of α-SMA was
measured. The protein expression levels of α-SMA and COL-I were
markedly increased by TGF-β and shikonin significantly decreased
TGF-β-induced protein expression (Fig.
1C and D). The results
indicated that shikonin markedly inhibited fibrosis.
Shikonin promotes apoptosis in
TGF-β-treated LX-2 cells
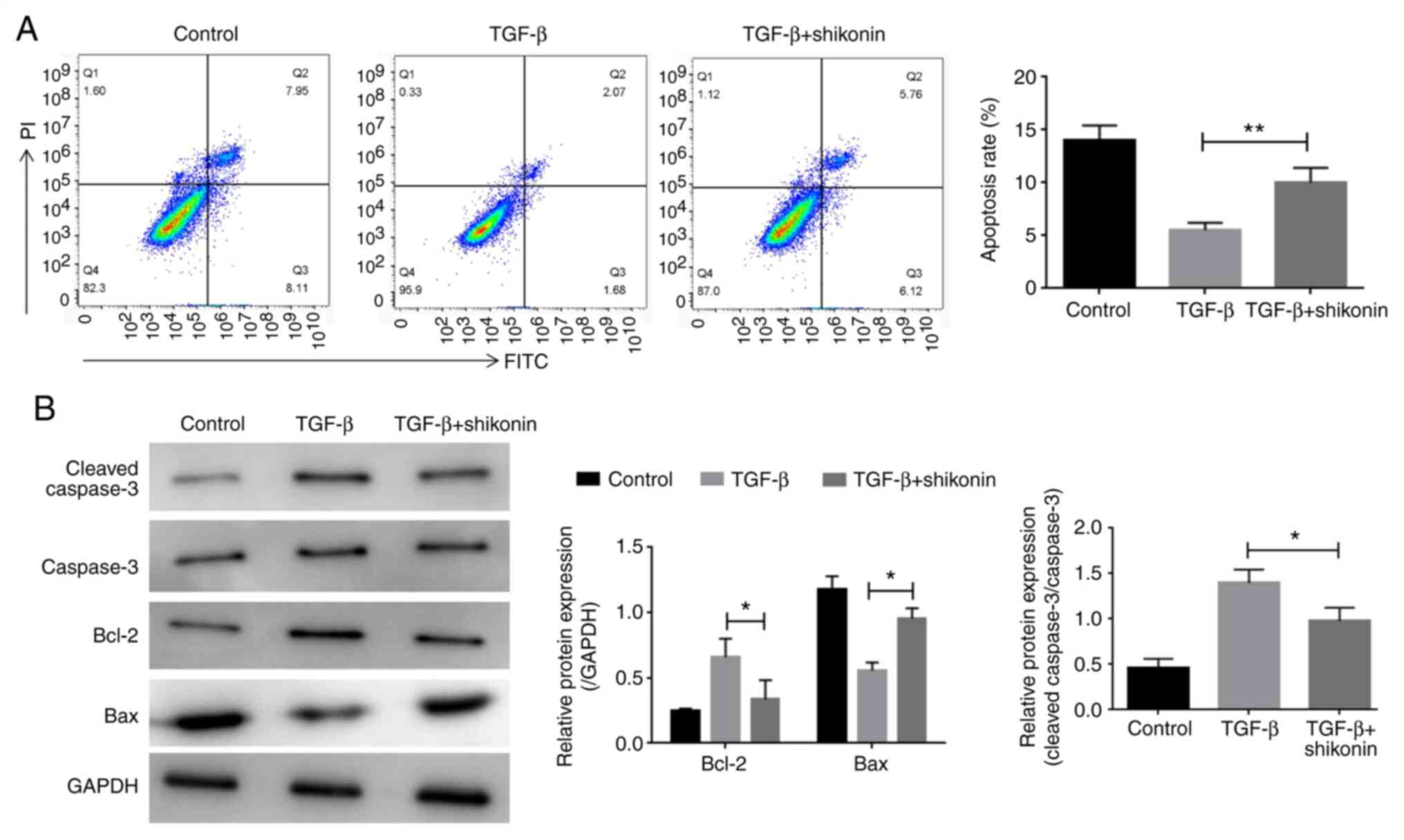
The effect of shikonin on cell apoptosis was
examined. LX-2 cells were treated with shikonin for 24 h and the
effect of shikonin on LX-2 cell apoptosis was determined by flow
cytometry. Compared with the control group, TGF-β treatment
decreased LX-2 cell apoptosis, whereas shikonin treatment
significantly decreased TGF-β-induced LX-2 cell apoptosis (Fig. 2A). Additionally, western blotting
was performed to detect the expression of apoptosis-related
proteins, including caspase-3, Bcl-2 and Bax. Compared with the
control group, TGF-β increased the expression levels of caspase-3
and Bcl-2, and decreased the expression levels of Bax. In contrast,
shikonin significantly decreased TGF-β-induced alterations in
protein expression (Fig. 2B).
