Introduction
Coronary artery bypass grafting (CABG) remains the
primary treatment for patients with unprotected left main and
multi-vessel coronary artery disease (1). Although total arterial
revascularization has been advocated, the autologous saphenous vein
is still the most commonly used graft owing to its availability and
length (2). However, the per-graft
occlusion rate of the vein graft is estimated to be up to 25%
during the first 12 months following CABG, and almost half of the
venous conduits fail at 10 years (3).
Clinically, vein graft stenotic disease leads to
considerable post-operative adverse events, including recurrent
angina, myocardial infraction and repeat revascularization
(4). To date, the
pathophysiological mechanisms underlying venous wall remodeling
remain to be fully understood. Thrombosis, neointimal hyperplasia
and superimposed atherosclerosis are generally regarded as primary
causes of vein graft failure that have attracted the most research
interest for therapeutic applications (5). To the best of our knowledge, however,
apart from statins and antiplatelet agents, no other interventions
have proven to be clinically effective (6). Considering the aging population and
the persistent increase in the incidence of patients with CABG
living with symptomatic vein graft stenotic disease in Europe and
USA (7-9),
further identification of novel biomarkers that can be effectively
targeted is urgently required (10).
There has been an increasing interest in the role of
non-coding RNAs in the pathogenesis of vascular disease (11). Based on transcript size, non-coding
RNAs are classified into small, medium and long non-coding RNAs
(lncRNAs) (12). The biogenesis and
functional properties of small RNAs, particularly microRNAs (miRNAs
or miRs), in the onset and progression of cardiovascular disease
have been well-described (13). For
example, the administration of miR-221 has been shown to
effectively inhibit endothelial cell proliferation at predilection
sites (14). However, the
expression profiles and potential functions of lncRNAs in the
progression of venous wall remodeling following bypass grafting
remain unknown. The present study aimed to investigate the dynamic
expression profiles of lncRNAs following bypass grafting in a rat
model, and to elucidate the potential association between lncRNAs
and vein graft stenotic disease.
Materials and methods
Rat model establishment
A total of 12 male Sprague Dawley rats (age, 6-8
weeks) were purchased from the Laboratory Animal Center of Sichuan
University (Chengdu, Sichuan, China). They were randomly divided
into four groups to ensure equal weight distribution (weight,
248±27 g). One group receiving sham surgery was set as the control
(day 0; n=3), whilst the other three groups were set as the
experimental groups according to different time points after vein
bypass grafting (7, 14 and 28 days after surgery, respectively; n=3
per group). For the experimental groups, the rats were anesthetized
with intraperitoneal injection of 10% chloral hydrate (350 mg/kg).
Heparin (100 U/kg) and cefazolin (50 mg/kg) were administered
intraperitoneally for anticoagulation and infection prevention,
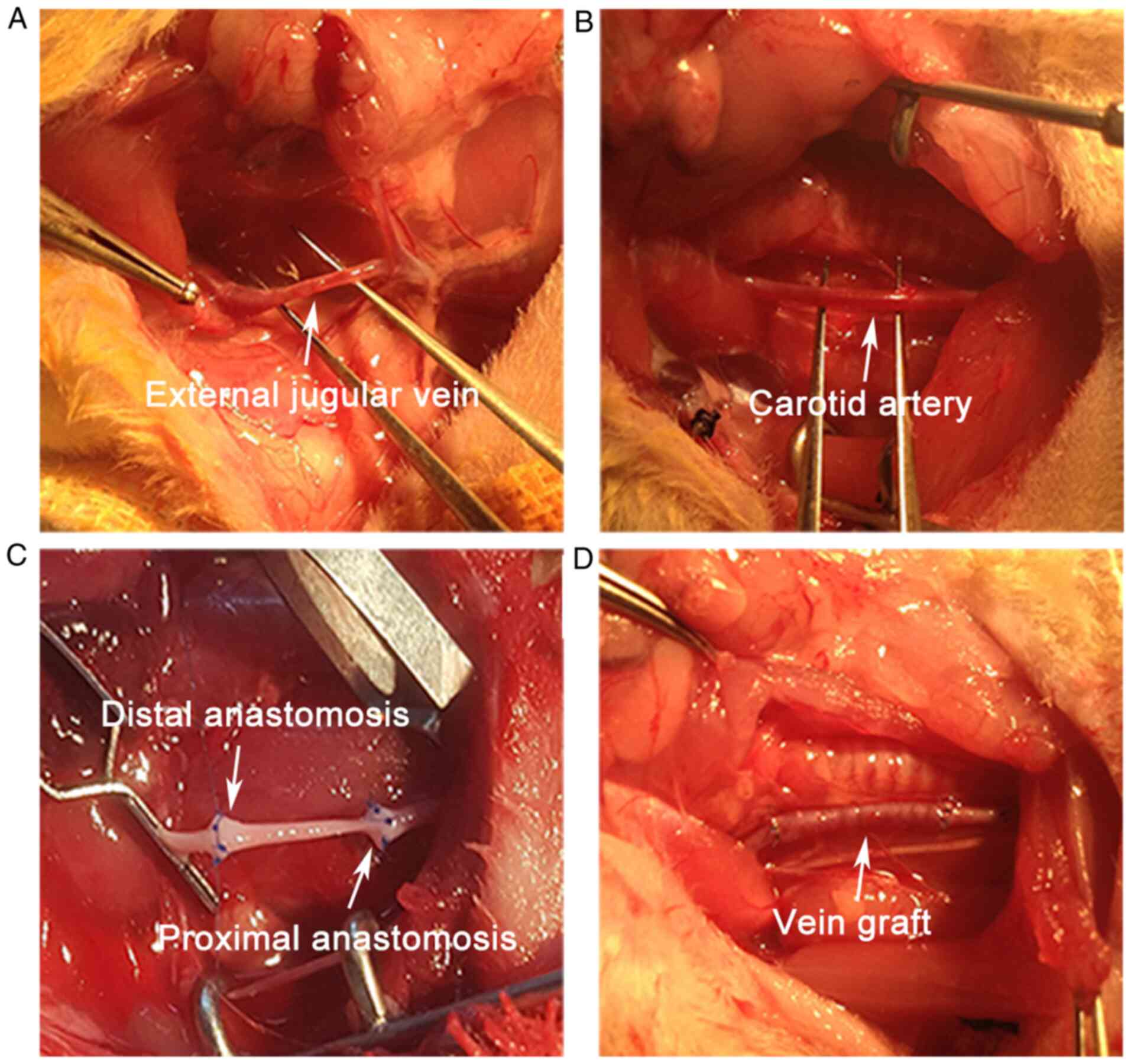
respectively. No signs of peritonitis were observed. A length of
vein graft (1 cm) was harvested from the external jugular vein and
preserved in heparinized saline (25 U/ml). The vein was
subsequently grafted to the carotid artery (Fig. 1). For the control group, all the
procedures performed were identical with those in the experimental
groups except vein bypass grafting was not peroformed. These rats
were housed in a temperature-controlled room at 20±2˚C and relative
humidity of 50%, with a 12-h light/dark cycle. Free access to food
and water was provided. These experiments were performed according
to the National Institutes of Health Guideline for the Care and Use
of Laboratory Animals (15). The
present study was approved by the Animal Ethical and Welfare of
Sichuan University, Sichuan, China (approval no. 2018103A).
