Introduction
Arteriosclerotic cardiovascular disease (ASCVD) is a
chronic common inflammatory disease mediated by the immune
response, and is closely associated with cytokine imbalance and
inflammatory cell activation (1).
ASCVD-associated mortalities account for >40% of all household
disease mortalities, and are one of the most important factors in
the global morbidity and mortality of different diseases (2). ASCVD is caused by atherosclerosis, a
complex disease associated with several risk factors, such as
hypertension, diabetes inflammation and low-density lipoprotein
cholesterol (3). Currently, the
molecular mechanism underlying atherosclerosis remains unclear, and
is considered to be associated with multiple factors, such as
migration of smooth muscle cells to the intima, blood lamina
activation, endothelial dysfunction and abnormal production of
inflammatory factors (4), among
which vascular endothelial cells have important value in
maintaining the normal structure and function of blood vessels
(5). Thus, understanding how to
protect vascular endothelial cells is important to improve
atherosclerosis, thereby decreasing the risk of ASCVD.
MicroRNAs (miRNAs/miRs) are non-coding RNA molecules
that are involved in the occurrence and development of
atherosclerosis plaques by regulating endothelial cell inflammation
to activate the phenotypic transformation of macrophages (6). In RAW264.7 cells, miR-301a-3p affected
p65 activity by regulating the expression of NF-κB repressing
factor (NKRF), thus regulating the mRNA expression levels of TNF-α,
IL-6 and monocyte chemoattractant protein-1 (MCP-1) (7). High levels of inflammatory factors
impair vascular endothelial function and promote vascular
endothelial cells to secrete adhesion factors, resulting in the
development of atherosclerosis (7).
Notoginsenoside R1 suppresses the NF-κB pathway to alleviate
inflammatory injury of ATDC5 cells induced by lipopolysaccharide
(LPS) by downregulating miR-301a expression (8). Overexpression of miR-301a-3p, induced
by the Japanese encephalitis virus, promotes the inflammatory
response by inhibiting NKRF production (9). Inhibition of miR-301a-3p relieved
LPS-induced chondrogenic cell injury by activating the NF-κB
signaling pathway (10). Thus, it
was speculated that miR-301a-3p may also affect inflammation in
atherosclerosis.
In the present study, the StarBase database
(http://starbase.sysu.edu.cn/index.php) identified the
ubiquitous Krueppel-like factor 7 (KLF7) as a target of
miR-301a-3p. KLF7 is highly expressed in several human tissues
(11). The present study aimed to
determine whether miR-301a-3p promoted oxidative stress,
inflammation and apoptosis in oxidized low-density lipoprotein
(ox-LDL)-induced human umbilical vein endothelial cells (HUVECs) by
decreasing KLF7 expression, aiming to provide the theoretical basis
for the clinical treatment of ASCVD.
Materials and methods
Cell culture and cell induction
HUVECs were purchased from the American Type Culture
Collection and maintained in DMEM supplemented with 10% fetal
bovine serum, 100 g/ml streptomycin and 100 U/ml penicillin (all
from Thermo Fisher Scientific, Inc.) at 37˚C in 5% CO2
and relative saturated humidity. Cells were cultured until fusion
and subsequently passaged at a 1:3 ratio. Cells were used for
experiments once they reached the logarithmic growth phase. HUVECs
were treated with ox-LDL (Bioss) at different concentrations (10,
20, 50, 100 and 200 mg/l) for 24 h.
Cell transfection
Following treatment of ox-LDL for 24 h,
ox-LDL-induced HUVECs were inoculated into six-well plates until
80% confluence. HUVECs were respectively transfected with miRNA
inhibitor-negative control (NC; 5 nM; cat. no. miR2N0000001-1-5;
Guangzhou RiboBio Co., Ltd.), miR-301a-3p inhibitor (5 nM; cat. no.
miR20000688-1-5; Guangzhou RiboBio Co., Ltd.), miRNA mimic-NC (5
nM; cat. no. miR1N0000001-1-5; Guangzhou RiboBio Co., Ltd.),
miR-301a-3p mimic (5 nM; cat. no. miR10000688-1-5; Guangzhou
RiboBio Co., Ltd.), small interfering RNA (si) R-NC (5 nM; cat. no.
siN0000002-1-5; Guangzhou RiboBio Co., Ltd.) and siKLF7 (5 nM; cat.
no. siG000008609A-1-5; Guangzhou RiboBio Co., Ltd.) using
Lipofectamine® 2000 transfection reagent (Invitrogen;
Thermo Fisher Scientific, Inc.) for 48 h at 37˚C. Subsequent
experiments were conducted 48 h after transfection.
Reverse transcription-quantitative
(RT-q)PCR
Following treatment of ox-LDL for 24 h, total RNA
was extracted from HUVECs using TRIzol® reagent
(Invitrogen; Thermo Fisher Scientific, Inc.). Total RNA (1 µg) was
reverse transcribed into cDNA with the Transcriptor First Strand
cDNA Synthesis kit (Roche Molecular Diagnostics) at 42˚C for 30 min
(Takara Biotechnology Co., Ltd.). qPCR was subsequently performed
using SYBR Green Master Mix (Beyotime). The following thermocycling
conditions were used for the qPCR: Initial denaturation at 95˚C for
30 sec; followed by 35 cycles of 95˚C for 20 sec, 60˚C for 30 sec
and 72˚C for 40 sec. The following primer pairs were used for the
qPCR: miR-301a-3p forward, 5'-ACACTCCAGCTGGGCAGTGCAATAGTATTGTC-3'
and reverse, 5'-CTCAACTGGTGTCGTGGA-3'; U6 forward,
5'-CTCGCTTCGGCAGCACA-3' and reverse 5'-AACGCT TCACGAATTTGCGT-3';
KLF7 forward, 5'-TTCCTGGCA GTCATCTGCAC-3' and reverse,
5'-GGGTCTGTTTGTTTG TCAGTCTGTC-3' and GAPDH forward, 5'-CCACAGTCC
ATGCCATCAC-3' and reverse, 5'-GCTTCACCACCT TCTTGATG-3'. Relative
expression levels were calculated using the 2-ΔΔCq
method (12) and normalized to the
internal reference gene GAPDH and U6.
