Introduction
Spinal cord injury (SCI) frequently causes
persistent functional deficits due to the absence of spontaneous
axon regeneration, formation of large cavities and glial scars that
interrupt the ascending and descending neural pathways (1,2). In
rodent models of SCI, levels of pro-inflammatory interleukins
(ILs), such as IL-6, peak acutely in the injured areas and lead to
the activation of the Janus kinase 1-signal transducer and STAT3
signaling pathway (3). STAT3
signaling is also upregulated in a number of neurodegenerative
diseases. For instance, in patients with amyotrophic lateral
sclerosis, the spinal cord microglia, reactive astrocytes and motor
neuron nuclei were all shown to exhibit increased levels of
phosphorylated (p-) STAT3 (p-STAT3) (4). Another study previously reported that
STAT3 is an important signaling component during reactive
astrogliosis through the Notch 1-STAT3-endothelin receptor type B
signaling axis (5). A growing body
of evidence suggests that STAT3 is an injury-induced signaling
mechanism that is critical for various aspects of nerve
regeneration (6-8).
In addition, in vitro suppression of STAT3(9) or its conditional deletion in
vivo (10) have been
demonstrated to induce neurogenesis and inhibit astrogliosis.
Therefore, STAT3 appears to be the key to enhancing neurogenesis
following SCI.
Neural stem cell (NSC) transplantation has been
demonstrated to be a promising treatment method for improving
tissue repair and functional recovery following SCI (11). Mechanistically, NSCs may
differentiate into neurons and glial cells to bridge the damaged
area, re-establish conduction pathway, form functional synapses by
expressing genes associated with axonal regeneration and by
secreting neurotrophic growth factors in the injured spinal cord
(12). However, accumulating
evidence has suggested that NSC-based therapy cannot achieve
optimal results, mainly due to the limited differentiation
potential of neurons and excessive differentiation of astrocytes,
which contributes to glial scarring (13). Natarajan et al (14) demonstrated that treatment with an
inhibitor of STAT3 promoted NSC differentiation into neurons and
suppressed glial differentiation in vitro, a finding that
was also subsequently confirmed by White et al (15). However, few studies have assessed
the effects of STAT3 silencing on NSCs transplantation and its
functional outcomes downstream in vivo.
In the present study, the effects of STAT3 RNA
interference (RNAi) on NSC survival and differentiation in
vitro and the potential mechanisms underlying these effects
were first explored. NSCs with STAT3 expression were the knocked
down before being injected into the injury site of a rat SCI model
to investigate its effects on neurodegeneration in vivo. The
focus was placed on exploring the effect of transplanting
STAT3-silenced NSCs on spinal cord injury to provide novel targets
for the treatment and rehabilitation of SCI in the future.
Materials and methods
Lentiviral vector construction
Green fluorescent protein (GFP) was also encoded by
the lentiviral vector with a sequence that specifically silenced
the STAT3 gene [STAT3-short hairpin RNA (shRNA); Shanghai GeneChem
Co., Ltd.]. The oligonucleotides were synthesized and ligated into
the GV248 plasmid-Hu6-MCS-Ubiquitin-IRES-puromycin plasmid (cat.
no. PC0NGC248028254; Shanghai GeneChem Co., Ltd.) and had the
following STAT3-shRNA sequence: 5'-CAGCAGATGCTGGAACAGCAT-3'. A
nontargeting sequence (5'-TTCTCCGAACGTGTCACGT-3') was also carried
by a control lentivirus (LV) vector. The LV particles were
generated as previously described (16) to a final titer of 1x108
TU/ml.
