Introduction
Colon cancer is a malignant tumor and is the third
most common cancer worldwide which is expected to increase by 60%
to >2.2 million new cases and 1.1 million deaths by
2030(1). Colon cancer is a serious
threat to public health due to its difficulty to operate on and its
ability to easily metastasize (1,2). A
number of factors may cause colon cancer, such as environment,
lifestyle (including lack of exercise, which can lead to obesity)
and eating habits (excessive intake of saturated fats and low
vegetable and fruit intake) (3,4). The
current treatment of colon cancer mainly involves surgery,
supplemented by chemotherapy and improved diet. Early diagnosis by
endoscopy is a primary goal in diagnosing and treating colon cancer
(5). Therefore, chemotherapy serves
an important role in the clinical treatment of colon cancer
(5).
Current commonly used chemotherapeutic drugs for
colon cancer include 5-fluorouracil (5-FU) (6), oxaliplatin (6), capecitabine (7) and irinotecan (8). These drugs are cytotoxic, often due to
insufficient selectivity of the drug to the site of colon cancer,
resulting in a series of adverse reactions, such as neutropenia,
anemia, diarrhea, gastrointestinal toxicity, mucositis, nausea and
vomiting, as well as blood system diseases and liver toxicity
(6-8).
In addition, drug resistance may also develop, which seriously
affects the quality of life and lifespan of patients (6-8).
Compared with traditional chemotherapeutics, several natural
biological compounds (9), including
artemisinin, celastrol and resveratrol, present unique advantages,
such as less toxicity, fewer side effects and pain relief for
patients with tumors. Therefore, researchers have focused on
discovering novel antitumor drugs from natural sources.
Plant extracts have demonstrated unique advantages
in preventing colon cancer. For example, a previous study (10) revealed that piperine can affect HT29
cells, which is a colon cancer cell line, and inhibit tumor cell
proliferation. Moreover, piperine has been indicated to increase
the production of reactive oxygen species, activate endoplasmic
reticulum emergency-associated proteins, inhibit the
phosphorylation of AKT and JNK and activate p38 MAPK, which can
eventually induce cells to undergo caspase-mediated mitochondrial
apoptosis (10). Other studies have
also demonstrated that anthocyanins are non-toxic to normal
intestinal cell lines in vitro (11,12),
and can inhibit human colon cancer cell growth and induce apoptosis
in a dose- and time-dependent manner (13).
Gallic acid (GA) is a type of natural polyphenol
compound that exists in nature, and it is widely found in several
Chinese medicines, such as green tea, Ganoderma lucidum and
other plants (14). GA has been
indicated to exert important anti-inflammatory (15) and antiviral (16) effects. Of note, GA can inhibit the
proliferation of tumor cells. For instance, several studies have
indicated that GA exhibited an inhibitory effect on lung cancer and
esophageal cancer cells (17,18).
However, to date, there are few systematic and comprehensive
studies on GA in colon cancer, and the underlying mechanism needs
to be further elucidated.
Therefore, the present study was designed to
investigate whether GA could be used to inhibit the proliferation
of colon cancer cells and determine the potential mechanisms via
in vivo and in vitro experiments.
Materials and methods
Cells and grouping
The human colon cancer cell lines HCT116 (cat. no.
BNCC337692) and HT29 (cat. no. BNCC100164) were purchased from BeNa
Culture Collection, Beijing Beina Chuanglian Institute of
Biotechnology. Cells were cultured in RPMI-1640 medium (Beijing
Solarbio Science & Technology Co., Ltd.) supplemented with 10%
FBS (Beijing Solarbio Science & Technology Co., Ltd.) and 1%
penicillin-streptomycin at 37˚C with 5% CO2. Cell growth
was observed under a light microscope, and when cells reached 80%
confluence, they were digested and passaged.
The aforementioned cells were grouped as follows: i)
Control group, no treatment; ii) GA high-dose group (GA-H), cells
stimulated with 60 µM GA (cat no. G131992; Shanghai Aladdin
Biochemical Technology Co., Ltd.) dissolved in PBS for 24 h; iii)
GA medium-dose group (GA-M), cells stimulated with 30 µM GA for 24
h; iv) GA low-dose group (GA-L), cells stimulated with 15 µM GA for
24 h (19); and v) positive drug
control group (5-FU), cells stimulated with 10 µM 5-FU (cat. no.
F0123.0005; Duchefa Biochemie) for 24 h (20). The experiments were performed in
three parallel groups per condition.
Subsequently, to verify whether GA could inhibit SRC
and EGFR phosphorylation to reduce colon cancer proliferation,
cells were divided into five groups as follows: i) Control group;
ii) GA-H group; iii) PP2 group, cells stimulated with 5 µM SRC
inhibitor PP2 (cat. no. P125361; Shanghai Aladdin Biochemical
Technology Co., Ltd.) for 24 h (21); iv) gefinitib group, cells stimulated
with 10 µM EGFR antagonist gefinitib (Nanjing Duly Biotechnology
Co. Ltd.) for 24 h (22); and v)
GA-H + PP2 group, cells simultaneously stimulated with 60 µM GA and
5 µM PP2 for 24 h.
Plate clone formation assay
When HCT116 and HT29 cells reached the logarithmic
growth phase, they were uniformly seeded in a six-well plate (500
cells/well) and cultured at 37˚C with 5% CO2 for 2-3
weeks. When the number of cells reached 50-150, the formed colony
is visible. Once formed colonies were visible, cells were fixed
with 4% paraformaldehyde for 15 min at room temperature. Cells were
subsequently stained with Giemsa (cat. no. D010-1-2; Nanjing
Jiancheng Bioengineering Institute) for 25 min at room temperature.
Following washing by PBS, the colonies were dried at room
temperature for 72 h. And image of each well was captured under
x100 magnification using a light microscopy (Olympus BX51; Olympus
Corporation). Finally, ImageJ v6.0 software (National Institutes of
Health) was used to analyze the colony number.
Flow cytometry
After HCT116 and HT29 cells were cultured for 24 h
in a 5% CO2 incubator at 37˚C, the supernatant was
collected and washed with PBS. A total of 2x105 cells
were collected by trypsin digestion. Cells were then resuspended in
100 µl binding buffer, followed by the addition of 400 µl PBS. The
assay was performed according to the manufacturer's instructions
(cat. no. G003-1-2; Nanjing Jiancheng Bioengineering Institute). A
total of 5 µl PI staining solution and 10 µl Annexin V-FITC were
added to each tube, and cells were incubated in the dark for 15 min
at room temperature to detect apoptosis by Coulter@
EPICS® XLTM flow cytometry (Beckman Coulter,
Inc.). The results were analyzed using CytExpert (version 2.0;
Beckman Coulter, Inc.) software.