Cell Counting Kit-8 (CCK-8) assay
Following treatment of ox-LDL for 24 h, HUVECs were
seeded into 96-well plates (5x104 cells/well) and
incubated with 10 µl CCK-8 solution (Beyotime Institute of
Biotechnology) at 37˚C for 4 h. The blank control well was zeroed,
and the absorbance was measured at a wavelength of 450 nm.
Lactate dehydrogenase (LDH)
leakage
Following ox-LDL induction or transfection for 24 h,
HUVECs were seeded into 96-well plates (5x104
cells/well) and LDH leakage was assessed using the LDH assay kit
(cat. no. C0016; Beyotime Institute of Biotechnology). The
supernatant was collected, and the absorbance was measured at a
wavelength of 450 nm using a Thermomax microplate reader (Thermo
Fisher Scientific, Inc.). The LDH leakage rate was calculated as
follows: LDH leakage (%) = (ODtreatment -
ODcontrol)/(ODstandard -
ODblank).
Detection of MCP-1 and IL-6
Following ox-LDL induction or transfection for 24 h,
the supernatant was collected, and MCP-1 and IL-6 secretion were
measured using MCP-1 (cat. no DCP00) and IL-6 (cat. no. D6050)
ELISA kits (both purchased from R&D Systems, Inc.).
Reactive oxygen species (ROS)
activity
Intracellular generation of ROS was assessed via the
2',7'-dichlorofluorescin diacetate (DCFH-DA) assay (Beyotime
Institute of Biotechnology). HUVECs were incubated with DCFH-DA at
37˚C for 30 min in the dark. Fluorescence was subsequently measured
using a fluorescence microplate reader.
Superoxide dismutase (SOD)
activity
HUVECs maintained in PBS were centrifuged at 1,500 x
g for 10 min at 4˚C. SOD activity in the supernatant was
subsequently detected using the SOD assay kit (cat. no. S0086;
Beyotime Institute of Biotechnology) at a wavelength of 450 nm
using a Thermomax microplate reader (Thermo Fisher Scientific,
Inc.).
Flow cytometric analysis
Following ox-LDL induction or transfection for 24 h,
HUVECs were digested using trypsin (Beyotime Institute of
Biotechnology) and subsequently adjusted to 1x106
cells/ml using ice pre-cooled PBS. The cell suspension (1 ml) was
centrifuged at 1,500 x g for 10 min at 4˚C and the cell precipitate
was collected, which was added to 200 µl binding buffer (Beyotime
Institute of Biotechnology), 5 µl Annexin V-FITC (Beyotime
Institute of Biotechnology) and 5 µl propidium iodide (Beyotime
Institute of Biotechnology) at room temperature for 15 min. Binding
buffer (300 µl) was added to each flow tube. A flow cytometry
software (BD FACSDiva software v6.1.3; BD Biosciences) and CYTOMICS
FC 500 flow cytometer (Beckman Coulter, Inc.) were used to detect
cell apoptosis within 1 h.
Western blotting
Following ox-LDL induction or transfection for 24 h,
HUVECs were treated with RIPA lysate (Beyotime Institute of
Biotechnology), which were centrifuged at 1,500 x g for 10 min at
4˚C to extract total protein. Total protein was quantified using a
BCA kit (cat. no. P0012S; Beyotime Institute of Biotechnology). A
total of 30 µg protein/lane was separated by 10% SDS-PAGE. The
separated proteins were transferred onto polyvinylidene fluoride
membranes and blocked with 5% skim milk powder for 1 h at room
temperature. The membranes were incubated with primary antibodies
against: IL-6 (cat. no. ab233706; 1:1,000; Abcam), MCP-1 (cat. no.
ab214819; 1:1,000; Abcam), Bcl2 (cat. no. ab182858; 1:2,000;
Abcam), Bax (cat. no. ab32503; 1:1,000; Abcam), poly (ADP-ribose)
polymerase (PARP; cat. no. ab191217; 1:1,000; Abcam), cleaved PARP
(cat. no. ab32064; 1:1,000; Abcam), pro-caspase3 (cat. no. ab32150;
1:1,000; Abcam), cleaved caspase-3 (cat. no. ab32042; 1:500; Abcam)
and GAPDH (cat. no. ab8245; 1:1,000; Abcam) overnight at 4˚C.
Membranes were washed with PBS three times and subsequently
incubated with horseradish peroxidase-conjugated secondary antibody
(cat. no. 7074; 1:1,000; Cell Signaling Technology, Inc.) at room
temperature for 1 h. Protein bands were visualized using ECL
reagent (EMD Millipore) and imaged on a Chemiluminescence Imaging
system (Tanon Science and Technology Co., Ltd.). Image-Pro Plus
software (version 6.0; Media Cybernetics, Inc.) was used for
densitometry.
Dual-luciferase reporter assay
HUVECs were co-transfected with pGL3-KLF7
3'-untranslated region (UTR) plasmid (containing mutant KLF7 3'-UTR
or wild-type KLF7 3'-UTR) and miR-301a-3p mimic or miR-NC vector
(Biovector NTCC Inc.) using Lipofectamine® 2000 reagent.
Following incubation for 48 h at 37˚C, a
Dual-Luciferase® Reporter Assay system (Promega
Corporation) was used to detect relative luciferase activities,
which was normalized to Renilla luciferase activity.
Statistical analysis
Three separate experiments were repeated for each
experiment. Statistical analysis was performed using SPSS 22.0
software (IBM Corp.). Data are presented as the mean ± SD. Unpaired
Student's t-test was used to compare differences between two
groups, while one-way ANOVA and Tukey's post hoc test were used to
compare differences between multiple groups. P<0.05 was
considered to indicate a statistically significant difference.
