Introduction
Atherosclerosis is a disease in which
atherosclerotic plaques are deposited in arterial wall and cause
arterial stenosis (1).
Atherosclerosis may lead to coronary artery disease, stroke and
peripheral artery disease (2).
Globally, 7.4 million deaths are caused by coronary heart disease
each year and 6.7 million patients die from stroke in 2015(3). The exact mechanism of atherosclerosis
is still not fully understood, but endothelial cell apoptosis is
known to be one of the important mechanisms underlying
atherosclerosis progression (4).
Endothelial cell injury, which is one of the
earliest pathophysiological changes in atherosclerosis promotes the
production of inflammatory mediators, such as interleukin-1β
(5) and free radicals including
reactive oxygen species and reactive nitrogen species (6) to form an inflammatory and an oxidative
stress environment (7). Normal
endothelium serves a role in regulating vascular tone, cell
adhesion, smooth muscle cell proliferation and in maintaining
vascular homeostasis (8). A
previous have found a significant increase in endothelial cell
apoptosis in atherosclerotic blood vessels and plaques (9). Apoptosis of endothelial cells allows
leukocytes and low-density lipoprotein (LDL) to pass through the
blood vessel wall more easily and continuous accumulation of LDL in
the endothelium, which results in plaque formation and
subsequently, atherosclerosis (10). In accordance, some factors that
cause atherosclerosis, such as high levels of LDL, elevated blood
glucose levels, reduced nitric oxide levels and increased oxidative
stress levels have been associated with an increase in endothelial
cell apoptosis (11). Apoptosis of
endothelial cells results in activation of the coagulation system,
which is followed by destruction of the vascular endothelium and
even, local thrombosis that can eventually lead to vascular
occlusion, unstable angina pectoris, heart attack and stroke
(12). Based on all these findings,
a growing number of researchers have been focusing on how
endothelial cell apoptosis can be inhibited.
MicroRNA (miR) is a ribonucleic acid molecule with a
length of 21-23 nucleotides that is widely present in eukaryotes
and can regulate the expression of other genes (13). Various miRNAs have been found to
serve an important role in the pathogenesis of atherosclerosis. For
example, miR-34 has a protective effect against oxidative stress in
endothelial cells (14) and miR-155
has the ability to destroys tight junctions and the integrity of
endothelial barriers, leading to an increased endothelial
permeability and enhanced atherosclerotic progression (15). In addition, miR-345-3p,
miRNA-26a-5p, miR-142-3p were found to regulate endothelial cell
apoptosis (16-19).
Additionally, previous studies found that miR-616-3p may be
involved in the development of coronary atherosclerosis. For
example, miR-616-3p was found to participate in the development of
atherosclerosis by directly acting on the 3'UTR of paraoxonase 1
(PON-1), but the specific mechanism is still unclear (20). Another study demonstrated that
miR-616-3p single nucleotide polymorphisms at PON1 could affect
genetic expression and that this was associated with an elevated
risk for ischemic stroke and subclinical atherosclerosis (21). Hence, the precise role of miR-616-3p
in the pathogenesis of coronary atherosclerosis is not clear
(21). It may be beneficial to
explore the mechanisms of miR-616-3p in endothelial cell injury, as
it may prove to be a potential treatment target for coronary
atherosclerosis in the future.
Based on all the previously reported findings, the
present study hypothesized that miR-616-3p is involved in the
pathogenesis of coronary atherosclerosis via its effect on
endothelial cell apoptosis. In addition, the present study aimed to
explore the potential mechanisms via which miR-616-3p may play a
role in endothelial cell apoptosis. The purpose of this study was
to elucidate the mechanism of atherosclerosis and to find new
therapeutic targets for atherosclerosis. miR-616-3p and XIAP may be
used as future therapeutic targets of atherosclerosis.
Materials and methods
Cell culture and treatment
HUVECs (ATCC® PCS-100-013™)
were purchased from ATCC and cultured in RPMI-1640 medium (Thermo
Fisher Scientific Inc.) containing 10% FBS (Hyclone; GE Healthcare
Life Sciences) and 1% penicillin-streptomycin solution. The cells
were cultured at 37˚C in a humidified incubator containing 5%
CO2 and passaged when they reached 90% confluence.
Subsequently, HUVECs were planted in 6-well plates at a density of
1x106 per well and were treated with RPMI-1640 medium
(with 10% FBS and 1% penicillin-streptomycin solution) containing
60 µg/ml oxidized low-density lipoprotein (ox-LDL) for 48 h at 37˚C
in a humidified incubator containing 5% CO2 (22). Untreated HUVECS served as the
control group. RNA was extracted from the treated cells for
subsequent reverse transcription-quantitative (RT-q) PCR.
Reverse transcription-quantitative
(RT-q) PCR
According to the manufacturer's protocol, RNAiso
(Takara Bio, Inc.) was used to extract total RNA from cells, and
NanoDrop2000 was used to measure RNA concentration and purity.
