Introduction
The incidence of thyroid cancer has significantly
increased in recent years (1).
Currently, ≥90% of patients with thyroid cancer are diagnosed with
differentiated thyroid cancer (DTC), which mainly includes
papillary thyroid cancer (PTC) and follicular thyroid cancer (FTC)
(1). Radioactive iodine
(131I) can be taken up by DTC cells and emit β-rays,
which exert a radiotherapeutic effect on DTC cells. Patients who
are resistant to 131I therapy, due to the inability to
take up 131I or the occurrence of radioresistance in
distant lesions, have a poor prognosis and shorter survival
(2-4).
The uptake of 131I by metastatic DTC is facilitated by a
series of iodine metabolism genes, among which the solute carrier
family 5 member 5 (NIS) protein has been reported to serve a key
role in 131I uptake and treatment (3). However, the current understanding of
radioresistance following 131I treatment in DTC remains
limited.
MicroRNAs (miRNAs/miRs) are a class of
mononucleotide, small non-coding RNAs that combine with the
3'-untranslated region (3'-UTR) of target genes to suppress target
gene expression and have a wide range of biological functions
(5). Notably, miR-221 and miR-222
share the same promoter, have a highly homologous sequence and
share the same seed site. The expression levels of miR-221 and
miR-222 in bladder cancer (6),
breast cancer (7) and other types
of tumor tissues or cell lines were found to be significantly
upregulated compared with those in normal tissues (8). In addition, miR-221 and miR-222 were
demonstrated to play important roles in regulating tumor cell
functions, such as invasion, metastasis, epithelial-to-mesenchymal
transition (EMT), proliferation and resistance to treatment, by
targeting target genes, such as suppressor of cytokine signaling 3
(SOCS3), transcriptional repressor GATA binding 1 and p27 (8,9). It
has also been demonstrated that the expression levels of miR-221
and miR-222 were significantly upregulated in DTC tissues compared
with those in normal thyroid tissues (10-12).
Furthermore, the expression levels of miR-221 and miR-222 in the
blood of patients with thyroid cancer were found to be associated
with the progression of DTC (13,14). A
recent large-scale meta-analysis revealed that the expression
levels of miR-221 and miR-222 could predict poor overall survival
in patients with cancer. In particular, miR-222 exhibited a
significant predictive value for secondary outcomes, including
disease-free and recurrence-free survival (15).
Signal transducer and activator of transcription 3
(STAT3) regulates a variety of cell functions, including EMT,
through its downstream gene signaling pathways. Treatment of
thyroid cancer cells with the STAT3 inhibitor, cucurbitacin I, was
shown to upregulate the expression levels of thyroid-specific genes
and significantly enhance 131I uptake; furthermore,
cucurbitacin I treatment enhanced the sensitivity of thyroid cancer
cells to radiation and chemotherapy (16). SOCS3 is a cytokine-inducible
negative regulator of cytokine signaling, which can bind to Janus
kinase (JAK)2 and inhibit the activity of the JAK/STAT signaling
pathway (9). The genetic silencing
of the miR-221/miR-222 cluster was shown to attenuate angiogenesis
in glioblastoma by inactivating the JAK/STAT signaling pathway via
the upregulation of SOCS3(9). This
finding suggested that miR-221 and miR-222 may activate the STAT3
signaling pathway by downregulating the expression levels of their
target gene, SOCS3. miR-221 and miR-222 were also found to be
expressed at high levels in DTC, which suggested that miR-221 and
miR-222 may be involved in the regulation of thyroid carcinoma
resistance to 131I treatment. However, the role and
corresponding underlying mechanism of miR-221 and miR-222 during
the 131I treatment of DTC remain unclear. Therefore, the
present study aimed to investigate the role and potential mechanism
underlying the upregulated expression levels of miR-221-3p and
miR-222-3p in the resistance of thyroid cancer to 131I therapy.
Materials and methods
Cell lines and culture
Nthy-ori-3, K1 and BCPAP cell lines were purchased
from the Cell Resource Center, Shanghai Institutes for Biological
Sciences of the Chinese Academy of Sciences. FTC133 and TPC1 cells
were purchased from Nanjing Branch Bai Biotechnology Co., Ltd. The
human thyroid cancer cell lines FTC133, TPC1 and K1 were cultured
at 37˚C in DMEM (Gibco; Thermo Fisher Scientific, Inc.)
supplemented with 10% FBS (Gibco; Thermo Fisher, Shanghai, China),
in a humidified incubator with 5% CO2 at 37˚C. The
Nthy-ori 3-1 human thyroid follicular epithelial cells and the
BCPAP human thyroid cancer cell line were cultured in RPMI-1640
medium (Gibco; Thermo Fisher Scientific, Inc.) supplemented with
10% FBS (Gibco; Thermo Fisher Scientific, Inc.), 1% non-essential
amino acids (Gibco; Thermo Fisher Scientific, Inc.), 1% sodium
pyruvate (100 mM) solution (Gibco; Thermo Fisher Scientific, Inc.),
100 U/ml penicillin and 100 g/ml streptomycin. The BCPAP and K1
cell lines were authenticated by comparing DNA short tandem repeats
data with the ATCC, DSMZ, JCRB RIKEN and EXPASY databases, and no
multiple alleles were found in these cell lines. The BCPAP cell
line was authenticated by Applied Biological Materials, Inc. The K1
cell line was authenticated by Jianlian Gene Technology (Beijing)
Co., Ltd. In experiments analyzing the expression levels of NIS,
FTC133, TPC1 and BCPAP cells were supplemented with 10 mU/ml
thyroid-stimulating hormone, bovine pituitary (cat. no. 609385-5;
Merck KGaA).
Cell transfection
BCPAP cells were transfected with 100 nM scrambled
oligonucleotide [negative control (NC)] or 50 nM miR-221-3p
inhibitor and 50 nM miR-222-3p inhibitor (Invitrogen; Thermo Fisher
Scientific, Inc.) to construct transient knockdown miR-221-3p and
miR-222-3p cell lines. The sequences were as follows: miR-221-3p
inhibitor, 5'-GAA ACCCAGCAGACAAUGUAGCU-3' (lot no. AS0299J7);
miR-222-3p inhibitor, 5'-ACCCAGUAGCCAGAUGUA GCU-3' (lot no.
AS0299J8); and NC, 5'-CAGUACUUUUGU GUAGUACAA-3', lot no. AS026V49).
Cell transfection was performed using Lipofectamine®
3000 (Invitrogen; Thermo Fisher Scientific, Inc.) in 6-well plates.
A total of 8 µl Lipofectamine® 3000 and 100 nM inhibitor
or NC were mixed in 2 ml RPMI-1640 medium and placed at room
temperature for 20 min. The cells were cultured at 37˚C with 5%
CO2 in the mixture for 6 h. Then the cells were cultured
in RPMI-1640 medium supplemented with 10% FBS. Total RNA was
extracted at 24 h or 48 h and protein was extracted at 48 h after
transfection with inhibitors or NC.