Results
miR-301a-3p is highly expressed in
ox-LDL-induced HUVECs
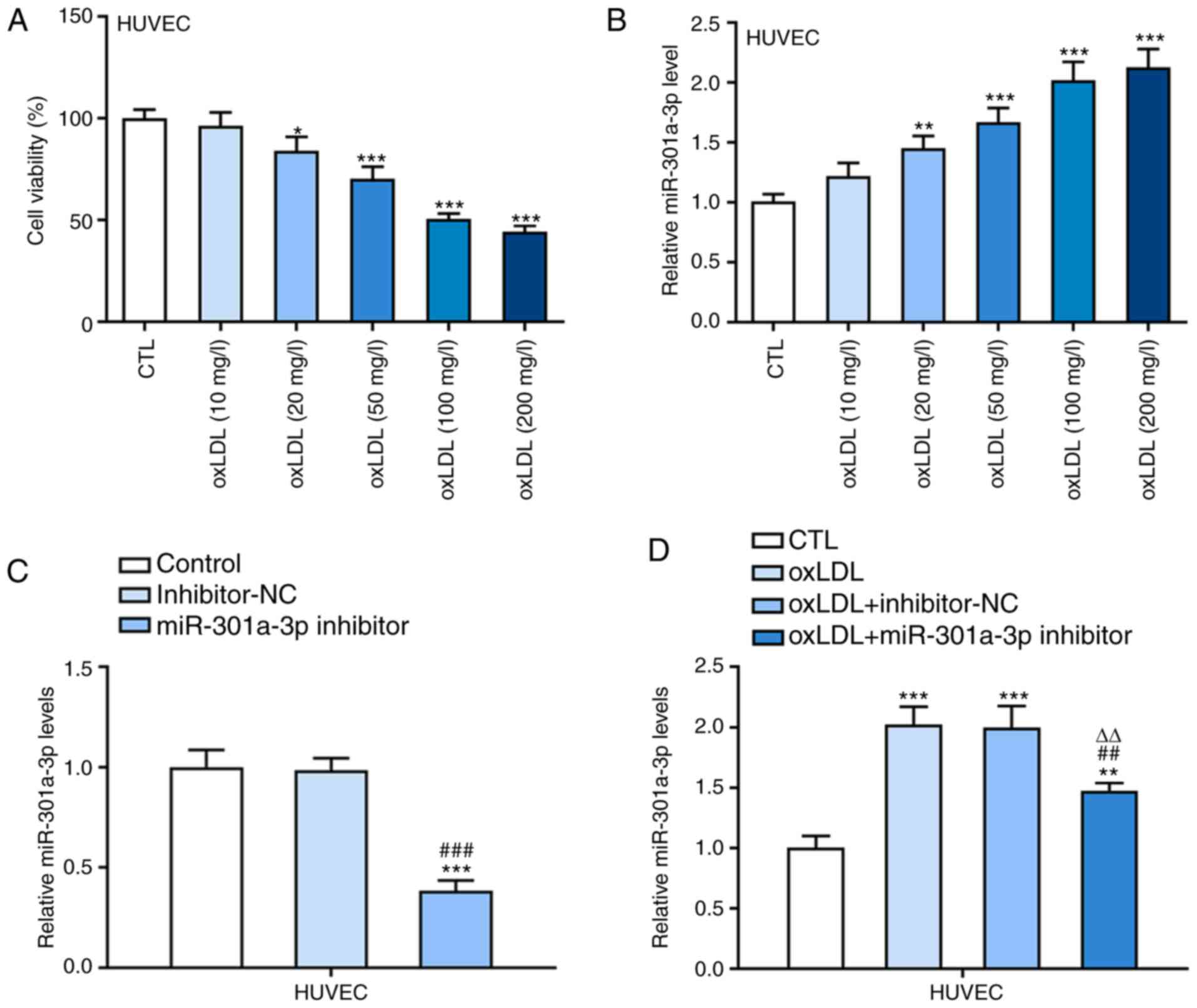
Following treatment with ox-LDL at different
concentrations, the viability of HUVECs gradually decreased upon
decreasing ox-LDL concentration (10-200 mg/l; Fig. 1A). Furthermore, miR-301a-3p
expression gradually increased in ox-LDL-induced HUVECs treated
with 10-100 mg/l ox-LDL (Fig. 1B).
No significant differences were observed in cell viability and
miR-301a-3p expression between 100 mg/l ox-LDL-induced HUVECs and
200 mg/l ox-LDL-induced HUVECs. Hence, 100 mg/l ox-LDL was selected
for subsequent experimentation. miR-301a-3p expression level was
notably decreased in HUVECs transfected with miR-301a-3p inhibitor
(Fig. 1C). As presented in Fig. 1D, miR-301a-3p expression
significantly increased in ox-LDL-induced HUVECs compared with
controls, while transfection with miR-301a-3p inhibitor
significantly downregulated miR-301a-3p expression in
ox-LDL-induced HUVECs.
miR-301a-3p knockdown inhibits
inflammation and oxidative stress in ox-LDL- induced HUVECs
ox-LDL significantly promoted the protein expression
levels of IL-6 and MCP-1, the effects of which were reversed
following miR-301a-3p knockdown (Fig.
2A). Similarly, IL-6 and MCP-1 expression levels increased in
ox-LDL-induced HUVECs, the effects of which were reversed following
miR-301a-3p knockdown (Fig. 2B). In
addition, ROS and SOD levels were upregulated in ox-LDL-induced
HUVECs, the effects of which were reversed following miR-301a-3p
knockdown (Fig. 2C and D).