PrimeScript™ RT Master Mix (Takara Bio, Inc.) was used
for reverse transcription according to the manufacturer's
instructions. The following protocol was used: 37˚C for 15 min
(reverse transcription reaction) and 85˚C for 5 sec (reverse
transcriptase inactivation reaction). The cDNA obtained by reverse
transcription was amplified on StepOne Plus (Thermo Fisher
Scientific Inc.) using the TB Green® Premix Ex
Taq™ kit (Takara Bio, Inc.) according to the
manufacturer's protocol. The thermocycling conditions used were as
follows: Initial denaturation at 95˚C for 30 sec, followed by 40
cycles of denaturation at 95˚C for 5 sec and finally annealing and
extension at 60˚C for 30 sec. The relative expression level was
calculated using the 2-ΔΔCq method (23). The following primers were used:
GAPDH forward, 5'-GGAGCCAAAAGGGTCAT-3' and reverse,
5'-GAGTCCTTCCACGATACCAA-3'; X-linked inhibitor of apoptosis protein
(XIAP) forward, 5'-GTGACTAGATGTCCACAAGG-3' and reverse,
5'-GTTGAGGAGTGTCTGGTAAG-3'; U6 forward, 5'-CTCGCTTCGGCAGCACA-3' and
reverse, 5'-AACGCTTCACGAATTTGCGT-3'; and miR-616-3p forward,
5'-ACACTCCAGCTGGGAGTCATTGGAGGGTTT-3' and reverse,
5'-TGGTGTCGTGGAGTCG-3'. GAPDH was used as the internal control for
XIAP and U6 was used as the internal control for miR-616-3p.
Cell counting kit-8
HUVECs were seeded into 96-well plates at a density
of 4,000 cells per well and the culture plate was placed in the
incubator for 24 h at 37˚C. Cell counting Kit-8 (CCK-8; Dojindo
Molecular Technologies Inc.) solution (10 µl) was added to each
well and the culture plate was placed in the incubator for 1 h at
37˚C. Absorbance was measured at 450 nm with a microplate reader
(iMark™ Microplate Absorbance Reader; Bio-Rad
Laboratories Inc.), and the measured optical density (OD) value was
used as an indicator of cell viability.
Transfection
miR-616-3p mimic, miR-616-3p mimic non-targeting
control, XIAP overexpression plasmid and non-targeting control
(empty vector) were designed and synthesized by Shanghai
Genepharma, Co. Ltd. miR-616-3p mimic (100 nM), miR-616-3p mimic
non-targeting control (100 nM), XIAP overexpression plasmid (4 µg)
and non-targeting control (4 µg) were transfected into HUVECs with
Lipofectamine 2000® (Invitrogen; Thermo Fisher
Scientific Inc.) at 37˚C for 48 h. Untransfected cells were used as
negative control (NC). RNA and total protein were extracted for
subsequent experiments 48 h after transfection. miR-616-3p mimic,
5'-AGUCAUUGGAGGGUUUGAGCAG-3'; and miR-616-3p mimic non-targeting
control, 5'-ACUACUGAGUGACAGUAGA-3'.
Flow cytometry analysis
HUVECs (1x106/well) from negative
control, miR-616-3p and miR-616-3p mimic control were collected and
washed with pre-cooled PBS and centrifuged at 725 x g for 10 min at
4˚C. The supernatant was discarded and the cells were resuspended
in 200 µl binding buffer. Subsequently, 10 µl of Annexin V-FITC and
10 µl propidium iodide (PI) was added (Dead Cell Apoptosis Kit with
Annexin V FITC and PI; Thermo Fisher Scientific Inc.) and mixed for
15 min at room temperature in the dark. Finally, 300 µl of binding
buffer was added and flow cytometry analysis was performed within 1
h on the FACSCalibur Flow Cytometry System (BD Biosciences), and
the results were analyzed using FlowJo v.8.0 software (Tree Star,
Inc.). Both early and late apoptosis were assessed.
TUNEL staining
HUVECs were seeded into 48-well plates at a density
of 104 cells per well. First, the cells were washed with
saline. Subsequently, the cells were fixed in PBS with 4% neutral
formaldehyde at room temperature for 15 min. Then, the cells were
washed once with PBS. After treatment with Enhanced Immunostaining
Permeabilization Solution (cat. no. P0097; Beyotime Institute of
Biotechnology) for 5 min at room temperature, the cells were washed
twice with PBS. Then, the slides were incubated with TUNEL reaction
mixture (One Step TUNEL Apoptosis Assay kit; cat. no. C1090;
Beyotime Institute of Biotechnology) for 60 min at 37˚C. The cells
were then immediately observed under a fluorescence microscope to
observe the red fluorescence (magnification, x400).
Western blotting
HUVECs (1x106/well) were washed once with
ice cold PBS and lysed with RIPA lysis buffer (cat. no. P0013B;
Beyotime Institute of Biotechnology) on ice for 30 min. The
homogenate was collected and centrifuged at 14,000 x g for 10 min
at 4˚C. The supernatant was the total protein and was used to
determine the protein concentration with the bicinchoninic acid
(BCA) method. Next, 40 µg of the extracted protein was added to
each well for 10% SDS-PAGE electrophoresis and the protein was
transferred to a PVDF membrane. The PVDF membrane was blocked with
5% skimmed milk containing TBS-0.1% Tween-20 (TBS-T) at room
temperature for 1 h and incubated with diluted primary antibodies
(all Abcam) against β-actin (1:1,000; cat. no. ab8227), XIAP
(1:1,000; cat. no. ab229050), cleaved caspase-3 (1:1,000; cat. no.
ab32042) and total caspase-3 (1:1,000; cat. no. ab32150) at 4˚C
overnight. The PVDF membrane was then washed with TBST and
incubated with the corresponding diluted secondary antibodies
(1:5,000; cat. no. ab205718; Abcam) at room temperature for 1 h.
BeyoECL Moon (Beyotime Institute of Biotechnology) was added to the
PVDF membrane to detect the chemiluminescence intensity. Image J
software v.2.1.4.7 (National Institutes of Health) was used to
analyze band intensity. β-actin was used as the loading
control.