Patient studies
A total of 15 tissue samples from patients with PTC
were obtained retrospectively under strict anonymity from the
tissue bank of Zhejiang Cancer Hospital (Hangzhou, China). The
patients, including 8 women and 7 men, with a median age of 43
years (range, 27-72 years), were treated for PTC in our hospital
from January 2018 to December 2018. Postoperative samples with
other pathological types, patients with a history of other tumors
or aged <18 years were excluded. All samples were fresh-frozen
in liquid nitrogen following surgery and stored at -80˚C. Frozen
tissue samples were homogenized using TissueRuptor II (Qiagen,
Inc.) prior to RNA extraction. The experimental protocols were
approved by the Ethics Committee of Zhejiang Cancer Hospital
(approval no. IRB-2020-337). Informed consent forms were signed by
all the patients prior to surgery.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) analysis
Total RNA was extracted from tissues and cell lines
using TRIzol® reagent (Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturer's protocol. Total
RNA concentration was determined using a NanoDrop spectrophotometer
(NanoDrop Technologies; Thermo Fisher Scientific, Inc.). For miRNA
expression, total RNA (1 µg) was reverse-transcribed into cDNA at
42˚C for 60 min and 70˚C for 10 min and qPCR was subsequently
performed using a Bulge-Loop miRNA RT-qPCR Starter kit (Guangzhou
RiboBio Co., Ltd.) on an ABI StepOnePlus quantitative PCR
instrument (Applied Biosystems; Thermo Fisher Scientific, Inc.)
with 2 µl cDNA as the template. Bulge-loop miRNA RT-qPCR Primer
sets (one RT primer and a pair of qPCR primers for each gene)
specific for miR-221-3p, miR-222-3p and U6 were designed by
Guangzhou RiboBio Co., Ltd. Due to the patented technology, the
company refused to disclose the sequence of primers. The following
thermocycling conditions were used for the qPCR: Initial
denaturation at 95˚C for 10 min; followed by 40 amplification
cycles at 95˚C for 10 sec, 60˚C for 20 sec and 70˚C for 20 sec, at
a ramp-rate of 1.6˚C/sec.
For mRNA expression, total RNA was
reverse-transcribed into cDNA using a M-MLV Reverse Transcriptase
for qPCR (Promega Corporation). qPCR was subsequently performed
using PowerUp™ SYBR™-Green Master Mix (Thermo Fisher Scientific,
Inc.) on an ABI StepOnePlus quantitative PCR instrument. The primer
sequences for SOCS3 and β-actin (Shanghai GenePharma Co., Ltd.)
were as follows: SOCS3 forward, 5'-TCGCCACCTACTGAACCCT-3' and
reverse, 5'-GGTCCAGGAACTCCCGAAT-3'; and β-actin forward,
5'-AGCGGGAAATCGTGCGTG-3' and reverse, 5'-GGTCCA GGAACTCCCGAAT-3'.
The following thermocycling conditions were used for the qPCR:
Initial denaturation at 95˚C for 5 min, followed by 40
amplification cycles at 95˚C for 5 sec and 60˚C for 31 sec, with a
ramp-rate of 1.6˚C/sec. The mRNA and miRNA expression levels were
calculated using the 2-ΔΔCq method (17). miRNA expression levels were
normalized to U6 snRNA and mRNA expression levels were normalized
to GAPDH.
Lentivirus infection
A total of 1x106 FTC133 or TPC1 cells
were seeded into 6-well plates using DMEM medium supplemented with
10% FBS overnight. FTC133 or TPC1 cells were then infected with
lentivirus vectors (MOI=1.0). Green fluorescent protein
(GFP)-miR-221-3p (HmiR0369), GFP-miR-222-3p (HmiR0370) and GFP
lentiviral vectors (pEZX-MR03, CmiR0001) (all from GeneCopoeia,
Inc.). The cells were re-infected with the lentivirus vectors
(MOI=1.0) 72 h after the first infection. The pEZX-MR03-GFP vector
was used as the NC. GFP expression was visualized under an inverted
fluorescence microscope (Olympus Corporation) at a magnification of
x200. The subsequent experiments were performed 72 h after the last
infection. During the subsequent experiments, cell lines were
cultured and passaged for no more than 3 generations.
Western blotting
Total protein was extracted from cells using RIPA
lysis buffer [50 mM Tris (pH 7.4), 150 mM NaCl, 1% Triton X-100, 1%
sodium deoxycholate and 0.1% SDS] supplemented with a protease
inhibitor cocktail (Sigma-Aldrich; Merck KGaA). Total protein was
quantified using the bicinchoninic acid (BCA) protein assay kit
(Beyotime Institute of Biotechnology) and 30 µg protein/lane was
separated via 10% SDS-PAGE and transferred onto nitrocellulose
membranes. The membranes were subsequently blocked with 5% non-fat
dry milk for 1 h at room temperature and incubated with the
following primary antibodies at 4˚C overnight: Anti-SOCS3 (1:500;
rabbit; cat. no. ab16030; Abcam), anti-phosphorylated (p)-STAT3
(1:1,000; rabbit; cat. no. ab76315; Abcam), anti-vimentin (1:1,000;
rabbit; cat. no. ab8069; Abcam), anti-E-cadherin (1:500; rabbit;
cat. no. ab133597; Abcam), anti-NIS (1:500; rabbit; cat. no.
ab83816; Abcam) and anti-GAPDH (1:2,000; mouse; cat. no. sc365062;
Santa Cruz Biotechnology, Inc.). Following primary antibody
incubation, the membranes were washed and incubated with a goat
anti-mouse (1:2,000; cat. no. sc2005; Santa Cruz Biotechnology,
Inc.) or anti-rabbit (1:2,000; cat. no. sc2004; Santa Cruz
Biotechnology, Inc.) antibody for 1 h at room temperature. Protein
bands were visualized using ECL (cat. no. sc-2048; Santa Cruz
Biotechnology, Inc.). Densitometric analysis was performed using
ImageJ software, version 7.0 (National Institutes of Health). The
relative protein expression levels were expressed as a ratio of the
gray value of the target protein band: GAPDH protein band.
Bioinformatics analysis
miRNA sequences and annotation were acquired form
the miRBase database (18). miRNA
target prediction software, TargetScanHuman v.7.1 was used to
identify miR-221-3p and miR-222-3p (MIMAT0000278 and MIMAT0000279)
putative targets (19). Microarray
data were extracted from the Oncomine database (20). Analysis of differential gene
expression was performed using the R statistical computing package
(http://www.r-project.org). All datasets were
logarithmically transformed and centered on the median per array,
and the standard deviations were normalized to one for each array
(20).
Colony formation assay
Cells were seeded into six-well plates at different
densities (400, 1,000, 2,000, 3,000 or 4,000 cells/well) and
incubated for 24 h. Cells were subsequently treated with different
X-ray doses (Siemens AG; 200 cGy/min). Following 12 days of
incubation, the cells were fixed with 10% formaldehyde at room
temperature for 10 min, stained with 1.0% crystal violet solution
at room temperature for 10 min and then the number of colonies
(>50 cells) was counted under an inverted microscope
(magnification, x200). Surviving fractions were estimated using the
following equation: (Colony number)/(cell number seeded x plating
efficiency) x 100% (21). Each
experiment was independently repeated in triplicate.