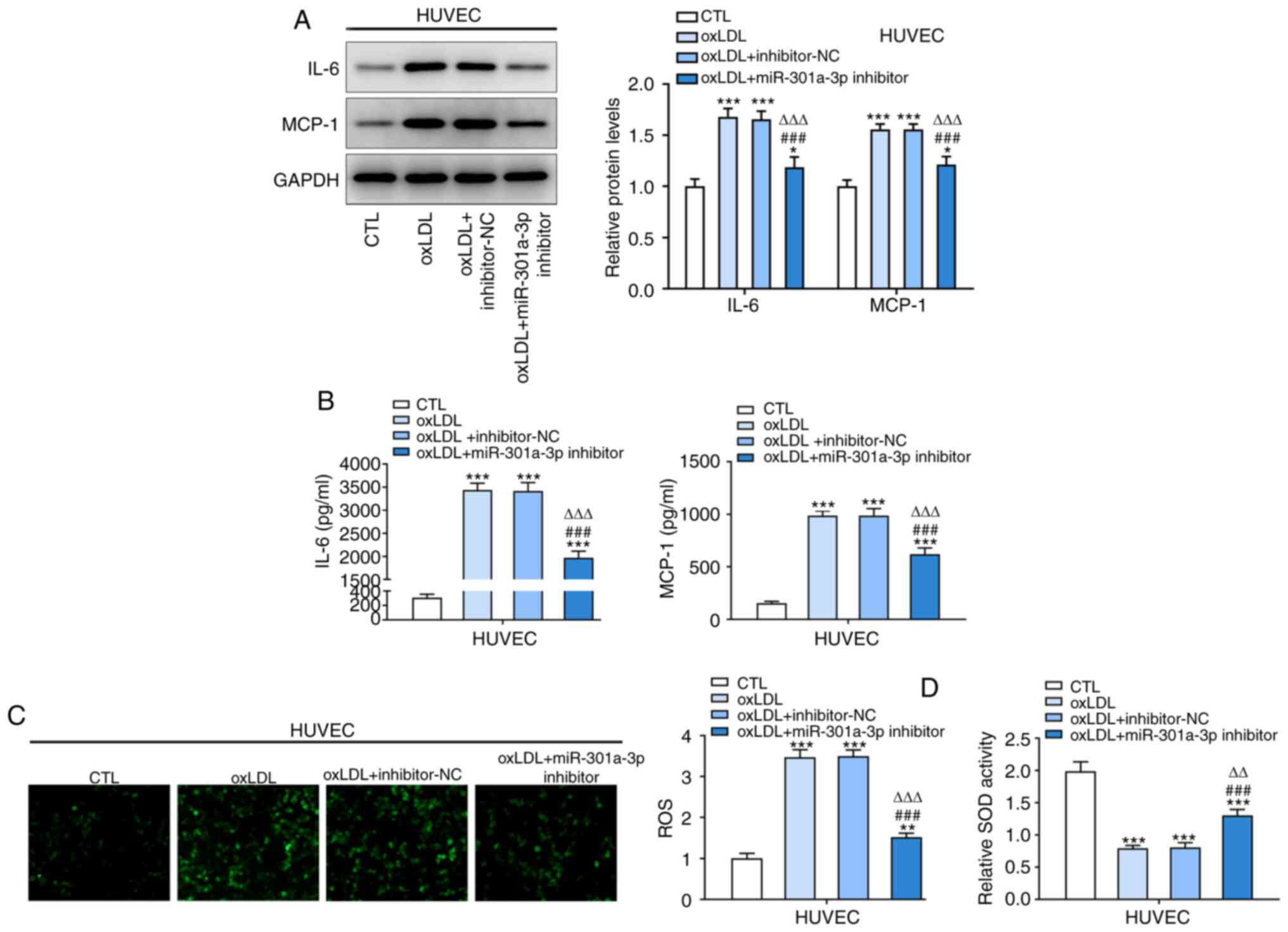 | Figure 2Inhibition of miR-301a-3p inhibits
inflammation and oxidative stress in ox-LDL-induced HUVECs. (A) The
expression of IL-6 and MCP-1 in HUVECs treated with ox-LDL and
transfected with miR-301a-3p inhibitor was analyzed by western blot
analysis. (B) The levels of IL-6 and MCP-1 in HUVECs treated with
ox-LDL and transfected with miR-301a-3p inhibitor was analyzed by
ELISA. (C) ROS level in HUVECs treated with ox-LDL and transfected
with miR-301a-3p inhibitor was determined by a
2',7'-dichlorofluorescin diacetate assay (magnification, x200). (D)
The SOD levels in HUVECs treated with ox-LDL and transfected with
miR-301a-3p inhibitor was determined by a SOD assay kit.
*P<0.05, **P<0.01 and
***P<0.001 vs. CTL group. ###P<0.001
vs. oxLDL group. ∆∆P<0.01 and ∆∆∆P<0.01
vs. oxLDL + miR-NC group. miR, microRNA; HUVEC, human umbilical
vein endothelial cells; oxLDL, oxidized low-density lipoprotein;
CTL, control; NC, negative control; MCP-1, monocyte chemoattractant
protein-1; ROS, reactive oxygen species; SOD, superoxide
dismutase. |
miR-301a-3p knockdown inhibits
apoptosis in ox-LDL-induced HUVECs
LDH leakage was increased and cell viability was
decreased in ox-LDL-induced HUVECs compared with the control group
(Fig. 3A and B). In addition, the expression levels of
Bax, cleaved PARP and cleaved caspase-3 was increased while Bcl-2
expression was decreased in ox-LDL-induced HUVECs (Fig. 3C). The apoptosis ratio of
ox-LDL-induced HUVECs was increased compared with controls
(Fig.3D and E). In addition, miR-301a-3p knockdown
reversed the aforementioned changes in HUVECs induced by
ox-LDL.