Total RNA extraction and array
hybridization
Following the intraperitoneal administration of 10%
chloral hydrate (350 mg/kg) for anesthetization, the rats were
sacrificed by cervical dislocation. Vein samples (~1 cm) were
harvested from the different groups. Total RNA was isolated using
TRIzol® reagent according to the manufacturer's
instructions (Invitrogen; Thermo Fisher Scientific, Inc.). The
purity of the RNA samples was analyzed using the Nanodrop
spectrophotometer (Implen GmbH), which measured absorbance at 230,
260 and 280 nm. The 260/280 and 260/230 absorbance ratios were
calculated to evaluate RNA purity (A260/A280
ratios are 1.8-2.2 and A260/A230 ratios are
over 1.7). 1.0% non-denaturing agarose gel electrophoresis was used
to determine the RNA integrity. The microarray analysis was
performed by Kangchen BioTech Co., Ltd. using Agilent Technologies
Rat LncRNA Microarray scanner (cat no. G2505C; Agilent
Technologies, Inc.). Sample labeling and microarray hybridization
were performed according to the manufacturer's protocols. In brief,
ribosomal RNA was removed from total RNA using a
mRNA-ONLY™ Eukaryotic mRNA Isolation kit (cat. no.
MOE51010; Zhongbei Linge Biotechnology, Ltd.; http://www.bjzblg.com/product/18395.html). The
remaining RNA was amplified and reverse transcribed into
fluorescent complementary (c)RNA using the PrimeScript™ RT Reagent
Kit (cat. no. RR037B; Takara bio, Inc.) at 37˚C for 15 min,
followed by 85˚C for 5 sec. The RNeasy Mini kit and NanoDrop
ND-1000 instrument were used to purify and measure the
concentration of the labeled cRNAs. A total of 5 µl 10X Blocking
Agent and 1 µl 25X Fragmentation Buffer were mixed with 1 µg
labeled cRNA and the mixture was then heated at 60˚C for 30 min.
Subsequently, 25 µl 2X GE Hybridization buffer was added to dilute
the labeled cRNA. The hybridization solution was incubated for 17 h
at 65˚C in an Agilent Technologies Hybridization oven. After
washing, the hybridized arrays were scanned with using an Agilent
microarray scanner (cat no. G2505C; Agilent Technologies,
Inc.).
Bioinformatics analysis of microarray
data
Raw data were extracted from the acquired array
images using Agilent Feature Extraction software 11.0.1.1 (Agilent
Technologies, Inc.). GeneSpring GX v12.1 software (Agilent
Technologies, Inc.) was used to normalize quantiles of raw data.
Genes with an adjusted P-value <0.01 and an absolute value of
log2 (fold change) >2.0 were considered to be
significantly differentially expressed. Hierarchical clustering
analysis was performed to observe the different clustering patterns
of lncRNAs and mRNAs between the control and experimental groups.
Scatter and volcano plots analysis were constructed to express the
differentially expressed genes. In order to examine the
distribution of differentially expressed lncRNAs, histograms and a
pie chart were generated to reflect the chromosomal distribution,
lengths and regulation mechanisms of the differentially expressed
lncRNAs.
Prediction and functional enrichment
analysis of lncRNA target genes
The protein-coding genes targeted by lncRNAs were
predicted via ce- and trans-regulation analyses,
respectively. In the ce-regulation analysis, lncRNAs compete
with shared miRNAs to regulate target protein-coding genes, which
are genomically neighboring within 10 kb. In the
trans-regulation analysis, lncRNAs regulate protein-coding
genes sequences matched with low complement energy (inclusion
criteria, G<-20). Kyoto Encyclopedia of Genes and Genomes (KEGG)
and Gene Ontology (GO) enrichment analyses were performed to
evaluate the functional pathways and biological relevance of the
lncRNA co-expressed mRNAs using Database for Annotation,
Visualization and Integrated Discovery (david.abcc.ncifcrf.gov/). All functional analyses were
performed using Enrichr (16).
Co-expression network analysis
The co-expression of lncRNAs and protein-coding
genes was assessed using Gephi 0.9.2(17) and Pearson's correlation test
(P<0.05). Upregulated lncRNAs were presented as orange nodes and
downregulated lncRNAs were presented as blue nodes. Protein-coding
genes targeted by lncRNAs were presented as green nodes.
Verification of microarray data by
reverse transcription-quantitative (RT-q)PCR
RT-qPCR was performed to validate the microarray
data. Samples were divided into control group and experimental
groups. Samples of control group were collected from rats receiving
sham surgery (n=3), samples of experimental groups were collected
from rats that received vein bypass grafting (7, 14 and 28 day
after surgery, respectively; n=3 per group). Total RNA was
extracted with TRIzol® reagent from the remaining
portion of tissues not used for the lncRNA microarray.
Subsequently, first-strand complementary DNAs were generated using
SuperScript™ III Reverse Transcriptase (Invitrogen; Thermo Fisher
Scientific, Inc.) following the manufacturer's protocols. The
RT-qPCR process was performed using a ViiA-7 RT-PCR System (Applied
Biosystems; Thermo Fisher Scientific, Inc.) and Arraystar
SYBR-Green Real-time qPCR Master Mix (cat no. AS-MR-005-25;
Arraystar, Inc.). The reactions were performed under the following
conditions: Initial denaturation at 95˚C for 30 sec, followed by 40
cycles of 95˚C for 5 sec and 60˚C for 30 sec. The relative
expression levels of lncRNAs were quantified using the
2-ΔΔCq method (14).
β-actin served as an internal control. The primers for RT-qPCR were
designed according to the lncRNA sequences in NONCODE (version
v6.0; http://www.noncode.org) and primer
sequences were synthesized and purified by Kangchen BioTech Co.,
Ltd. The primer sequences are presented in Table I. All experiments were performed in
triplicate for error reduction.
 | Table IPCR primers used for expression
analysis. |
Table I
PCR primers used for expression
analysis.
| Gene | Primer sequence
(5'→3') | Length, bp |
|---|
| AF062402 | F:
TGGCCCAGAACGGAAATATCA | 116 |
| | R:
ACTAGCCCGGAGTTTGACCAT | |
| BC091437 | F:
GGCACCAGAGCTACCATTCC | 167 |
| | R:
CGTGGACTTGGCAGGAGAAA | |
| BC166461 | F:
GCACAAGGTTCAAAAAGGCAA | 97 |
| | R:
GCTTGGTAGCCATACTTCTGGAA | |
| Src | F:
GAACCCGAGAGGGACCTTC | 61 |
| | R:
GAGGCAGTAGGCACCTTTTGT | |
| Edg1 | F:
ATGGTGTCCACTAGCATCCC | 112 |
| | R:
CGATGTTCAACTTGCCTGTGTAG | |
| Mcam | F:
CCCAAACTGGTGTGCGTCTT | 220 |
| | R:
GGAAAATCAGTATCTGCCTCTCC | |
| β-actin | F:
CGAGTACAACCTTCTTGCAGC | 202 |
| | R:
ACCCATACCCACCATCACAC | |
Cells and cell culture
After the rats which were used for lncRNA microarray
were sacrificed by cervical dislocation, primary vascular smooth
muscle cells (VSMCs) were obtained from normal external jugular
vein. All procedures were performed according to a previously
published protocol (18). In brief,
external jugular vein was isolated from adherent tissue, and
adventitia and endothelium were denuded. Pieces of vein (1
mm2) were cultured in serum-free DMEM/F12 (Gibco; Thermo
Fisher Scientific, Inc.) supplemented with 20% FBS (Gibco; Thermo
Fisher Scientific, Inc.) and 1% penicillin-streptomycin at 37˚C in
a 5% CO2 humidified incubator. Cells migrated from the
sections of aorta after 7-9 days. Vein pieces were then removed and
VSMCs were left to proliferate. Experiments used VSMCs from
passages 3-6 at 80-90% confluence.