NSC isolation and culture
3-month old pregnant Sprague-Dawley rats (Laboratory
Animal Center of Sun Yat-sen University; n=3) were sacrificed and
NSCs were extracted from the brain tissues of the fetuses (n=14) on
embryonic day 14 as previously described (17). The animals were housed three to a
cage with free access to food/water and kept under standardized
atmosphere (temperature, 22˚C; humidity, 55%; 12/12 h light/dark
cycle). Rats were anesthetized with 1% pentobarbital sodium (40-45
mg/kg) to minimize pain and sacrificed by CO2 with the
displacement rate at 20%/min). Fetal brain tissue was mechanically
cut and removed in Hanks' balanced salt solution (Beijing Solarbio
Science & Technology Co., Ltd.), following which the cell
suspension was centrifuged at 300 x g and 4˚C for 5 min. The
supernatant was discarded and the cell pellet was diluted into
DMEM/F-12 (Thermo Fisher Scientific, Inc.) as a single-cell
suspension. NSCs were then plated in a T25 culture flask (Corning,
Inc.) containing DMEM/F-12 (Thermo Fisher Scientific, Inc.)
nutrient mixture, 1% L-glutamine (Thermo Fisher Scientific, Inc.),
2% B27, 1% penicillin/streptomycin, 20 ng/ml fibroblast growth
factor-2 (FGF2; PeproTech, Inc.) and 20 ng/ml epidermal growth
factor (EGF; PeproTech, Inc.). NSCs were cultured in 5%
CO2 at 37˚C and passaged weekly by digesting with
Accutase (EMD Millipore) in the aforementioned medium. NSCs at
passages 2-4 were used for subsequent experiments (15,16).
NSC transfection and
differentiation
In the DMEM/F12 nutrient mixture, NSCs form
neurospheres spontaneously. For cell transfection, 2nd passage
neurospheres were dissociated into a single-cell suspension at a
density of 1x105 cells/ml and plated onto coverslips
coated with 0.01% poly-L-lysine (Merck KGaA) in 12-well plates
(Corning, Inc.). The aforementioned culture medium was then added
to the 12-well plate to rejuvenate the cells. Cells were
subsequently divided into the following three groups: No
transfected NSCs group (CL, n=5), Lv-control shRNA transfected NSCs
group (LV-CL, n=5), Lv-STST3 shRNA transfected NSCs group
(LV-STAT3, n=5). The medium was then changed to fresh medium after
24 h. After 72 h transfection, GFP expression was visualized by
fluorescence microscopy (x40 magnification; AG Axio Observer Z1;
Zeiss AG). To quantify the suppressive effects of RNAi on STAT3
expression, STAT3 expression in each group was examined by reverse
transcription-quantitative PCR (RT-qPCR) and western blotting. To
assess the differentiation capacity of NSCs, the culture medium was
changed to the differentiation medium (culture medium without
growth factors EGF and FGF2) for 1 week after transfection.
Western blotting
Protein lysates (RIPA Lysis and Extraction Buffer;
cat. no. 89901; Thermo Fisher Scientific, Inc.) were extracted from
the cells used in the in vitro study (n=5 per group,
harvested 3 days after transfection). Bicinchoninic acid assay
(Beyotime Institute of Biotechnology) was used to measure protein
concentration, which was equilibrated prior to loading. Protein
samples in each group were separated by SDS-PAGE (10% Bis-Tris
gel), transferred onto PVDF membranes (EMD Millipore) and blocked
with 5% BSA (Merck KGaA) for 1 h at 24˚C, followed by incubation
with primary antibodies at 4˚C overnight. Primary antibodies
targeting STAT3 (cat. no. ab68153; Abcam), p-STAT3 (cat. no. 9145;
Cell Signaling Technology, Inc.), mTOR (cat. no. 2983; Cell
Signaling Technology, Inc.), p-mTOR (cat. no. 2971; Cell Signaling
Technology, Inc.) and GAPDH (cat. no. 5174; Cell Signaling
Technology, Inc.) were used at a 1:1,000 dilution. After washing
the membranes in Tris-HCl buffer containing 0.2% Tween-20 (TBST; pH
7.5) twice, the membranes were incubated with a filtered and
horseradish peroxidase-conjugated goat anti-rabbit IgG antibody
(1:5,000; cat. no. ab6721; Abcam) for 1 h at room temperature and
then washed three times in TBST. Finally, the bands were visualized
using enhanced chemiluminescence development system (EMD
Millipore). For analysis of the Western blot images ImageJ 1.50
(National Institutes of Health) was used.