Dual-luciferase reporter assay
Based on the StarBase 3.0 (http://starbase.sysu.edu.cn/) prediction, miR-616-3p
was found to have a potential binding site for the 3'UTR of XIAP
mRNA. Luciferase reporter plasmids containing the wild-type (wt) or
mutant (mut) 3'untranslated region (UTR) sequence of XIAP.
pmirGLO-XIAP-wt and pmirGLO-XIAP-mut plasmids were constructed by
Shanghai Gene Pharma Co. Ltd. HUVECs were seeded in 24-well plates
at a density of 2x105 cells/well. miR-616-3p mimic
(5'-AGUCAUUGGAGGGUUUGAGCAG-3') and miR-616-3p non-targeting control
(5'-ACUACUGAGUGACAGUAGA-3') were synthesized by Shanghai Gene
Pharma Co. Ltd. According to the manufacturer's instructions,
Lipofectamine 2000® (Invitrogen; Thermo Fisher
Scientific Inc.) was used to transfect miR-616-3p mimic, miR-616-3p
non-targeting control, pmirGLO-XIAP-wt and pmirGLO-XIAP-mut into
cells. After 48 h of transfection, the medium was removed and
fluorescence intensity was detected using the Dual Luciferase
Reporter Gene Assay kit (Beyotime Institute of Biotechnology).
Renilla luciferase activity was used as the normalization
control.
Statistical analysis
All experiments were repeated 3 times. All results
are expressed as mean ± SD. Statistical analysis was performed
using SPSS 19.0 (IBM Corp.). Paired Student's t-test was used for
comparison between 2 groups. ANOVA followed by a post hoc Tukey's
test was used for comparison between multiple groups. P<0.05 was
considered to indicate a statistically significant difference.
Results
ox-LDL treatment increases miR-616-3p
levels in HUVECs
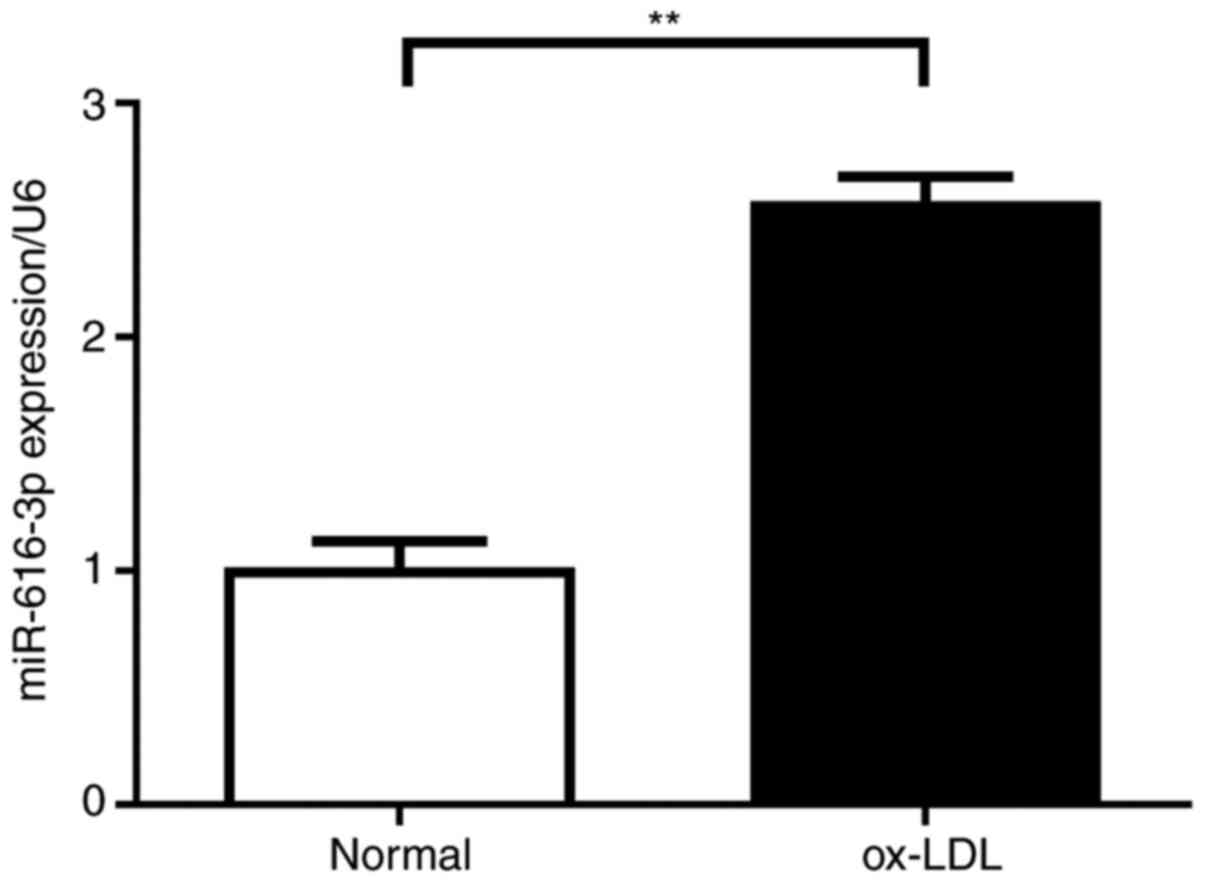
RT-qPCR analysis of HUVECs treated with ox-LDL
revealed that compared with normal HUVECs, ox-LDL treatment
resulted in an increase in the expression of miR-616-3p (Fig. 1).