Infection with lentiviral vector and
cell proliferation assay
VSMCs suspensions were seeded into six-well plates
at a density of 4x105 cells/well. DMEM/F12 (Gibco;
Thermo Fisher Scientific, Inc.) with 10% v/v FBS (Gibco; Thermo
Fisher Scientific, Inc.) was removed the following day and replaced
with virus concentrated liquid (Shanghai OBiO Technology Co.,
Ltd.). Stable overexpression of Src was achieved by construction of
the GL107 pSLenti-EF1-EGFP-P2A-Puro-CMV-MCS-3xFLAG-WPRE lentivirus
(Shanghai OBiO Technology Co., Ltd.), containing the AF062402, one
of the significantly upregulated lncRNAs, gene vector with green
fluorescence protein (GFP; Shanghai OBiO Technology Co., Ltd.) to
detect transfection efficacy. Polybrene reagent (Shanghai OBiO
Technology Co., Ltd.) was diluted to a final concentration of 5
µg/ml. Multiplicity of infection (MOI) values (1, 2, 5, 10, 30 and
50) were then tested on VSMCs. A total of 4x106
lentivral particles were used for transfection and the optimal MOI
value was 10 in the present study. At this value, the efficiency of
infection reached 80% 3 days post-infection. After swirling the
plate gently to mix the cells, the plate was placed in an incubator
with 5% CO2 at 37˚C for 24 h. After 24 h, the virus
concentrated medium was removed and replaced by DMEM/F12 medium
(Gibco; Thermo Fisher Scientific, Inc.) with 10% FBS (Gibco; Thermo
Fisher Scientific, Inc.). At 3 days post-transfection, GFP
expression was observed as the lentivirus was integrated into the
VSMC genome, in five randomly-selected fields using a fluorescence
microscope (magnification, x200). Cells with a transfection
efficiency >80% on day 3 were selected for subsequent analysis.
VSMCs transfected with AF062402 lentiviral expression vector were
set as the AF062402 intervention group. Non-transfected cells were
set as the blank control and lentivirus without the AF062402 gene
was transfected as the negative control (NC). At 72 h following
transfection, RT-qPCR was performed to verify the expression levels
of AF062402 in the different groups. Moreover, Cell Counting Kit-8
(CCK-8) assay was performed using a kit from Abcam. VSMCs were
digested into single cell suspensions and seeded into 96-well
plates (5x103 cells/ml). After 48 h, CCK-8 solution (10
µl/well) was added and the plates were incubated for 2.5 h at 37˚C
before the optical density (OD) value was detected at 460 nm using
a plate reader. All experiments were performed in triplicate.
Statistical analysis
Feature Extraction software (Agilent Technologies,
version 11.0.1.1) was used to analyze the acquired array images.
Differentially expressed lncRNAs and mRNAs were identified by
adjusted P-value <0.01 and absolute value of the log2
(fold change) >2.0. The results of CCK-8 assay and RT-qPCR are
expressed as the mean ± SD (n=3). Statistical comparisons between
groups were performed using one-way analysis of variance with
Tukey's post hoc multiple comparison test. P<0.05 was considered
to indicate a statistically significant difference.
Results
Overview of microarray data and
expression patterns of lncRNAs
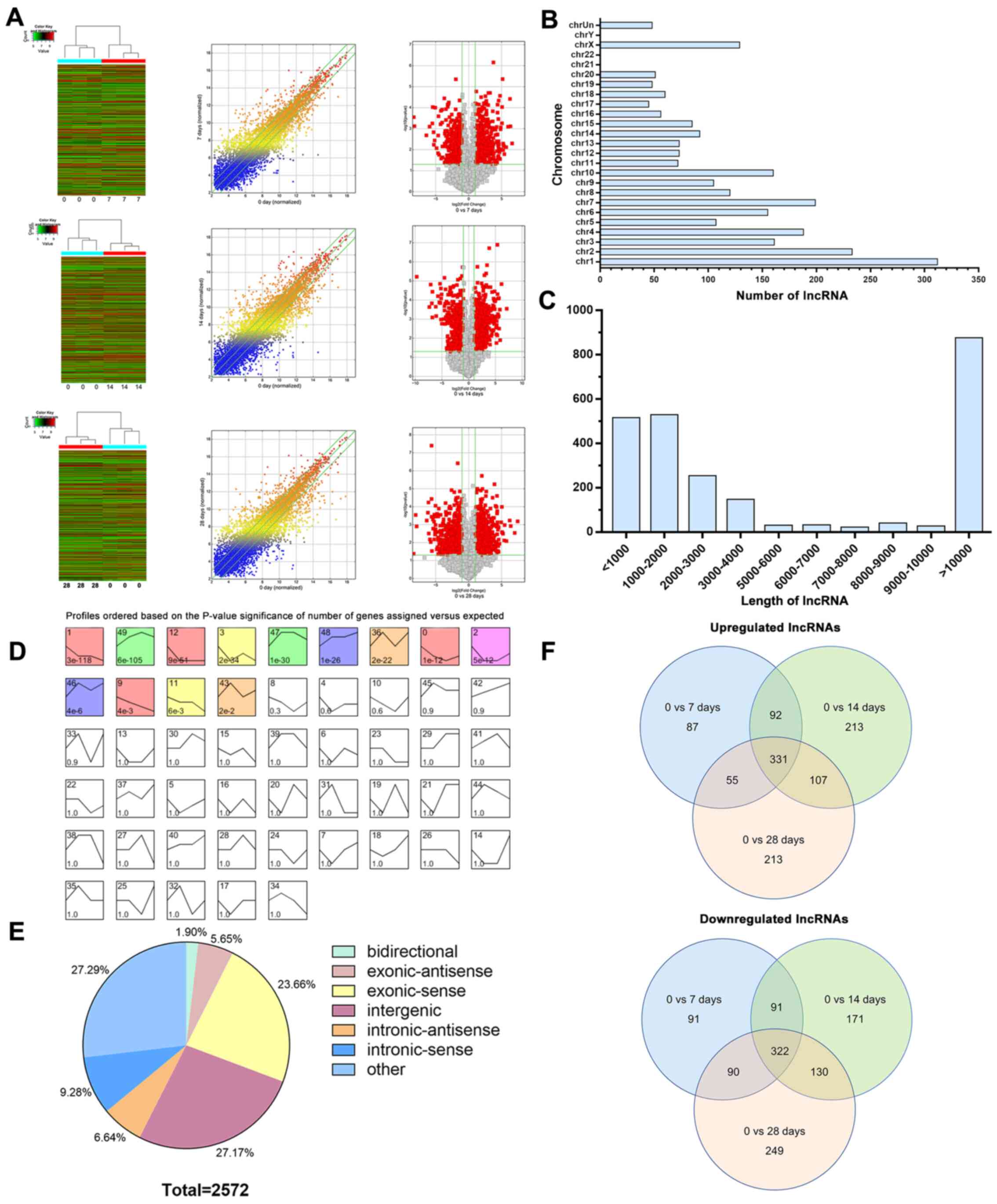
A total of 8,041 lncRNAs and 13,001 mRNAs were
detected. Box plots and hierarchical clustering analysis (Fig. S1) provided an overview of all
normalized genes in all samples. As shown in the hierarchical
clustering analyses, scatter and volcano plots, distinct expression
profiles of lncRNAs and mRNAs at different post-operative time
points were observed (Figs. 2A and
S2). The lncRNAs were then
classified based on chromosomal distribution and length (Fig. 2B and C). Subsequently, 13 lncRNA profiles with
statistical significance were selected for further investigation
(Fig. 2D). A total of 2,572 lncRNAs
and 1,652 associated mRNAs with significant changes in expression
levels (P<0.01; fold change >2) were detected. lncRNAs were
categorized into five subgroups (Fig.