RNA extraction and RT-qPCR
Total cell RNA (n=5 per group, harvested 3 days
after transfection) was extracted using TRIzol® reagent
(Invitrogen; Thermo Fisher Scientific, Inc.). RNA was reverse
transcribed into cDNA using a reverse transcription system
(GoScript™ Reverse Transcription System; cat. no. A5000; Promega
Corporation). The process used was as follows: Combine RNA and cDNA
primers at 70˚C for 5 min, followed by adding reverse transcription
mix into primer/RNA mix at 25˚C for 5 min, 42˚C for 60 min and 70˚C
for 15 min. qPCR was performed on an ABI 7900 PCR detection system
(Thermo Fisher Scientific, Inc.) using SYBR™ Green PCR Master Mix
(cat. no. 4309155, Thermo Fisher Scientific, Inc.). The process
used was as follows: 95˚C for 5 min, followed by 40 cycles (95˚C
for 10 sec and 60˚C for 30 sec). Parallel amplification of the
GAPDH housekeeping gene was used to normalize gene expression. PCR
primer sequences are listed below: STAT3 forward,
5'-AATATAGCCGATTCCTGCAAGAG-3' and reverse,
5'-TGGCTTCTCAAGATACCTGCTC-3' and GAPDH forward,
5'-TGACGCTGGGGCTGGCATTG-3' and reverse, 5'-GGCTGGTGGTCCAGGGGTCT-3'.
The relative expression level of target mRNA was calculated using
the 2-∆∆Cq method (18).
Surgical procedures and cell
transplantation
All experimental animal procedures were approved by
the Care and Use Committee of Sun Yat-Sen University (approval no.
SYXX2012-0081; November 9, 2016) and performed following the Guide
for the Care and Use of Experimental Animals provided by the
National Research Council (1996, USA) (19). All the animals were housed three to
a cage with free access to food and water, where they were kept
under a standardized atmosphere (temperature, 22˚C; humidity, 55%;
12-h light/dark cycle).
Spinal cord surgery was performed on 60 adult female
SD rats (weight range 200-250 g laboratory Animal Center of Sun
Yat-sen University). All rats were healthy and did not undergo any
previous procedures. The rats were randomly divided into the
following four groups: i) Sham (spinal cord exposure only, n=10);
ii) SCI (n=15); iii) STAT3-shRNA-transfected NSC (LS group, n=20);
and iv) Control-shRNA-transfected NSC (LC group, n=15). In the Sham
group, the rats underwent laminectomy without transection of the
spinal cord. In the SCI group, rats underwent surgery with complete
transection of the spinal cord. In the LC group, NSCs transfected
with control LV were injected immediately into the rats following
complete transection of the spinal cord. In the LS group, NSCs
transfected with STAT3 LV were injected immediately into the rat's
complete transection of the spinal cord. Briefly, animals were
intraperitoneally injected with 1% sodium pentobarbital (40-45
mg/kg) for anesthesia. Laminectomy was then performed at the level
of the 10th thoracic vertebra (T10). Next, the spinal cord was cut
twice using scissors (once caudal to T10 and once rostral to T10)
to achieve complete transection, following which a 2-mm block of
the spinal cord was removed. Following hemostasis, the rats were
injected with 5 µl control-shRNA-transfected NSCs or
STAT3-shRNA-transfected NSCs at a density of 1x105
cells/µl. Two injections were performed at the depth of 1 mm
rostral, as well as 1 mm caudal to the injured site using a
microsyringe at a rate of 0.5 µl/min. Finally, following
hemostasis, 5-0 sutures were used to suture the muscle and skin. To
prevent infection and dehydration, 1 ml saline containing
1x105 units of penicillin was intraperitoneally injected
daily for 1 week (20). The
postsurgical care of SCI rats included massaging the urinary
bladder twice a day for urination until bladder function was
restored. Surgery was performed in a blinded manner. At the 7th
week after surgery, 5 rats were randomly selected from each group
for retrograde tracking experiments [injection of Fluorogold (FG)].