miR-616-3p inhibits viability and
promotes apoptosis of HUVECs
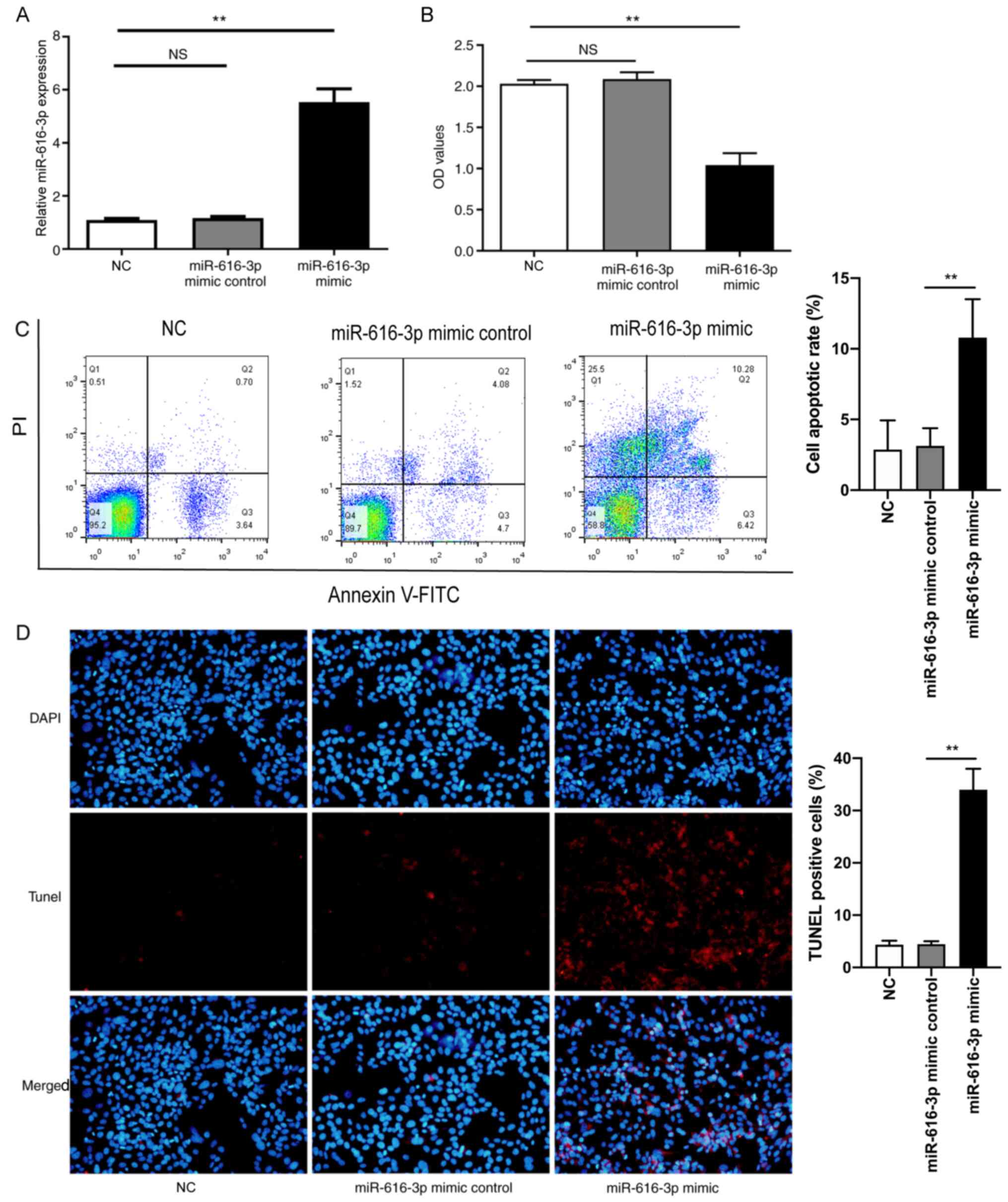
Firstly, HUVECs were transfected with miR-616-3p
mimic and the corresponding non-targeting control. The results
confirmed that compared with miR-616-3p mimic non-targeting
control, miR-616-3p mimic increased miR-616-3p expression (Fig. 2A). Subsequently, the viability of
HUVECs was assessed using the CCK-8 assay. The results demonstrated
that compared with miR-616-3p mimic non-targeting control,
miR-616-3p mimic significantly inhibited the viability of HUVECs
(Fig. 2B). Flow cytometry analysis
and TUNEL staining were used to analyze apoptosis of HUVECs. Flow
cytometry analysis demonstrated that compared with miR-616-3p mimic
control, miR-616-3p mimic significantly increased cell apoptosis
(Fig. 2C). The results of the TUNEL
staining experiment were consistent with the results of flow
cytometry analysis (Fig. 2D).
