Introduction
Myocardial infarction occurs when blood flow
decreases, causing damage to heart muscle (1). Acute myocardial infarction (AMI)
frequently induces cardiac dysfunction, eventually resulting in
heart failure, which is characterized by cardiac dysfunction and
cell death (2,3). During AMI, the myocardium undergoes a
series of structural alterations, including decreased proliferation
or abnormal apoptosis of myocardial cells (4,5).
Therefore, studying H9c2 cell viability and apoptosis following
hypoxic treatment was considered to be a suitable model of AMI for
the present study.
MicroRNAs (miRNAs/miRs), a class of non-coding small
endogenous RNAs 18-22 nucleotides in length, regulate the
expression of target genes by post-transcriptionally binding to the
3' untranslated region (3'UTR) of their target genes (6,7).
Numerous miRNAs have been reported to participate in the
development of various disorders, including diseases of the
cardiovascular system (8-10).
The miR-126 locus gives rise to two mature miRNAs: miR-126-3p and
miR-126-5p (11). Additionally,
deletion of miR-126 affects vascular integrity and angiogenesis
(12,13). A previous study demonstrated that
miR-126 expression was downregulated in patients with AMI following
the onset of symptoms (14),
indicating that miR-126 may function as a potential biomarker for
the diagnosis of AMI. However, the possible roles and mechanisms
underlying miR-126-5p in AMI are not completely understood.
Interleukin (IL)-17A is one of 30 known types of ILs
and one of the 6 known members of the IL-17 cytokine family
(15). IL-17A is an early promoter
of T-cell-induced inflammatory response, which can be amplified by
promoting the release of pre-inflammatory cytokines (16). IL-17A serves a crucial role in the
inflammatory reaction, tissue damage and immunological defense
(17-19).
A previous study revealed that IL-17A may be involved in H9c2 cell
development (20). However, the
association between miR-126-5p and IL-17A and their functions in
AMI are not completely understood.
Materials and methods
Serum samples
A total of 40 serum samples from patients with AMI
(male, 27; female, 13; age, 43-62 years; mean age, 55.3±3.8 years)
and 40 serum samples from healthy volunteers (male, 24; female, 16;
age, 40-67 years; mean age, 56.2±4.3 years) were collected from the
Taizhou People's Hospital of Jiangsu Province (Taizhou, China)
between December 2015 and January 2017. The inclusion criteria were
as follows: i) Electrocardiogram with characteristic alterations
including the emergence of Q, the spread of ST segment elevation
and the dynamic evolution of ST-T; ii) elevated serum biomarkers
for myocardial necrosis; iii) myocardial necrosis detected by serum
biomarkers; and iv) an intracoronary thrombus identified by
angiography. The exclusion criteria were as follows: i) Severe
renal dysfunction; ii) myocarditis; iii) rhabdomyolysis or
myositis; iv) cardiomyopathy; v) acute pulmonary embolism; vi)
cardiothoracic surgery, cardiac procedure or chest-wall trauma;
and/or vii) central nervous system pathology.
The present study was approved by the Ethics
Committee of Taizhou People's Hospital of Jiangsu Province. Written
informed consent was obtained from each patient or volunteer prior
to sample collection. All samples were stored at -80˚C until
further experimentation.
Cell culture
The rat myocardial H9c2 cell line (Shanghai Yanji
Biological Technology Co., Ltd.) was cultured in DMEM (Hyclone;
Cytiva) supplemented with 10% FBS (Gibco; Thermo Fisher Scientific,
Inc.) and 100 µg/ml streptomycin (Gibco; Thermo Fisher Scientific,
Inc.) in a humidified incubator with 5% CO2 at 37˚C.
H9c2 cells were seeded (seeding density, 1x106
cells/well) into six-well plates and transfected with 0.4 nM
miR-126-5p mimics or negative controls (NC) using
Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.) for 24 h at 37˚C until use. miR-126-5p mimics
(5'-CGCGTACCA AAAGTAATAATG-3') and NC (5'-GTGTAACACGTCTATACG
CCCA-3') were designed and synthesized by Shanghai GenePharma Co.,
Ltd.
Plasmid construction and
transfection
To produce IL-17A overexpression pcDNA3.1-IL-17A
plasmids, IL-17A coding sequence lacking the 3'UTR was cloned into
the pcDNA3.1 vector (Guangzhou RiboBio Co., Ltd.). Subsequently,
H9c2 cells (seeding density, 1x105 cells/well) were
transfected with 40 nM pcDNA3.1 and 40 nM pcDNA3.1-IL-17A plasmids
using Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.) for 2 h at 37˚C. Following this, H9c2 cells were
cultured at 37˚C for a further 24 h until subsequent analysis.
Dual-luciferase reporter assay
TargetScan (version no. 7.2; targetscan.org/vert_72) predicted that IL-17A was a
target of miR-126-5p. Subsequently, 2x104 cells/well
H9c2 cells were co-transfected with 40 nM wild-type or mutant
IL-17A 3'UTR plasmids (Guangzhou RiboBio Co., Ltd.) and miR-126-5p
mimics or NCs using Lipofectamine® 2000 (Invitrogen;
Thermo Fisher Scientific, Inc.). Following incubation for 24 h at
room temperature, luciferase activities were measured using a
Dual-Luciferase Reporter Assay system (Promega Corporation).
Firefly luciferase activities were normalized to Renilla
luciferase activities.
Hypoxic treatment
Control cells were cultured in DMEM (Hyclone;
Cytiva) supplemented with 10% FBS (Gibco; Thermo Fisher Scientific,
Inc.) and 100 µg/ml streptomycin (Gibco; Thermo Fisher Scientific,
Inc.) in a humidified incubator with 5% CO2 at 37˚C
until use. For hypoxic treatment, H9c2 cells were cultured in a
tri-gas incubator (Thermo Fisher Scientific, Inc.) containing 94%
N2, 5% CO2 and 1% O2. Following
incubation for 48 h at room temperature, H9c2 cells were harvested
for cell viability and apoptosis analysis.
Cell viability assay
Cell viability was determined by performing Cell
Counting Kit-8 (CCK-8) assays (Dojindo Molecular Technologies,
Inc.), according to the manufacturer's protocol. Following hypoxic
treatment, transfected cells were seeded (seeding density,
1x105 cells/well) into 96-well plates and cultured with
5% CO2 at room temperature. Following incubation for 12,
24 or 48 h, 10 µl CCK-8 solution was added to each well for 2 h and
incubated at room temperature until analysis. Subsequently, the
absorbance of each well was measured at a wavelength of 450 nm
using a multi-mode microplate reader (Biotek Instruments,
Inc.).