After the experiment was completed, the rats were not sacrificed
and continued to be raised for 1 week. On the 8th week, 5 rats were
randomly selected from each group for the SCEP recording
experiment. After the SCEP recording experiment, rats were
anesthetized with 1% pentobarbital sodium (40-45 mg/kg) and then
sacrificed by CO2 inhalation, using a displacement rate
of 20%/min, for H&E staining (n=5 per group), fluorescent
immunohistochemistry (n=5 per group) and neuron retrograde tracing
staining (n=5 per group).
Basso-Beattie-Bresnahan (BBB)
test
The BBB locomotor rating scale is considered to be a
reliable tool for evaluating impairment and recovery of motor
abilities in hindlimbs following spinal cord injury (21) A total score of 21 points indicate
that the locomotor ability had not been affected, while 0 points
represents a severe deficit. The rats were placed in an open
experimental field and allowed to move freely for 5 min. During
this period, the lower limb movement ability of the rats was
observed and the movement of each rat was scored based on the BBB
score scale. Hindlimb motor behavior was evaluated weekly for 8
weeks, with tests performed at the same time each day and grading
performed by two investigators (XZ and JD) blinded to grouping.
Spinal cord-evoked potential (SCEP)
recording
At 8 weeks post-injury, 20 rats (n=5 in each group)
were anesthetized with 1% pentobarbital sodium (40-45 mg/kg) and
stereotaxically fixed. The T5-L1 vertebrae were then completely
exposed. Finally, a stimulation electrode was inserted into the
T5-T6 interspinous ligaments, following which a pair of needle
electrodes were inserted into the interspinous ligaments of T12-L1
for SCEP recording. The electrodes were connected to a BL-420N
series biological signal acquisition and analysis system (Chengdu
Thai UNITA Co., Ltd.). The parameters of the SCEP signals were set
as follows: i) Gain, 2,000; ii) Time constant, 0.01 sec; and iii)
Filtering, 300 Hz. To elicit a SCEP, a single-pulse stimulation (50
msec in duration at a frequency of 5.1 Hz and a voltage increase of
1 mV) was transmitted through the electrodes until a mild twitch of
the vertebral body of the animal was observed. To obtain
high-quality waveforms for the SCEP signals, 100 SCEP responses per
rat were averaged.
Retrograde axonal tract tracing
Animals (n=5 per group) were submitted for neuron
retrograde tracing using 4% FG (cat. no. sc-358883; dilution, 1:25;
Santa Cruz Biotechnology, Inc.) 7 weeks after SCI. Briefly, a
dorsal laminectomy was performed under anesthesia (40-45 mg/kg
pentobarbital) at T12 before 0.5 µl FG was injected into the spinal
cord using a microsyringe. A week after injection, the animals were
perfused after CO2 euthanasia and the T7 segment of the
spinal cord was collected to detect FG-labelled neurons (22).
Perfusion and cryosection
Rats were subjected to anesthesia with 1%
pentobarbital sodium (40-45 mg/kg) 8 weeks after spinal cord
surgery and sacrificed by CO2 using a displacement rate
of 20%/min. Rats were then perfused transcardially with 0.9% normal
saline and 400 ml 4% paraformaldehyde (PFA). T8-L1 segments of the
spinal cord were removed from rats without Fluorogold (FG)
labelling and T7-L1 segments were removed from Fluorogold labelled
rats. These segments were then fixed with 4% PFA at room
temperature overnight and transferred to 30% sucrose for
cryoprotection for 2-3 days at 4˚C following collection. Tissue
were embedded into OCT Tissue-Tek (cat. no. 4583; Sakura Finetek
USA, Inc.). Embedded tissues were then sliced transversely or
longitudinally at a thickness of 10 µm, mounted onto
polylysine-coated glass slides and stored at -20˚C to be used in
subsequent experiments.