miR-616-3p directly inhibits XIAP
expression
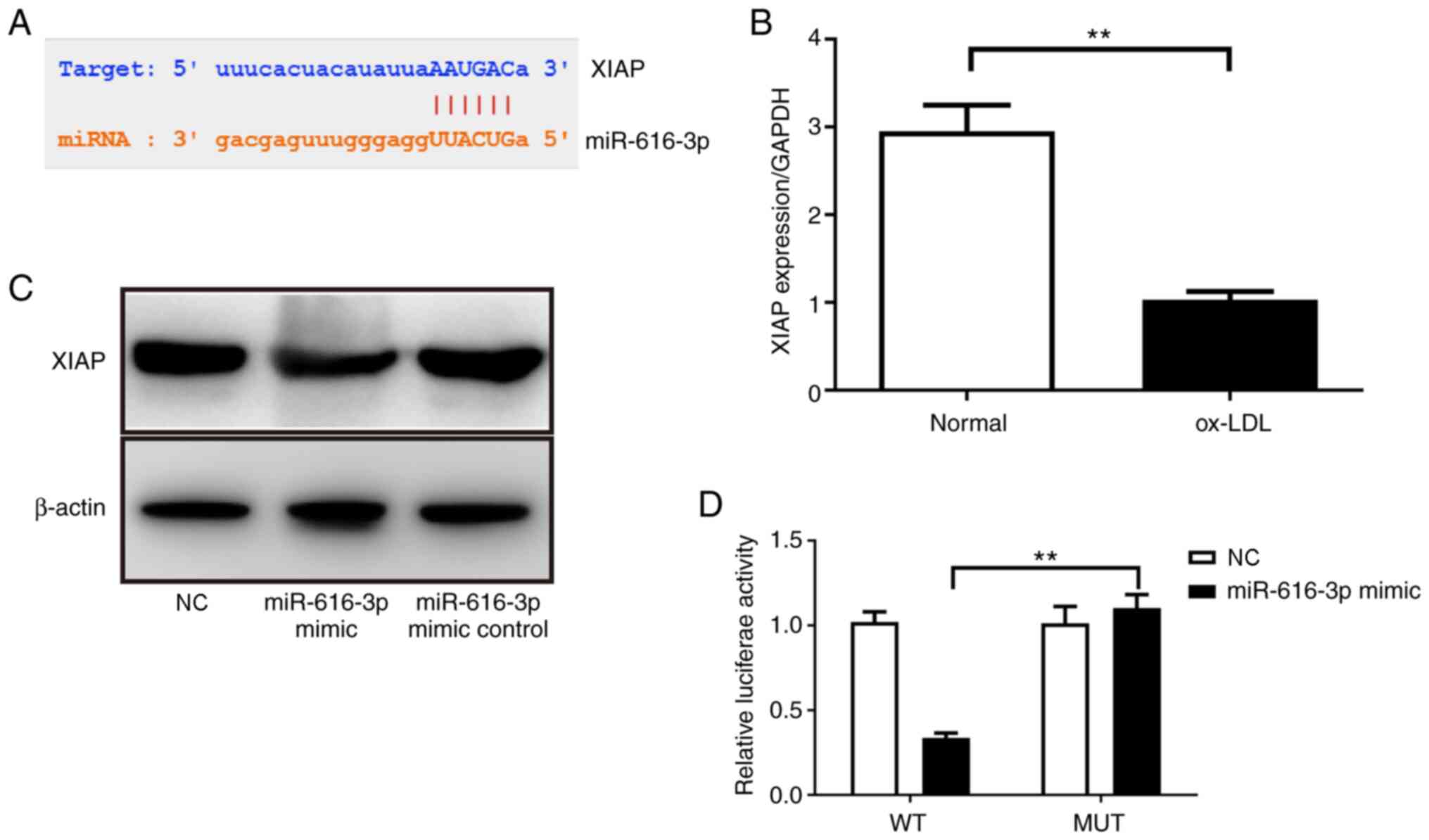
Next, the present study explored the mechanism via
which miR-616-3p inhibits viability and promotes apoptosis in
HUVECs. Based on the StarBase 3.0 prediction, miR-616-3p was found
to have a potential binding site for the 3'UTR of XIAP mRNA
(Fig. 3A). Subsequently, whether
ox-LDL treatment would cause changes in XIAP expression was
assessed. Using RT-qPCR, it was demonstrated that contrary to its
effect on miR-616-3p, ox-LDL treatment inhibited the expression of
XIAP (Fig. 3B). Next, the effect of
miR-616-3p on XIAP in HUVECs was assessed using western blotting.
The results demonstrated that compared with miR-616-3p mimic
non-targeting control, miR-616-3p mimic inhibited XIAP protein
expression (Fig. 3C). Finally, the
direct interaction between miR-616-3p and XIAP was demonstrated
through dual-luciferase experiments. The results revealed that
compared with co-transfection of miR-616-3p mimic and
pmirGLO-XIAP-mut, the fluorescence intensity of cells treated with
pmirGLO-XIAP-wt and the miR-616-3p mimic decreased significantly
(Fig. 3D). This indicated that
miR-616-3p directly acts on XIAP mRNA to inhibit XIAP gene
expression (Fig. 3D).
Inhibition of miR-616-3p expression
inhibits the expression of cleaved caspase-3 protein in HUVECs
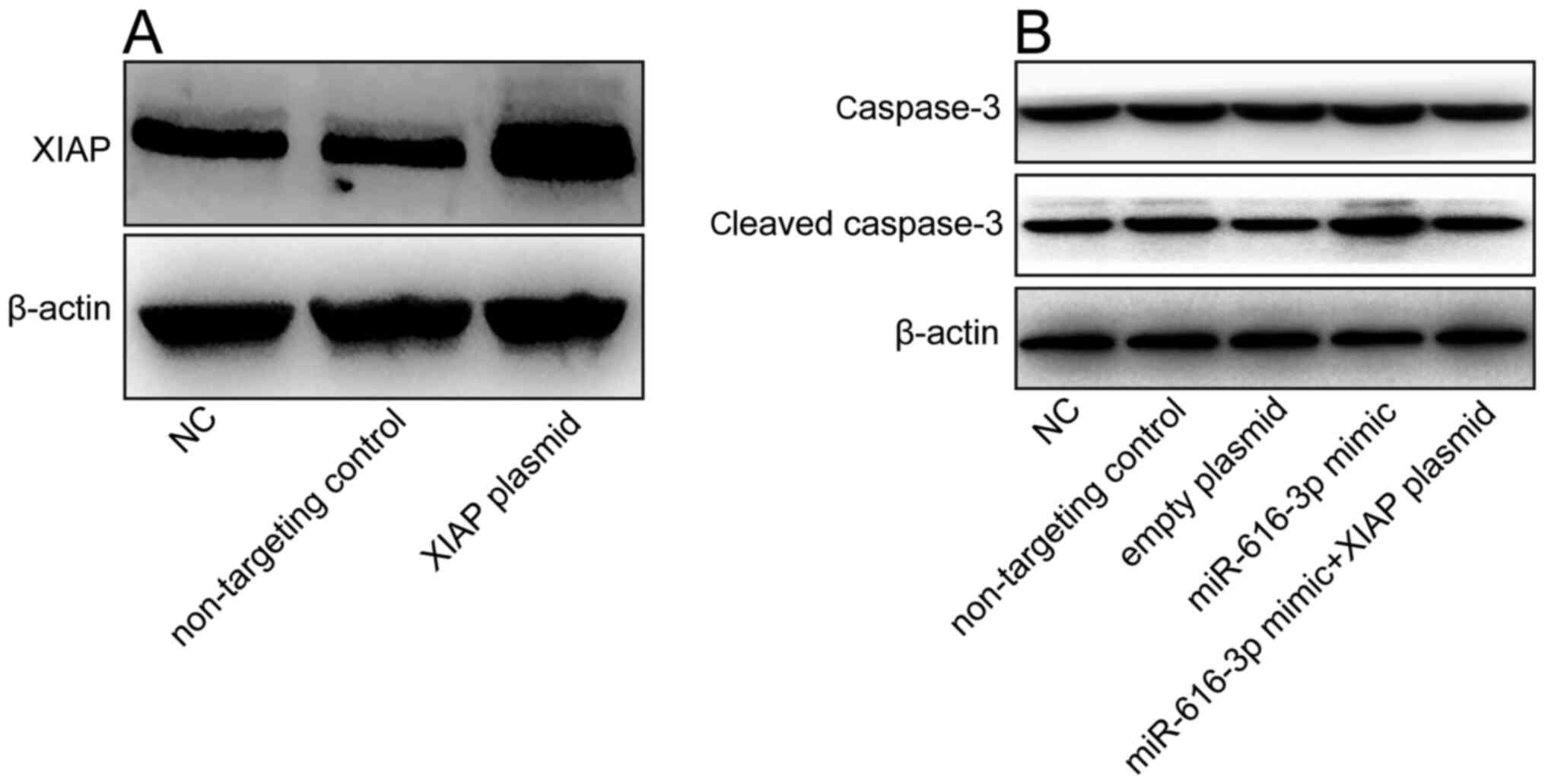
Cleaved caspase-3 is an apoptosis-related protein
(24). Since mR-616-3p was found to
promote apoptosis in HUVECs, western blotting analysis was used to
determine whether miR-616-3p affects the expression of cleaved
caspase-3 protein. Compared with non-targeting control, XIAP
overexpression plasmid significantly increase the expression of
XIAP (Fig. 4A). Co-transfection of
miR-616-3p mimic and XIAP overexpression plasmid inhibited cleaved
caspase-3 protein expression in HUVECs (Fig. 4B). This indicated that the
miR-616-3p mimic promoted apoptosis of HUVECs by inhibiting
XIAP.
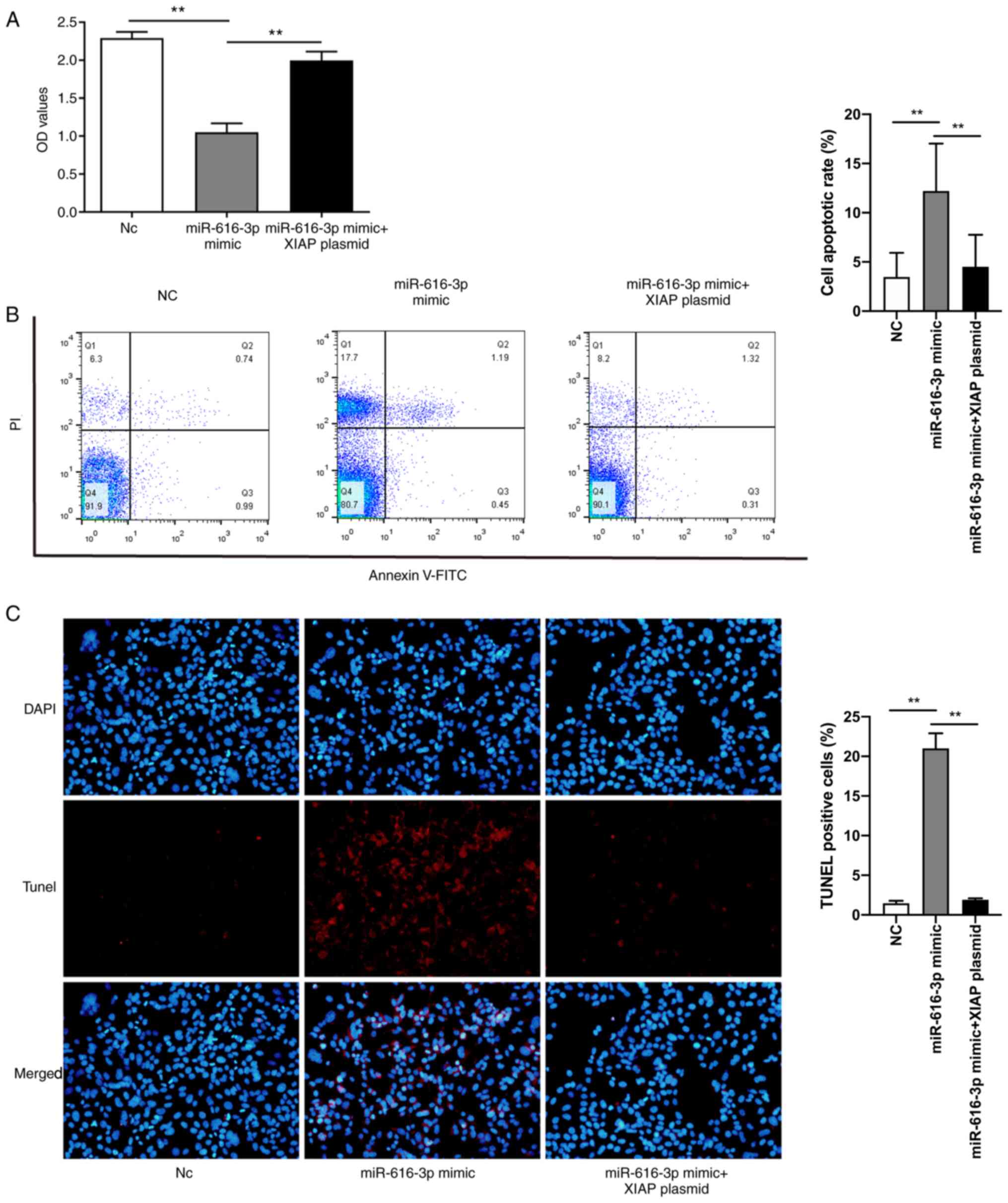
XIAP overexpression plasmid
counteracts the effect of the miR-616-3p mimic on the viability and
apoptosis of HUVECs
Using the CCK-8 assay it was demonstrated that
miR-616-3p mimic inhibited HUVEC viability and this effect was
partially counteracted by the XIAP overexpression plasmid (Fig. 5A). Similarly, in the flow cytometric