Shikonin inhibits autophagy in
TGF-β-stimulated LX-2 cells
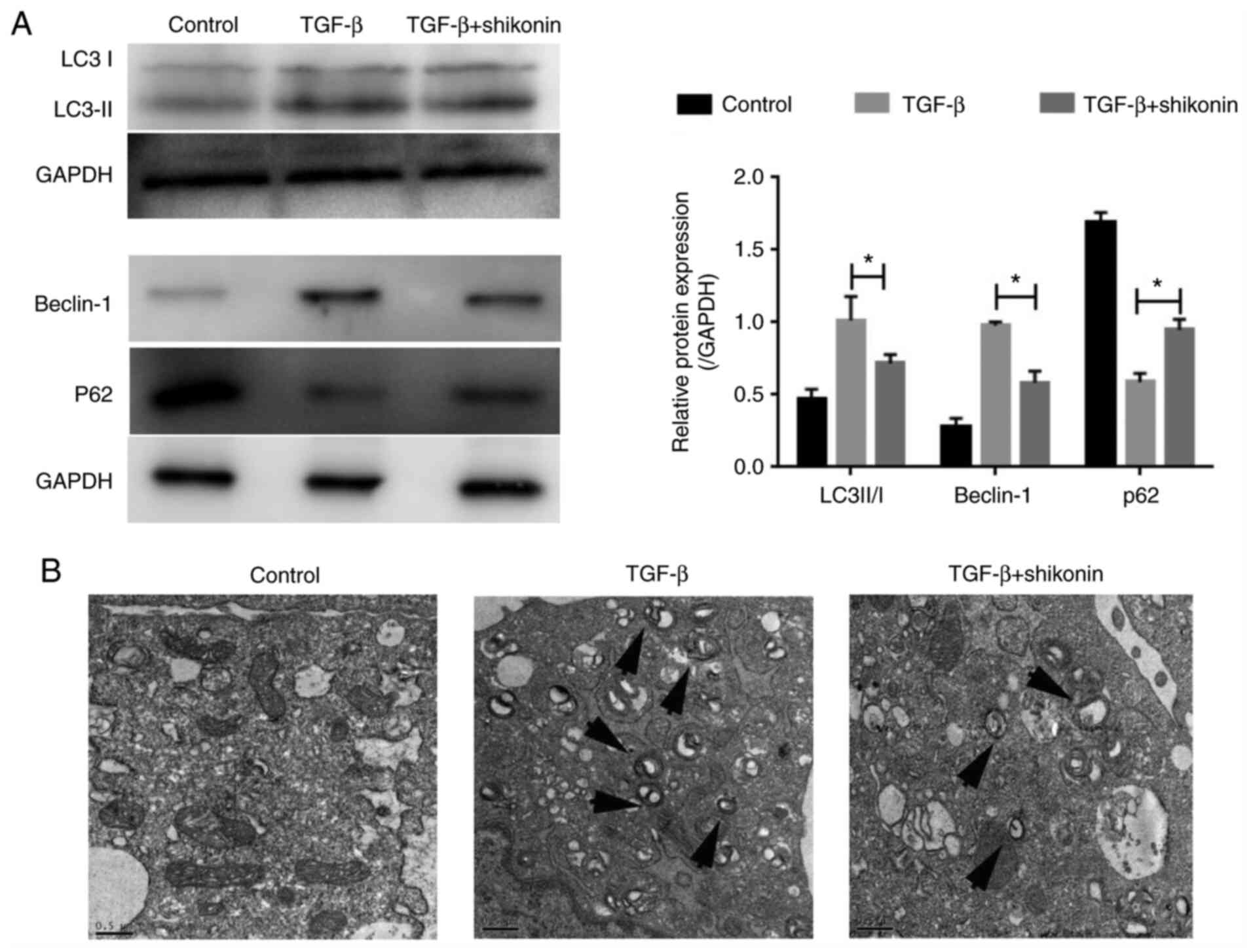
Autophagy serves an important role in fibrosis
(10). Therefore, whether shikonin
exerted a regulatory effect on autophagy was investigated. The
expression of autophagy-related proteins (LC3-I, LC3-II, Beclin-1
and P62) was determined. The expression levels of LC3-II/I and
Beclin-1 were significantly increased, whereas P62 expression
levels were significantly decreased in the TGF-β group compared
with the control group (Fig. 3A).