2E) according to their association with adjacent genes as
follows: i) Intergenic lncRNAs, 27.17%; ii) exonic-antisense and
exonic-sense lncRNAs, 5.65 and 23.66%, respectively; iii)
intronic-antisense and intronic-sense lncRNAs, 6.64 and 9.28%,
respectively; iv) lncRNAs transcribed on the opposite strand of a
coding gene, 1.90% and v) the remaining lncRNAs transcribed in
other patterns, 27.29%. The numbers of lncRNAs with significantly
upregulated or downregulated expression at different post-operative
time points are presented in Fig.
2F. Moreover, the top 10 lncRNAs with significant changes in
expression levels at different post-operative time points are
presented in Table II.
| Table IITop 10 significantly differential
expressed lncRNAs at days 7, 14 and 28 vs. day 0. |
Table II
Top 10 significantly differential
expressed lncRNAs at days 7, 14 and 28 vs. day 0.
| A, Day 7 |
|---|
| Upregulated | Downregulated |
|---|
| lncRNA | P-value | FC | lncRNA | P-value | FC |
|---|
| MRAK132609 | 0.0050 | 95.50 | XR_006421 | 0.0003 | 320.62 |
| EF199812 | 0.0008 | 79.79 | BC086433 | 0.0008 | 298.86 |
| MRAK142484 | 0.0000 | 76.34 | MRuc008egb | 0.0021 | 85.27 |
| BC158825 | 0.0008 | 62.77 | XR_008627 | 0.0008 | 60.58 |
| AJ005396 | 0.0076 | 51.12 | BC169049 | 0.0006 | 45.32 |
| MRAK005319 | 0.0069 | 40.62 | AB049626 | 0.0001 | 43.74 |
| XR_007646 | 0.0061 | 38.35 | MRuc008lzo | 0.0015 | 39.94 |
| X89963 | 0.0051 | 37.14 | MRAK046251 | 0.0058 | 38.96 |
| MRuc007pas | 0.0000 | 35.18 | MRAK005337 | 0.0036 | 35.81 |
| BC167061 | 0.0001 | 33.34 | BC166501 | 0.0001 | 33.57 |
| B, Day 14 |
| Upregulated | Downregulated |
| lncRNA | P-value | FC | lncRNA | P-value | FC |
| EF199812 | 0.0003 | 138.51 | BC086433 | 0.0000 | 848.08 |
| MRAK132609 | 0.0000 | 70.79 | XR_008627 | 0.0002 | 164.20 |
| MRAK142484 | 0.0003 | 56.92 | AJ517196 | 0.0000 | 138.75 |
| XR_007646 | 0.0031 | 51.58 | MRAK005337 | 0.0001 | 118.56 |
| BC167061 | 0.0001 | 50.03 | XR_006421 | 0.0008 | 112.10 |
| MRAK143033 | 0.0007 | 49.40 | MRuc008egb | 0.0000 | 110.95 |
| BC158675 | 0.0001 | 44.93 | MRuc008lzo | 0.0002 | 71.61 |
| XR_008202 | 0.0005 | 44.29 | BC169049 | 0.0007 | 67.66 |
| EF673689 | 0.0008 | 42.76 | AB049626 | 0.0001 | 54.08 |
| BC158638 | 0.0000 | 42.24 | MRuc007smc | 0.0001 | 45.26 |
| C, Day 28 |
| Upregulated | Downregulated |
| lncRNA | P-value | FC | lncRNA | P-value | FC |
| MRAK132609 | 0.0007 | 170.96 | XR_006421 | 0.0003 | 262.57 |
| MRAK020463 | 0.0029 | 125.46 | XR_006359 | 0.0012 | 237.66 |
| AJ005396 | 0.0012 | 82.50 | MRAK149123 | 0.0006 | 82.12 |
| X89963 | 0.0006 | 74.27 | MRAK084152 | 0.0002 | 72.11 |
| NR_027324 | 0.0082 | 52.56 | BC086433 | 0.0001 | 68.54 |
| EF199812 | 0.0011 | 43.63 | MRAK148606 | 0.0031 | 65.84 |
| AF130879 | 0.0075 | 41.29 | MRAK037975 | 0.0067 | 58.59 |
| MRAK164133 | 0.0032 | 36.60 | XR_007815 | 0.0094 | 53.91 |