Histopathological analysis
To assess the cavity area in the spinal cord, rats
were sacrificed for H&E staining 8 weeks after SCI. OCT
Component (Tissue-Tek Frozen Embedding Agent; cat. no. 4583;
Sakura)-embedded T8-T11 longitudinal spinal cord sections from each
group were fixed in 4% paraformaldehyde for 30 min at room
temperature, rehydrated in a graded series of ethanol (100% twice,
95% twice, 90% and 85% for 5 min at room temperature), then the
tissue was treated with hematoxylin solution for 5 min and eosin
solution (cat. no. C0105S; Beyotime Institute of Biotechnology) for
1 min for H&E staining at room temperature or treated with
Nissl solution (cat. no. C0117; Beyotime Institute of
Biotechnology) for 5 min for Nissl staining at room temperature,
rehydrated in the graded series of ethanol and xylene (50%, 100%
twice for 1 min) at room temperature and observed using a light
microscope at x20 magnification. The cavity area of these images
was evaluated using NIH ImageJ software (1.50; National Institutes
of Health).
Fluorescence immunohistochemistry
Frozen sections of spinal cord tissue sections or
cells were fixed in 4% PFA for 30 min and permeabilised with 0.3%
Triton X-100 for 30 min, both at 4˚C. Blocking was performed with
5% normal goat serum (cat. no. C0265; Beyotime Institute of
Biotechnology) for 1 h at 4˚C and tissue sections were incubated
with primary antibodies targeting the following proteins at 4˚C
overnight: Nestin (dilution, 1:200; cat. no. 33475; Cell Signaling
Technology, Inc.), β3-tubulin (dilution, 1:200; cat. no. 5568; Cell
Signaling Technology, Inc.), GFAP (dilution, 1:300; cat. no. 80788;
Cell Signaling Technology, Inc.) and microtubule-associated protein
2 (MAP2; dilution, 1:200; cat. no. 8707; Cell Signaling Technology,
Inc.). Goat anti-rabbit IgG (H+L) Cross-Adsorbed Secondary
Antibody, Alexa Fluor 555 (cat. no. A-21428; Thermo Fisher
Scientific, Inc.), was used at a concentration of 2 µg/ml in PBS
containing 0.2% BSA for 45 min at room temperature. ProLong Gold
antifade reagent containing DAPI (dilution: 1:5000; cat. no. D1306;
Thermo Fisher Scientific, Inc.) was used for nuclear staining at
room temperature in spinal cord tissue sections. The total area of
target marker-positive cells was then evaluated in each visual
field under fluorescence (x40 magnification; Carl Zeiss Axio
Observer Z1; Carl Zeiss AG) using ImageJ software (1.50, National
Institutes of Health). A total of five random fields per section
and five sections per group were examined independently by two
observers (XZ and JD) blinded to grouping. The percent of
β3-tubulin or GFAP-positive cells was calculated by dividing the
number of β3-tubulin or GFAP-positive cells by the total number of
cells.
Statistical analysis
All statistical analyses were performed using
GraphPad Prism 6 software (GraphPad Software, Inc.). Shapiro-Wilk
test was first used to evaluate the normality of all data in the
present study. Experiments were repeated 5 times in vitro
study, All data are expressed as the mean ± SD and analyzed using
one-way analysis of variance followed by Bonferroni post hoc tests
for multiple comparisons or Student t-test for pairwise
comparisons. P<0.05 was considered to indicate a statistically
significant difference.