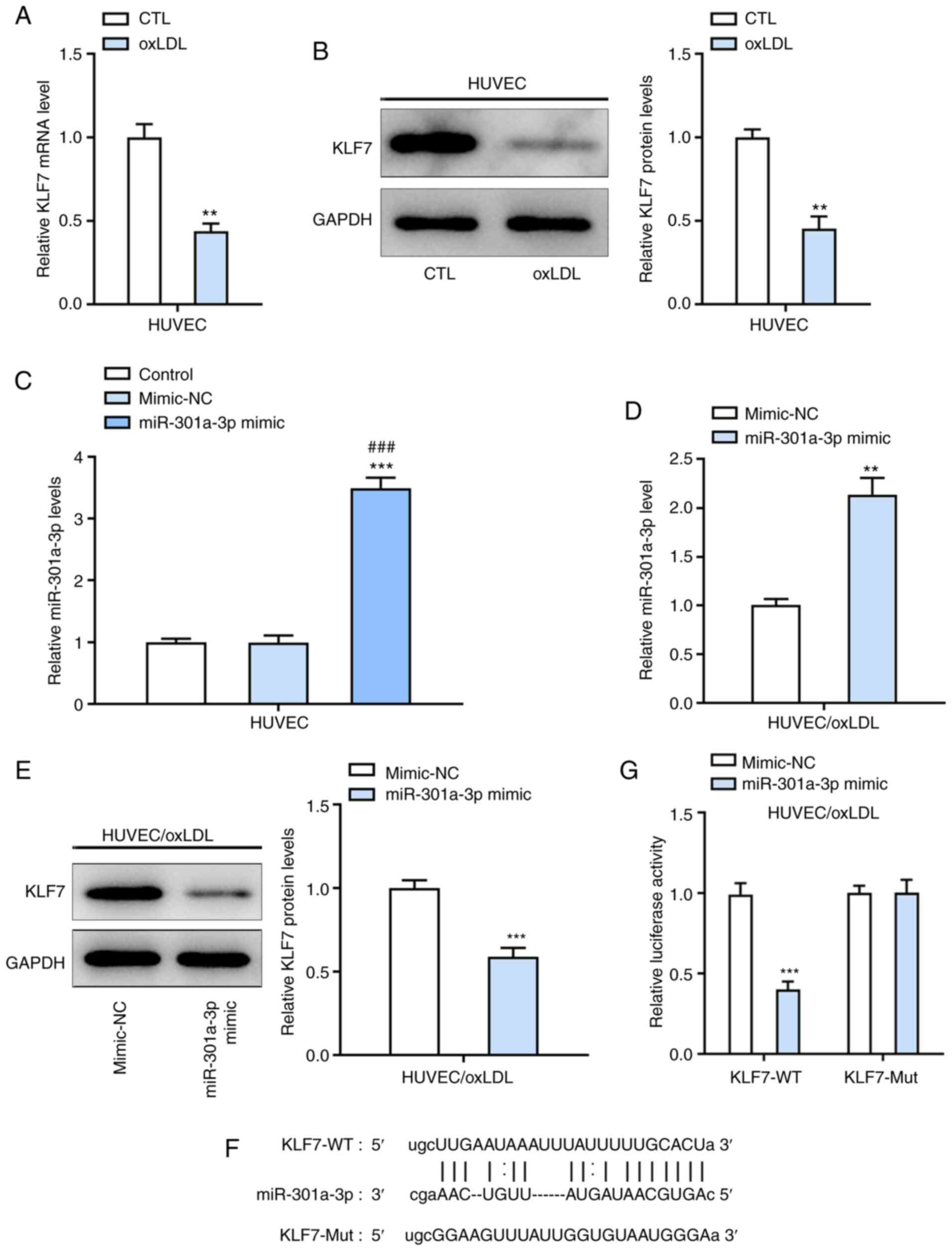
miR-301a-3p targets KLF7 in
ox-LDL-induced HUVECs
Both mRNA and protein levels of KLF7 were
significantly decreased in ox-LDL-induced HUVECs compared with
controls (Fig. 4A and B). Following transfection with miR-301a-3p
mimic, miR-301a-3p expression significantly increased in
ox-LDL-induced HUVECs compared with the miR-NC group (Fig. 4C). In addition, overexpression of
miR-301a-3p suppressed KLF7 expression in ox-LDL-induced HUVECs
(Fig. 4D and E). The binding sites between miR-301a-3p
and KLF7 are presented in Fig. 4F.
Relative luciferase activity significantly decreased in
ox-LDL-induced HUVECs co-transfected with miR-301a-3p mimic and
KLF7-WT (Fig. 4G).
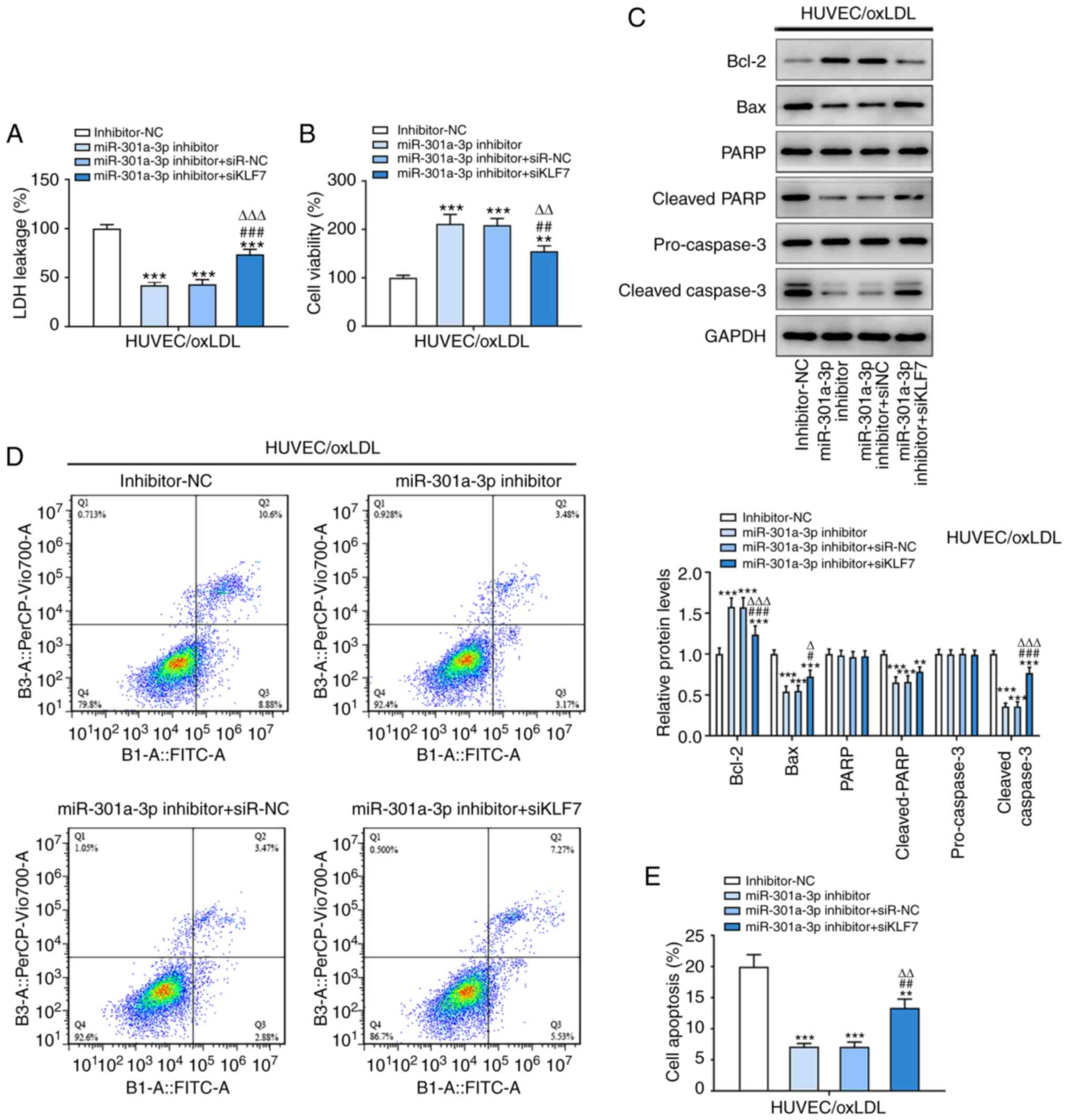
Inhibition of KLF7 weakens the
inhibitory effects of miR-301a-3p inhibition on inflammation and
oxidative stress in ox-LDL-induced HUVECs
KLF7 expression decreased following transfection of
ox-LDL-induced HUVECs with siKLF7 (Fig.