Statistical analysis
Statistical analysis was performed using SPSS
software, version 22.0 (IBM Corp.). The significance of the
differences between groups were determined by paired or unpaired
Student's t-test (for comparison between two groups) or one way
ANOVA followed by Bonferroni's post hoc test. Histograms were used
to check for the distribution of the data. The data in this study
exhibited normal distribution. The experiments were performed in
triplicate. Data are presented as the mean ± SD. P<0.05 was
considered to indicate a statistically significant difference.
Results
SOCS3 is a potential target gene of
miR-221-3p and miR-222-3p in thyroid cancer
miR-221-3p and miR-222-3p target genes were
predicted using TargetScan (Fig.
1A). Microarray data was extracted from the Oncomine database.
Genes expressed at low levels in thyroid cancer were screened as
potential target genes. The mRNA expression levels of SOCS3 were
downregulated in different subtypes of DTC, including PTC,
follicular variant PTC (FV-PTC), tall-cell variant PTC (TCV-PTC)
and FTC (*P<0.05, ***P<0.001; Fig. 1B-E). The expression levels of the
STAT3 inhibitor, SOCS3, were of particular interest, as STAT3 not
only promotes EMT, but also participates in the regulation of
iodine uptake (16). Thus, RT-qPCR
was used to analyze the differences in SOCS3 gene expression
between 15 thyroid cancer and 5 normal thyroid tissues; the results
further confirmed that the expression levels of SOCS3 were
significantly downregulated in thyroid cancer specimens
(**P<0.01; Fig.
1F).
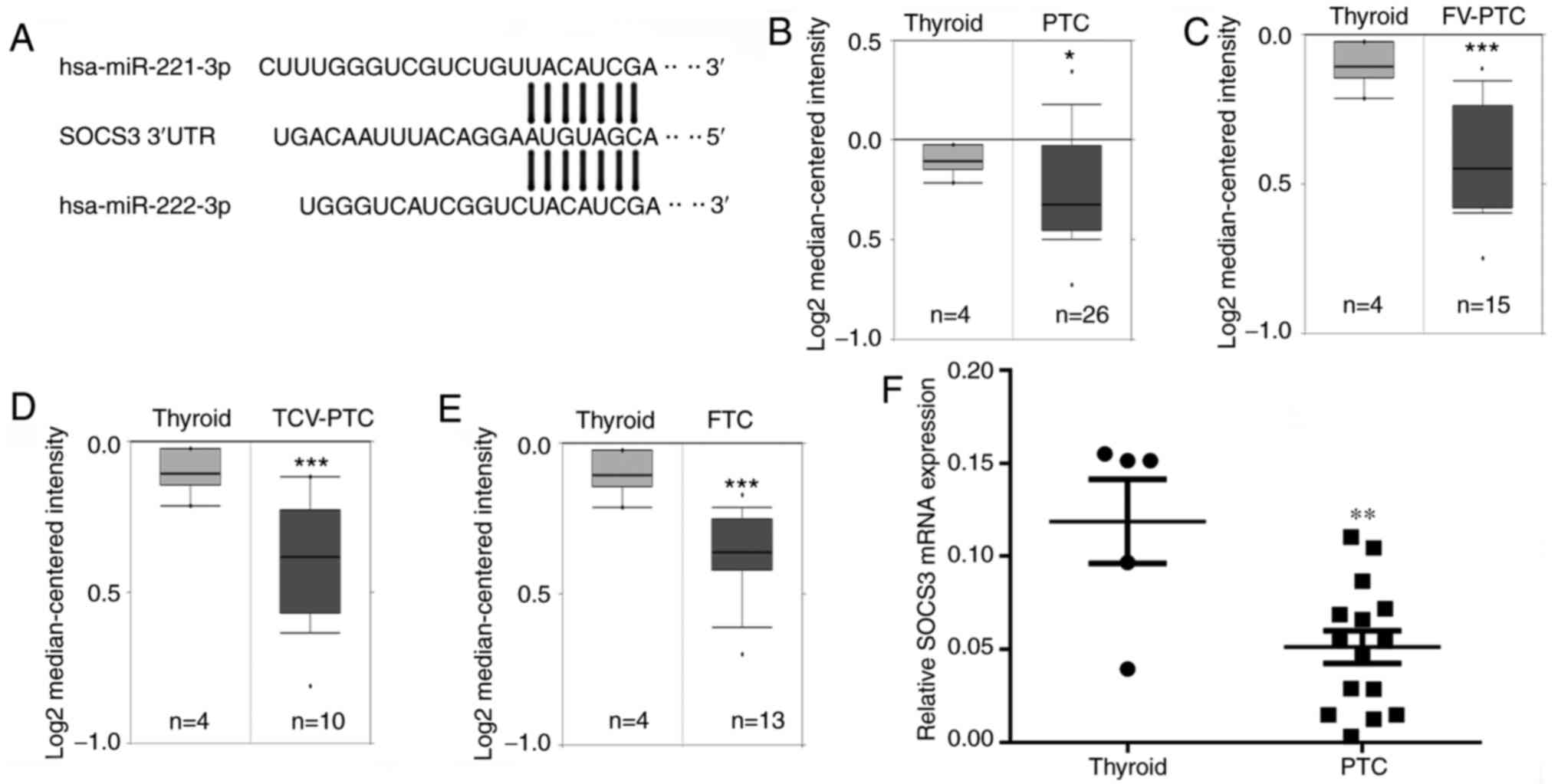 | Figure 1SOCS3 is a potential target gene of
miR-221-3p and miR-222-3p. (A) Complementary sequence of miR-221-3p
and miR-222-3p to the seed region in the SOCS3 3'- UTR. mRNA
expression levels of SOCS3 in (B) PTC, (C) FV-PTC, (D) TCV-PTC and
(E) FTC tissues obtained from the Oncomine database. (F) Reverse
transcription-quantitative polymerase chain reaction was used to
analyze the differences in SOCS3 mRNA expression levels between
thyroid cancer and normal thyroid tissues. The results are
presented as the mean ± SD of three independent experiments. The
relative mRNA levels were normalized to β-actin.