| L38717 | 0.0027 | 33.82 | MRAK080917 | 0.0000 | 49.53 |
| U57362 | 0.0026 | 32.63 | MRAK080330 | 0.0000 | 49.43 |
lncRNA target prediction and
functional annotation
In order to identify the potential functional
pathways of lncRNAs with significant changes in expression levels,
ce- and trans-regulation analysis results were
combined and overlapping mRNAs were selected for further ‘GO’ and
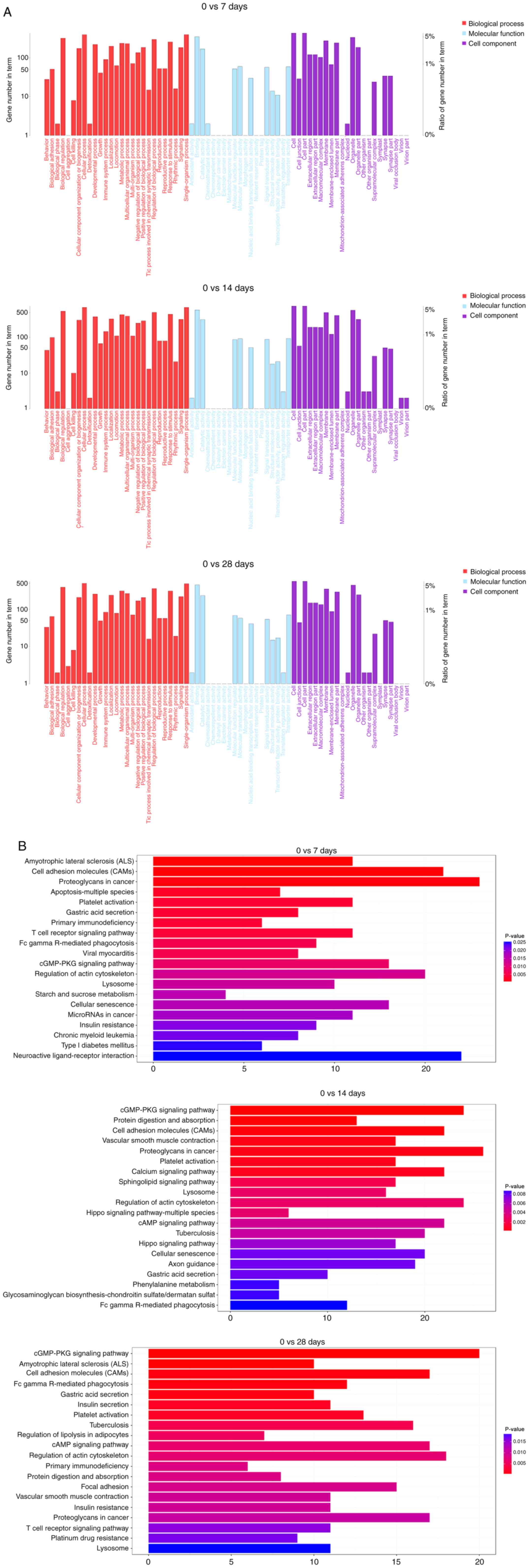
‘KEGG’ analyses. ‘GO’ analysis revealed a significant enrichment of
4,993 ‘GO’ terms (P<0.01), of which ‘cellular process’, ‘protein
binding’ and ‘cell/cell part’ were the top-listed terms involved in
biological process, molecular function and cell component,
respectively (Fig. 3A). ‘KEGG’
analysis revealed that 298 biological pathways were enriched
(P<0.01), of which several were associated with vascular
remodeling, including ‘cell adhesion molecules’, ‘platelet
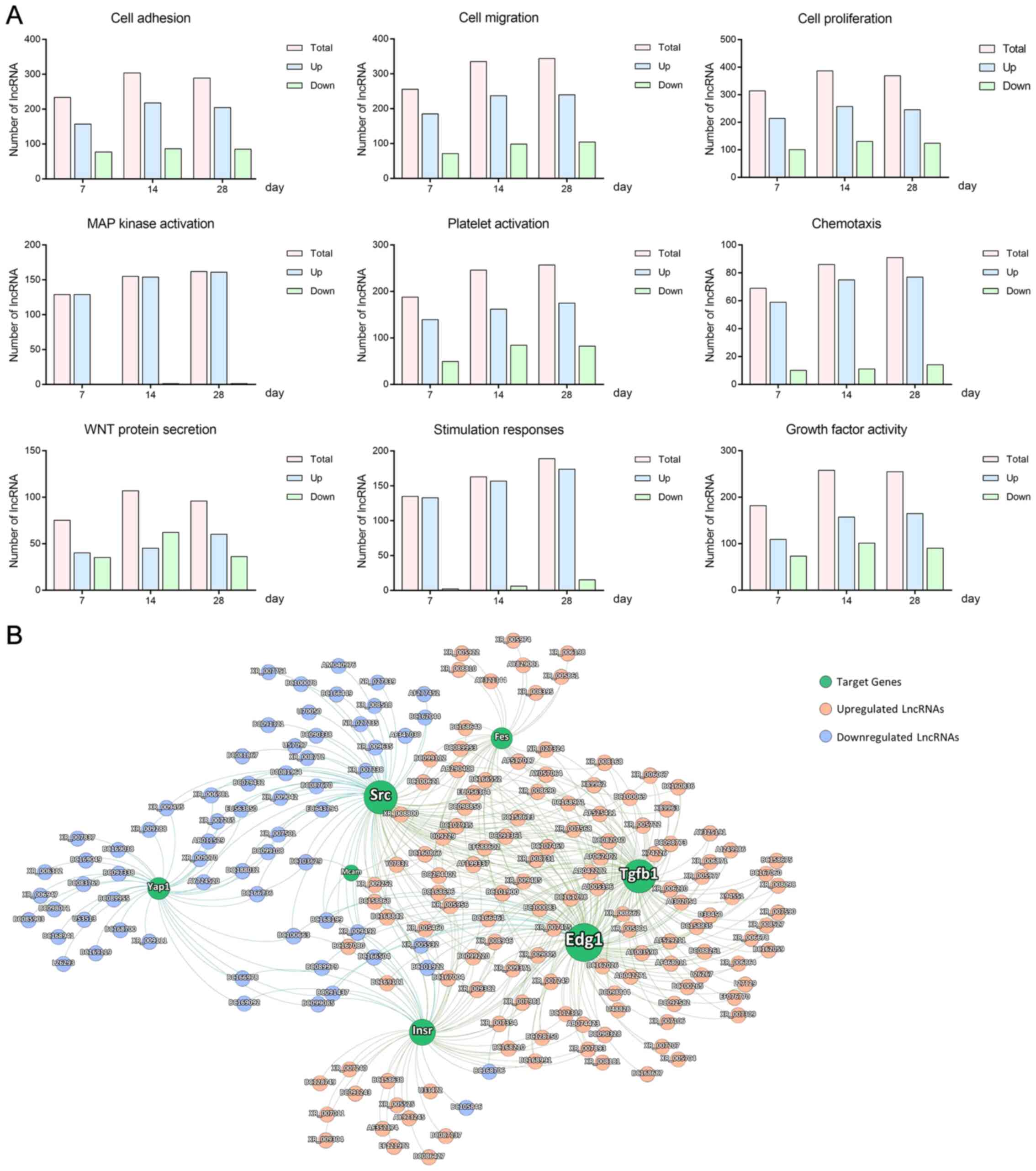
activation’ and ‘vascular smooth muscle contraction’ (Fig. 3B). Among these, nine biological
processes associated with vascular remodeling process were selected
for further analysis, including ‘cell adhesion’, ‘cell migration’,
‘cell proliferation’, ‘MAP kinase activation’, ‘platelet
activation’, ‘chemotaxis’, ‘WNT protein secretion’, ‘stimulation
responses’ and ‘growth factor activity’. The changes in the numbers
of lncRNAs in these selected biological processes are presented in
Fig. 4A.
Co-expression network analysis of
lncRNAs and associated protein-coding genes
Based on the results of functional analysis, a total
of 360 lncRNAs and 135 associated protein-coding genes were
predicted to be involved in the vascular remodeling process (Table
SI). Among these, 194 lncRNAs and seven previously reported
associated protein-coding genes (19-32)
involved in mediating vascular disease were selected to construct a
co-expression network. Each protein-coding gene was targeted by
multiple lncRNAs (Fig. 4B).
RT-qPCR validation and stimulation of
VSMC proliferation via overexpression of AF062402
A total of three lncRNAs and associated
protein-coding genes were randomly selected to verify the
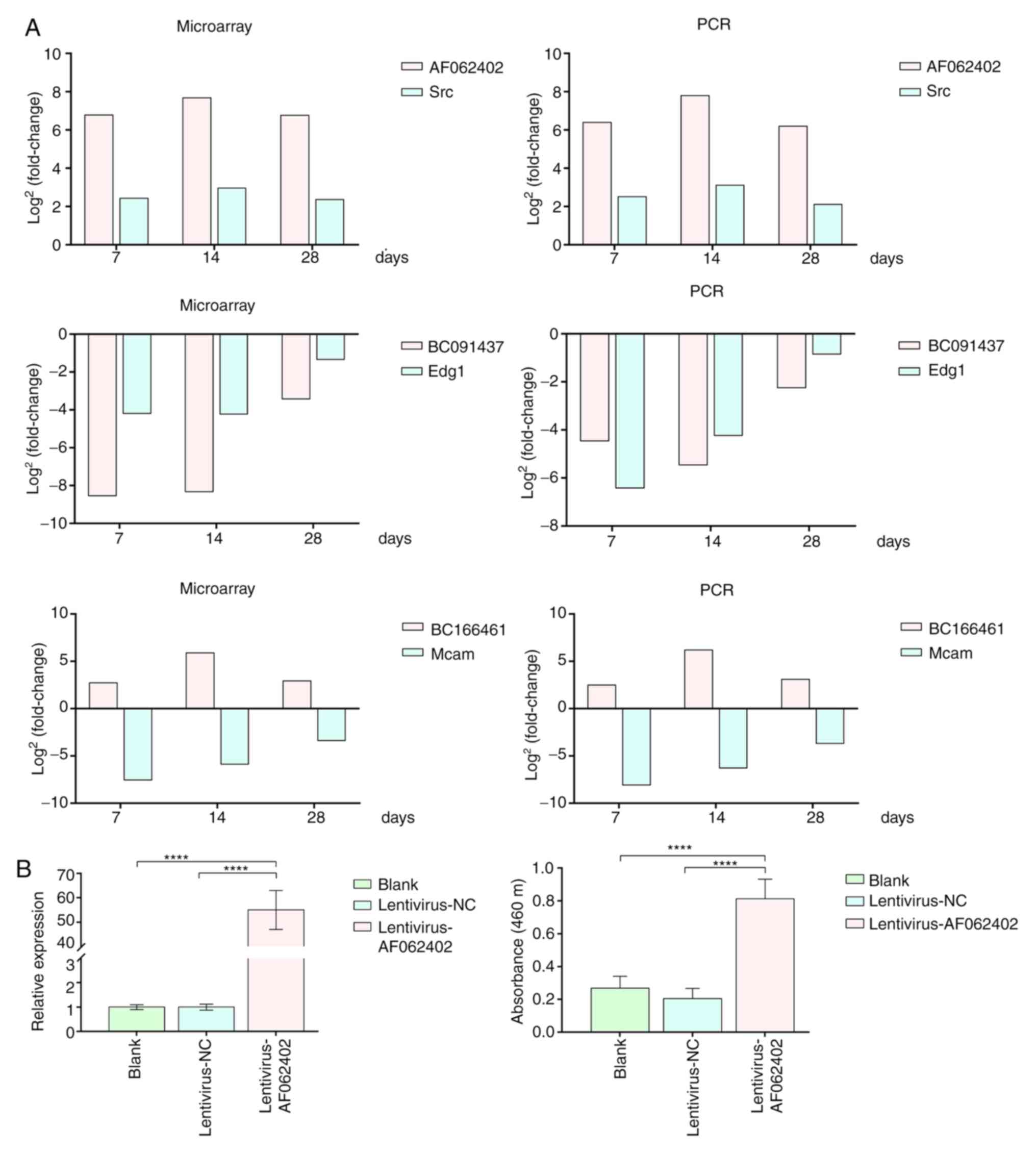
microarray data by RT-qPCR. AF062402 and Src were upregulated,
whereas BC091437 and Edg1 were downregulated (Fig. 5A). Moreover, the upregulation of
BC166461 was accompanied by downregulation of melanoma cell
adhesion molecule (Mcam). The relative expression levels of the
selected lncRNAs and associated protein-coding genes were
consistent with the microarray data. Moreover, RT-qPCR confirmed
that the relative expression of AF062402 in the Lentivirus-AF062402
group was significantly higher than that in the blank and
Lentivirus-NC groups. The overexpression of AF062402 stimulated the
proliferation of primary VSMCs compared with the other two groups
(Fig. 5B).