Flow cytometry assay
Cell apoptosis was detected via flow cytometry using
a Annexin V-FITC/propidium iodide (PI) reagent kit (Sigma-Aldrich;
Merck KGaA), according to the manufacturer's protocol. Early and
late apoptosis was assessed accordingly. Following hypoxic
treatment, transfected cells were seeded (seeding density,
3x104 cells/well) into 12-well plates and cultured at 5%
CO2 for 48 h at 37˚C. Subsequently, cells were labeled
with Annexin V-FITC and PI for 10 min in the dark at room
temperature. Cell apoptosis was assessed using a FACSCalibur flow
cytometer (BD Biosciences) and FlowJo software (version no. 10; BD
Biosciences).
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was extracted from serum samples and cells
using TRIzol® (Invitrogen; Thermo Fisher Scientific,
Inc.), according to the manufacturer's protocol. Total RNA (2 µl)
was reverse transcribed into cDNA using a First-strand cDNA
synthesis kit (Invitrogen; Thermo Fisher Scientific, Inc.) for 2 h
at 37˚C. Subsequently, qPCR was performed using a
SYBR®-Green I kit (cat. no. S9430; Sigma-Aldrich; Merck
KGaA) using the ABI Prism 7700 sequence detection system (Applied
Biosystems; Thermo Fisher Scientific, Inc.), according to the
manufacturer's protocol. The following thermocycling conditions
were used for qPCR: Initial denaturation or 3 min at 94˚C; 30
cycles of annealing at 62˚C for 10 sec and extension at 72˚C for 10
sec; and final extension at 72˚C for 10 min. Primers were purchased
from Sigma-Aldrich, Merck KGaA and the sequences used were as
follows: miR-126-5p forward, 5'-GGAATGTAAGGAAGT GTG-3' and reverse,
5'-GAGCAGGCTGGAGAA-3'; IL-17A forward, 5'-TCCCACGAAATCCAGGATGC-3'
and reverse, 5'-GGATGTTCAGGTTGACCATCAC-3'; U6 forward, 5'-CTT
CGGCAGCACATATAC-3' and reverse, 5'-GAACGCTTCACG AATTTGC-3'; and
GAPDH forward, 5'-ACATGTTCCAATAT GATTCC-3' and reverse,
5'-TGGACTCCACGACGTACTCAG-3'. miRNA and mRNA expression levels were
calculated using the 2-ΔΔCq method (21) and normalized to the internal
reference genes U6 and GAPDH, respectively.
Western blotting
Total protein was extracted from serum samples and
cells using RIPA buffer (Sigma-Aldrich; Merck KGaA). Total protein
was quantified using a bicinchoninic acid protein assay kit
(Beyotime Institute of Biotechnology). Proteins (10 µg/lane) were
separated via 10% SDS-PAGE (cat. no. P1200-25T; Beijing Solarbio
Science & Technology Co., Ltd.) and transferred to PVDF
membranes (Beijing Solarbio Science & Technology Co., Ltd.).
The membranes were blocked with 5% non-fat milk in TBST (containing
0.1% Tween-20; cat. no. T1081-500; Beijing Solarbio Science &
Technology Co., Ltd.) for 50 min at room temperature. Subsequently,
the membranes were incubated at 4˚C overnight with the following
primary antibodies: Rabbit anti-IL-17A (1:1,000; cat. no. ab136668;
Abcam), rabbit anti-B-cell lymphoma 2 (Bcl-2; 1:1,000; cat. no.
ab32124; Abcam), rabbit anti-AKT (1:500; cat. no. ab8805; Abcam),
rabbit anti-phosphorylated (p)-AKT (1:1,000; cat. no. 4060; Cell
Signaling Technology, Inc.), rabbit anti-PI3K (1:1,000; cat. no.
ab32089; Abcam), rabbit anti-p-PI3K (1:1,000; cat. no. ab182651;
Abcam), rabbit anti-Bcl-2 associated X protein (Bax; 1:1,000; cat.
no. ab32503; Abcam), rabbit anti-cleaved caspase-3 (1:500; cat. no.
ab13847; Abcam) and rabbit anti-GAPDH (1:2,500; cat. no. ab9485;
Abcam). After washing with 0.1% TBST, the membranes were incubated
with a goat anti-rabbit IgG H&L secondary antibodies (1:1,000;
cat. no. ab6940; Abcam) for 50 min at room temperature. Protein
bands were visualized using an enhanced chemiluminescence kit
(Thermo Fisher Scientific, Inc.). Protein expression levels were
quantified using ImageJ software (version no. 1.16; National
Institutes of Health) with GAPDH as the loading control.
Statistical analysis
Statistical analyses were performed using SPSS
software (version no. 21.0; IBM Corp.). Data are presented as the
mean ± standard deviation. Comparisons between two groups were
analyzed using unpaired Student's t-test. Comparisons among
multiple groups were analyzed using one-way ANOVA followed by
Tukey's post-hoc test. P<0.05 was considered to indicate a
statistically significant difference. All experiments were
performed in triplicate.