Western blotting
Total protein from each group of HCT116 and HT29
cells or tumor tissues was extracted using RIPA buffer containing
protein inhibitors (cat. no. W062-1-1; Nanjing Jiancheng
Bioengineering Institute). Following centrifugation at 12,000 x g
for 10 mins at 4˚C, the protein concentration of the supernatant
was quantified using a bicinchoninic acid protein assay kit
(Nanjing Jiancheng Bioengineering Institute). In total, 50 µg
protein samples were separated by 10% SDS-PAGE and then transferred
to PVDF membranes for 2 h. After blocking with 5% non-fat dry milk
at room temperature for 2 h, primary antibodies against cleaved
caspase-9 (cat. no. orb227889; 1:1,000; Biorbyt Ltd.),
pro-caspase-9 (ab138412; 1:800), cleaved caspase-3 (cat. no.
ab49822; 1:500), pro-caspase-3 (ab32150; 1:1,000; all from Abcam),
Ki-67 (cat. no. orb475270; 1:200; Biorbyt Ltd.), rabbit anti-human
phosphorylated (p)-EGFR (cat. no. ab32578; 1:1,000), EGFR (cat. no.
ab52894; 1:1,000), p-AKT (cat. no. ab38449; 1:1,000), AKT (cat. no.
ab8805; 1:500), p-SRC (cat. no. ab32078; 1:5,000), SRC (cat. no.
ab109381; 1:10,000), p-STAT3 (cat. no. ab76315; 1:2,000), STAT3
(cat. no. ab68153; 1:1,000) and β-actin (cat. no. ab8227; 1:1,000;
all from Abcam) were added to the membranes and incubated overnight
at 4˚C. The membranes were washed with 1X TBST (containing 0.1%
Tween-20) and incubated with goat anti-rabbit horseradish
peroxidase conjugated IgG secondary antibody (cat. no. ab6721;
1:2,000; Abcam) for 1 h at room temperature. The membranes were
washed again with 1X TBS-T and protein bands were exposed using an
ECL kit (cat. no. W028-2-1; Nanjing Jiancheng Bioengineering
Institute). The relative expression of target proteins was analyzed
by Image J v6.0 software (National Institutes of Health) and
calculated based on β-actin as the internal reference.
Immunofluorescence
HCT116 and HT29 cells were fixed with 4%
paraformaldehyde for 20 min at room temperature. The cells were
then permeabilized with 0.5% Triton X-100 for 20 min and blocked
with 5% BSA (Wuhan Boster Biological Technology, Ltd.) for 40 min,
both at room temperature. Subsequently, the cells were incubated
with p-STAT3 (cat. no. ab76315; 1:500), p-SRC (cat. no. ab32078;
1:50), p-EGFR (cat. no. ab32578; 1:25; all from Abcam) and p-AKT
(cat. no. orb312153; 1:100; Biorbyt Ltd.) antibodies overnight at
4˚C. After washing with PBS, the cells were incubated with goat
anti-rabbit IgG H&L (Alexa Fluor® 488; cat. no.
ab150077; 1:200; Abcam) at 37˚C for 1.5 h. Following three washes
with PBS, the nuclei were stained with 5 µg/ml DAPI for 5 min at
room temperature. Immunofluorescence was observed by confocal
microscopy (Nikon Corporation) with the magnification of x400 and
the mean fluorescence intensity were quantified using Image J v6.0
software (National Institutes of Health).
Animals
A total of 48 specific-pathogen-free BALB/c female
nude mice (16-18 g; 4 weeks old) were purchased from Beijing
Weitong Lihua Laboratory Animal Technology Co., Ltd. [SCXK
(Beijing) 20160006]. Operations were performed after 7 days of
adaptive feeding. Animal experiments in the present study were
agreed and approved by the Animal Protection and Use Committee of
Yantai University (Yantai, China). The animals were raised at
26-28˚C, 40-60% humidity, light/dark cycle for 12-h and free access
to water and food. The food, drinking water and bedding materials
were sterilized. Animal experiments followed the National
Institutes of Health guidelines (NIH publication no. 85-23, revised
1996).
Xenograft tumor model
HCT116 and HT29 cells were suspended with RPMI-1640
medium without 10% FBS. A total of 0.2 ml HCT116 and HT29 cell
suspended with PBS (5x107 cells/ml) were inoculated
subcutaneously into the left backs of nude mice in the
corresponding group. If a rice-sized tumor (~50 mm3)
appeared at the inoculation site on the 7th day, the inoculation
was considered to be successful. Nude mice were randomly divided
into four groups (n=6): i) Model, mice inoculated with cancer
cells; ii) GA group, following inoculation of cancer cells, mice
were injected daily with 80 mg/kg/day GA (23); iii) PP2 group, following inoculation
of cancer cells, mice were injected daily with 10 mg/kg/day
PP2(24); iv) gefitinib group,
following inoculation of cancer cells, mice were injected daily
with 50 mg/kg/day gefitinib (25);
and v) GA + PP2 group, following inoculation of cancer cells, mice
were injected daily with 80 mg/kg/day GA and 10 mg/kg/day PP2.
Continuous administration was performed for 28 days. Tumor volumes
were recorded every 7 days. At the end of the experiment, the mice
were anesthetized by intraperitoneal injection of 3% sodium
pentobarbital (40 mg/kg) and sacrificed by cervical dislocation.
Death was confirmed by cardiac arrest. Tumors were excised,
photographed and weighed.
TUNEL assay
Tumor tissues were fixed with 4% paraformaldehyde
for 24 h at room temperature, dehydrated and embedded in paraffin.
The slices (5 µm) were heated at 60-70˚C for 2 h, dewaxed with
xylene and then rehydrated using a descending ethanol gradient. The
tissue slices were incubated in 3%
H2O2-methanol for 10 min at room temperature
and rinsed with PBS three times (3 min each time). Subsequently,
the slices were stained with TUNEL solution (cat. no. G002-2-2;
Nanjing Jiancheng Bioengineering Institute). A sealing film was
used to perform the reaction in a dark wet box for 1 h at 37˚C.
After TUNEL-staining, DAB solution was added for 5 min at room
temperature. After washing with PBS for three times, the slices
were re-stained using hematoxylin at room temperature for 60 sec,
then dehydrated in ascending series of ethanol and cleared in
xylene. Finally, the slices were sealed with neutral gum and
observed under a light microscope (magnification, x400; Olympus
BX51; Olympus Corporation) from five randomly selected fields. The
number of positive-stained cells was analyzed by ImageJ v6.0
software (National Institutes of Health). Cell apoptosis rate was
calculated using the following formula: Apoptosis rate = (number of
apoptotic cells/total number of cells) x100%.
Statistical analysis
All data were analyzed with SPSS v19.0 (IBM Corp.).
Numerical data are presented as the mean ± SD. Differences among
multiple groups were evaluated by one-way ANOVA followed by Tukey's
post hoc test. P<0.05 was considered to indicate a statistically
significant difference.