*P<0.05, **P<0.01,
***P<0.001 vs. normal thyroid tissue. SOCS3,
suppressor of cytokine signaling 3; miR, microRNA; PTC, papillary
thyroid cancer; FV, follicular variant; TCV, tall-cell variant;
FTC, follicular thyroid cancer; UTR, untranslated region. |
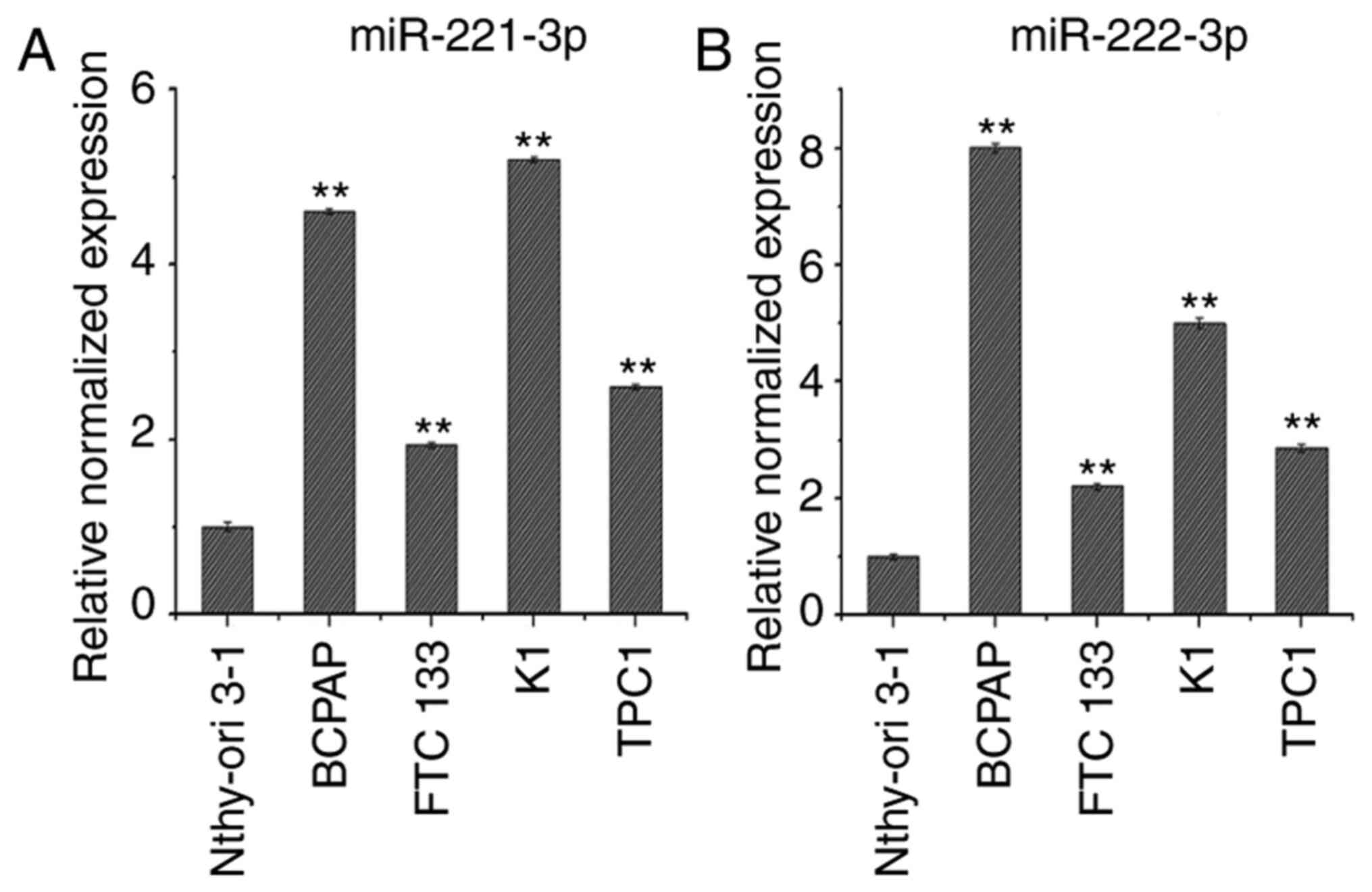
Expression levels of miR-221-3p and
miR-222-3p in thyroid cancer cell lines
RT-qPCR analysis was used to determine miR-221-3p
and miR-222-3p expression levels in the thyroid cancer cell lines
BCPAP, FTC133, K1 and TPC1, compared with Nthy-ori 3-1 cells. As
shown in Fig. 2A and B, the expression levels of miR-221-3p and
miR-222-3p were upregulated in BCPAP, FTC133, K1 and TPC1 cells
compared with Nthy-ori3-1 cells. Of note, FTC133 and TPC1 cells
exhibited lower expression levels of miR-221-3p and miR-222-3p
compared with K1 and BCPAP cells; therefore, the BCPAP cell line
was selected to establish knockdown cell models and FTC133 and TPC1
cell lines were selected to establish overexpression cell
models.
SOCS3 expression levels in thyroid
cancer cells are downregulated by miR-221-3p and miR-222-3p
BCPAP cells were transfected with a miR-221-3p
inhibitor, miR-222-3p inhibitor or NC, and total RNA was extracted
at 24 and 48 h post-transfection. RT-qPCR analysis revealed that
the co-transfection with the miR-221-3p and miR-222-3p inhibitors
could effectively downregulate the expression levels of miR-221-3p
and miR-222-3p compared with the NC group at 24 and 48 h
post-transfection in BCPAP cells (Fig.
3A). In addition, compared with the NC group, co-transfection
of BCPAP cells with miR-221-3p and miR-222-3p inhibitors
upregulated the protein expression levels of SOCS3 (Fig. 3B).
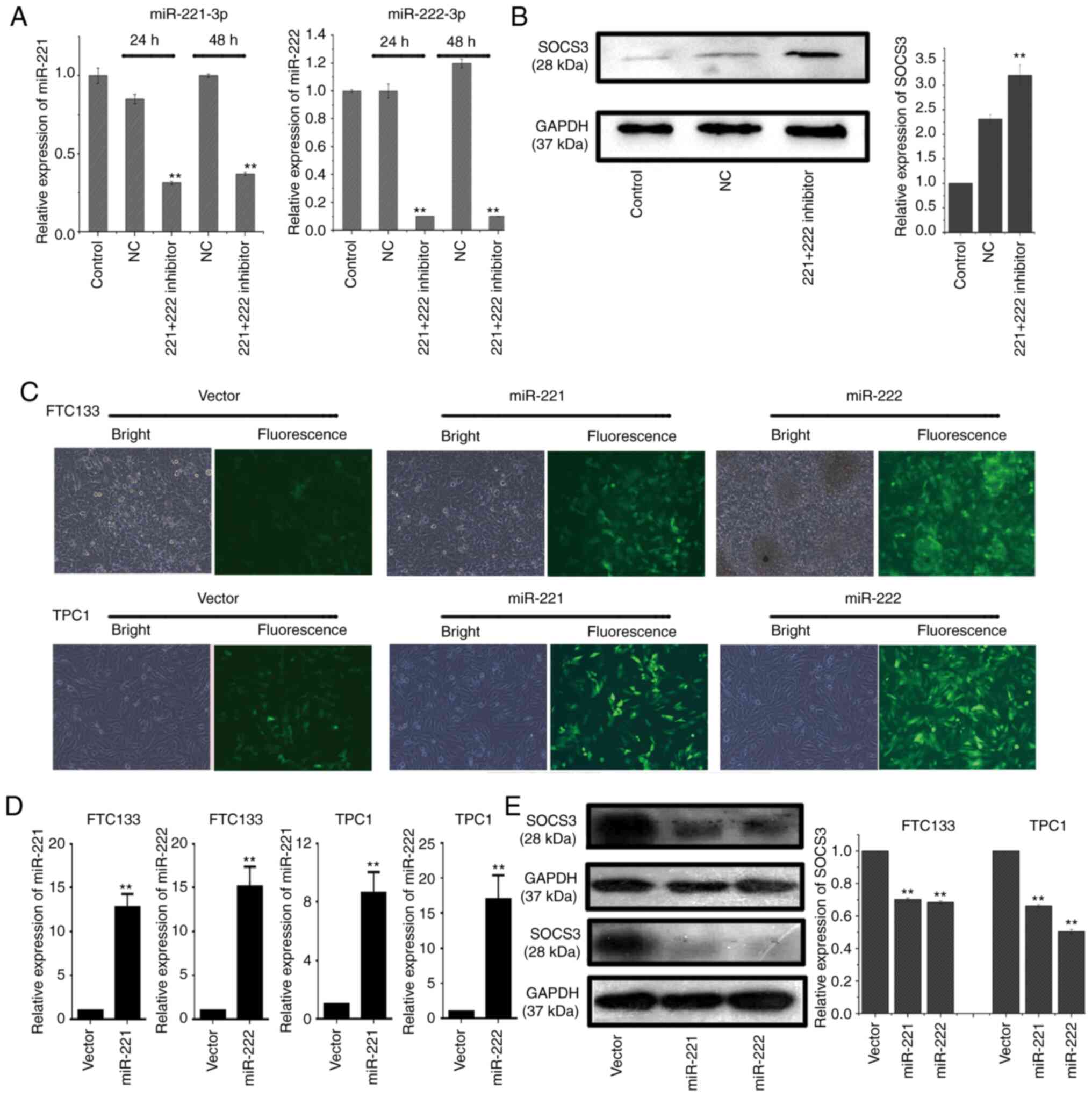 | Figure 3Downregulation of SOCS3 expression by
miR-221-3p and miR-222-3p in thyroid cancer cell lines. (A) RT-qPCR
was used to analyze endogenous miR-221-3p and miR-222-3p expression
levels in BCPAP cells following co-transfection with miR-221-3p and
miR-222-3p inhibitors for 24 or 48 h. Each reaction was performed
in triplicate. **P<0.01 vs. NC. (B) BCPAP cells were
co-transfected with a miR-221-3p and miR-222-3p inhibitor and
western blotting was used to analyze the expression levels of
SOCS3. Relative expression levels of SOCS3/GAPDH were analyzed by
ImageJ software. Each band was measured in triplicate.