Moreover, shikonin significantly inhibited autophagy, as indicated
by reduced expression levels of LC3-II/I and Beclin-1, and
increased P62 expression level in the TGF-β + shikonin group
compared with the TGF-β group. Compared with the TGF-β group, the
number of autophagosomes in the shikonin group was notably reduced
(Fig. 3B). The results indicated
that the inhibitory effect of shikonin on fibrosis in LX-2 cells
may be mediated via suppressing autophagy.
Shikonin reduces the expression of PAF
in LX-2 cells
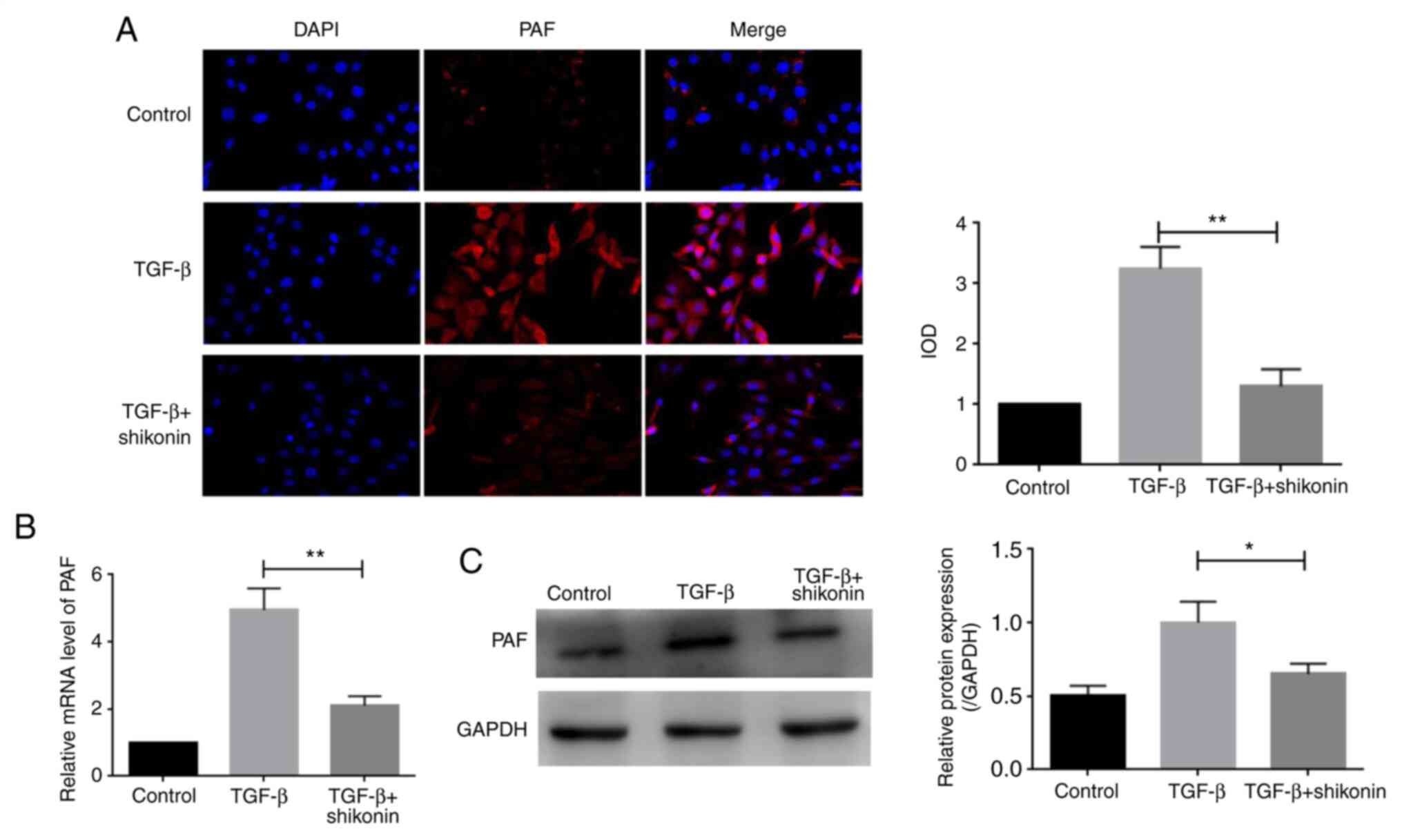
PAF serves a key role in cell apoptosis and
autophagy (5). The expression of
PAF was determined in the present study. PAF levels were higher in
the TGF-β group compared with the control group. PAF expression was
significantly decreased in the shikonin + TGF-β group compared with
the TGF-β group (Fig. 4A).
Additionally, the western blotting and RT-qPCR results indicated an
inhibitory effect of shikonin on PAF expression (Fig. 4B and C).
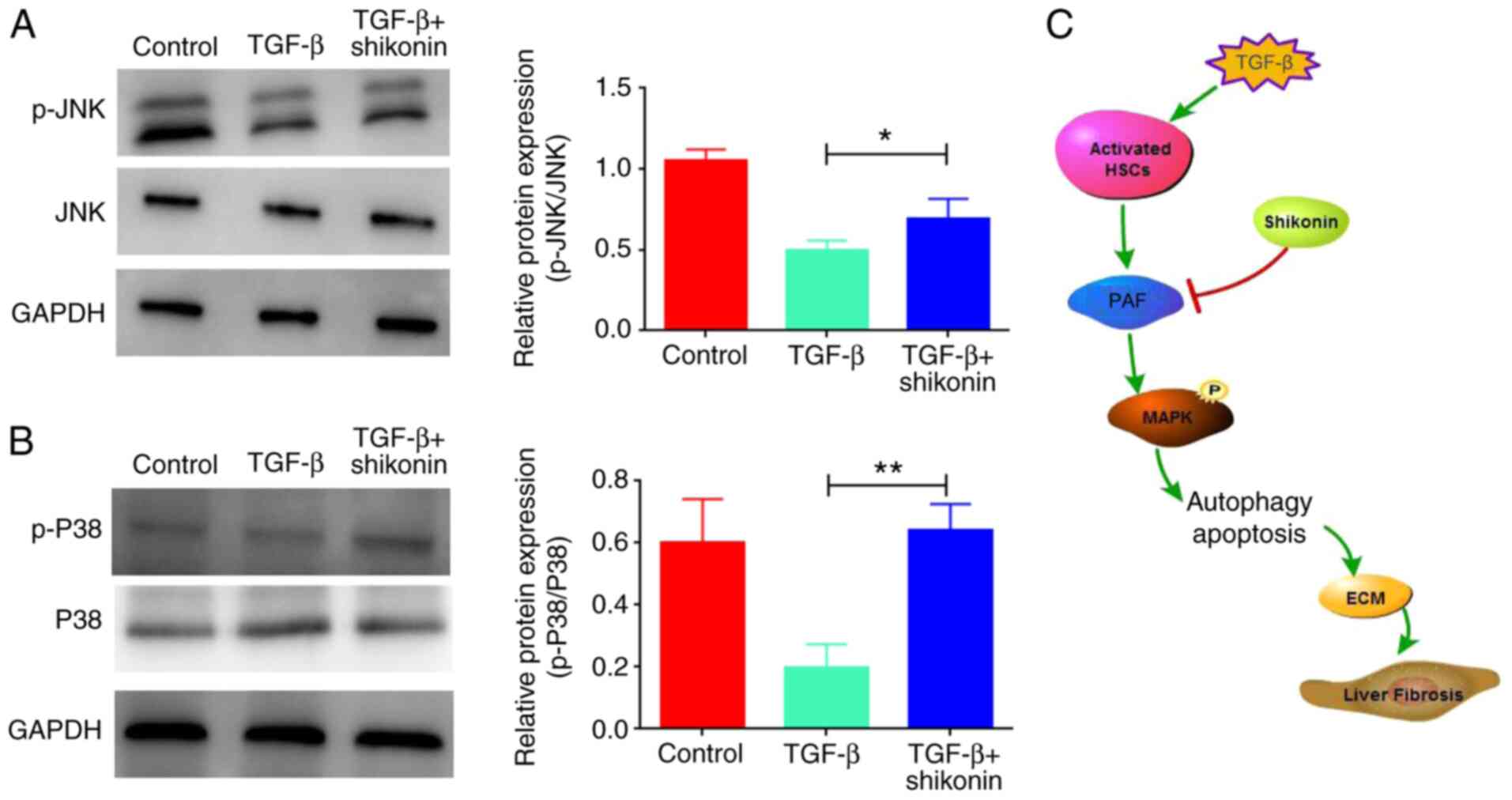
Shikonin activates MAPK signaling by
upregulating the expression levels of p-JNK and p-P38
The mechanisms of action underlying the effects of
shikonin on cell apoptosis and autophagy were evaluated. A previous
study reported that PAF regulated cell apoptosis via the MAPK
signaling pathway (5). Therefore,
whether shikonin affected MAPK signaling was investigated. The
results indicated that TGF-β inhibited the MAPK signaling pathway,
as indicated by downregulated p-JNK and p-P38 expression levels in