Discussion
Vein graft restenosis is a primary long-term
complication which leads to a poor prognosis of patients with CABG
(10). However, apart from statins
and antiplatelet agents, few therapeutic strategies have been
proven to have significant clinical efficacy (6). Multiple studies have demonstrated that
lncRNAs serve key roles in the pathophysiology of various diseases
by participating in numerous regulatory processes, such as DNA
methylation, histone modification, basal transcription,
post-transcriptional processes and protein binding (33,34),
indicating that lncRNAs may be potential intervention targets for
vein graft restenosis.
The present study systemically screened a
genome-wide microarray profile of lncRNA and mRNA expression at
different time points following the establishment of rat autogenous
vein bypass grafts. A total of 8,041 lncRNAs and 13,001 mRNAs were
detected. In total, 2,572 dysregulated lncRNAs were screened
following vein bypass grafting and lncRNA-associated protein coding
genes were predicted by ce- and trans-regulation
analysis. Through enrichment analysis, nine biological processes
associated with the vascular remodeling process were selected.
Moreover, co-expression network analysis revealed an association
between lncRNAs and their target genes, which were reported to play
roles in mediating vascular disorders (19-32).
Furthermore, it was found that AF062402, one of the significantly
upregulated lncRNAs, may serve a role in inducing vein graft
restenosis.
Various pathophysiological changes secondary to
vascular bypass grafting, such as inflammation, platelet
activation, stimulation responses, leukocyte chemotaxis and
mechanical intimal injury, are regarded as primary causes of vein
graft restenosis (35-37).
The inflammatory and thrombotic cascade responsible for early graft
thrombosis and failure can be caused by direct endothelial injury
or endothelial activation (38).
Intimal hyperplasia and atherosclerosis are also considered to be
primary causes of lumen remodeling of the graft, which are
associated with the processes of leukocyte adhesion and vascular
endothelial cells and VSMC proliferation (35). Several studies have implicated the
production of reactive oxygen species in disease progression in
grafts (39,40). There is increasing evidence to
demonstrate a pivotal role of MAPK activation cascades in vascular
restenosis, which contribute to proliferation of VSMCs and
neointima formation via downstream proteins (41). It is also known that leukocyte
chemotaxis serves a key role in a complex mechanism, involving an
association between inflammation factors, platelets and vascular
disease (36). Moreover, exposure
to high pressure of arterial circulation results in early, diffuse
intimal thickening as a compensatory response termed
arterialization (37). With
enhanced understanding of the functions of lncRNAs in regulating
transcription, lncRNA-mRNA pairs were predicted to be involved in
vein graft failure following CABG in the present study. The
association between lncRNA and mRNA was the focus of the present
study.
As shown in the present study, four protein-coding
genes were targeted by both upregulated and downregulated lncRNAs.
Edg1, a specific cell surface G-protein coupled receptor,
stimulates the activation of multiple cellular events, such as cell
proliferation and migration, and the expression of adhesion
molecules via S1P signaling (42).
c-Src, a member of the Src family, is involved in multiple
signaling mechanisms; it not only serves a critical role in
mediating the migration and proliferation of VSMCs via the
c-Src/protein kinase C (PKC)/MAPK signaling pathway, but is also
essential in inducing the expression of Edg1, which is hypothesized
to contribute to the progression of intimal thickening following
vascular injury in VSMCs (20,21).
Overexpression of AF062402 in the present study stimulated the
proliferation of VSMCs, verifying the potential therapeutic effect
of AF062402-Src in mediating VSMC proliferation. For patients with
CABG with diabetes mellitus or hyperlipemia, insulin receptor
(Insr) may be a therapeutic target. According to a previous study,
overexpression of Insr-A, one of the isoforms of Insr, confers a
proliferative advantage on VSMCs in the early stages of the
atherosclerotic process (43).
Furthermore, decreased Insr-A/Insr-B ratio may also contribute to
apoptosis of VSMCs and increased risk of atherosclerotic plaque
rupture (22). Mcam, an endothelial
transmembrane protein of the immunoglobulin superfamily, has been
reported to be involved in several important signaling pathways in
vascular disease, including PI3K/AKT, p38 and PKC (23). The interaction between Mcam and
VEGFR-2 on the endothelial cell surface stimulates cell migration
via the AKT and p38 pathways (24).
Mcam can also interact with galectin-1 to protect against
galectin-1-induced endothelial cell apoptosis (25). Moreover, a previous study revealed a
causative role of Mcam in activating lipid-loaded macrophages in
plaques, indicating the potential ability of Mcam to be a novel
therapeutic target in vascular remodeling following CABG (26).
TGFβ-1 and Fes are two protein-coding genes shown to
be targeted by upregulated lncRNAs in the present study. TGFβ-1 is
the central regulator of the canonical SMAD2/3 pathway in mediating
the deposition of extracellular matrix, which leads to fibrosis in
vascular injury (27). A previous
study also indicated that TGFβ-1 is involved in regulating the
expression of CD40, which serves as the central regulator of
various immune responses (28).
TGFβ-1 may be responsible for heterogeneous expression of CD40 at
the site of vascular bifurcations or curvatures, particularly in
the vein grafts of patients with CABG. Fes, one of the known
members of a subfamily of the non-receptor tyrosine kinase family,
was targeted by 32 upregulated lncRNAs in the present study.
Activated Fes kinase has been reported to shift the equilibrium of
STAT3, STAT5 and extracellular-signal regulated kinase/MAPK
activation to facilitate the adhesion and chemotaxis of
inflammatory cells and regulate the proliferation of vascular
endothelial cells. Upon phenotypic transition, Fes assists in
smooth muscle cell migration to the neo-intima (29).