**P<0.01 vs. control and NC. (C) GFP, GFP-miR-221-3p
or GFP-miR-222-3p lentiviral vectors were infected into FTC133 and
TPC1 cells and visualized using a fluorescence microscope.
(magnification, x200). (D) RT-qPCR was used to analyze the
endogenous expression levels of miR-221-3p or miR-222-3p in FTC133
and TPC1 cells following transfection with lentiviral vectors. Each
reaction was performed in triplicate. GFP lentiviral vectors served
as the NC. **P<0.01 vs. vector. (E) FTC133 (upper two
bands) and TPC1 (lower two bands) cells were infected with
GFP-miR-221-3p, GFP-miR-222-3p or GFP lentiviral vectors, and
western blotting was used to analyze the expression levels of
SOCS3. Grouping of FTC133 and TPC1 images from different parts of
the same gel. Relative expression levels of SOCS3/GAPDH were
analyzed using ImageJ software. Each band was measured in
triplicate. **P<0.01 vs. vector. SOCS3, suppressor of
cytokine signaling 3; miR, microRNA; RT-qPCR, reverse
transcription-quantitative polymerase chain reaction; GFP, green
fluorescent protein; NC, negative control; vector, GFP lentiviral
vector. |
FTC133 and TPC1 cells were subsequently infected
with lentiviral vectors to construct cell models overexpressing
miR-221-3p and miR-222-3p. The fluorescence of the lentiviral
vector was observed under a fluorescence microscope at 72 h
post-transfection, and the results demonstrated that the lentiviral
vectors were successfully infected into the cells (Fig. 3C). RT-qPCR analysis also revealed
that the infection with miR-221-3p and miR-222-3p lentiviral
vectors markedly upregulated the expression levels of miR-221-3p
and miR-222-3p at 72 h post-infection (Fig. 3D). The western blotting results also
demonstrated that the protein expression levels of SOCS3 were
significantly downregulated following overexpression of miR-221-3p
or miR-222-3p (Fig. 3E).
miR-221-3p and miR-222-3p regulate the
expression levels of p-STAT3, EMT-related markers and NIS in
thyroid cancer cells
The western blotting results revealed that,
following inhibition of miR-221-3p and miR-222-3p expression in
BCPAP cells, the expression levels of p-STAT3 and vimentin were
downregulated, while the expression levels of E-cadherin and NIS
were upregulated (Fig. 4A and
B). Conversely, following
overexpression of miR-221-3p or miR-222-3p in FTC133 and TPC1
cells, the expression levels of p-STAT3 and vimentin were
upregulated, while the expression levels of E-cadherin and NIS were
significantly downregulated (Fig.
4A, C and D).
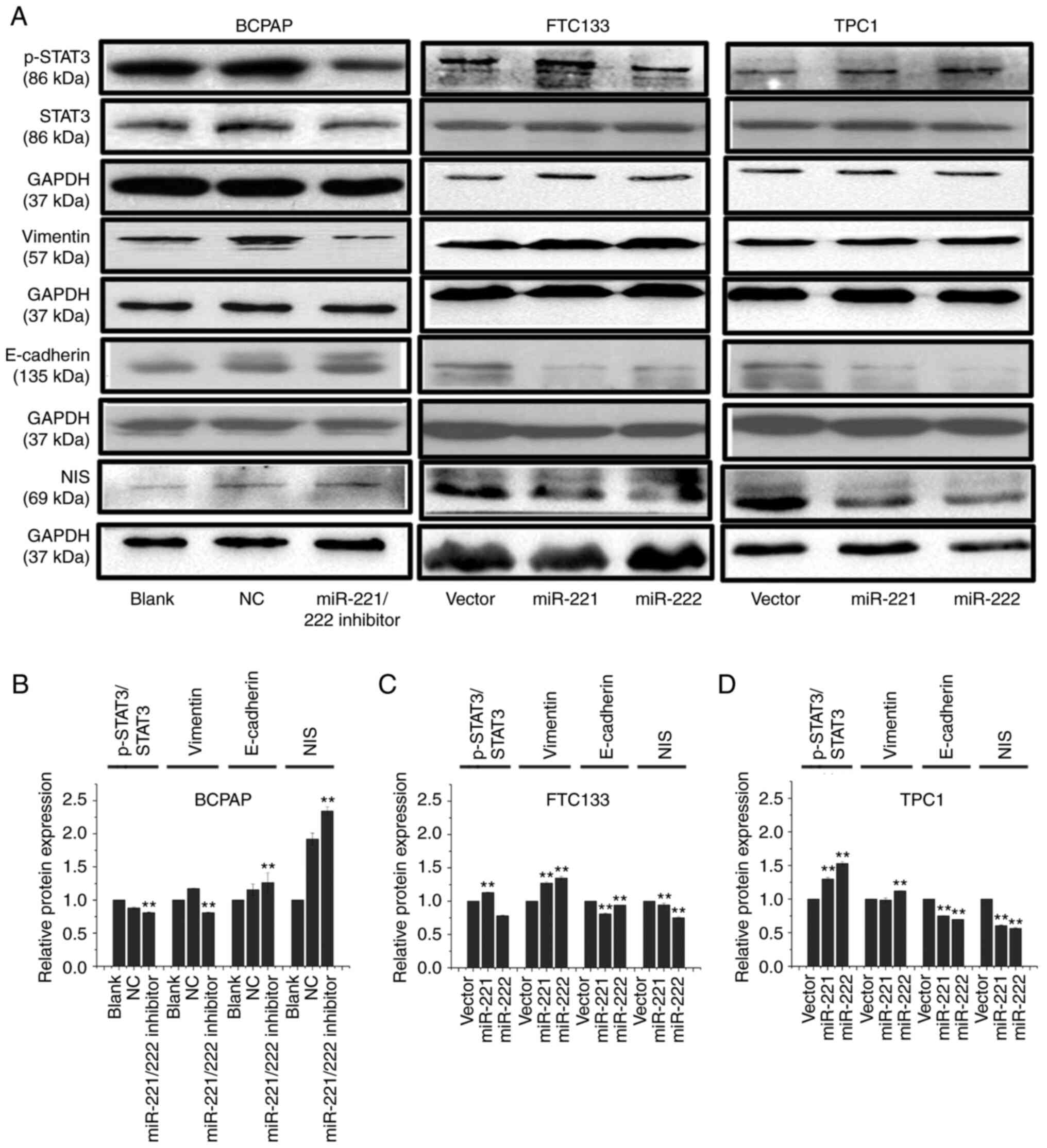 | Figure 4miR-221-3p and miR-222-3p upregulate
the protein expression levels of p-STAT3 and vimentin and
downregulate the expression of NIS and E-cadherin. (A) BCPAP cells
were transfected with miR-221-3p, miR-222-3p inhibitor and NC.