the TGF-β group compared with the control group, which were
significantly upregulated by shikonin treatment (Fig. 5).
Discussion
Liver fibrosis is a compensatory reaction in the
process of tissue repair following liver injury and inflammation
caused by various chronic pathogenic factors (1). During chronic liver injury, increased
ECM secretion and decreased degradation can lead to excessive
deposition of ECM, which eventually results in liver fibrosis
(1). Activated HSCs are the major
producers of ECM (1). In the
present study, TGF-β was used to induce LX-2 cell activation to
establish a hepatic fibrosis model. Shikonin induced apoptosis and
inhibited autophagy, thereby inhibiting liver fibrosis development.
Moreover, shikonin inhibited PAF expression and may serve a role in
inhibiting liver fibrosis via the MAPK signaling pathway.
Shikonin, a compound extracted from natural
lithospermum, has been reported to have a wide range of biological
and pharmacological properties. Previous studies have demonstrated
that shikonin exerts antibacterial, antitumor, anti-inflammatory
and antifibrosis effects (11-16).
Certain studies (16-19)
demonstrated that shikonin improved isoproterenol-induced
myocardial injury by inhibiting myocardial fibrosis, inflammation,
apoptosis and endoplasmic reticulum stress. Wang et al
(13) revealed that shikonin
inhibited the occurrence and development of pancreatic cancer via
the NF-κB signaling pathway. Jing et al (20) reported that shikonin induced HaCaT