Results
miR-126-5p expression is downregulated
and IL-17A expression is upregulated in the serum of patients with
AMI and in H9c2 cells following hypoxic treatment
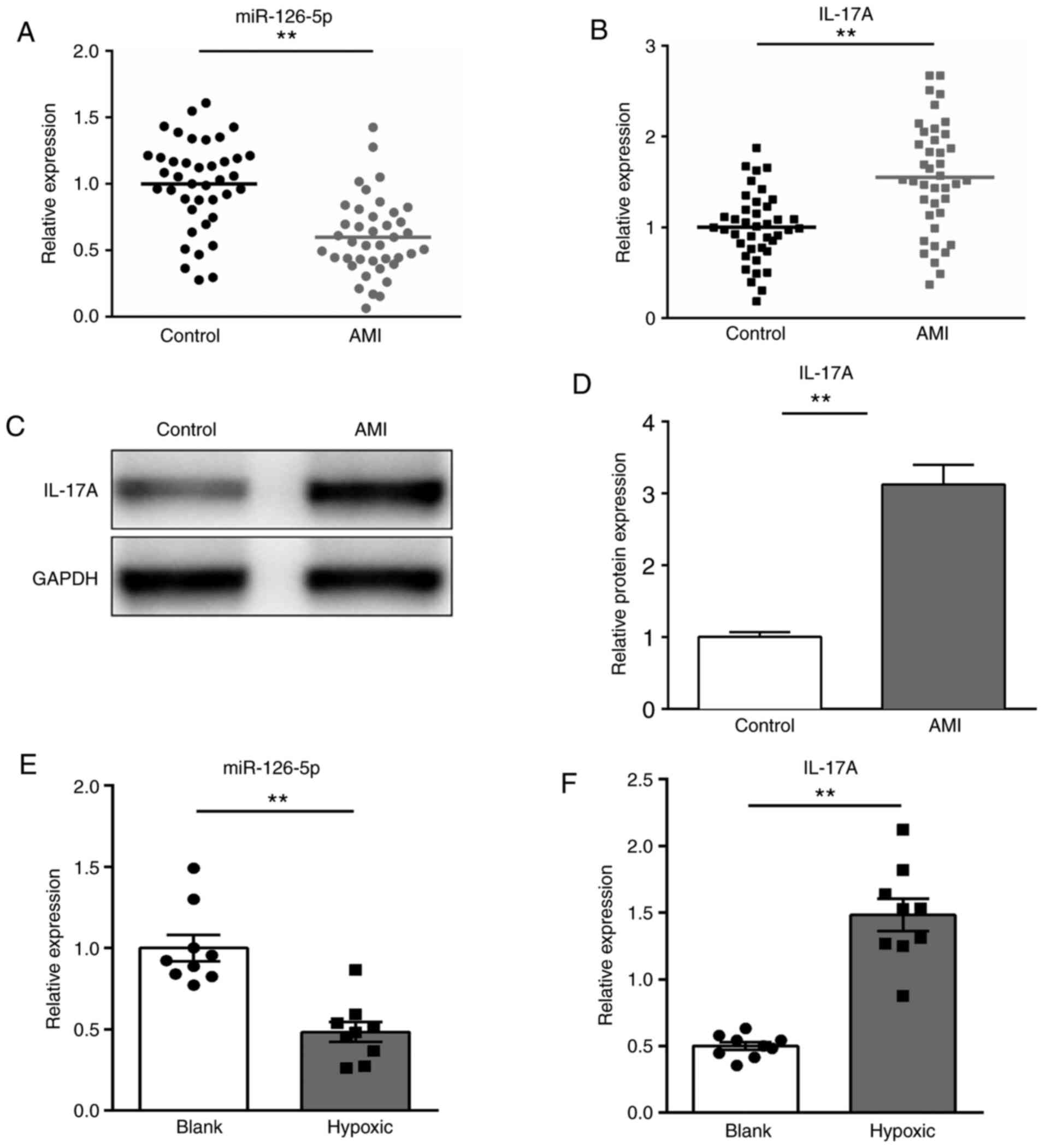
To verify whether miR-126-5p participated in the
pathogenesis of AMI, the expression of miR-126-5p and its predicted
target, IL-17A, in the serum of patients with AMI and healthy
individuals was determined via RT-qPCR. The expression of
miR-126-5p was significantly downregulated, whereas IL-17A mRNA and
protein expression levels were significantly upregulated in
patients with AMI compared with healthy individuals (P<0.01;
Fig. 1A-D). Furthermore, the
expression levels of miR-126-5p and IL-17A in H9c2 cells following
hypoxic treatment were also determined. The expression of
miR-126-5p was significantly decreased, whereas IL-17A expression
levels were significantly increased after hypoxic treatment
compared with the control group (P<0.01; Fig. 1E and F). The results indicated that miR-126-5p
may participate in AMI progression.
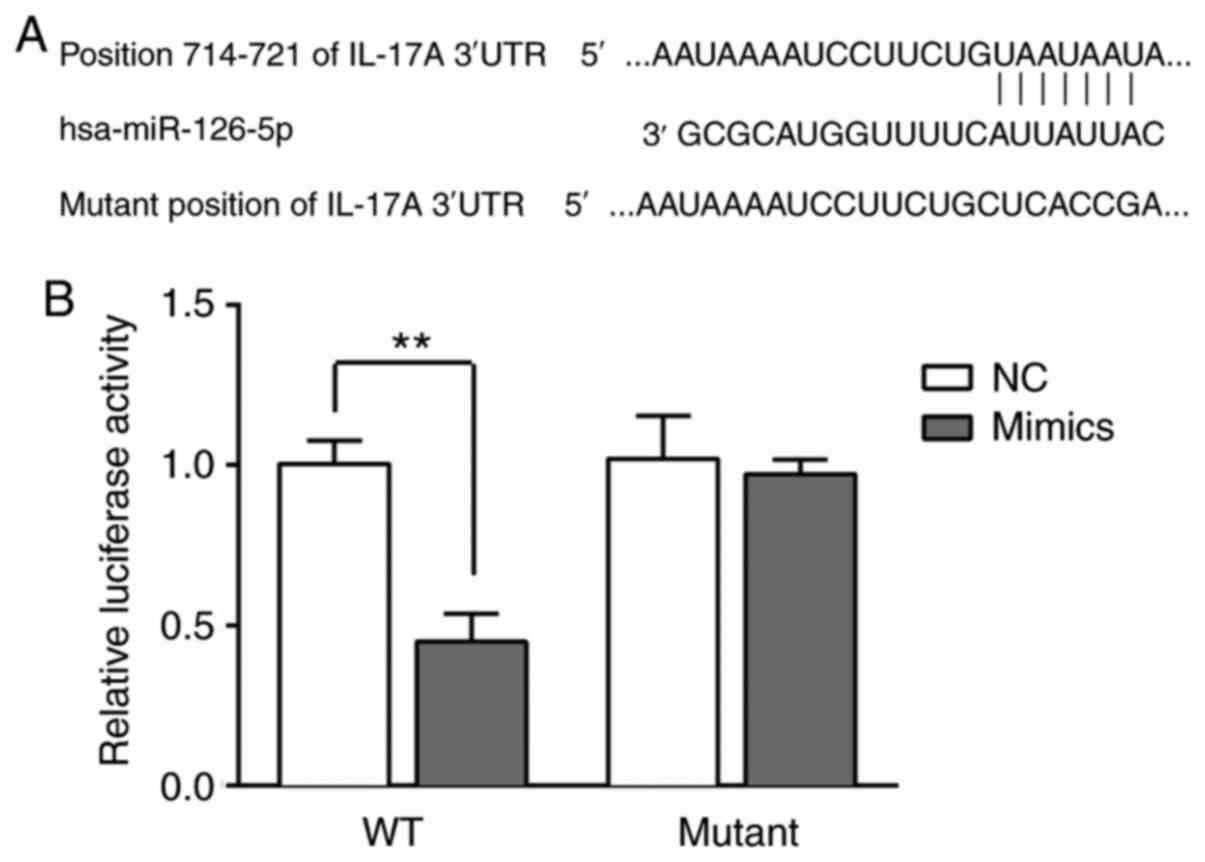
IL-17A is a target of miR-126-5p
The IL-17A 3'UTR contained predicted miR-126-5p
binding sites (Fig. 2A). miR-126-5p
mimics significantly decreased the luciferase activity of wild-type
IL-17A 3'UTR compared with the NC group (P<0.01). However, the
luciferase activity of mutant IL-17A 3'UTR was not significantly
altered by miR-126-5p mimics compared with NC (P>0.05; Fig. 2B).