Results
GA promotes apoptosis and inhibits
colon cancer cell proliferation
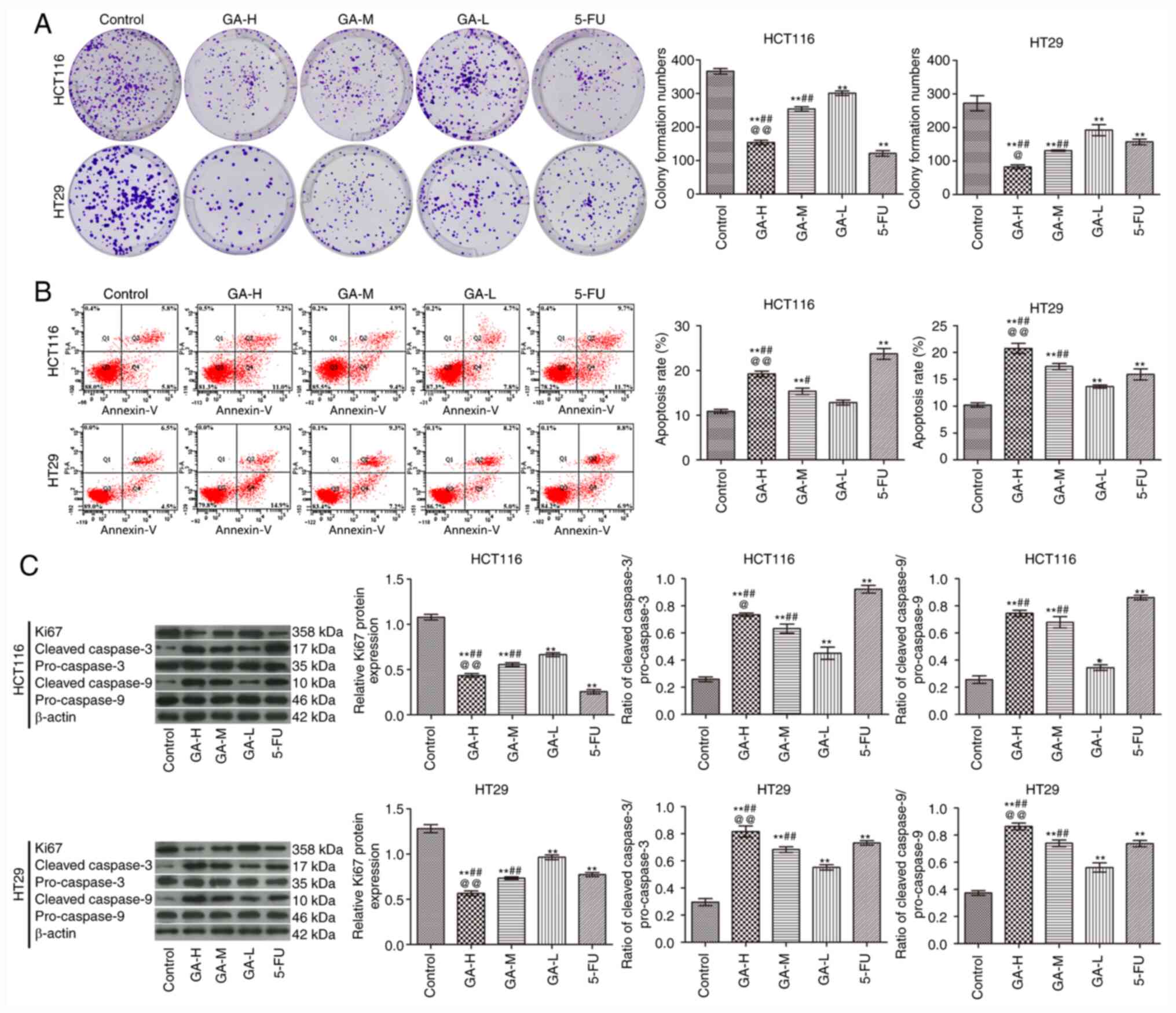
Compared with the control group, GA inhibited the
proliferation (Fig. 1A) and
promoted the apoptosis (Fig. 1B) of
HCT116 and HT29 cells in a concentration-dependent manner. In
addition, GA increased the ratio of cleaved caspase-3/pro-caspase-3
and cleaved caspase-9/pro-caspase-9 and inhibited Ki-67 expression
when compared with the control group (Fig. 1C). Notably, the effects of GA-H
treatment were similar with those observed with 5-FU treatment.
These data demonstrated that GA administration promoted apoptosis
and inhibited proliferation in colon cancer cells.
GA inhibits the phosphorylation of SRC
in colon cancer cells
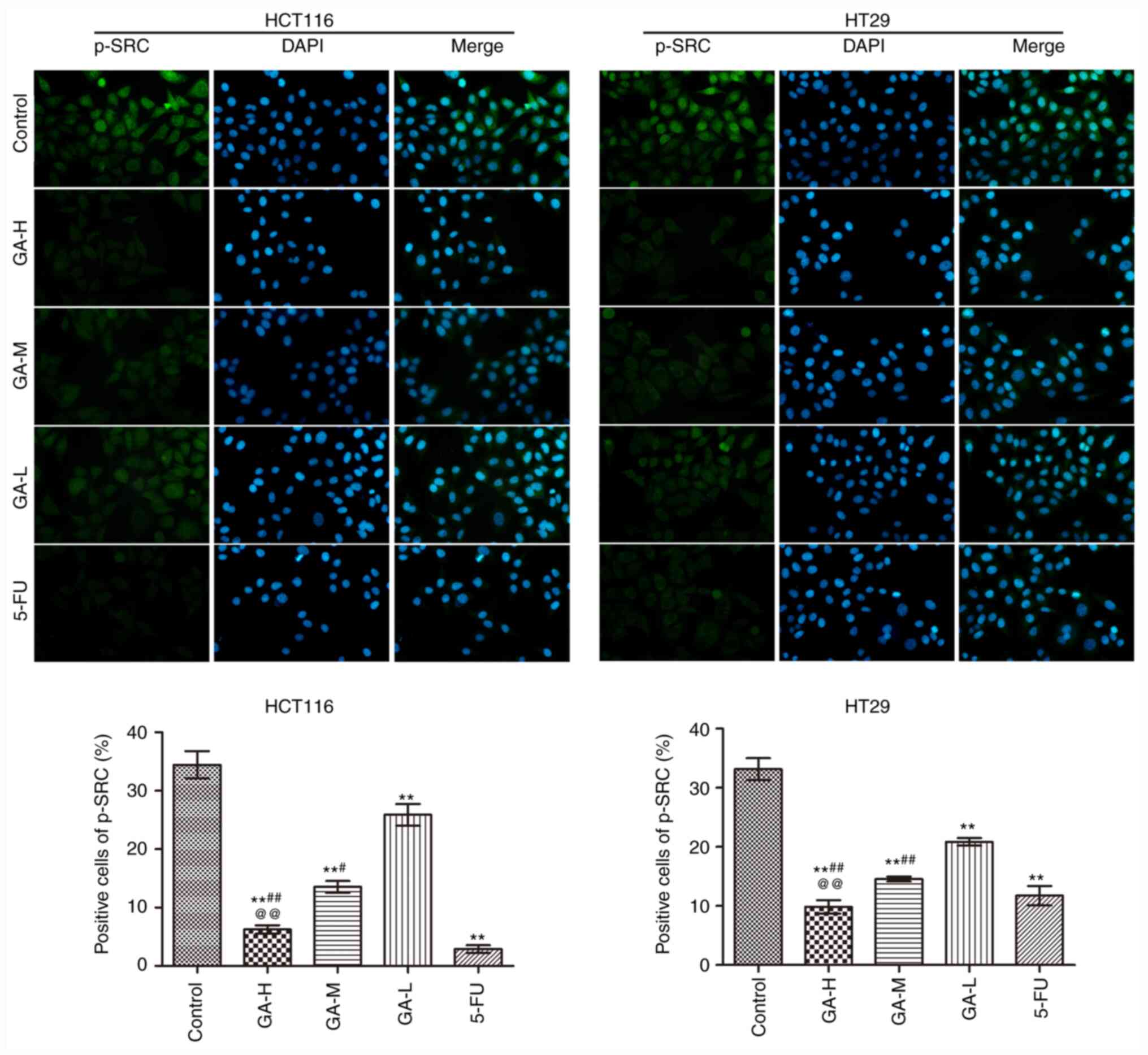
The effects of GA on SRC phosphorylation in HCT116
and HT29 cells were analyzed using immunofluorescence (Fig. 2). GA and 5-FU intervention
significantly reduced the positive p-SRC staining in HCT116 and
HT29 cells compared with the control group, especially in 5-FU.
Moreover, high GA concentrations exerted the strongest inhibitory
effect on SRC phosphorylation in HCT116 cells and HT29 cells. These
data suggested that GA suppressed SRC phosphorylation.