cell apoptosis via the mitochondrial, Erk and Akt signaling
pathways. In the present study, the results indicated that shikonin
may inhibit the development of liver fibrosis by inducing HSC
apoptosis, indicating that shikonin may serve as a therapeutic
agent for liver fibrosis.
Autophagy is an important strategy to maintain cell
homeostasis (21). Autophagy is
active in liver cells and maintains the normal physiological
function of liver cells by eliminating easily accumulated proteins
and damaged mitochondria (21-23).
Additionally, autophagy is an important regulator implicated in
various liver diseases (22).
Although autophagy mediates the adaptive mechanism against injury
via degradation of the damaged organelles and proteins produced in
liver diseases, it may also serve as a promoter triggering the
development and progression of liver disease (22). Moreover, it has been demonstrated
that autophagosomes in HSCs promote the decomposition of lipid
droplets into free fatty acids in patients with liver fibrosis,
indicating that autophagy is involved in the physiological process
of liver fibrosis (22). Shen et
al (16) reported that
astaxanthin improved liver fibrosis by regulating TGF-1 expression
and the process of autophagy. In the present study, shikonin
significantly reduced the protein expression levels of LC3-II/I and
Beclin-1, and the TEM results indicated that the number of
autophagosomes was significantly reduced, suggesting that shikonin
may delay the process of liver fibrosis by inhibiting
autophagy.