5A and B). miR-301a-3p
knockdown downregulated the protein expression levels of IL-6 and
MCP-1 in ox-LDL-induced HUVECs (Fig.
5C and D). In addition,
miR-301a-3p knockdown downregulated ROS expression and upregulated
SOD expression in ox-LDL-induced HUVECs (Fig. 5E-G). Collectively, these results
suggested that inhibition of KLF7 may reverse the inhibitory
effects of miR-301a-3p on ox-LDL-induced HUVECs.
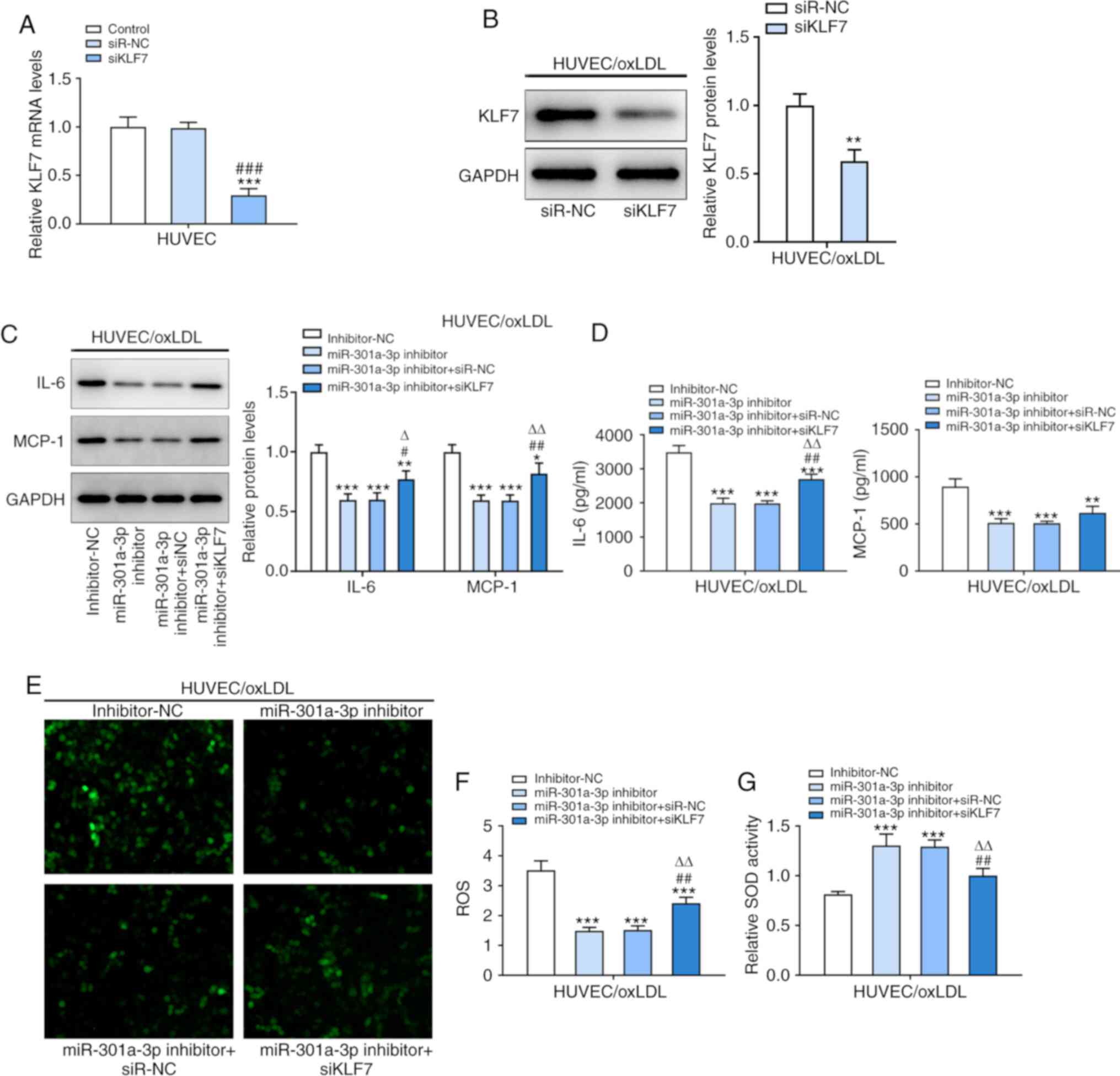 | Figure 5Inhibition of KLF7 weakens the
inhibitory effects of miR-301a-3p inhibition on inflammation and
oxidative stress in ox-LDL-induced HUVECs. (A) KLF7 protein
expression in ox-LDL-induced HUVEC following transfection of
miR-301a-3p inhibitor transfection and siKLF7 was analyzed by
western blot analysis. ***P<0.001 vs. Control group.