GA decreases STAT3 and AKT
phosphorylation level via inhibiting SRC and EGFR
phosphorylation
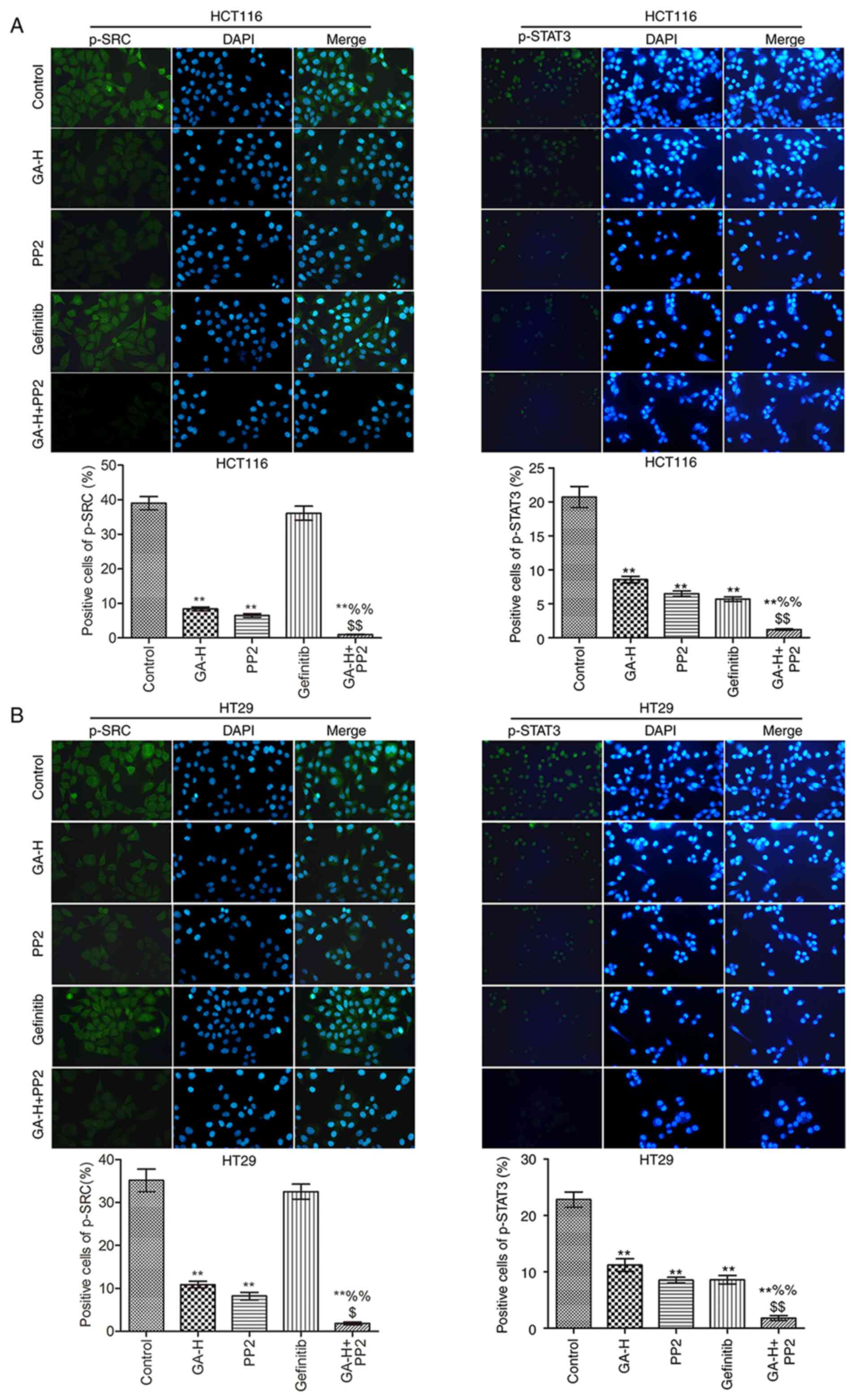
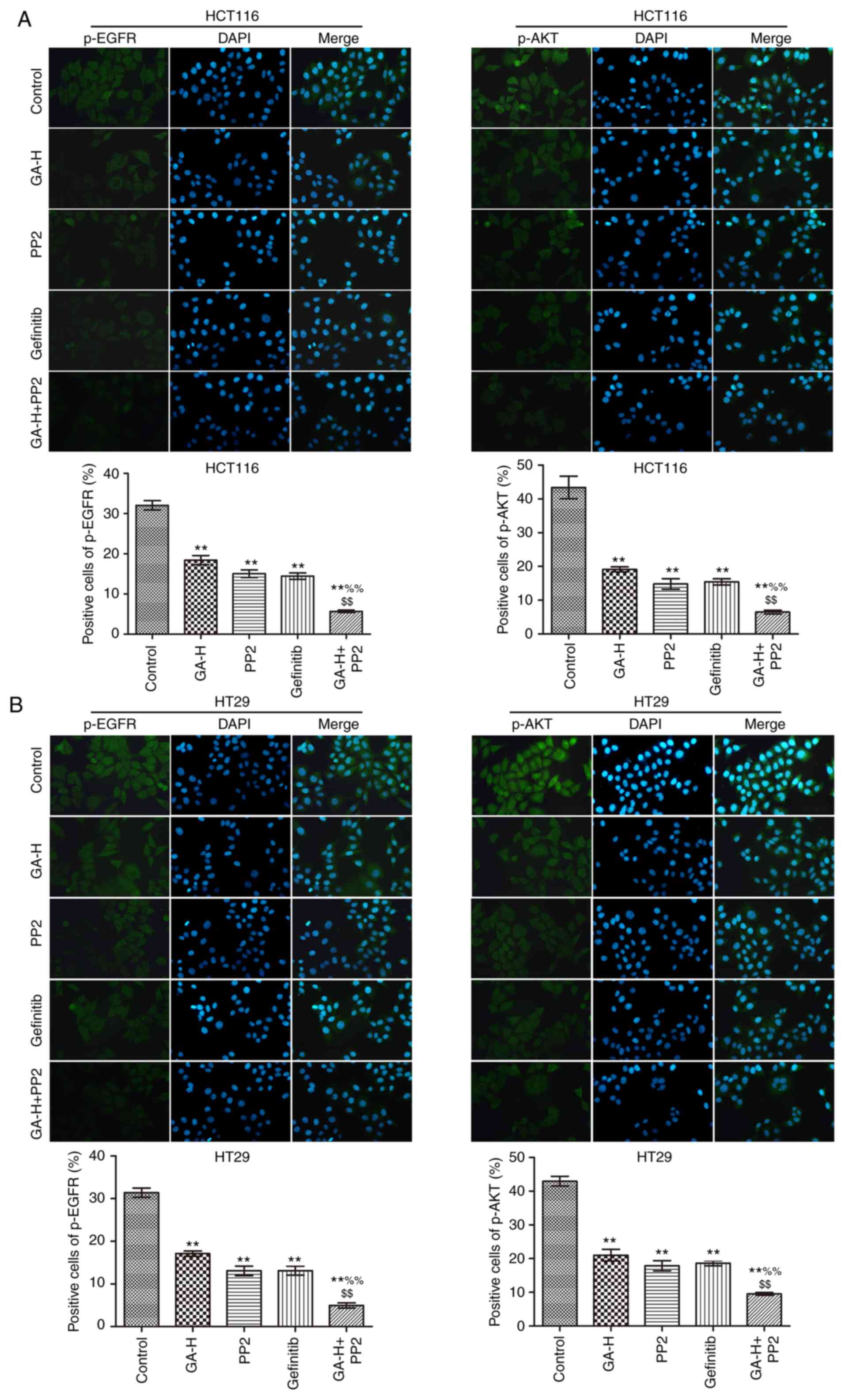
The levels of p-SRC, p-STAT3 (Fig. 3), p-EGFR and p-AKT (Fig. 4) were examined using
immunofluorescence. Compared with the control group, GA-H and PP2
markedly inhibited the level of p-STAT3, p-EGFR, p-AKT and p-SRC,
and gefitinib significantly reduced the level of p-STAT3, p-EGFR
and p-AKT, but had no effect on p-SRC level. When GA-H and PP2 were
used in combination, the phosphorylation level of the
aforementioned proteins was further decreased compared with the
GA-H or PP2 groups alone. These results indicated that GA could
decrease SRC and EGFR phosphorylation level to inhibit STAT3 and
AKT phosphorylation.