PAF is a bioactive lipid neurotransmitter that acts
on various cells and tissues and serves an important role in the
pathogenesis of several diseases (24). Liang et al (24) reported that lncRNA pyruvate
formate-lyase regulated the expression of PAF by serving as a
competitive endogenous let-7d RNA, thereby regulating the
progression of myocardial fibrosis. Lv et al (25) demonstrated that rupatadine inhibited
pulmonary fibrosis by acting on PAF. The present study revealed
that shikonin significantly reduced the expression of PAF via the
MAPK signaling pathway in a cell model of liver fibrosis,
indicating that shikonin may inhibit liver fibrosis by regulating
the expression of PAF.
In summary, the results reported that shikonin
induced HSC apoptosis and inhibited autophagy by acting on the PAF
protein, thus blocking the development of liver fibrosis.
Therefore, the present study indicated a potential novel
therapeutic strategy for liver fibrosis. However, animal
experiments are required to verify the effect of shikonin in liver
fibrosis.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Wuhan Health
Bureau Research Project (grant no. WX09C07).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
MS and HZ designed and performed the experiments,
analyzed data and wrote the manuscript. ZC and JY performed the
experiments. JLi and SS performed parts of the experiments. JLiu
participated in the experimental design, provided financial support
and revised the manuscript. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Friedman SL: Evolving challenges in
hepatic fibrosis. Nat Rev Gastroenterol Hepatol. 7:425–436.
2010.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Zhang X, Cui JH, Meng QQ, Li SS, Zhou W
and Xiao S: Advance in anti-tumor mechanisms of shikonin, alkannin
and their derivatives. Mini Rev Med Chem. 18:164–172.
2018.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Hernandez-Gea V and Friedman SL:
Pathogenesis of liver fibrosis. Annu Rev Pathol. 6:425–456.
2011.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Ma X, Yu M, Hao C and Yang W: Shikonin
induces tumor apoptosis in glioma cells via endoplasmic reticulum
stress, and Bax/Bak mediated mitochondrial outer membrane
permeability. J Ethnopharmacol. 263(113059)2020.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Guo C, He J, Song X, Tan L, Wang M, Jiang
P, Li Y, Cao Z and Peng C: Pharmacological properties and
derivatives of shikonin-A review in recent years. Pharmacol Res.
149(104463)2019.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Nie Y, Yang Y, Zhang J, Cai G, Chang Y,
Chai G and Guo C: Shikonin suppresses pulmonary fibroblasts
proliferation and activation by regulating Akt and p38 MAPK
signaling pathways. Biomed Pharmacother. 95:1119–1128.
2017.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Ding H, Jiang L, Xu J, Bai F, Zhou Y, Yuan
Q, Luo J, Zen K and Yang J: Inhibiting aerobic glycolysis
suppresses renal interstitial fibroblast activation and renal
fibrosis. Am J Physiol Renal Physiol. 313:F561–F575.
2017.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Fan C, Dong Y, Xie Y, Su Y, Zhang X,
Leavesley D and Upton Z: Shikonin reduces TGF-β1-induced collagen
production and contraction in hypertrophic scar-derived human skin
fibroblasts. Int J Mol Med. 36:985–991. 2015.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Yang J, Wang Z and Chen DL: Shikonin
ameliorates isoproterenol (ISO)-induced myocardial damage through
suppressing fibrosis, inflammation, apoptosis and ER stress. Biomed
Pharmacother. 93:1343–1357. 2017.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Wang H and Zhang G: Activation of
CaMKKβ-AMPK-mTOR pathway is required for autophagy induction by
β,β-dimethylacrylshikonin against lung adenocarcinoma cells.
Biochem Biophys Res Commun. 517:477–483. 2019.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Tian R, Li Y and Gao M: Shikonin causes
cell cycle arrest and induces apoptosis by regulating the
EGFR/NF-κB signaling pathway in human epidermoid carcinoma A431
cells. Biosci Rep. 35(e00189)2015.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Wei Y, Li M, Cui S, Wang D, Zhang CY, Zen
K and Li L: Shikonin inhibits the proliferation of human breast
cancer cells by reducing tumor-derived exosomes. Molecules.