###P<0.001 vs. siR-NC group. (B) The expression of
KLF7 in ox-LDL-induced HUVECs following transfection of siKLF7 was
analyzed by western blot analysis. **P<0.01 vs.
siR-NC group. (C) The levels of IL-6 and MCP-1 in ox-LDL-induced
HUVECs following transfection of miR-301a-3p inhibitor and siKLF7
was analyzed by western blot analysis. (D) The levels of IL-6 and
MCP-1 in ox-LDL-induced HUVECs following transfection of
miR-301a-3p inhibitor and siKLF7 was analyzed by ELISA. (E and F)
The ROS level in ox-LDL-induced HUVECs following transfection of
miR-301a-3p inhibitor and siKLF7 was determined by a
2',7'-dichlorofluorescin diacetate assay. (G) The SOD levels in
ox-LDL-induced HUVECs following transfection of miR-301a-3p
inhibitor and siKLF7 was determined by a SOD assay kit.
*P<0.05, **P<0.01 and
***P<0.001 vs. miR-NC group. #P<0.05
and ##P<0.01 vs. miR-301a-3p inhibitor group.
∆P<0.05 and ∆∆P<0.01 vs. miR-301a-3p
inhibitor + siR-NC group. miR, microRNA; HUVEC, human umbilical
vein endothelial cells; oxLDL, oxidized low-density lipoprotein;
CTL, control; NC, negative control; KLF7, Krueppel-like factor 7;
MCP-1, monocyte chemoattractant protein-1; ROS, reactive oxygen
species; SOD, superoxide dismutase; si, small interfering. |
Inhibition of KLF7 weakens the
inhibitory effects of miR-301a-3p inhibition on apoptosis in
ox-LDL-induced HUVECs
miR-301a-3p knockdown decreased LDH leakage and
increased cell viability in ox-LDL-induced HUVECs (Fig. 6A and B). In addition, the expression levels of
Bax, cleaved PARP and cleaved caspase-3 was decreased, while Bcl-2
expression was increased in ox-LDL-induced HUVECs transfected with
miR-301a-3p inhibitor (Fig. 6C),
and inhibition of miR-301a-3p suppressed the ratio of apoptosis of
ox-LDL-induced HUVECs (Fig. 6D and
E). In addition, inhibition of KLF7
could reverse the aforementioned changes in ox-LDL-induced HUVECs
caused by miR-301a-3p inhibition.
Discussion
In the present study, an ox-LDL-induced HUVEC model
was used to simulate atherosclerosis that leads to ASCVD. It was
found that inhibition of miR-301a-3p suppressed oxidative stress,
inflammation and apoptosis in ox-LDL-induced HUVEC, which was
reversed by KLF7 inhibition. Studies have demonstrated that >50%
of cardiovascular diseases, including atherosclerosis, are
inextricably associated with genetics (13,14).
miRNAs are involved in regulating several physiological and
pathological mechanisms, including inflammation and cardiovascular
disease (15).
Several studies have suggested that miRNAs play an
important role in the progression of atherosclerosis and
inflammatory response. For example, miR-21 expression is
significantly increased in the miRNA expression profile of
atherosclerosis (16). miR-21 is
abundantly expressed in macrophages, the lack of which aggravates
the progression of atherosclerosis (17). miR-21 decreased IL-6 expression via
the Toll-like receptor 4 and NF-κB pathways, while increasing IL-10
expression to decrease LPS-induced lipid deposition in macrophages.
Hence, miR-21 may reverse the bacterial infection-induced
pathological process of atherosclerosis (18). In addition, severe burns inhibit
miR-301a-3p expression, which suppresses the apoptosis of HUVECs
and stabilizes endothelial barrier permeability (19). miR-301a-3p expression is upregulated
in obesity-related inflammation, and miR-301a decreases the
production of proinflammatory cytokines in 3T3-L1 cells (20). miR-301a-3p boosts intestinal mucosal
inflammation by increasing the expression levels of IL-17A and
TNF-α in inflammatory bowel disease (21). The results of the present study
demonstrated that miR-301a-3p expression was increased in
ox-LDL-induced HUVECs, and inhibition of miR-301a-3p suppressed
oxidative stress, inflammation and apoptosis in ox-LDL-induced
HUVECs.
Macrophages participate in the immune response of
the vascular wall, and thus promote the formation and development
of atherosclerosis (22). KLF7
expression is downregulated in some injury diseases, thus KLF7 may
inhibit disease progression. Inhibition of KLF7 notably suppressed
the proliferation and migration of Schwann cells (23). Adult corticospinal tract (CST)
neurons failed to promote KLF7 expression due to axon injury, while
overexpression of VP16-KLF7 in vivo promoted both sprouting and
regenerative axon growth in the CST of adult mice (24). The results of the present study
demonstrated that KLF7 expression decreased in ox-LDL-induced
HUVECs, and inhibition of KLF7 reversed the effect of inhibition of
miR-301a-3p and promoted oxidative stress, inflammation and
apoptosis in ox-LDL-induced HUVECs.
In conclusion, the results of the present study
demonstrated that miR-301a-3p expression was upregulated and KLF7
expression was downregulated in ox-LDL-induced HUVECs. In addition,
miR-301a-3p induced oxidative stress, inflammation and apoptosis in
ox-LDL-induced HUVECs by decreasing KLF7 expression. However, the
present study only performed cell experiments. In vivo experiments
will be conducted in the future to support the present results.
Acknowledgements
Not applicable.
Funding
Funding: This study was supported by the Fujian Provincial
Health and Family Planning Project Funding Plan for Rural and Urban
Communities to Promote Appropriate Technology in 2018 (grant no.
2018025).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
JL designed the experiments. HJ performed the
experiments and collected the data. HJ analyzed the experimental
data and drafted the manuscript. JL revised the manuscript. JL and
HJ confirm the authenticity of the raw data. All authors read and
approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare they have no competing
interests.
References
|
1
|
Brown WV, Remaley AT and Ridker PM: JCL
Roundtable: is inflammation a future target in preventing
arteriosclerotic cardiovascular disease. J Clin Lipidol. 9:119–128.