FTC133 and TPC1 cells were infected with GFP-miR-221-3p,
GFP-miR-222-3p or GFP lentiviral vectors. Western blotting was used
to analyze the expression levels of p-STAT3, STAT3, vimentin,
E-cadherin and NIS. Grouping of BCPAP images from different gels
than those of FTC133 and TPC1. Grouping of FTC133 and TPC1 images
from different parts of the same gel. Bands of p-STAT3 and STAT3
from different gels. (B-D) Relative expression levels of
p-STAT3/STAT3, vimentin/GAPDH, E-cadherin/GAPDH and NIS/GAPDH were
analyzed using ImageJ 7.0 software in each cell model. Each band
was measured in triplicate. **P<0.01 vs. NC or
vector. miR, microRNA; p-, phosphorylated; STAT, signal transducer
and activator of transcription; NIS, solute carrier family 5 member
5; GFP, green fluorescent protein; NC, negative control; vector,
GFP lentiviral vector. |
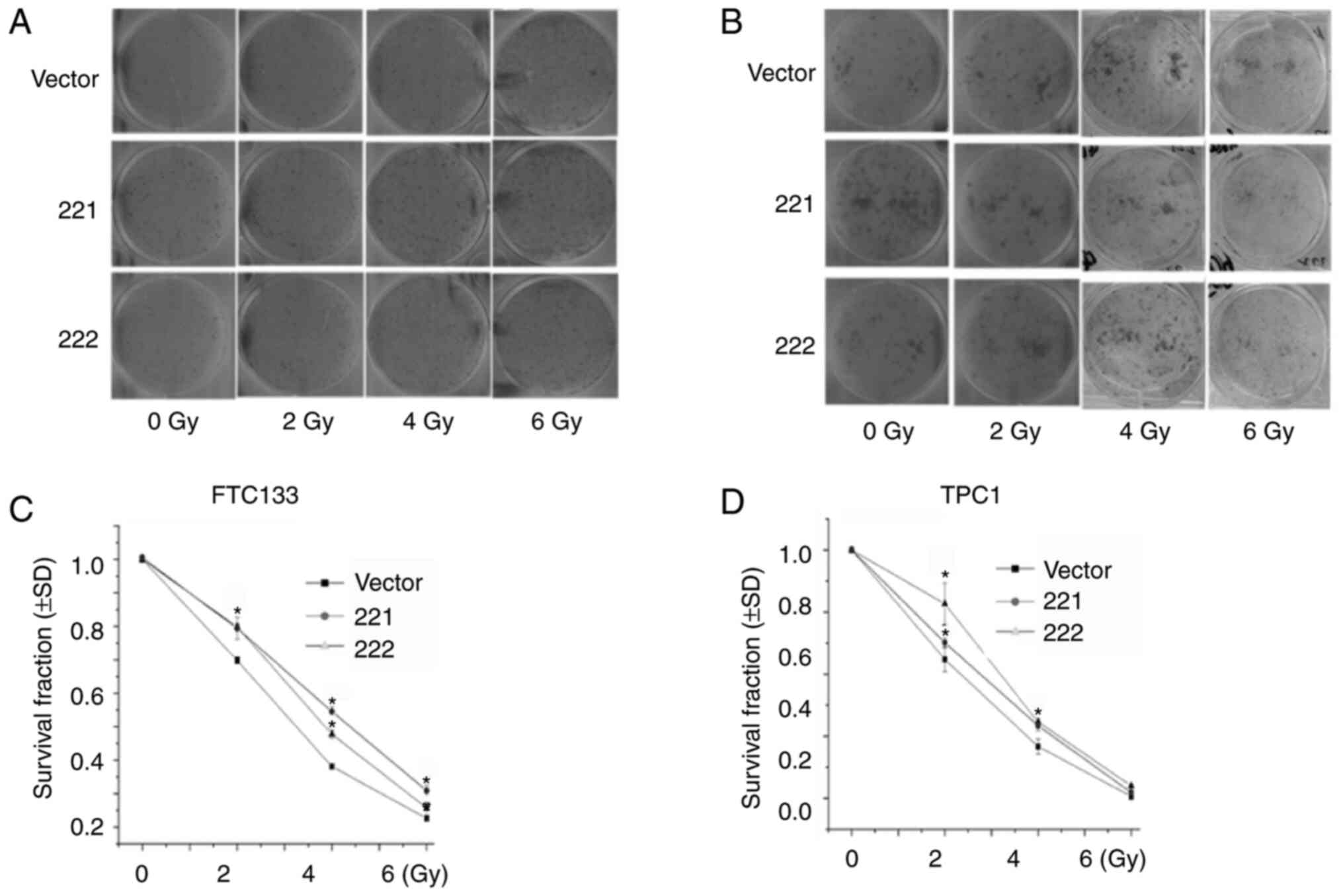
miR-221-3p and miR-222-3p reduce
radiosensitivity in thyroid cancer cells
It was previously reported that EMT was associated
with decreased cancer sensitivity to radiation therapy (22-25).
Therefore, a colony formation assay was used to determine the role
of miR-221-3p and miR-222-3p in radiosensitivity. FTC133 (Fig. 5A and C) and TPC1 (Fig. 5B and D) cells were infected with lentivirus
vectors, treated with different doses of X-ray irradiation and
cultured for 12 days. The results demonstrated that the
overexpression of miR-221-3p or miR-222-3p significantly increased
the surviving fraction of FTC133 (at 2, 4 and 6 Gy; Fig. 5C) and TPC1 (at 2 and 4 Gy; Fig. 5D) cells. These results indicated
that overexpression of miR-221-3p or miR-222-3p may increase
radioresistance in DTC cell lines.
Discussion
131I is an important method
used for the treatment of distant metastatic DTC. However, ~50% of
distant metastatic DTCs are resistant to 131I therapy,
which is currently the main cause of DTC-related mortality
(2,4,26).