21(777)2016.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Wang Y, Zhou Y, Jia G, Han B, Liu J, Teng
Y, Lv J, Song Z, Li Y, Ji L, et al: Shikonin suppresses tumor
growth and synergizes with gemcitabine in a pancreatic cancer
xenograft model: Involvement of NF-κB signaling pathway. Biochem
Pharmacol. 88:322–333. 2014.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Wang L, Gai P, Xu R, Zheng Y, Lv S, Li Y
and Liu S: Shikonin protects chondrocytes from
interleukin-1beta-induced apoptosis by regulating PI3K/Akt
signaling pathway. Int J Clin Exp Pathol. 8:298–308.
2015.PubMed/NCBI
|
|
15
|
Wada N, Kawano Y, Fujiwara S, Kikukawa Y,
Okuno Y, Tasaki M, Ueda M, Ando Y, Yoshinaga K, Ri M, et al:
Shikonin, dually functions as a proteasome inhibitor and a
necroptosis inducer in multiple myeloma cells. Int J Oncol.
46:963–972. 2015.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Shen M, Chen K, Lu J, Cheng P, Xu L, Dai
W, Wang F, He L, Zhang Y, Chengfen W, et al: Protective effect of
astaxanthin on liver fibrosis through modulation of TGF-β1
expression and autophagy. Mediators Inflamm.
2014(954502)2014.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Zhu XF, Un XF, Liu GH and Chen FF: Effects
of shikonin on the proliferation, migration and differentiation of
HaCaT cells mediated by IL-22. J Clin Dermatol. 43:215–218.
2014.
|
|
18
|
Qisen Li, Zeng J, Su M, He Y and Zhu B:
Acetylshikonin from Zicao attenuates cognitive impairment and
hippocampus senescence in D-galactose-induced aging mouse model via
upregulating the expression of SIRT1. Brain Res Bulletin.
137:311–318. 2018.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Wang H and Zhang G: Endoplasmic reticulum
stress-mediated autophagy protects against
β,β-dimethylacrylshikonin-induced apoptosis in lung adenocarcinoma
cells. Cancer Sci. 109:1889–1901. 2018.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Jing H, Sun W, Fan J, Zhang Y, Yang J, Jia
J, Li J, Guo J, Luo S and Zheng Y: Shikonin induces apoptosis of
HaCaT cells via the mitochondrial, Erk and Akt pathways. Mol Med
Rep. 13:3009–3016. 2016.PubMed/NCBI View Article : Google Scholar
|
|
21
|
He C and Klionsky DJ: Regulation
mechanisms and signaling pathways of autophagy. Annual Rev Genet.
43:67–93. 2009.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Mallat A, Lodder J, Teixeira-Clerc F,
Moreau R, Codogno P and Lotersztajn S: Autophagy: A multifaceted
partner in liver fibrosis. Biomed Res Int 2014: 869390, 2014.
urihttps://doi.org/10.1155/2014/869390simplehttps://doi.org/10.1155/2014/869390.
|
|
23
|
Zhang XW, Zhou JC, Peng D, Hua F, Li K, Yu
JJ, Lv XX, Cui B, Liu SS, Yu JM, et al: Disrupting the TRIB3-SQSTM1
interaction reduces liver fibrosis by restoring autophagy and
suppressing exosome-mediated HSC activation. Autophagy. 16:782–796.
2020.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Liang H, Pan Z, Zhao X, Liu L, Sun J, Su
X, Xu C, Zhou Y, Zhao D, Xu B, et al: LncRNA PFL contributes to
cardiac fibrosis by acting as a competing endogenous RNA of let-7d.
Theranostics. 8:1180–1194. 2018.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Lv XX, Wang XX, Li K, Wang ZY, Li Z, Lv Q,
Fu XM and Hu ZW: Rupatadine protects against pulmonary fibrosis by
attenuating PAF-mediated senescence in rodents. PLoS One.
8(e68631)2013.PubMed/NCBI View Article : Google Scholar
|