2015.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Zhou M, Wang H, Zeng X, Yin P, Zhu J, Chen
W, Li X, Wang L, Wang L, Liu Y, et al: Mortality, morbidity, and
risk factors in China and its provinces, 1990-2017: A systematic
analysis for the Global Burden of Disease Study 2017. Lancet.
394:1145–1158. 2019.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Roth EM: Alirocumab for low-density
lipoprotein cholesterol lowering. Future Cardiol. 15:17–29.
2019.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Khosravi M, Poursaleh A, Ghasempour G,
Farhad S and Najafi M: The effects of oxidative stress on the
development of atherosclerosis. Biol Chem. 400:711–732.
2019.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Lankin VZ, Sharapov MG, Goncharov RG,
Tikhaze AK and Novoselov VI: Natural dicarbonyls inhibit peroxidase
activity of peroxiredoxins. Dokl Biochem Biophys. 485:132–134.
2019.PubMed/NCBI View Article : Google Scholar
|
|
6
|
de Lucia C, Komici K, Borghetti G,
Femminella GD, Bencivenga L, Cannavo A, Corbi G, Ferrara N, Houser
SR, Koch WJ and Rengo G: microRNA in cardiovascular aging and
age-related cardiovascular diseases. Front Med (Lausanne).
4(74)2017.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Lin D, Xiu-Qing H, Tao S, Lin-Fang LI,
Jian LI and Amp BH: miR-301a regulates the expression of
inflammatory factors in macrophages. Chin J Arterioscler.
25:447–451. 2017.
|
|
8
|
Dong Y, Yan X, Yang X, Yu C, Deng Y, Song
X and Zhang L: Notoginsenoside R1 suppresses miR-301a via NF-κB
pathway in lipopolysaccharide-treated ATDC5 cells. Exp Mol Pathol.
112(104355)2020.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Hazra B, Chakraborty S, Bhaskar M,
Mukherjee S, Mahadevan A and Basu A: miR-301a regulates
inflammatory response to japanese encephalitis virus infection via
suppression of NKRF activity. J Immunol. 203:2222–2238.
2019.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Chen H, Qi J, Bi Q and Zhang S:
Suppression of miR-301a alleviates LPS-induced inflammatory injury
in ATDC5 chondrogenic cells by targeting Sirt1. Int J Clin Exp
Pathol. 10:8991–9000. 2017.PubMed/NCBI
|
|
11
|
Guan F, Kang Z, Zhang JT, Xue NN, Yin H,
Wang L, Mao BB, Peng WC, Zhang BL, Liang X, et al: KLF7 promotes
polyamine biosynthesis and glioma development through
transcriptionally activating ASL. Biochem Biophys Res Commun.
514:51–57. 2019.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) Method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Wysocka A, Cybulski M, Berbeć H,
Wysokiński A, Stążka J, Daniluk J and Zapolski T: Dynamic changes
of paraoxonase 1 activity towards paroxon and phenyl acetate during
coronary artery surgery. BMC Cardiovasc Disord.
17(92)2017.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Marenberg ME, Risch N, Berkman LF,
Floderus B and de Faire U: Genetic susceptibility to death from
coronary heart disease in a study of twins. N Engl J Med.
330:1041–1046. 1995.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Schober A and Weber C: Mechanisms of
MicroRNAs in Atherosclerosis. Annu Rev Pathol. 11:583–616.
2016.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Pordzik J, Pisarz K, De Rosa S, Jones AD,
Eyileten C, Indolfi C, Malek L and Postula M: The potential role of
platelet-related micrornas in the development of cardiovascular
events in high-risk populations, including diabetic patients: a
review. Front Endocrinol (Lausanne). 9(74)2018.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Canfrán-Duque A, Rotllan N, Zhang X,
Fernández-Fuertes M, Ramírez-Hidalgo C, Araldi E, Daimiel L, Busto
R, Fernández-Hernando C and Suárez Y: Macrophage deficiency of
miR-21 promotes apoptosis, plaque necrosis, and vascular
inflammation during atherogenesis. EMBO Mol Med. 9:1244–1262.
2017.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Feng J, Li A, Deng J, Yang Y, Dang L, Ye
Y, Li Y and Zhang W: miR-21 attenuates lipopolysaccharide-induced
lipid accumulation and inflammatory response: Potential role in
cerebrovascular disease. Lipids Health Dis. 13(27)2014.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Liu L, Yin H, Hao X, Song H, Chai J, Duan
H, Chang Y, Yang L, Wu Y, Han S, et al: Down-Regulation of
miR-301a-3p Reduces Burn-Induced Vascular Endothelial Apoptosis by
potentiating hMSC-Secreted IGF-1 and PI3K/Akt/FOXO3a Pathway.
iScience. 23(101383)2020.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Li H, Xue M, Xu J and Qin X: MiR-301a is
involved in adipocyte dysfunction during obesity-related
inflammation via suppression of PPARγ. Pharmazie. 71:84–88.
2016.PubMed/NCBI
|
|
21
|
He C, Shi Y, Wu R, Sun M, Fang L, Wu W,
Liu C, Tang M, Li Z, Wang P, et al: miR-301a promotes intestinal
mucosal inflammation through induction of IL-17A and TNF-α in IBD.
Gut. 65:1938–1950. 2016.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Domínguez-Andrés J and Netea MG: Long-term
reprogramming of the innate immune system. J Leukoc Biol.
105:329–338. 2019.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Li WY, Zhang WT, Cheng YX, Liu YC, Zhai
FG, Sun P, Li HT, Deng LX, Zhu XF and Wang Y: Inhibition of
KLF7-Targeting MicroRNA 146b Promotes Sciatic Nerve Regeneration.
Neurosci Bull. 34:419–437. 2018.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Blackmore MG, Wang Z, Lerch JK, Motti D,
Zhang YP, Shields CB, Lee JK, Goldberg JL, Lemmon VP and Bixby JL:
Krüppel-like Factor 7 engineered for transcriptional activation
promotes axon regeneration in the adult corticospinal tract. Proc
Natl Acad Sci USA. 109:7517–7522. 2012.PubMed/NCBI View Article : Google Scholar
|