Reversing the 131I resistance of distant metastatic
lesions to 131I therapy has been associated with the
upregulated expression of NIS, which increases radiosensitivity and
may represent a valuable research focal point for the clinical
treatment of DTC (26). miRNAs are
a class of short-stranded RNAs that play important roles in
regulating the biological functions of tumor cells, including the
response to cancer therapy. However, research into the role of
miRNAs in the resistance of thyroid cancer to 131I
remains limited.
The expression levels of miR-221 and miR-222 were
previously found to be upregulated in DTC (27,28).
The RT-qPCR results obtained in the present study also revealed
that the expression levels of miR-221-3p and miR-222-3p were
upregulated in thyroid cancer cell lines compared with those in
normal thyroid cells. Few previous studies have investigated the
mechanism of miR-221-3p and miR-222-3p in the resistance to
131I therapy. In the present study, through
bioinformatics analysis prediction, SOCS3 was identified as a
potential target gene of miR-221-3p and miR-222-3p. Analysis of
thyroid cancer tissues from the Oncomine database revealed that the
expression levels of SOCS3 were downregulated in thyroid cancer.
Several previous studies have also confirmed that the
miR-221/miR-222 cluster can bind to the 3'-UTR of SOCS3 and
negatively regulate the translation of SOCS3 (9,29-31).
The results of the present study demonstrated that the
overexpression of miR-221-3p or miR-222-3p inhibited the expression
of SOCS3 in thyroid cancer cells, and inhibiting the expression of
miR-221-3p and miR-222-3p upregulated the expression levels of
SOCS3. Whether miR-221-3p and miR-222-3p directly target 3'-UTR of
SOCS3 and suppress the expression of SOCS3 was not been further
verified in the present study. SOCS3 has been previously observed
to negatively regulate the JAK/STAT signaling pathway (9,29,32).
The JAK/STAT signaling pathway participates in the regulation of
EMT in tumor cells and was found to inhibit 131I uptake
in thyroid cancer (16,33). Furthermore, the upregulated
expression of SOCS3 was shown to inhibit metastasis by
downregulating p-STAT3 and vimentin expression and upregulating
E-cadherin expression (32,34).
In the present study, the expression levels of
miR-221-3p and miR-222-3p were found to be higher in the BCPAP cell
line compared with those in the FTC and TPC1 cell lines. Therefore,
BCPAP cells were transfected with miR-221-3p and miR-222-3p
inhibitors or NC to establish miR-221-3p and miR-222-3p knockdown
cell models. The results revealed that the simultaneous inhibition
of the expression of miR-221-3p and miR-222-3p downregulated the
expression levels of p-STAT3 and vimentin, and upregulated
E-cadherin expression levels. In addition, the simultaneous
inhibition of miR-221-3p and miR-222-3p restored the expression of
NIS, which is a key gene required for the uptake of 131I
by thyroid cancer cells (26). In
addition, FTC133 and TPC1 cells were transfected with lentiviral
vectors to establish miR-221-3p and miR-222-3p overexpression cell
models. Following overexpression of miR-221-3p or miR-222-3p, the
expression levels of p-STAT3 and vimentin were upregulated, while
the expression levels of E-cadherin and NIS were significantly
downregulated. These findings further suggested that the
upregulated expression of miR-221-3p and miR-222-3p may be
associated with the mechanisms underlying iodine metabolism
dysfunction in thyroid cancer cells. In the miR-221-3p and
miR-222-3p overexpression cell models, the radiosensitivity of the
cells was verified using a colony formation assay. The results
demonstrated that the overexpression of miR-221-3p or miR-222-3p in
FTC133 and TPC1 cells could significantly reduce their
radiosensitivity.
A limitation of the present study is that rescue
assay or dual-luciferase reporter gene assay would be necessary to
further verify the role of the miR-221/222-3p-SOCS3 axis.
Furthermore, although overexpression of miR-221/222-3p decreased
NIS expression, this result must be verified by iodine uptake
test.
In conclusion, the results of the present study
indicated that the overexpression of miR-221-3p and miR-222-3p in
thyroid cancer cells may downregulate NIS expression levels and
thereby reduce radiosensitivity, which indicates that the
overexpression of miR-221-3p and miR-222-3p in DTC may
simultaneously inhibit iodine metabolism and reduce
radiosensitivity. The potential underlying mechanism was
hypothesized to be associated with the miR-221-3p- and
miR-222-3p-targeted inhibition of SOCS3, which may subsequently
lead to the activation of the STAT3 signaling pathway in DTC
cells.
Acknowledgements
Not applicable.
Funding
Funding: The present study was supported by the National Natural
Science Foundation of China (grant no. 81502318) and the Zhejiang
Province Natural Science Foundation of China (grant no.
LY20H180007).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
HY conceived and designed the study, contributed to
the experiments and drafted the manuscript. TY and LZ performed all
the experiments. XY and JL performed the data analysis. LL
contributed to the conception and designed the study. TY and HY
have seen and can confirm the authenticity of all the raw data. All
the authors have read and approved the final manuscript.
Ethics approval and consent to
participate
The experimental protocols were approved by the
Ethics Committee of Zhejiang Cancer Hospital (approval no.
IRB-2020-337). All patients provided consent for the publication of
their data.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018.PubMed/NCBI View Article : Google Scholar : Erratum in: CA
Cancer J Clin 70, 313, 2020.
|
|
2
|
Song HJ, Qiu ZL, Shen CT, Wei WJ and Luo
QY: Pulmonary metastases in differentiated thyroid cancer: Efficacy
of radioiodine therapy and prognostic factors. Eur J Endocrinol.
173:399–408. 2015.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Agrawal N, Akbani R, Aksoy BA, Ally A,
Arachchi H, Asa SL, Auman JT, Balasundaram M, Balu S, Baylin SB, et
al: Cancer Genome Atlas Research Network: Integrated genomic
characterization of papillary thyroid carcinoma. Cell. 159:676–690.
2014.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Van Nostrand D: Radioiodine refractory
differentiated thyroid cancer: Time to update the classifications.
Thyroid. 28:1083–1093. 2018.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Treiber T, Treiber N and Meister G:
Regulation of microRNA biogenesis and its crosstalk with other
cellular pathways. Nat Rev Mol Cell Biol. 20:5–20. 2019.PubMed/NCBI View Article : Google Scholar : Erratum in: Nat Rev
Mol Cell Biol 19, 808, 2018.
|
|
6
|
Tsikrika FD, Avgeris M, Levis PK, Tokas T,
Stravodimos K and Scorilas A: miR-221/222 cluster expression
improves clinical stratification of non-muscle invasive bladder
cancer (TaT1) patients' risk for short-term relapse and
progression. Genes Chromosomes Cancer. 57:150–161. 2018.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Chen WX, Hu Q, Qiu MT, Zhong SL, Xu JJ,
Tang JH and Zhao JH: miR-221/222: Promising biomarkers for breast
cancer. Tumour Biol. 34:1361–1370. 2013.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Amini S, Abak A, Sakhinia E and Abhari A:
MicroRNA-221 and microRNA-222 in common human cancers: Expression,
function, and triggering of tumor progression as a key modulator.