Results
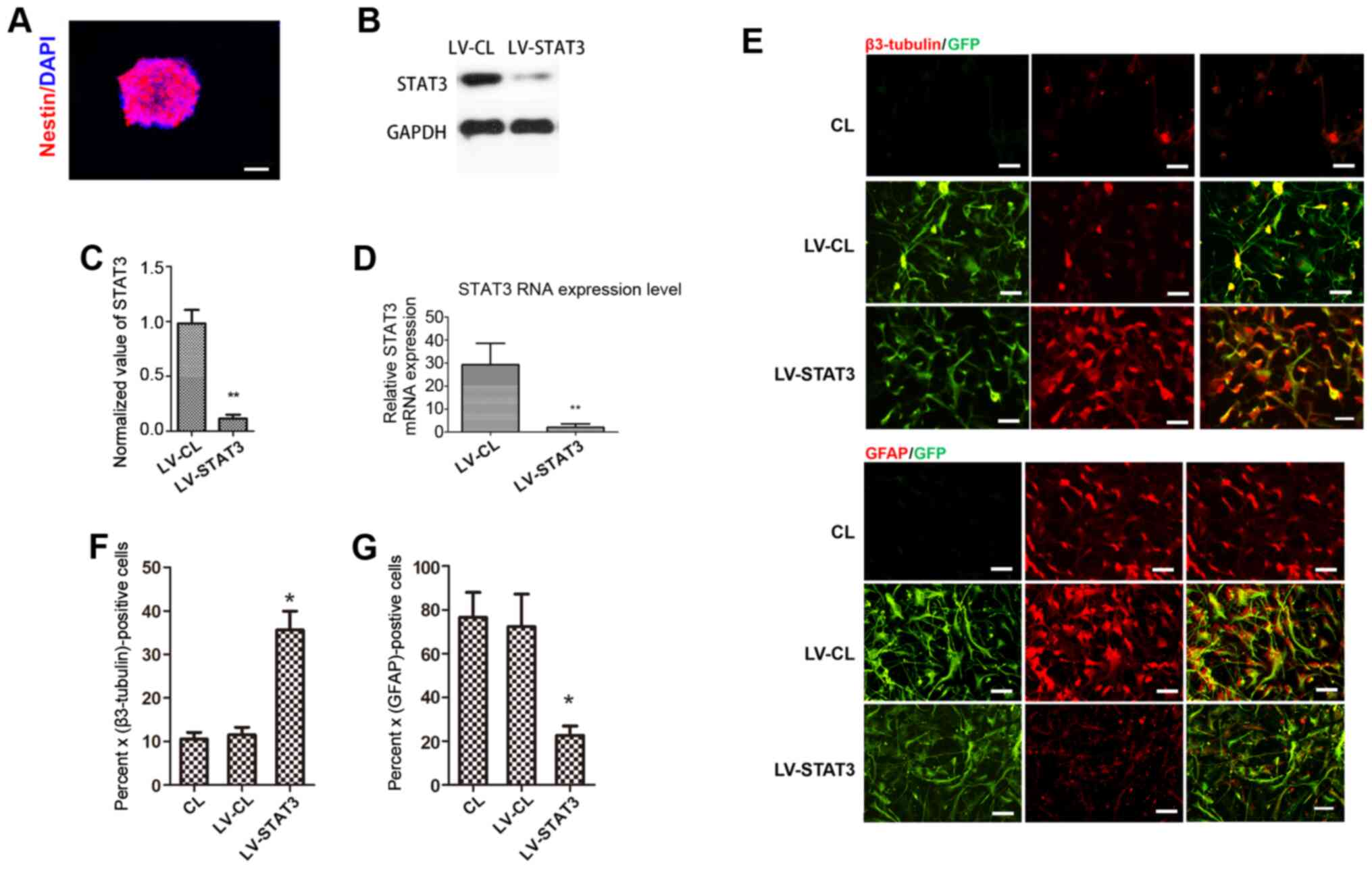
Knockdown of STAT3 promotes neuronal
differentiation in NSCs
NSCs were passaged and allowed to grow for an
additional 3-5 days to form a 100-µm-diameter neurosphere from
single cells. Immunostaining of the neurospheres showed that
nestin, a surface marker of neural stem and precursor cells
(23), was strongly expressed in
the neurosphere (Fig. 1A). To
achieve a specific knockdown of the STAT3 gene in NSCs, NSCs were
transfected with STAT3 shRNA-expressing LV. GFP expression was
observed by fluorescence microscopy 72 h after LV infection.