Yes1 associated transcriptional regulator (Yap) was
a protein-coding gene targeted by downregulated lncRNAs in the
present study. As a downstream effector, Yap is one of the vital
components of the Hippo/Yap pathway, which has been reported to
serve roles in mediating vascular restenosis in rodent models
(30). Overexpression of Yap
combined with intima-media thickness has also been observed in a
rat carotid artery balloon injury model (31). In addition, elevated activity of Yap
has been reported to mediate the switch of the VSMC phenotype to
the synthetic state, promoting neointima formation (32). Moreover, thromboxane A2-specific
agonists have been found to induce VSMC migration and proliferation
by activating Yap (19). These
results demonstrate a potential therapeutic approach for vascular
remodeling by activating the Hippo pathway or inhibiting the
expression of Yap.
Several limitations of the present study need to be
acknowledged. First, since microarray can only detect known gene
sequences, certain lncRNAs involved in the vascular remodeling
process remain to be investigated. Second, lack of knowledge of the
properties and functional mechanisms of lncRNAs involving gene
expression meant the role of a specific lncRNA in an overt
biological process according to its expression level could not be
accurately predicted. Third, although several lncRNAs with
potential association in vein graft failure were identified via
co-expression network analysis, the detailed mechanisms of these
lncRNAs remain unclear; further validation experiments, including
western blot analysis, immunohistochemistry and immunofluorescence
staining, are required to determine the casual association between
these lncRNAs and protein-coding genes overlapped with mRNAs.
In conclusion, the present study provides
preliminary evidence of lncRNA expression profiles in vein graft
restenosis following vascular grafting. Even though further
validation experiments are still required, lncRNAs and target
protein-coding genes predicted in the present study may serve as
potential therapeutic targets for vein graft restenosis following
CABG.
Supplementary Material
Microarray data quality control. (A)
Box plots showing the symmetry of data and the degree of
dispersion. The ordinate represents the fold change
(log2) in signal values of probes. (B) Heatmaps
revealing all differentially expressed lncRNAs and mRNAs. Red in
heatmap indicates increased expression; green in heatmap indicates
decreased expression. Purple, green, blue and red on the top of
heatmap indicate day 0, 7, 14 and 28, respectively. lncRNAs, long
non-coding RNAs.
Heatmaps showing mRNAs that were
differentially expressed between groups. Scatter and volcano plots
were generated to show the variations in mRNA expression between
different time points (0 vs. 7, 14 and 28 day). Red and green in
heatmap indicate increased and decreased expression, respectively;
blue on the top of heatmap indicates the control group (day 0); red
on the top of heatmap indicates the experimental group (day 7, 14
and 28, respectively); orange and blue in scatter plots indicate
significantly and non-significantly expressed mRNAs; red and grey
in volcano plots indicate significantly and non-significantly
expressed mRNAs.
Acknowledgements
Not applicable.
Funding
Funding: The present study was funded by the National Natural
Science Foundation of China (grant nos. 81670327 and 81300155), the
Commission of Science Technology of Sichuan Province (grant nos.
2019YJ0046 and 2018SZ0384) and the Clinical Research Incubation
Project of West China Hospital of Sichuan University (grant no.
2019HXFH027).
Availability of data and materials
The datasets generated and/or analyzed during the
current study are available in the NCBI Gene Expression Omnibus
repository, accession no. GSE152538.
Authors' contributions
HZ and JS confirm the authenticity of all the raw
data. HZ established the animal model. YZ performed the cell
culture experiments. JS and CL analyzed the data and wrote the
manuscript. JG and JH contributed to put forward the conception and
helped to revise the manuscript. All authors read and approved the
final manuscript.
Ethics approval and consent to
participate
The present study was approved by the Animal Ethical
and Welfare of Sichuan University, Sichuan, China (approval no.
2018103A).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Bhatt DL: CABG the clear choice for
patients with diabetes and multivessel disease. Lancet.
391:913–914. 2018.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Collet C, Capodanno D, Onuma Y, Banning A,
Stone GW, Taggart DP, Sabik J and Serruys PW: Left main coronary
artery disease: Pathophysiology, diagnosis, and treatment. Nat Rev
Cardiol. 15:321–331. 2018.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Caliskan E, de Souza DR, Böning A,
Liakopoulos OJ, Choi YH, Pepper J, Gibson CM, Perrault LP, Wolf RK,
Kim KB and Emmert MY: Saphenous vein grafts in contemporary
coronary artery bypass graft surgery. Nat Rev Cardiol. 17:155–169.
2020.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Aranki SF and Tatooles AJ: Disconnect
between vein graft failure and clinical events after coronary
artery bypass graft surgery. Circulation. 130:1439–1441.
2014.PubMed/NCBI View Article : Google Scholar
|
|
5
|
de Vries MR, Simons KH, Jukema JW, Braun J
and Quax PH: Vein graft failure: From pathophysiology to clinical
outcomes. Nat Rev Cardiol. 13:451–470. 2016.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Petrie MC, Jhund PS, She L, Adlbrecht C,
Doenst T, Panza JA, Hill JA, Lee KL, Rouleau JL, Prior DL, et al:
Ten-Year outcomes after coronary artery bypass grafting according
to age in patients with heart failure and left ventricular systolic
dysfunction: An analysis of the extended Follow-Up of the STICH
trial (surgical treatment for ischemic heart failure). Circulation.
134:1314–1324. 2016.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Escaned J: Secondary revascularization
after CABG surgery. Nat Rev Cardiol. 9:540–549. 2012.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Lenzen MJ, Boersma E, Bertrand ME, Maier
W, Moris C, Piscione F, Sechtem U, Stahle E, Widimsky P, de Jaegere
P, et al: Management and outcome of patients with established
coronary artery disease: The Euro Heart Survey on coronary
revascularization. Eur Heart J. 26:1169–1179. 2005.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Brilakis ES, Rao SV, Banerjee S, Goldman
S, Shunk KA, Holmes DR Jr, Honeycutt E and Roe MT: Percutaneous
coronary intervention in native arteries versus bypass grafts in
prior coronary artery bypass grafting patients: A report from the
National Cardiovascular Data Registry. JACC Cardiovasc Interv.
4:844–850. 2011.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Harskamp RE, Alexander JH, Schulte PJ,
Brophy CM, Mack MJ, Peterson ED, Williams JB, Gibson CM, Califf RM,
Kouchoukos NT, et al: Vein graft preservation solutions, patency,
and outcomes after coronary artery bypass graft surgery: Follow-up
from the PREVENT IV randomized clinical trial. JAMA Surg.
149:798–805. 2014.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Fiedler J, Baker AH, Dimmeler S, Heymans
S, Mayr M and Thum T: Non-coding RNAs in vascular disease-from
basic science to clinical applications: Scientific update from the
Working Group of Myocardial Function of the European Society of
Cardiology. Cardiovasc Res. 114:1281–1286. 2018.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Quinn JJ and Chang HY: Unique features of
long non-coding RNA biogenesis and function. Nat Rev Genet.
17:47–62. 2016.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Halushka PV, Goodwin AJ and Halushka MK:
Opportunities for microRNAs in the crowded field of cardiovascular
biomarkers. Annu Rev Pathol. 14:211–238. 2019.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Wang XW, He XJ, Lee KC, Huang C, Hu JB,
Zhou R, Xiang XY, Feng B and Lu ZQ: MicroRNA-221 sponge therapy
attenuates neointimal hyperplasia and improves blood flows in vein
grafts. Int J Cardiol. 208:79–86. 2016.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Garber JC, Barbee RW, Bielitzki JT,
Clayton LA, Donovan JC, Hendriksen CF, Kohn DF, Lipman NS, Locke
PA, Melcher J, et al: Guide for the Care and Use of Laboratory
Animals. 8th edition. National Research Council of The National
Academies, Washington, DC, 2011.
|
|
16
|
Kuleshov MV, Jones MR, Rouillard AD,
Fernandez NF, Duan Q, Wang Z, Koplev S, Jenkins SL, Jagodnik KM,
Lachmann A, et al: Enrichr: A comprehensive gene set enrichment
analysis web server 2016 update. Nucleic Acids Res. 44:W90–W97.