analysis and TUNEL staining experiments, it was found that the
miR-616-3p mimic promoted HUVEC apoptosis and the XIAP
overexpression plasmid counteracted this effect (Fig. 5B and C).
Discussion
The present study investigated the role of
miR-616-3p in endothelial cell apoptosis and the potential
mechanisms that may be involved in the context of atherosclerosis.
The results of flow cytometry and TUNEL staining in the present
study demonstrated that miR-616-3p significantly promoted HUVEC
apoptosis. This finding of the present study confirmed the role of
miR-616-3p in atherosclerosis via promotion of apoptosis in
endothelial cells. Using StarBase3.0, the present study predicted
that miR-616-3p may bind to the XIAP mRNA to induce its effects on
HUVECs. Shin et al (25)
demonstrated that miR-513a-5p mediates tumor necrosis-α and
lipopolysaccharide induced apoptosis via downregulation of XIAP in
HUVECs. Another study by Li et al (26) demonstrated that miR-122 promotes
endothelial cell apoptosis by targeting XIAP.
Hence, the present study examined whether miR-616-3p
promoted apoptosis of endothelial cells by directly acting on the
3'UTR of XIAP and inhibiting the expression of XIAP. In the present
study, ox-LDL treatment resulted in an increase in miR-616-3p
expression and decrease in XIAP expression in HUVECs. In addition,
dual-luciferase experiments performed in the present study
demonstrated that miR-616-3p mimic can directly target the XIAP
3'UTR. Flow cytometry and TUNEL staining experiments performed in
the present study also confirmed that miR-616-3p mimic can promote
apoptosis of HUVECs and this effect can be partially reversed by
the XIAP overexpression plasmid.
Caspase-3 is the most important terminal cleavage
enzyme in the process of apoptosis and cleaved-caspase-3 is the
activated form of caspase-3(27).
Through western blotting changes in the expression of the
apoptosis-related protein cleaved caspase-3 were found in the
present study. In the present study, compared with miR-616-3p
non-targeting control, miR-616-3p mimic increased the expression of
cleaved caspase-3 protein and this effect was partially reversed by
the XIAP overexpression plasmid.
The present study had several limitations. Firstly,