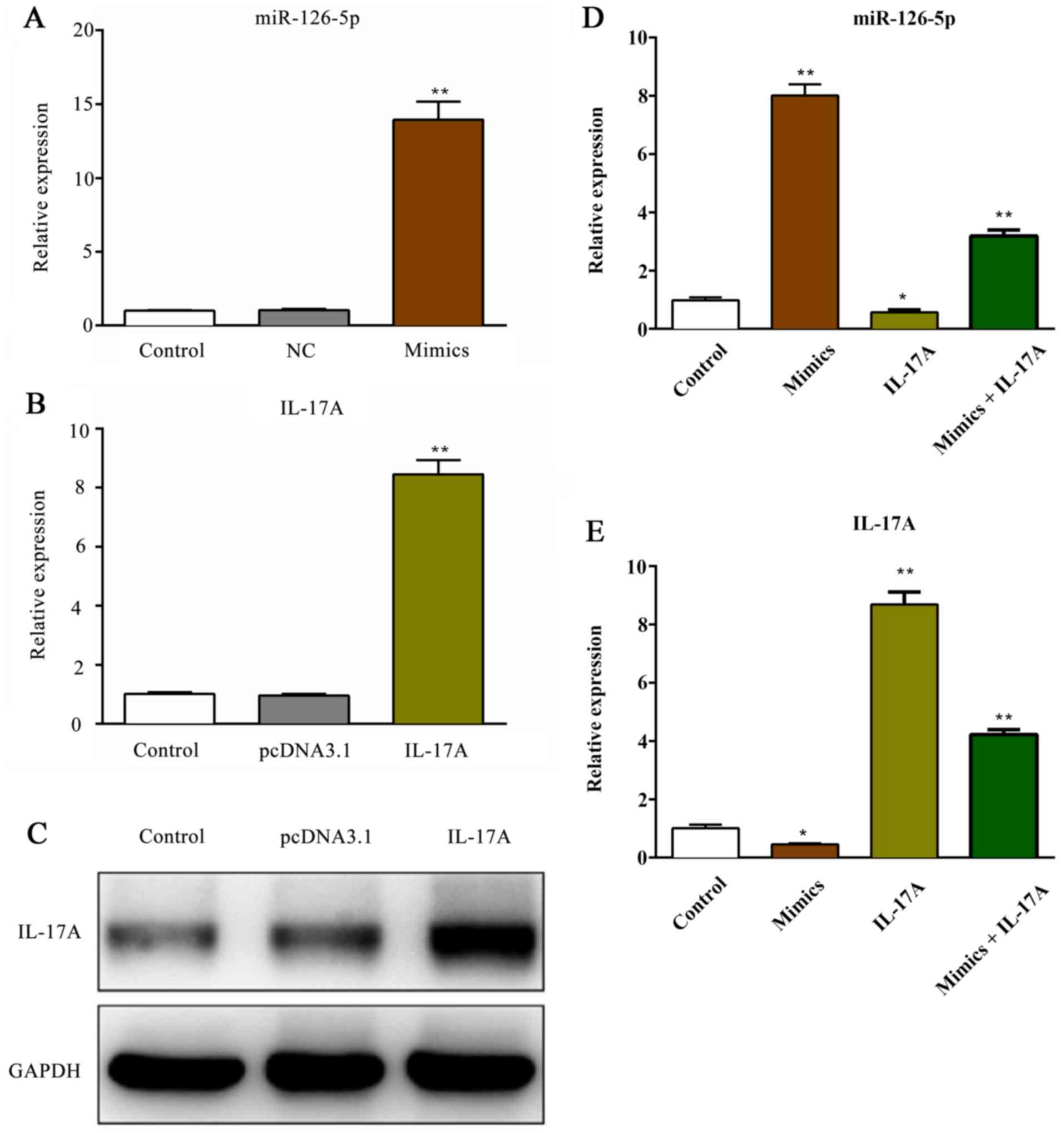
Transfection efficiency of miR-126-5p
and IL-17A
The expression of miR-126-5p was significantly
increased following transfection with miR-126-5p mimics compared
with the NC group (P<0.01). However, the expression of
miR-126-5p was not significantly altered in the NC group compared
with the control group (P>0.05; Fig.
3A). Additionally, the mRNA and protein expression levels of
IL-17A were significantly increased following transfection with
pcDNA3.1-IL-17A vector (P<0.01; Fig.
3B and C) compared with the
pcDNA3.1 group. Following co-transfection with miR-126-5p mimics
and pcDNA3.1-IL-17A, the expression levels of miR-126-5p and IL-17A
were measured. miR-126-5p expression levels were significantly
reduced in the IL-17A group compared with the control group,
whereas co-transfection with miR-126-5p mimics partially reversed
the effects of IL-17A overexpression (P<0.01; Fig. 3D) compared with the IL-17 group.
Meanwhile, IL-17A expression was significantly reduced in the
mimics group compared with the control group, co-transfection with
pcDNA3.1-IL-17 vector partially reversed the effects of miR-126-5p
overexpression (P<0.01; Fig. 3E)
compared with the mimics group.