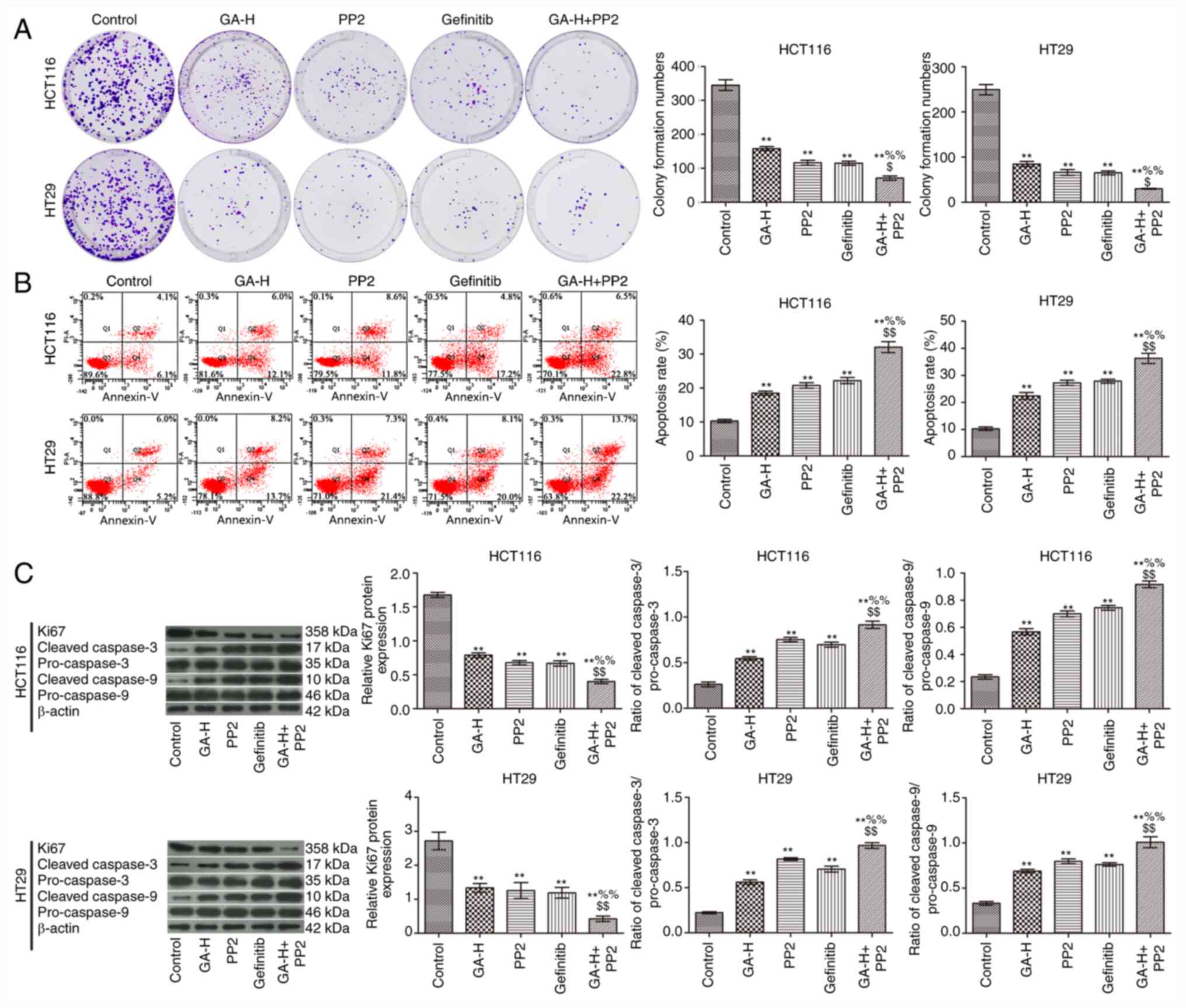
GA decreases SRC and EGFR
phosphorylation level to promote colon cancer cell apoptosis and
inhibit proliferation
To verify whether GA could inhibit SRC and EGFR
phosphorylation to reduce colon cancer cell proliferation, cells
were also treated with PP2 and gefitinib. As indicated in Fig. 5, GA-H, PP2 and gefitinib significantly
inhibited cell proliferation (Fig.
5A) and promoted cell apoptosis (Fig. 5B) compared with the control group.
The combined administration of GA-H and PP2 further promoted cell
apoptosis and inhibited proliferation compared with the GA-H or PP2
groups. In addition, western blot analysis demonstrated that GA-H,
PP2 and gefitinib increased the ratio of cleaved
caspase-3/pro-caspase-3 and cleaved caspase-9/pro-caspase-9, and
decreased Ki-67 protein expression level compared with the control
group. The combined administration of GA-H and PP2 further enhanced
the ratio of cleaved caspase-3/pro-caspase-3 and cleaved
caspase-9/pro-caspase-9, and reduced Ki-67 protein expression level
compared with the GA-H or PP2 groups (Fig. 5C). These results revealed that GA
promoted colon cancer cell apoptosis and inhibited proliferation
via inhibiting SRC and EGFR phosphorylation.