Western blotting showed that STAT3 expression was significantly
lower in the LV-STAT3 group compared with that in the LV-CL group
(Fig. 1B and C). This observation was also confirmed by
RT-qPCR analysis (Fig. 1D).
Immunofluorescence staining analysis showed that the targeted
inhibition of STAT3 induced the differentiation of NSCs into
neurons, as the neuronal marker β3-tubulin (24) was found to be expressed more
frequently in the LV-STAT3 group compared with that in the CL and
LV-CL groups. However, GFAP, a specific marker of astrocytes
(23), was expressed at
significantly lower levels in the LV-STAT3 group compared with
those in the other two groups (Fig.
1E-G). These results suggest that a specific knockdown of STAT3
gene expression was achieved in NSCs, which supports the hypothesis
that targeted inhibition of STAT3 promotes neuronal differentiation
in NSCs, consistent with data from a previous study.
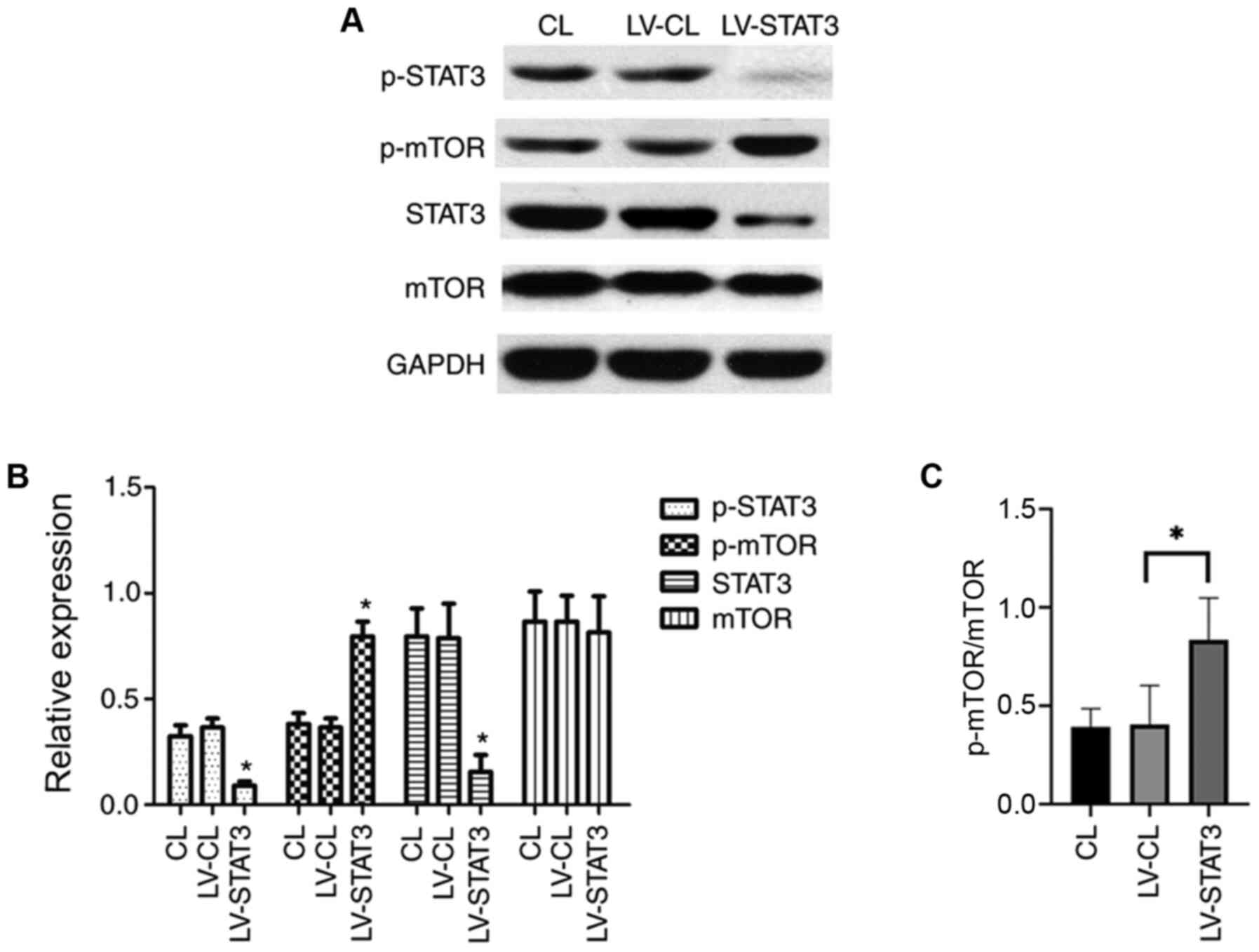
Targeted inhibition of STAT3 promotes
mTOR activation
mTOR signaling is important for the maintenance and
differentiation of NSC development, where the activation of mTOR
promotes the involvement of NSCs in neurogenesis (25). To explore the relationship between
the mTOR and STAT3 signaling pathways in NSC differentiation, the
STAT3 signaling pathway was inhibited by treating NSCs that were
transfected with shRNA-expressing LV. NSCs were cultured in a
differentiation medium following transfection. On day 7, western
blotting was performed to examine the expression of STAT3 and mTOR.
The expression of STAT3 itself and STAT3 phosphorylation was
significantly lower in the LV-STAT3 group compared with that in the
LV-CL group. Of note, although no difference in total mTOR