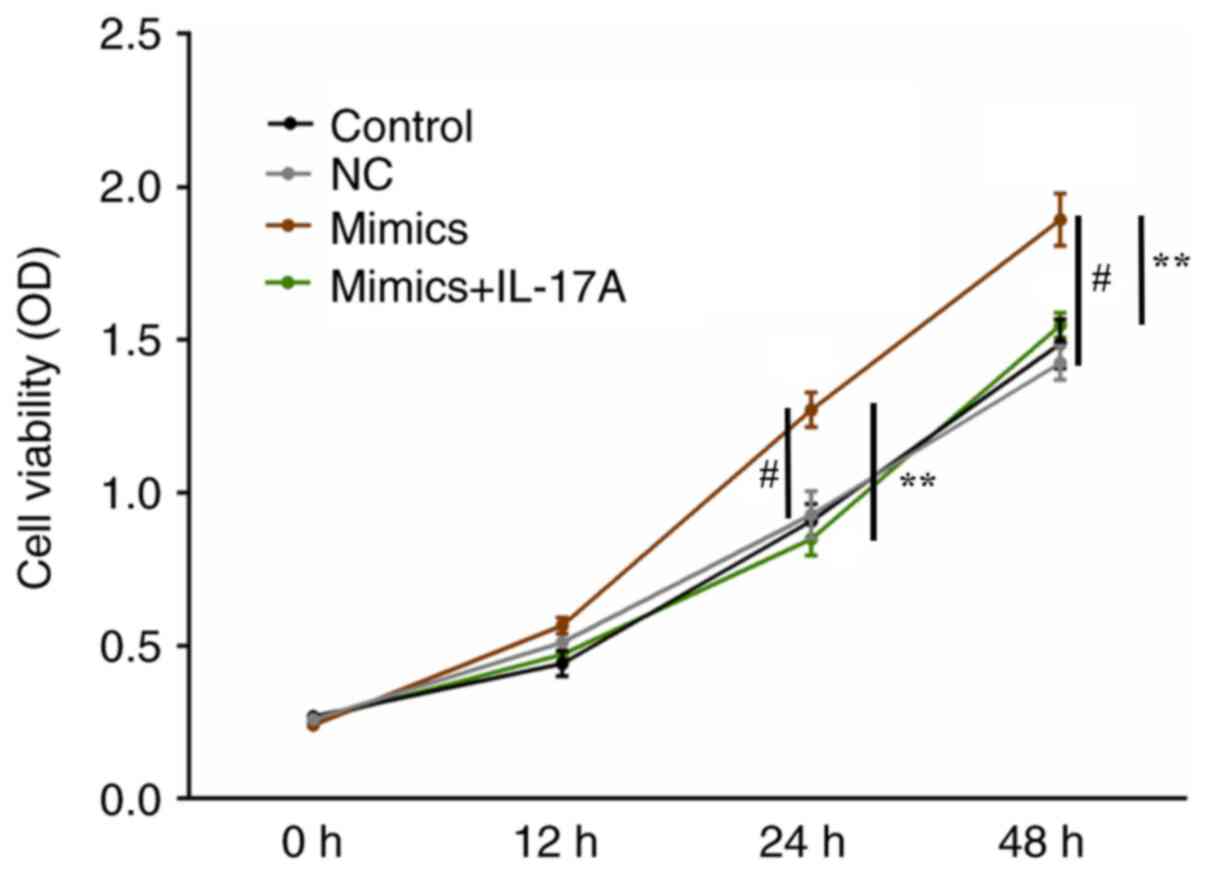
miR-126-5p overexpression promotes
H9c2 cell survival under hypoxic conditions
Under hypoxic conditions, H9c2 cell viability was
significantly increased in the miR-126-5p mimics group compared
with the NC group (P<0.01). The results also indicated that
co-transfection with pcDNA3.1-IL-17A reversed the effects of
miR-126-5p mimics on cell viability compared with the mimics group
(P<0.01; Fig. 4).
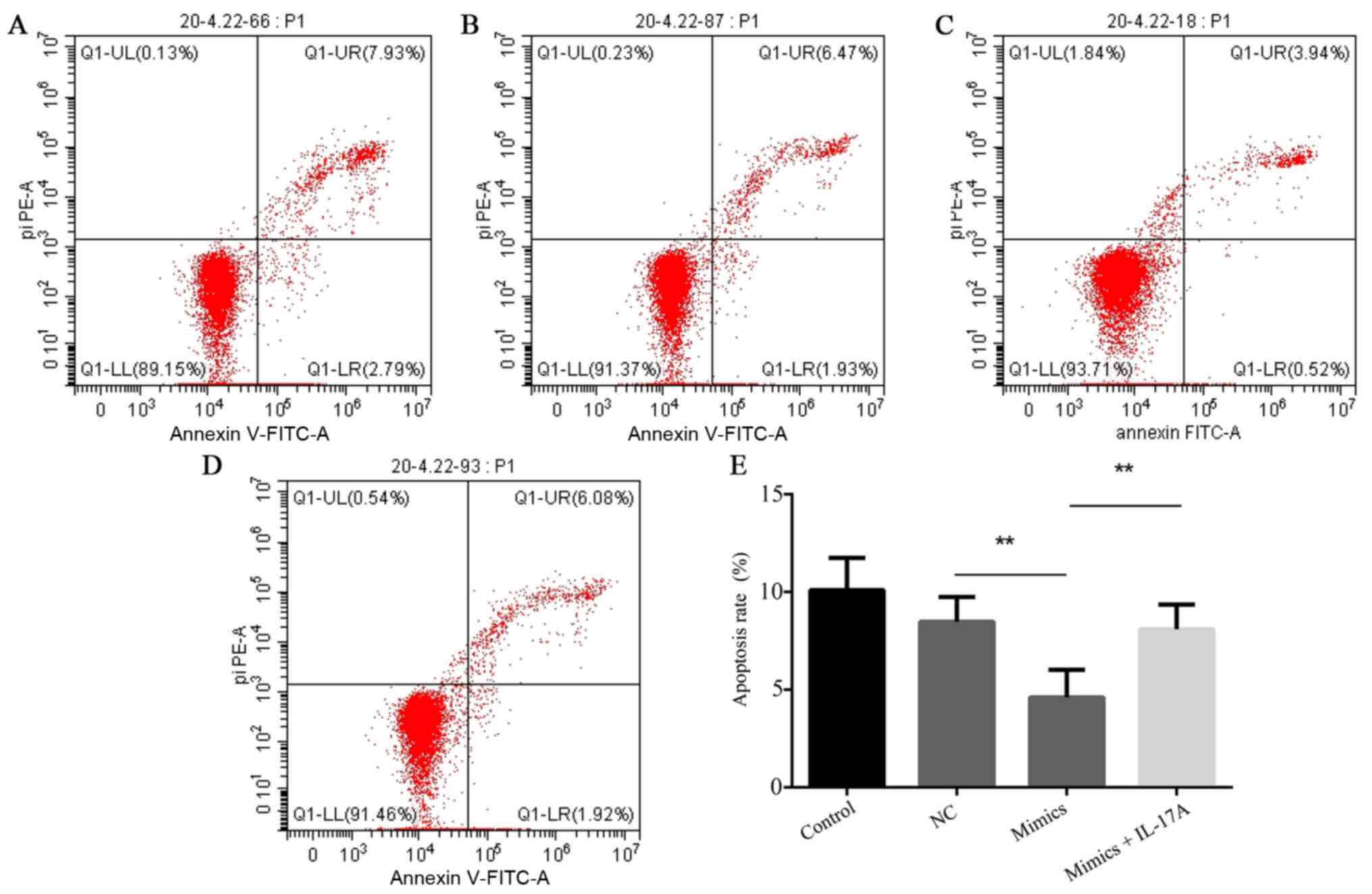
miR-126-5p overexpression suppresses
H9c2 cell apoptosis under hypoxic conditions
Under hypoxic conditions, H9c2 cell apoptosis was
significantly decreased in the miR-126-5p mimics group compared
with the NC group (P<0.01), which was reversed by
co-transfection with pcDNA3.1-IL-17A (P<0.01; Fig. 5).