the specificity of miR-166-3p and the causal relationship between
miR-166-3p and endothelial cell apoptosis need to be further
verified. Secondly, only in vitro cell experiments were
conducted in the present study and future in vivo
experiments are needed to verify the findings of the present
study.
In summary, the present study found that miR-616-3p
can directly act on the 3'UTR of XIAP to promote apoptosis of
HUVECs. The present study provides a new basis for the pathogenesis
of atherosclerosis and indicates that miR-616-3p may have potential
as a treatment target in the future.
Acknowledgements
Not applicable.
Funding
Funding: This study was supported by a grant from the Natural
Science Foundation of Inner Mongolia (grant no. 2018MS08069). The
funding body did not play a role in the design of the study;
collection, analysis, and interpretation of data and manuscript
writing.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
XZ designed the experiments. HC, XL, YW, XWu, XWen
and YL performed the experiments. HC collected and analyzed the
data. All authors confirmed the authenticity of the raw data. HC
and XZ wrote the manuscript. All authors have read and approved the
final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Boucher P, Matz RL and Terrand J:
atherosclerosis: Gone with the Wnt? Atherosclerosis. 301:15–22.
2020.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Shao C, Wang J, Tian J and Tang YD:
Coronary artery disease: From mechanism to clinical practice. Adv
Exp Med Biol. 1177:1–36. 2020.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Libby P, Buring JE, Badimon L, Hansson GK,
Deanfield J, Bittencourt MS, Tokgözoğlu L and Lewis EF:
Atherosclerosis. Nat Rev Dis Primers. 5(56)2019.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Grechowa I, Horke S, Wallrath A, Vahl CF
and Dorweiler B: Human neutrophil elastase induces endothelial cell
apoptosis by activating the PERK-CHOP branch of the unfolded
protein response. FASEB J. 31:3868–3881. 2017.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Gomez D, Baylis RA, Durgin BG, Newman AAC,
Alencar GF, Mahan S, St HC, Müller W, Waisman A, Francis SE, et al:
Interleukin-1β has atheroprotective effects in advanced
atherosclerotic lesions of mice. Nat Med. 24:1418–1429.