2016.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Lucaciu O, Băciuţ M, Băciuţ G, Câmpian RS
and Ionel A: The Gephi Network visualisation of the guided bone
regeneration process induced with tissue engineered grafts. In: Key
Engineering Materials. Trans Tech Publ. 695:215–221. 2016.
|
|
18
|
Zhao Y, Tang N, Xi D, Huang Z, Zhang T,
Liu Y, Wang L, Tang Y, Zhong H and He F: Calcilytic NPS2143
promotes proliferation and inhibits apoptosis of spontaneously
hypertensive rat vascular smooth muscle cells via activation of the
renin-angiotensin system. Exp Ther Med. 20:818–829. 2020.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Feng X, Liu P, Zhou X, Li MT, Li FL, Wang
Z, Meng Z, Sun YP, Yu Y, Xiong Y, et al: Thromboxane A2 Activates
YAP/TAZ protein to induce vascular smooth muscle cell proliferation
and migration. J Biol Chem. 291:18947–18958. 2016.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Simo-Cheyou ER, Vardatsikos G and
Srivastava AK: Src tyrosine kinase mediates endothelin-1-induced
early growth response protein-1 expression via MAP kinase-dependent
pathways in vascular smooth muscle cells. Int J Mol Med.
38:1879–1886. 2016.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Thiel G, Mayer SI, Müller I, Stefano L and
Rössler OG: Egr-1-A Ca(2+)-regulated transcription factor. Cell
Calcium. 47:397–403. 2010.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Beneit N, Martín-Ventura JL, Rubio-Longás
C, Escribano Ó, García-Gómez G, Fernández S, Sesti G, Hribal ML,
Egido J, Gómez-Hernández A and Benito M: Potential role of insulin
receptor isoforms and IGF receptors in plaque instability of human
and experimental atherosclerosis. Cardiovasc Diabetol.
17(31)2018.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Wang Z and Yan X: CD146, a
multi-functional molecule beyond adhesion. Cancer Lett.
330:150–162. 2013.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Jiang T, Zhuang J, Duan H, Luo Y, Zeng Q,
Fan K, Yan H, Lu D, Ye Z, Hao J, et al: CD146 is a coreceptor for
VEGFR-2 in tumor angiogenesis. Blood. 120:2330–2339.
2012.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Colomb F, Wang W, Simpson D, Zafar M,
Beynon R, Rhodes JM and Yu LG: Galectin-3 interacts with the
cell-surface glycoprotein CD146 (MCAM, MUC18) and induces secretion
of metastasis-promoting cytokines from vascular endothelial cells.
J Biol Chem. 292:8381–8389. 2017.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Luo Y, Duan H, Qian Y, Feng L, Wu Z, Wang
F, Feng J, Yang D, Qin Z and Yan X: Macrophagic CD146 promotes foam
cell formation and retention during atherosclerosis. Cell Res.
27:352–372. 2017.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Xie Y, Ostriker AC, Jin Y, Hu H, Sizer AJ,
Peng G, Morris AH, Ryu C, Herzog EL, Kyriakides T, et al: LMO7 is a
negative feedback regulator of transforming growth factor β
signaling and fibrosis. Circulation. 139:679–693. 2019.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Cooley BC, Nevado J, Mellad J, Yang D, St
Hilaire C, Negro A, Fang F, Chen G, San H, Walts AD, et al: TGF-β
signaling mediates endothelial-to-mesenchymal transition (EndMT)
during vein graft remodeling. Sci Transl Med.
6(227ra234)2014.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Greer P: Closing in on the biological
functions of Fps/Fes and Fer. Nat Rev Mol Cell Biol. 3:278–289.
2002.PubMed/NCBI View
Article : Google Scholar
|
|
30
|
He J, Bao Q, Yan M, Liang J, Zhu Y, Wang C
and Ai D: The role of Hippo/yes-associated protein signalling in
vascular remodelling associated with cardiovascular disease. Br J
Pharmacol. 175:1354–1361. 2018.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Ono H, Ichiki T, Ohtsubo H, Fukuyama K,
Imayama I, Hashiguchi Y, Sadoshima J and Sunagawa K: Critical role
of Mst1 in vascular remodeling after injury. Arterioscler Thromb
Vasc Biol. 25:1871–1876. 2005.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Wang X, Hu G, Gao X, Wang Y, Zhang W,
Harmon EY, Zhi X, Xu Z, Lennartz MR, Barroso M, et al: The
induction of yes-associated protein expression after arterial
injury is crucial for smooth muscle phenotypic modulation and
neointima formation. Arterioscler Thromb Vasc Bio. 32:2662–2669.
2012.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Wu G, Cai J, Han Y, Chen J, Huang ZP, Chen
C, Cai Y, Huang H, Yang Y, Liu Y, et al: LincRNA-p21 regulates
neointima formation, vascular smooth muscle cell proliferation,
apoptosis, and atherosclerosis by enhancing p53 activity.
Circulation. 130:1452–1465. 2014.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Wang Z, Zhang XJ, Ji YX, Zhang P, Deng KQ,
Gong J, Ren S, Wang X, Chen I, Wang H, et al: The long noncoding
RNA Chaer defines an epigenetic checkpoint in cardiac hypertrophy.
Nat Med. 22:1131–1139. 2016.PubMed/NCBI View
Article : Google Scholar
|
|
35
|
Yahagi K, Kolodgie FD, Otsuka F, Finn AV,
Davis HR, Joner M and Virmani R: Pathophysiology of native
coronary, vein graft, and in-stent atherosclerosis. Nat Rev
Cardiol. 13:79–98. 2016.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Storey RF: Exploring mechanisms of graft
occlusion toward improved outcomes in coronary artery bypass graft
surgery. J Am Coll Cardiol. 57:1078–1080. 2011.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Jelaković B, Nikolić J, Radovanović Z,
Nortier J, Cosyns JP, Grollman AP, Bašić-Jukić N, Belicza M, Bukvić
D, Čavaljuga S, et al: Consensus statement on screening, diagnosis,
classification and treatment of endemic (Balkan) nephropathy.
Nephrol Dial Transplant. 29:2020–2027. 2014.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Sabik JF II: Understanding saphenous vein
graft patency. Circulation. 124:273–275. 2011.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Lee R, Margaritis M, Channon KM and
Antoniades C: Evaluating oxidative stress in human cardiovascular
disease: Methodological aspects and considerations. Curr Med Chem.
19:2504–2520. 2012.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Tóth A, Balogh E and Jeney V: Regulation
of vascular calcification by reactive oxygen species. Antioxidants
(Basel). 9(963)2020.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Muslin AJ: MAPK signalling in
cardiovascular health and disease: Molecular mechanisms and
therapeutic targets. Clin Sci (Lond). 115:203–218. 2008.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Garcia JG, Liu F, Verin AD, Birukova A,
Dechert MA, Gerthoffer WT, Bamberg JR and English D: Sphingosine
1-phosphate promotes endothelial cell barrier integrity by
Edg-dependent cytoskeletal rearrangement. J Clin Invest.
108:689–701. 2001.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Ullrich A, Gray A, Tam AW, Yang-Feng T,
Tsubokawa M, Collins C, Henzel W, Le Bon T, Kathuria S, Chen E, et
al: Insulin-like growth factor I receptor primary structure:
Comparison with insulin receptor suggests structural determinants
that define functional specificity. EMBO J. 5:2503–2512.
1986.PubMed/NCBI
|