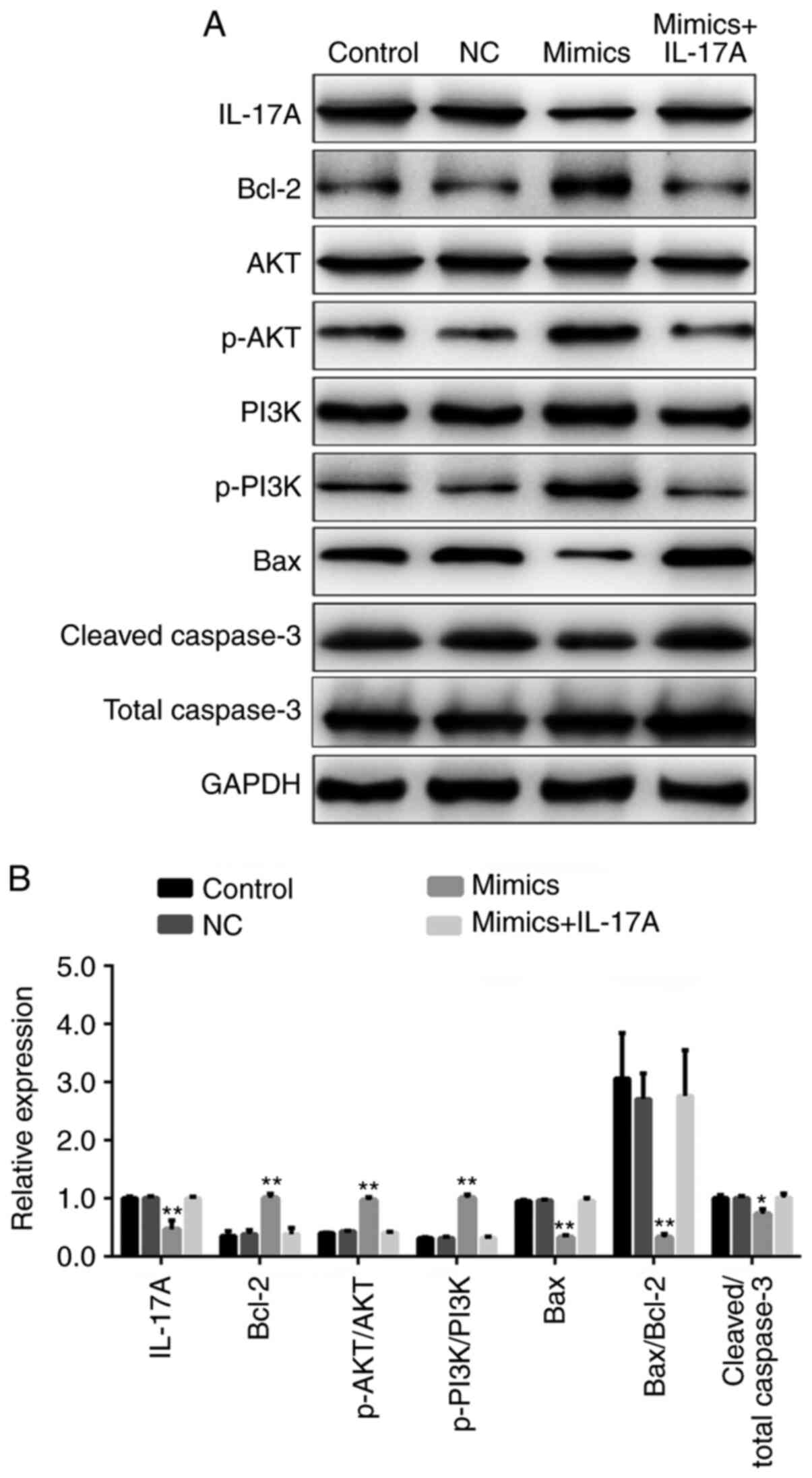
miR-126-5p regulates H9c2 cell
apoptosis via the PI3K/AKT signaling pathway
Under hypoxic conditions, the expression levels of
IL-17A, Bax and cleaved caspase-3 were significantly decreased,
whereas p-PI3K, p-AKT and Bcl-2 were significantly increased in the
miR-126-5p mimics group compared with the NC group (P<0.05).
However, co-transfection with pcDNA3.1-IL-17A reversed miR-126-5p
mimic-mediated effects on protein expression (Fig. 6).
Discussion
The present study indicated that miR-126-5p
expression was significantly reduced in the serum of patients with
AMI compared with healthy volunteers. Previous studies have
demonstrated that miR-126-5p was downregulated in H9c2 cells and
may inhibit H9c2 cell apoptosis, inflammation and angiogenesis
following an antioxidant stimulus (22,23). A
recent study reported that the expression of miR-126-5p was
significantly decreased in patients with AMI (24), which was consistent with the results
of the present study.
IL-17A is a proinflammatory cytokine that is
primarily produced by Th17 cells (25). Accumulating evidence has confirmed
that IL-17A may serve as a potential diagnostic marker for the
development of various acute or chronic diseases (26,27).
Furthermore, the IL-17A gene is highly expressed in patients with
AMI (28). Bcl-2 and Bax are
members of the Bcl-2 family that regulate cell apoptosis by
inhibiting or inducing apoptosis in a wide variety of cellular
activities (29). Previous studies
have reported that Bcl-2 overexpression may suppress
hypoxia-induced H9c2 cell apoptosis (30,31),
whereas Bax knockdown may protect H9c2 cells against
hypoxia-induced apoptosis (32).
Further studies reported that IL-17A inhibition increased the
expression of Bcl-2 and decreased the expression of Bax (33-35).