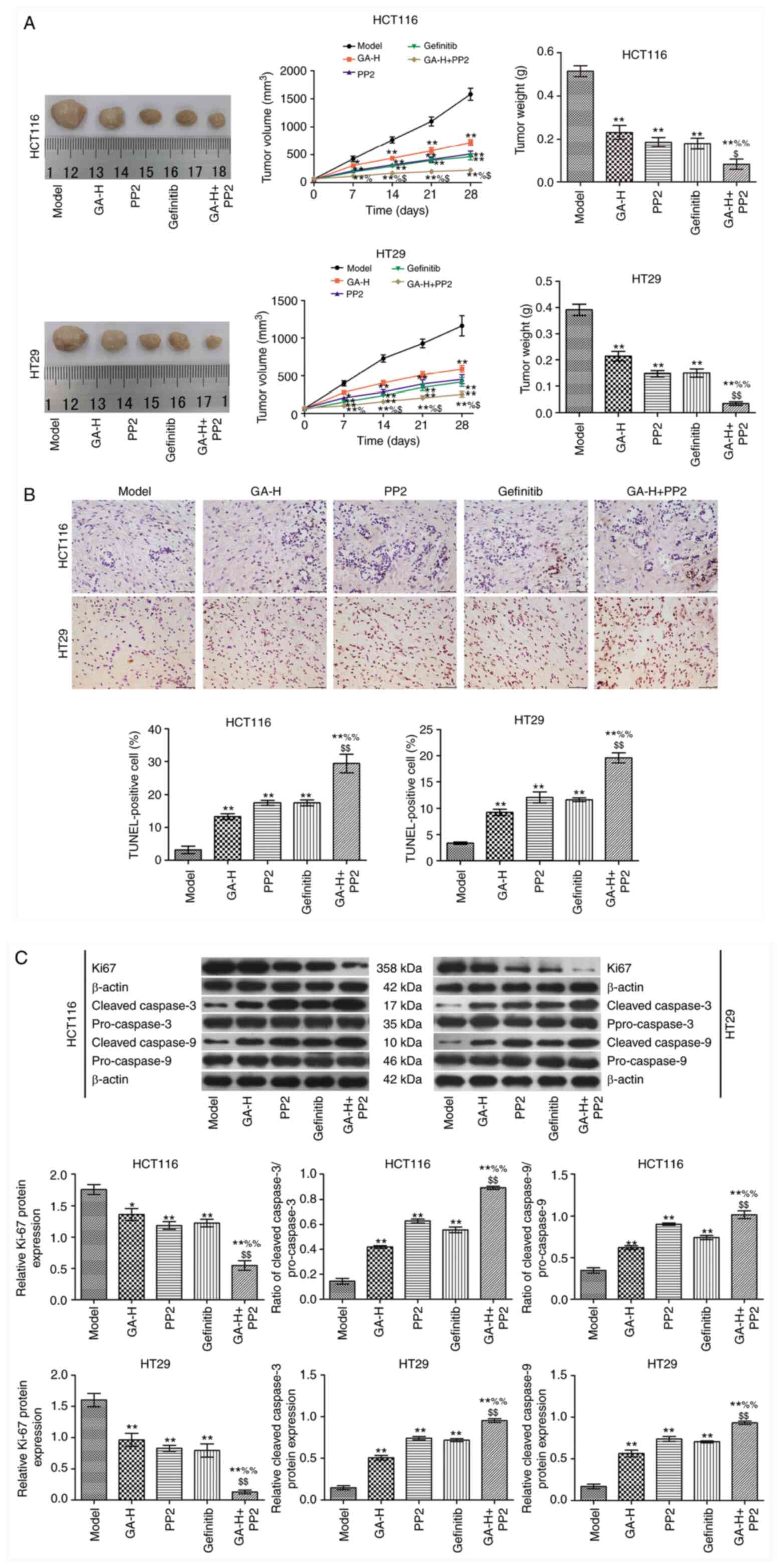
GA decreases SRC and EGFR
phosphorylation level to inhibit tumor growth
In the present study, the maximum tumor diameter was
1.56 cm, and the maximum tumor volume was 1,900.4 mm3.
GA-H, PP2 and gefitinib inhibited tumor growth compared with the
model group (Fig. 6A). When GA-H
and PP2 were administered in combination, tumor growth was
significantly reduced compared with the GA or PP2 groups alone. The
results in Fig. 6B demonstrated
that GA-H, PP2 and gefitinib significantly increased the apoptosis
rate of tumor cells in vivo compared with the model group.
This effect was enhanced in the GA-H + PP2 group compared with the
GA or PP2 groups. The expression level of Ki67 and the ratio of
cleaved caspase-3/pro-caspase-3 and cleaved caspase-9/pro-caspase-9
are presented in Fig. 6C. The
results revealed that GA-H, PP2 and gefitinib decreased Ki-67
expression level and increased the ratio of cleaved
caspase-3/pro-caspase-3 and cleaved caspase-9/pro-caspase-9
compared with the model group. These effects were further enhanced
when GA-H and PP2 were administered concurrently.
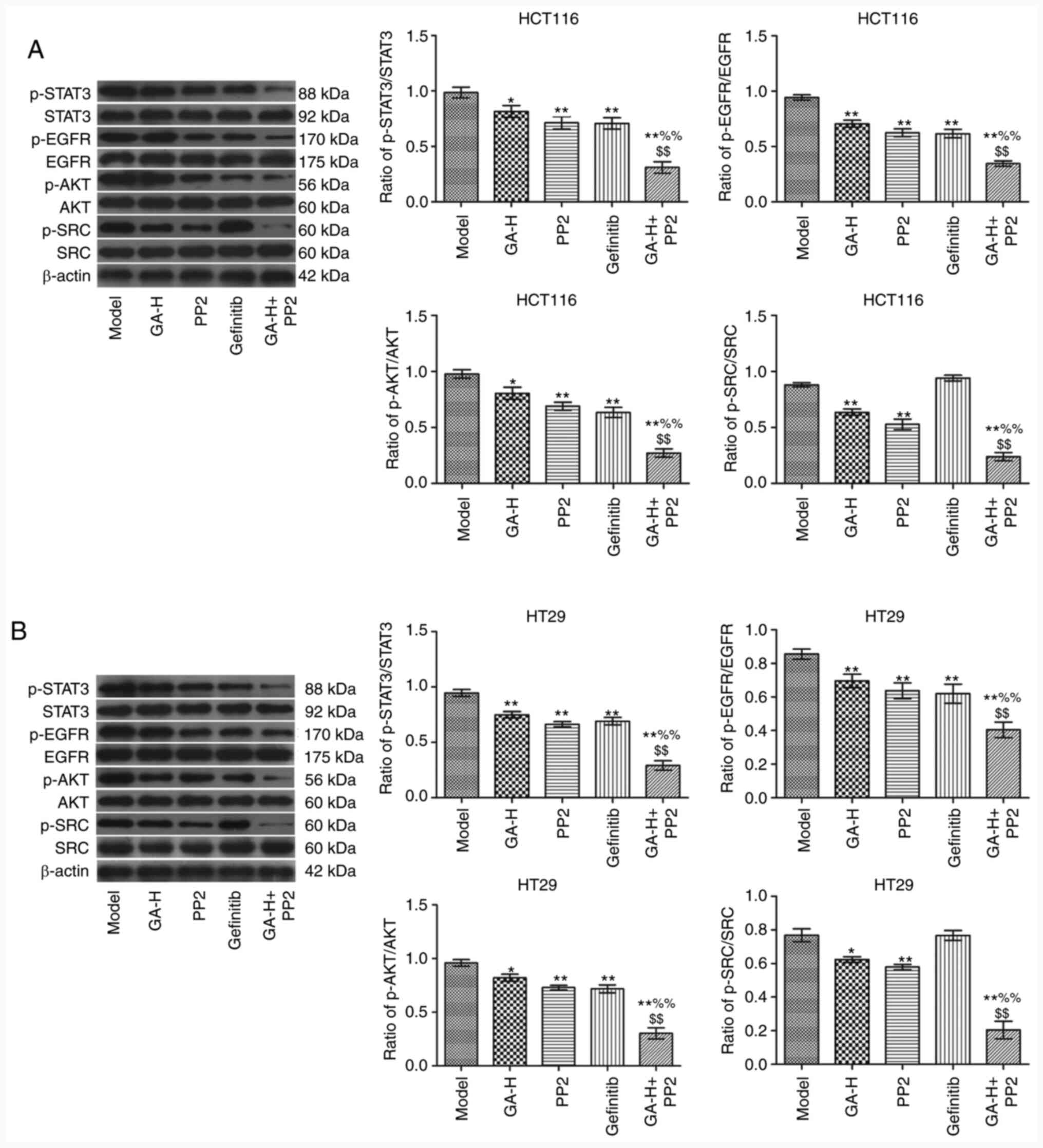
GA inhibits SRC and EGFR
phosphorylation to regulate STAT3 and AKT in vivo
The relative levels of p-STAT3/STAT3, p-EGFR/EGFR,
p-AKT/AKT and p-SRC/SRC in tumor tissues was analyzed by western
blotting (Fig. 7). The results
indicated that the administration of GA-H and PP2 significantly
decreased the ratio of p-STAT3/STAT3, p-EGFR/EGFR, p-AKT/AKT and
p-SRC/SRC in tumor tissues compared with the model group (Fig. 7). Gefitinib significantly reduced
the level of p-STAT3/STAT3, p-EGFR/EGFR and p-AKT/AKT, but did not
affect p-SRC level compared with the model group. Furthermore,
following the combined treatment of GA-H and PP2, the
phosphorylation level of the aforementioned four proteins was
significantly lower compared with the GA-H or PP2 groups.