Lab Med. 50:333–347. 2019.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Xu CH, Liu Y, Xiao LM, Chen LK, Zheng SY,
Zeng EM, Li DH and Li YP: Silencing microRNA-221/222 cluster
suppresses glioblastoma angiogenesis by suppressor of cytokine
signaling-3-dependent JAK/STAT pathway. J Cell Physiol.
234:22272–22284. 2019.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Jikuzono T, Kawamoto M, Yoshitake H,
Kikuchi K, Akasu H, Ishikawa H, Hirokawa M, Miyauchi A, Tsuchiya S,
Shimizu K, et al: The miR-221/222 cluster, miR-10b and miR-92a are
highly upregulated in metastatic minimally invasive follicular
thyroid carcinoma. Int J Oncol. 42:1858–1868. 2013.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Zhang X, Mao H and Lv Z: MicroRNA role in
thyroid cancer pathogenesis. Front Biosci. 18:734–739.
2013.PubMed/NCBI View
Article : Google Scholar
|
|
12
|
Braun J, Hoang-Vu C, Dralle H and
Hüttelmaier S: Downregulation of microRNAs directs the EMT and
invasive potential of anaplastic thyroid carcinomas. Oncogene.
29:4237–4244. 2010.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Yu S, Liu Y, Wang J, Guo Z, Zhang Q, Yu F,
Zhang Y, Huang K, Li Y, Song E, et al: Circulating microRNA
profiles as potential biomarkers for diagnosis of papillary thyroid
carcinoma. J Clin Endocrinol Metab. 97:2084–2092. 2012.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Gómez-Pérez AM, Cornejo Pareja IM, García
Alemán J, Coín Aragüez L, Sebastián Ochoa A, Alcaide Torres J,
Molina Vega M, Clu Fernández C, Mancha Doblas I and Tinahones FJ:
New molecular biomarkers in differentiated thyroid carcinoma:
Impact of miR-146, miR-221 and miR-222 levels in the evolution of
the disease. Clin Endocrinol (Oxf). 91:187–194. 2019.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Ravegnini G, Cargnin S, Sammarini G,
Zanotti F, Bermezo JL, Hrelia P, Terrazzino S and Angelini S:
Prognostic role of miR-221 and miR-222 expression in cancer
patients: A systematic review and meta-analysis. Cancers (Basel).
11(970)2019.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Tseng LM, Huang PI, Chen YR, Chen YC, Chou
YC, Chen YW, Chang YL, Hsu HS, Lan YT, Chen KH, et al: Targeting
signal transducer and activator of transcription 3 pathway by
cucurbitacin I diminishes self-renewing and radiochemoresistant
abilities in thyroid cancer-derived CD133+ cells. J
Pharmacol Exp Ther. 341:410–423. 2012.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) Method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Kozomara A, Birgaoanu M and
Griffiths-Jones S: miRBase: From microRNA sequences to function.
Nucleic Acids Res. 47 (D1):D155–D162. 2019.PubMed/NCBI View Article : Google Scholar
|
|
19
|
TargetScanHuman 5.2 Custom. http://www.targetscan.org/vert_50/seedmatch.html.
Accessed April 14, 2021.
|
|
20
|
Rhodes DR, Kalyana-Sundaram S, Mahavisno
V, Varambally R, Yu J, Briggs BB, Barrette TR, Anstet MJ,
Kincead-Beal C, Kulkarni P, et al: Oncomine 3.0: Genes, pathways,
and networks in a collection of 18,000 cancer gene expression
profiles. Neoplasia. 9:166–180. 2007.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Franken NA, Rodermond HM, Stap J, Haveman
J and van Bree C: Clonogenic assay of cells in vitro. Nat Protoc.
1:2315–2319. 2006.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Wu C, Guo E, Ming J, Sun W, Nie X, Sun L,
Peng S, Luo M, Liu D, Zhang L, et al: Radiation-induced DNMT3B
promotes radioresistance in nasopharyngeal carcinoma through
methylation of p53 and p21. Mol Ther Oncolytics. 17:306–319.
2020.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Xie C, Wu Y, Fei Z, Fang Y, Xiao S and Su
H: MicroRNA-1275 induces radiosensitization in oesophageal cancer
by regulating epithelial-to-mesenchymal transition via
Wnt/β-catenin pathway. J Cell Mol Med. 24:747–759. 2020.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Zhou S, Zhang M, Zhou C, Wang W, Yang H
and Ye W: The role of epithelial-mesenchymal transition in
regulating radioresistance. Crit Rev Oncol Hematol.
150(102961)2020.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Gomez-Casal R, Bhattacharya C, Ganesh N,
Bailey L, Basse P, Gibson M, Epperly M and Levina V: Non-small cell
lung cancer cells survived ionizing radiation treatment display
cancer stem cell and epithelial-mesenchymal transition phenotypes.
Mol Cancer. 12(94)2013.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Zhao Y, Zhong L and Yi H: A review on the
mechanism of iodide metabolic dysfunction in differentiated thyroid
cancer. Mol Cell Endocrinol. 479:71–77. 2019.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Marini F, Luzi E and Brandi ML: MicroRNA
role in thyroid cancer development. J Thyroid Res.
2011(407123)2011.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Li S, Li Q, Lü J, Zhao Q, Li D, Shen L,
Wang Z, Liu J, Xie D, Cho WC, et al: Targeted inhibition of
miR-221/222 promotes cell sensitivity to cisplatin in
triple-negative breast cancer MDA-MB-231 cells. Front Genet.
10(1278)2020.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Ying X, Wu Q, Wu X, Zhu Q and Wang X,
Jiang L, Chen X and Wang X: Epithelial ovarian cancer-secreted
exosomal miR-222-3p induces polarization of tumor-associated
macrophages. Oncotarget. 7:43076–43087. 2016.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Xie J, Wen JT, Xue XJ, Zhang KP, Wang XZ
and Cheng HH: miR-221 inhibits proliferation of pancreatic cancer
cells via down regulation of SOCS3. Eur Rev Med Pharmacol Sci.
22:1914–1921. 2018.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Zhou QY, Peng PL and Xu YH: miR-221
affects proliferation and apoptosis of gastric cancer cells through
targeting SOCS3. Eur Rev Med Pharmacol Sci. 24(7565)2020.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Wang H, Zhan M, Liu Q and Wang J:
Glycochenodeoxycholate promotes the metastasis of gallbladder
cancer cells by inducing epithelial to mesenchymal transition via
activation of SOCS3/JAK2/STAT3 signaling pathway. J Cell Physiol.
235:1615–1623. 2020.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Riesco-Eizaguirre G, Rodríguez I, De la
Vieja A, Costamagna E, Carrasco N, Nistal M and Santisteban P: The
BRAFV600E oncogene induces transforming growth factor beta
secretion leading to sodium iodide symporter repression and
increased malignancy in thyroid cancer. Cancer Res. 69:8317–8325.
2009.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Zhou QX, Jiang XM, Wang ZD, Li CL and Cui
YF: Enhanced expression of suppresser of cytokine signaling 3
inhibits the IL-6-induced epithelial-to-mesenchymal transition and
cholangiocarcinoma cell metastasis. Med Oncol.
32(105)2015.PubMed/NCBI View Article : Google Scholar
|