Caspase-3 is a member of the caspase family and serves a crucial
role in the execution phase of cell apoptosis (36). Caspase-3 knockdown may protect
against hypoxia-induced H9c2 cell apoptosis (37). Several studies have revealed that
cleaved caspase-3 expression could be suppressed by IL-17A
inhibition (38,39). Hence, miR-126-5p may promote H9c2
cell viability and protect against hypoxia-induced H9c2 apoptosis
by downregulating Bax and cleaved caspase-3 expression, and
upregulating Bcl-2 expression, which was in accordance with present
study. PI3K/AKT is an intracellular signaling pathway that is
important for the modulation of the cell cycle and is directly
associated with cell viability and apoptosis (40,41).
Several studies have demonstrated that activation of the PI3K/AKT
signaling pathway attenuates hypoxia-induced H9c2 cell apoptosis
(42-44).
The present study indicated that compared with the NC group,
miR-126-5p overexpression promoted H9c2 cell viability and
protected against hypoxia-induced H9c2 apoptosis by targeting
IL-17A via the PI3K/AKT signaling pathway. To verify the
interaction between miR-126-5p and IL-17A, miR-126-5p and IL-17A
were overexpressed in the same cells in the present study.
Nonetheless, future studies should perform IL-17A knockdown
experiments to further verify the mechanism identified in the
present study.
In conclusion, the present study identified a
potential mechanism underlying AMI development associated with
miR-126-5p. However, further studies are required to confirm the
results of the present study.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
YR, RB, ZG and JK drafted the manuscript. YR, RB and
ZG designed and conceptualized the current study. YR, CGC and ZL
contributed to data acquisition and supervision. JK, CGC and ZL
performed data analysis, interpretation and revised the manuscript.
All authors read and approved the final manuscript.
Ethics approval and consent for
participation
The present study was approved by the Ethics
Committee of Taizhou People's Hospital of Jiangsu Province,
Taizhou, China. Written informed consent was obtained from all
patients and volunteers prior to sample collection.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Orlic D, Kajstura J, Chimenti S, Jakoniuk
I, Anderson SM, Li B, Pickel J, McKay R, Nadal-Ginard B, Bodine DM,
et al: Bone marrow cells regenerate infarcted myocardium. Nature.
410:701–705. 2001.PubMed/NCBI View
Article : Google Scholar
|
|
2
|
Bissessor N and White H: Valsartan in the
treatment of heart failure or left ventricular dysfunction after
myocardial infarction. Vasc Health Risk Manag. 3:425–430.
2007.PubMed/NCBI
|
|
3
|
DeBusk R, Drory Y, Goldstein I, Jackson G,
Kaul S, Kimmel SE, Kostis JB, Kloner RA, Lakin M, Meston CM, et al:
Management of sexual dysfunction in patients with cardiovascular
disease: Recommendations of The Princeton Consensus Panel. Am J
Cardiol. 86:175–181. 2000.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Schaper J and Schaper W: Reperfusion of
ischemic myocardium: Ultrastructural and histochemical aspects. J
Am Coll Cardiol. 1:1037–1046. 1983.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Fein FS, Kornstein LB, Strobeck JE,
Capasso JM and Sonnenblick EH: Altered myocardial mechanics in
diabetic rats. Circ Res. 47:922–933. 1980.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Montgomery RL and van Rooij E: Therapeutic
advances in MicroRNA targeting. J Cardiovasc Pharmacol. 57:1–7.
2011.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Calin GA and Croce CM: MicroRNA signatures
in human cancers. Nat Rev Cancer. 6:857–866. 2006.PubMed/NCBI View
Article : Google Scholar
|
|
8
|
Wu W: MicroRNA: Potential targets for the
development of novel drugs? Drugs RD. 10:1–8. 2010.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Ai J, Zhang R, Li Y, Pu J, Lu Y, Jiao J,
Li K, Yu B, Li Z, Wang R, et al: Circulating microRNA-1 as a
potential novel biomarker for acute myocardial infarction. Biochem
Biophys Res Commun. 391:73–77. 2010.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Pan ZW, Lu YJ and Yang BF: MicroRNAs: A
novel class of potential therapeutic targets for cardiovascular
diseases. Acta Pharmacol Sin. 31:1–9. 2010.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Villain G, Poissonnier L, Noueihed B,
Bonfils G, Rivera JC, Chemtob S, Soncin F and Mattot Y: miR-126-5p
promotes retinal endothelial cell survival through SetD5 regulation
in neurons. Development. 145(dev156232)2018.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Fish JE, Santoro MM, Morton SU, Yu S, Yeh
RF, Wythe JD, Ivey KN, Bruneau BG, Stainier DY and Srivastava D:
miR-126 regulates angiogenic signaling and vascular integrity. Dev
Cell. 15:272–284. 2008.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Wang S, Aurora AB, Johnson BA, Qi X,
McAnally J, Hill JA, Richardson JA, Bassel-Duby R and Olson EN: The
endothelial-specific microRNA miR-126 governs vascular integrity
and angiogenesis. Dev Cell. 15:261–271. 2008.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Long G, Wang F, Duan Q, Chen F, Yang S,
Gong W, Wang Y, Chen C and Wang DW: Human circulating microRNA-1
and microRNA-126 as potential novel indicators for acute myocardial
infarction. Int J Biol Sci. 8:811–818. 2012.PubMed/NCBI View Article : Google Scholar
|
|
15
|
von Stebut E, Boehncke WH, Ghoreschi K,
Gori T, Kaya Z, Thaci D and Schäffler A: IL-18A in psoriasis and
beyond: Cardiovascular and metabolic implications. Front Immunol.
10(3096)2020.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Bai H, Gao X, Zhao L, Peng Y, Yang J, Qiao
S, Zhao H, Wang S, Fan Y, Joyee AG, et al: Respective IL-17A
production by γδ T and Th17 cells and its implication in host
defense against chlamydial lung infection. Cell Mol Immunol.
14:850–861. 2017.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Kuwabara T, Ishikawa F, Kondo M and
Kakiuchi T: The role of IL-17 and related cytokines in inflammatory
autoimmune diseases. Mediators Inflamm.