Discussion
The present study demonstrated that GA significantly
suppressed the proliferation of HCT116 and HT29 colon cancer cells
and accelerated their apoptotic process, which was consistent with
the results of previous studies. In previous studies, GA has been
indicated to inhibit cell proliferation and invasion of different
tumors, such as non-small cell lung cancer (NSCLC) (26,27)
and bladder cancer (28). The
occurrence of colon cancer is closely associated with the abnormal
expression of multiple genes (29).
Therefore, regulating the expression of certain key genes, such as
SRC, during tumorigenesis to inhibit malignant transformation is an
effective means to control tumor growth.
The SRC protein is encoded by the proto-oncogene
Src and is a member of the non-receptor tyrosine protein
kinase family, which can activate multiple signaling pathways, such
as PI3K/AKT, MAPK and STAT3(30).
SRC activity is enhanced in 80% of patients with colon cancer, and
is significantly associated with the occurrence and development of
colon cancer (31,32). In the present study, GA was
administered to HCT116 and HT29 colon cancer cells and colon cancer
animal models. It was revealed that GA treatment significantly
inhibited the proliferation and promoted the apoptosis of colon
cancer cells, and this effect was associated with p-SRC level.
The EGFR signal transduction pathway is also widely
considered to serve a key role in tumor formation and development
and is one of the most important targets for numerous cancers
(33,34). It often exhibits abnormal activation
during cancer progression and is closely associated with poor
prognosis (33). Nam et al
(34) demonstrated that GA induced
apoptosis in EGFR-mutant NSCLC via acceleration of EGFR turnover,
which suggested that GA-mediated inhibition of cancer growth may be
associated with EGFR signaling. Previously, SRC was indicated to
activate EGFR via different pathways to promote tumor proliferation
and metastasis (35). In addition,
STAT3 has been also revealed to be continuously activated in
tumors, which serves crucial roles in regulating the proliferation,
migration and survival of cells and can be also regulated by SRC
(36). In present study, GA was
indicated to inhibit SRC and EGFR phosphorylation resulting in a
decrease in STAT3 and AKT phosphorylation level. When the SRC
inhibitor PP2 and the EGFR inhibitor gefitinib were administered
in vitro or in vivo, these results were further
confirmed. However, Chen et al (37) demonstrated that constitutively
active AKT and MEK-1 or SRC overexpression reversed the inhibitory
effect of GA on the EGF-induced MMP-9 upregulation in breast cancer
cells. It is possible that the reported effects are the results of
the effect of GA on different, independent pathways and not part of
the mechanism mediating the effect on tumor growth.
Few studies have investigated the mechanism of GA in
colon cancer in vivo and in vitro. The present study
demonstrated that GA inhibited colon cancer cell proliferation and
induced apoptosis via inhibiting SRC and EGFR phosphorylation. The
results of the present study may provide novel ideas, strategies
and treatment options for the prevention and targeted therapy of
colon cancer.
Acknowledgements
Not applicable.
Funding
Funding: No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
XL, LH and PL conceived the study and performed
literature research, data analysis, experiments, manuscript writing
and review; GW and YG designed the experiments and were involved in
the data acquisition and statistical analysis; TW was involved in
data acquisition, manuscript preparation and data analysis; KC
performed study design, literature research, experiments and
manuscript editing. XL and GW confirmed the authenticity of the raw
data. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
Animal experiments followed the NIH guidelines (NIH
publication no. 85-23, revised 1996) and were approved by the
Animal Protection and Use Committee of Yantai University (Yantai,
China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Arnold M, Sierra MS, Laversanne M,
Soerjomataram I, Jemal A and Bray F: Global patterns and trends in
colorectal cancer incidence and mortality. Gut. 66:683–691.
2017.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Favoriti P, Carbone G, Greco M, Pirozzi F,
Pirozzi RE and Corcione F: Worldwide burden of colorectal cancer: A
review. Updates Surg. 68:7–11. 2016.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Giftson JS, Jayanthi S and Nalini N:
Chemopreventive efficacy of gallic acid, an antioxidant and
anticarcinogenic polyphenol, against 1,2-dimethyl hydrazine induced
rat colon carcinogenesis. Invest New Drugs. 28:251–259.
2010.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Carole M, Livine D, Daniel K, Järvinen JR,
Annelise L and Muller CD: Impact of procyanidins from different
berries on caspase 8 activation in colon cancer. Oxid Med Cell
Longev. 2015(154164)2015.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Grothey A and Venook AP: Optimizing
adjuvant therapy for localized colon cancer and treatment selection
in advanced colorectal cancer. J Natl Compr Canc Netw. 16:611–615.
2018.PubMed/NCBI View Article : Google Scholar
|
|
6
|
No authors listed: Adjuvant chemotherapy
with oxaliplatin, in combination with fluorouracil plus leucovorin
prolongs disease-free survival, but causes more adverse event
cancer Abstracted from: Andre T, Boni C, Mounedji-Boudiaf L, et
al. Multicenter international study of
oxaliplatin/5-fluorouracil/leucovorin in the adjuvant treatment of
colon cancer (MOSAIC) investigators. Oxaliplatin, fluorouracil, and
leucovorin as adjuvant treatment for colon cancer. N Engl J Med
350, 2343-2351, 2004. Cancer Treat Rev 30, 711-713, 2004.
|
|
7
|
Schmoll HJ, Tabernero J, Maroun J, de
Braud F, Price T, Van Cutsem E, Hill M, Hoersch S, Rittweger K and
Haller DG: Capecitabine plus oxaliplatin compared with
fluorouracil/folinic acid as adjuvant therapy for stage III colon
cancer: Final results of the NO16968 randomized controlled phase
III trial. J Clin Oncol. 33:3733–3740. 2015.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Liu X, Jiang J, Chan R, Ji Y, Lu J, Liao
YP, Okene M, Lin J, Lin P, Chang CH, et al: Improved efficacy and
reduced toxicity using a custom-designed irinotecan-delivering
silicasome for orthotopic colon cancer. ACS Nano. 13:38–53.
2019.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Deng S, Shanmugam MK, Kumar AP, Yap CT,
Sethi G and Bishayee A: Targeting autophagy using natural compounds
for cancer prevention and therapy. Cancer. 125:1228–1246.
2019.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Yaffe PB, Power Coombs MR, Doucette CD,
Walsh M and Hoskin DW: Piperine, an alkaloid from black pepper,
inhibits growth of human colon cancer cells via G1 arrest and
apoptosis triggered by endoplasmic reticulum stress. Mol Carcinog.
54:1070–1085. 2015.PubMed/NCBI View
Article : Google Scholar
|
|
11
|
Venancio VP, Cipriano PA, Kim H, Antunes
LM, Talcott ST and Mertens-Talcott SU: Cocoplum (Chrysobalanus
icaco L.) anthocyanins exert anti-inflammatory activity in
human colon cancer and non-malignant colon cells. Food Funct.