2018.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Singh R, Devi S and Gollen R: Role of free
radical in atherosclerosis, diabetes and dyslipidaemia:
Larger-than-life. Diabetes Metab Res Rev. 31:113–126.
2015.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Gimbrone MA Jr and García-Cardeña G:
Endothelial cell dysfunction and the pathobiology of
atherosclerosis. Circ Res. 118:620–636. 2016.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Rajendran P, Rengarajan T, Thangavel J,
Nishigaki Y, Sakthisekaran D, Sethi G and Nishigaki I: The vascular
endothelium and human diseases. Int J Biol Sci. 9:1057–1069.
2013.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Stoneman VE and Bennett MR: Role of
apoptosis in atherosclerosis and its therapeutic implications. Clin
Sci (Lond). 107:343–354. 2004.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Werner N, Wassmann S, Ahlers P, Kosiol S
and Nickenig G: Circulating CD31+/annexin V+ apoptotic
microparticles correlate with coronary endothelial function in
patients with coronary artery disease. Arterioscler Thromb Vasc
Biol. 26:112–116. 2006.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Libby P, Okamoto Y, Rocha VZ and Folco E:
Inflammation in atherosclerosis: Transition from theory to
practice. Circ J. 74:213–220. 2010.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Paone S, Baxter AA, Hulett MD and Poon
IKH: Endothelial cell apoptosis and the role of endothelial
cell-derived extracellular vesicles in the progression of
atherosclerosis. Cell Mol Life Sci. 76:1093–1106. 2019.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Krol J, Loedige I and Filipowicz W: The
widespread regulation of microRNA biogenesis, function and decay.
Nat Rev Genet. 11:597–610. 2010.PubMed/NCBI View
Article : Google Scholar
|
|
14
|
Zhong X, Li P, Li J, He R, Cheng G and Li
YL: Downregulation of microRNA-34a inhibits oxidized low-density
lipoprotein-induced apoptosis and oxidative stress in human
umbilical vein endothelial cells. Int J Mol Med. 42:1134–1144.
2018.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Zheng B, Yin WN, Suzuki T, Zhang XH, Zhang
Y, Song LL, Jin LS, Zhan H, Zhang H, Li JS and Wen JK:
Exosome-mediated miR-155 transfer from smooth muscle cells to
endothelial cells induces endothelial injury and promotes
atherosclerosis. Mol Ther. 25:1279–1294. 2017.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Wei Q, Tu Y, Zuo L, Zhao J, Chang Z, Zou Y
and Qiu J: miR-345-3p attenuates apoptosis and inflammation caused
by oxidized low-density lipoprotein by targeting TRAF6 via
TAK1/p38/NF-kB signaling in endothelial cells. Life Sci.
241(117142)2020.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Jing R, Zhong QQ, Long TY, Pan W and Qian
ZX: Downregulated miRNA-26a-5p induces the apoptosis of endothelial
cells in coronary heart disease by inhibiting PI3K/AKT pathway. Eur
Rev Med Pharmacol Sci. 23:4940–4947. 2019.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Zhong X, Zhang L, Li Y, Li P, Li J and
Cheng G: Kaempferol alleviates ox-LDL-induced apoptosis by
up-regulation of miR-26a-5p via inhibiting TLR4/NF-κB pathway in
human endothelial cells. Biomed Pharmacother. 108:1783–1789.
2018.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Qin B, Shu Y, Long L, Li H, Men X, Feng L,
Yang H and Lu Z: MicroRNA-142-3p induces atherosclerosis-associated
endothelial cell apoptosis by directly targeting rictor. Cell
Physiol Biochem. 47:1589–1603. 2018.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Wang Z, Chen S, Zhu M, Zhang W, Zhang H,
Li H and Zou C: Functional SNP in the 3'UTR of PON1 is associated
with the risk of calcific aortic valve stenosis via MiR-616. Cell
Physiol Biochem. 45:1390–1398. 2018.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Liu ME, Liao YC, Lin RT, Wang YS, His E,
Lin HF, Chen KC and Juo SH: A functional polymorphism of PON1
interferes with microRNA binding to increase the risk of ischemic
stroke and carotid atherosclerosis. Atherosclerosis. 228:161–167.
2013.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Feng Y, Cai ZR, Tang Y, Hu G, Lu J, He D
and Wang S: TLR4/NF-κB signaling pathway-mediated and oxLDL-induced
up-regulation of LOX-1, MCP-1, and VCAM-1 expressions in human
umbilical vein endothelial cells. Genet Mol Res. 13:680–695.
2014.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Zhuo EQ, Cai CQ, Liu WZ, Li KS and Zhao
WZ: Downregulated microRNA-140-5p expression regulates apoptosis,
migration and invasion of lung cancer cells by targeting zinc
finger protein 800. Oncol Lett. 20(390)2020.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Shin S, Moon KC, Park KU and Ha E:
MicroRNA-513a-5p mediates TNF-α and LPS induced apoptosis via
downregulation of X-linked inhibitor of apoptotic protein in
endothelial cells. Biochimie. 94:1431–1436. 2012.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Li Y, Yang N, Dong B, Yang J, Kou L and
Qin Q: MicroRNA-122 promotes endothelial cell apoptosis by
targeting XIAP: Therapeutic implication for atherosclerosis. Life
Sci. 232(116590)2019.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Jiang M, Qi L, Li L and Li Y: The
caspase-3/GSDME signal pathway as a switch between apoptosis and
pyroptosis in cancer. Cell Death Discov. 6(112)2020.PubMed/NCBI View Article : Google Scholar
|