2017(3908061)2017.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Shaikh SB, Bhat SG and Bhandary YP:
Curcumin attenuates IL-17A mediated pulmonary SMAD dependent and
non-dependent mechanism during acute lung injury in vivo. Mol Biol
Rep. 47:5643–5649. 2020.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Yang P, Qian FY, Zhang MF, Xu AL, Wang X,
Jiang BP and Zhou LL: Th17 cell pathogenicity and plasticity in
rheumatoid arthritis. J Leukoc Biol. 106:1233–1240. 2019.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Huang KD, Shen Y, Wei X, Zhang FQ, Liu YY
and Ma L: Inhibitory effect of microRNA-27b on interleukin 17
(IL-17)-induced monocyte chemoattractant protein-1 (MCP1)
expression. Genet Mol Res. 15(2)2016.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Luo Q, Guo D, Liu G, Chen G, Hang M and
Jin M: Exosomes from miR-126-overexpressing adscs are therapeutic
in relieving acute myocardial ischaemic injury. Cell Physiol
Biochem. 44:2105–2116. 2017.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Li B, Tao Y and Huang Q: Effect and
mechanism of miR-126 in myocardial ischemia reperfusion. Genet Mol
Res. 14:18990–18998. 2015.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Xue S, Liu D, Zhu W, Su Z, Zhang L, Zhou C
and Li P: Circulating miR-17-5p, miR-126-5p and miR-145-3p are
novel biomarkers for diagnosis of acute myocardial infarction.
Front Physiol. 10(123)2019.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Butcher MJ, Wu CI, Waseem T and Galkina
EV: CXCR6 regulates the recruitment of pro-inflammatory
IL-17A-producing T cells into atherosclerotic aortas. Int Immunol.
28:255–261. 2016.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Erbel C, Chen L, Bea F, Wangler S, Celik
S, Lasitschka F, Wang Y, Böckler D, Katus HA and Dengler TJ:
Inhibition of IL-17A attenuates atherosclerotic lesion development
in apoE-deficient mice. J Immunol. 183:8167–8175. 2009.PubMed/NCBI View Article : Google Scholar
|
|
27
|
de Boer OJ, van der Meer JJ, Teeling P,
van der Loos CM, Idu MM, van Maldegem F, Aten J and van der Wal AC:
Differential expression of interleukin-17 family cytokines in
intact and complicated human atherosclerotic plaques. J Pathol.
220:499–508. 2010.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Chen XM, Zhang T, Qiu D, Feng JY, Jin ZY,
Luo Q, Wang XY and Wu XL: Gene expression pattern of TCR repertoire
and alteration expression of IL-17A gene of γδ T cells in patients
with acute myocardial infarction. J Transl Med.
16(189)2018.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Edlich F: BCL-2 proteins and apoptosis:
Recent insights and unknowns. Biochem Biophys Res Commun.
500:26–34. 2018.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Liu Y, Yang L, Yin J, Su D, Pan Z, Li P
and Wang X: MicroRNA-15b deteriorates hypoxia/reoxygenation-induced
cardiomyocyte apoptosis by downregulating Bcl-2 and MAPK3. J
Investig Med. 66:39–45. 2018.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Liu N, Shi YF, Diao HY, Li YX, Cui Y, Song
XJ, Tian X, Li TY and Liu B: MicroRNA-135a regulates apoptosis
induced by hydrogen peroxide in rat cardiomyoblast cells. Int J
Biol Sci. 13:13–21. 2017.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Zhou YL, Sun Q, Zhang L and Li R: miR-208b
targets Bax to protect H9c2 cells against hypoxia-induced
apoptosis. Biomed Pharmacother. 106:1751–1759. 2018.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Olsson Åkefeldt S, Maisse C, Belot A,
Mazzorana M, Salvatore G, Bissay N, Jurdic P, Aricò M,
Rabourdin-Combe C, Henter JI, et al: Chemoresistance of human
monocyte-derived dendritic cells is regulated by IL-17A. PLoS One.
8(e56865)2013.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Sui G, Qiu Y, Yu H, Kong Q and Zhen B:
Interleukin-17 promotes the development of cisplatin resistance in
colorectal cancer. Oncol Lett. 17:944–950. 2019.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Chen X, Yu X, Li X, Li L, Li F, Guo T,
Guan C, Miao L and Cao G: miR-126 targets IL-17A to enhance
proliferation and inhibit apoptosis in high-glucose-induced human
retinal endothelial cells. Biochem Cell Biol. 98(13)2019.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Fan TJ, Han LH, Cong RS and Liang J:
Caspase family proteases and apoptosis. Acta Biochim Biophys Sin
(Shanghai). 37:719–727. 2005.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Jiang YQ, Chang GL, Wang Y, Zhang DY, Cao
L and Liu J: Geniposide prevents hypoxia/Reoxygenation-induced
apoptosis in H9c2 cells: Improvement of mitochondrial dysfunction
and activation of GLP1R and the PI3K/AKT signaling pathway. Cell
Physiol Biochem. 39:407–421. 2016.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Cruz A, Ludovico P, Torrado E, Gama JB,
Sousa J, Gaifem J, Appelberg R, Rodrigues F, Cooper AM, Pedrosa J,
et al: IL-17A promotes intracellular growth of mycobacterium by
inhibiting apoptosis of infected macrophages. Front Immunol.
6(498)2015.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Li N, Gao S, Wang J, Zhu Y and Shen X:
Anti-apoptotic effect of interleukin-17 in a mouse model of
oxygen-induced retinopathy. Exp Eye Res. 187(107743)2019.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Mao Y, Xi L, Li Q, Cai Z, Lai Y, Zhang X
and Yu C: Regulation of cell apoptosis and proliferation in
pancreatic cancer through PI3K/Akt pathway via Polo-like kinase 1.
Oncol Rep. 36:49–56. 2016.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Maurya AK and Vinayak M: PI-103 attenuates
PI3K-AKT signaling and induces apoptosis in murineT-cell lymphoma.
Leuk Lymphoma. 58:1153–1161. 2017.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Liu MH, Li GH, Peng LJ, Qu SL, Zhang Y,
Peng J, Luo XY, Hu HJ, Ren Z, Liu Y, et al: PI3K/Akt/FoxO3a
signaling mediates cardioprotection of FGF-2 against hydrogen
peroxide-induced apoptosis in H9c2 cells. Mol Cell Biochem.
414:57–66. 2016.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Li L, Zhou Y, Li Y, Wang L, Sun L, Zhou L,
Arai H, Qi Y and Xu Y: Aqueous extract of Cortex Dictamni protects
H9c2 cardiomyocytes from hypoxia/reoxygenation-induced oxidative
stress and apoptosis by PI3K/Akt signaling pathway. Biomed
Pharmacother. 89:233–244. 2017.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Zhang Z, Li H, Chen S, Li Y, Cui Z and Ma
J: Knockdown of microRNA-122 protects H9c2 cardiomyocytes from
hypoxia-induced apoptosis and promotes autophagy. Med Sci Monit.
23:4284–4290. 2017.PubMed/NCBI View Article : Google Scholar
|