8:307–314. 2017.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Kubow S, Iskandar MM, Melgar-Bermudez E,
Sleno L, Sabally K, Azadi B, How E, Prakash S, Burgos G and Felde
TZ: Effects of simulated human gastrointestinal digestion of two
purple-fleshed potato cultivars on anthocyanin composition and
cytotoxicity in colonic cancer and non-tumorigenic cells.
Nutrients. 9(953)2017.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Mazewski C, Liang K and de Mejia EG:
Inhibitory potential of anthocyanin-rich purple and red corn
extracts on human colorectal cancer cell proliferation in vitro. J
Funct Foods. 34:254–265. 2017.
|
|
14
|
Niemetz R and Gross GG: Enzymology of
gallotannin and ellagitannin biosynthesis. Phytochemistry.
66:2001–2011. 2005.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Kroes BH, van den Berg AJ, Quarles van
Ufford HC, van Dijk H and Labadie RP: Anti-inflammatory activity of
gallic acid. Planta Med. 58:499–504. 1992.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Choi HJ, Song JH, Bhatt LR and Baek SH:
Anti-human rhinovirus activity of gallic acid possessing
antioxidant capacity. Phytother Res. 24:1292–1296. 2010.PubMed/NCBI View
Article : Google Scholar
|
|
17
|
You BR and Park WH: Gallic acid-induced
lung cancer cell death is related to glutathione depletion as well
as reactive oxygen species increase. Toxicol In Vitro.
24:1356–1362. 2010.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Sun GL and Wang D: Gallic acid from
terminalia chebula inhibited the growth of esophageal carcinoma
cells by suppressing the Hippo signal pathway. Iran J Basic Med
Sci. 23:1401–1408. 2020.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Lee HL, Lin CS, Kao SH and Chou MC: Gallic
acid induces G1 phase arrest and apoptosis of triple-negative
breast cancer cell MDA-MB-231 via p38 mitogen-activated protein
kinase/p21/p27 axis. Anticancer Drugs. 28:1150–1156.
2017.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Li J, Hou N, Faried A, Tsutsumi S,
Takeuchi T and Kuwano H: Inhibition of autophagy by 3-MA enhances
the effect of 5-FU-induced apoptosis in colon cancer cells. Ann
Surg Oncol. 16:761–771. 2009.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Kundu J, Choi BY, Jeong CH, Kundu JK and
Chun KS: Thymoquinone induces apoptosis in human colon cancer
HCT116 cells through inactivation of STAT3 by blocking JAK2- and
Src-mediated phosphorylation of EGF receptor tyrosine kinase. Oncol
Rep. 32:821–828. 2014.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Agelaki S, Spiliotaki M, Markomanolaki H,
Kallergi G, Mavroudis D, Georgoulias V and Stournaras C: Caveolin-1
regulates EGFR signaling in MCF-7 breast cancer cells and enhances
gefitinib-induced tumor cell inhibition. Cancer Biol Ther.
8:1470–1477. 2009.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Li ZJ, Liu M, Dawuti G, Dou Q, Ma Y, Liu
HG and Aibai S: Antifungal activity of gallic acid in vitro and in
vivo. Phytother Res. 31:1039–1045. 2017.PubMed/NCBI View
Article : Google Scholar
|
|
24
|
Lou L, Yu Z, Wang Y, Wang S and Zhao Y:
c-Src inhibitor selectively inhibits triple-negative breast cancer
overexpressed vimentin in vitro and in vivo. Cancer. 109:1648–1659.
2018.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Bruzzese F, Di Gennaro E, Avallone A, Pepe
S, Arra C, Caraglia M, Tagliaferri P and Budillon A: Synergistic
antitumor activity of epidermal growth factor receptor tyrosine
kinase inhibitor gefitinib and IFN-alpha in head and neck cancer
cells in vitro and in vivo. Clin Cancer Res. 12:617–625.
2006.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Zhang T, Ma L, Wu P, Li W, Li T, Gu R, Dan
X, Li Z, Fan X and Xiao Z: Gallic acid has anticancer activity and
enhances the anticancer effects of cisplatin in non-small cell lung
cancer A549 cells via the JAK/STAT3 signaling pathway. Oncol Rep.
41:1779–1788. 2019.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Phan AN, Hua TN, Kim MK, Vo VT, Choi JW,
Kim HW, Rho JK, Kim KW and Jeong Y: Gallic acid inhibition of
Src-Stat3 signaling overcomes acquired resistance to EGF receptor
tyrosine kinase inhibitors in advanced non-small cell lung cancer.
Oncotarget. 7:54702–54713. 2016.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Liao CC, Chen SC, Huang HP and Wang CJ:
Gallic acid inhibits bladder cancer cell proliferation and
migration via regulating fatty acid synthase (FAS). J Food Drug
Anal. 26:620–627. 2018.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Yang H, Wu J, Zhang J, Yang Z, Jin W, Li
Y, Jin L, Yin L, Liu H and Wang Z: Integrated bioinformatics
analysis of key genes involved in progress of colon cancer. Mol
Genet Genomic Med. 7(e00588)2019.PubMed/NCBI View
Article : Google Scholar
|
|
30
|
Chen J, Elfiky A, Han M, Chen C and Saif
MW: The role of Src in colon cancer and its therapeutic
implications. Clin Colorectal Cancer. 13:5–13. 2014.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Hurwitz H, Fehrenbacher L, Novotny W,
Cartwright T, Hainsworth J, Heim W, Berlin J, Baron A, Griffing S,
Holmgren E, et al: Bevacizumab plus irinotecan, fluorouracil, and
leucovorin for metastatic colorectal cancer. N Engl J Med.
350:2335–2342. 2004.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Xie G, Peng Z and Raufman JP: Src-mediated
aryl hydrocarbon and epidermal growth factor receptor cross talk
stimulates colon cancer cell proliferation. Am J Physiol
Gastrointest Liver Physiol. 302:G1006–G1015. 2012.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Atmaca A, Werner D, Pauligk C, Steinmetz
K, Wirtz R, Altmannsberger HM, Jäger E and Al-Batran SE: The
prognostic impact of epidermal growth factor receptor in patients
with metastatic gastric cancer. BMC Cancer. 12(524)2012.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Nam B, Rho JK, Shin DM and Son J: Gallic
acid induces apoptosis in EGFR-mutant non-small cell lung cancers
by accelerating EGFR turnover. Bioorg Med Chem Lett. 26:4571–4575.
2016.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Liu ZM and Huang HS: As2O3-induced
c-Src/EGFR/ERK signaling is via Sp1 binding sites to stimulate
p21WAF1/CIP1 expression in human epidermoid carcinoma A431 cells.
Cell Signal. 18:244–255. 2006.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Sen B, Saigal B, Parikh N, Gallick G and
Johnson FM: Sustained Src inhibition results in signal transducer
and activator of transcription 3 (STAT3) activation and cancer cell
survival via altered Janus-activated kinase-STAT3 binding. Cancer
Res. 69:1958–1965. 2009.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Chen YJ, Lin KN, Jhang LM, Huang CH, Lee
YC and Chang LS: Gallic acid abolishes the
EGFR/Src/Akt/Erk-mediated expression of matrix metalloproteinase-9
in MCF-7 breast cancer cells. Chem Biol Interact. 252:131–140.
2016.PubMed/NCBI View Article : Google Scholar
|