expression was observed among the three groups, the levels of the
activated form of mTOR (p-mTOR) was significantly higher in the
LV-STAT3 group, compared with that in the LV-CL groups (Fig. 2A and B). These results suggest that STAT3
inhibition activated the mTOR signaling pathway, which may promote
neuronal differentiation of NSCs.
Transplantation of
STAT3-RNAi-transfected NSCs enhances functional recovery following
SCI
To investigate if transplantation with NSCs with
STAT3 expression knocked down promotes the recovery of motor
function following SCI in rats, the rats were divided into four
groups. On week 8 after surgery, hindlimb weight-bearing
experiments were performed in the four groups of rats. The rats in
the sham group were able to use their hindlimbs to walk normally,
whilst rats in the LC and SCI groups exhibited poor hindlimb
strength during walking. By contrast, rats in the LS group could
sometimes stand on their hind legs to support their weight and walk
slowly (Fig. 3A). Results from BBB
functional score curve also showed that the score in the LS group
was significantly higher compared with that in the LC group from
week 5 onwards after SCI (Fig. 3B).
The electrophysiological restoration of SCEP responses was also
subsequently explored. The SCEP waveforms in the LC and SCI group
were significantly different from the SCEP waveform of the sham
group, exhibiting a significantly prolonged latency and decreased
amplitude. However, compared with that in the LC and SCI group, the
LS group had a significantly shorter SCEP latency and significantly
greater amplitude (Fig. 3C-E).
Taken together, these results suggest that transplantation with
STAT3-knockdown NSCs promoted functional and neurological recovery
in rats following SCI.
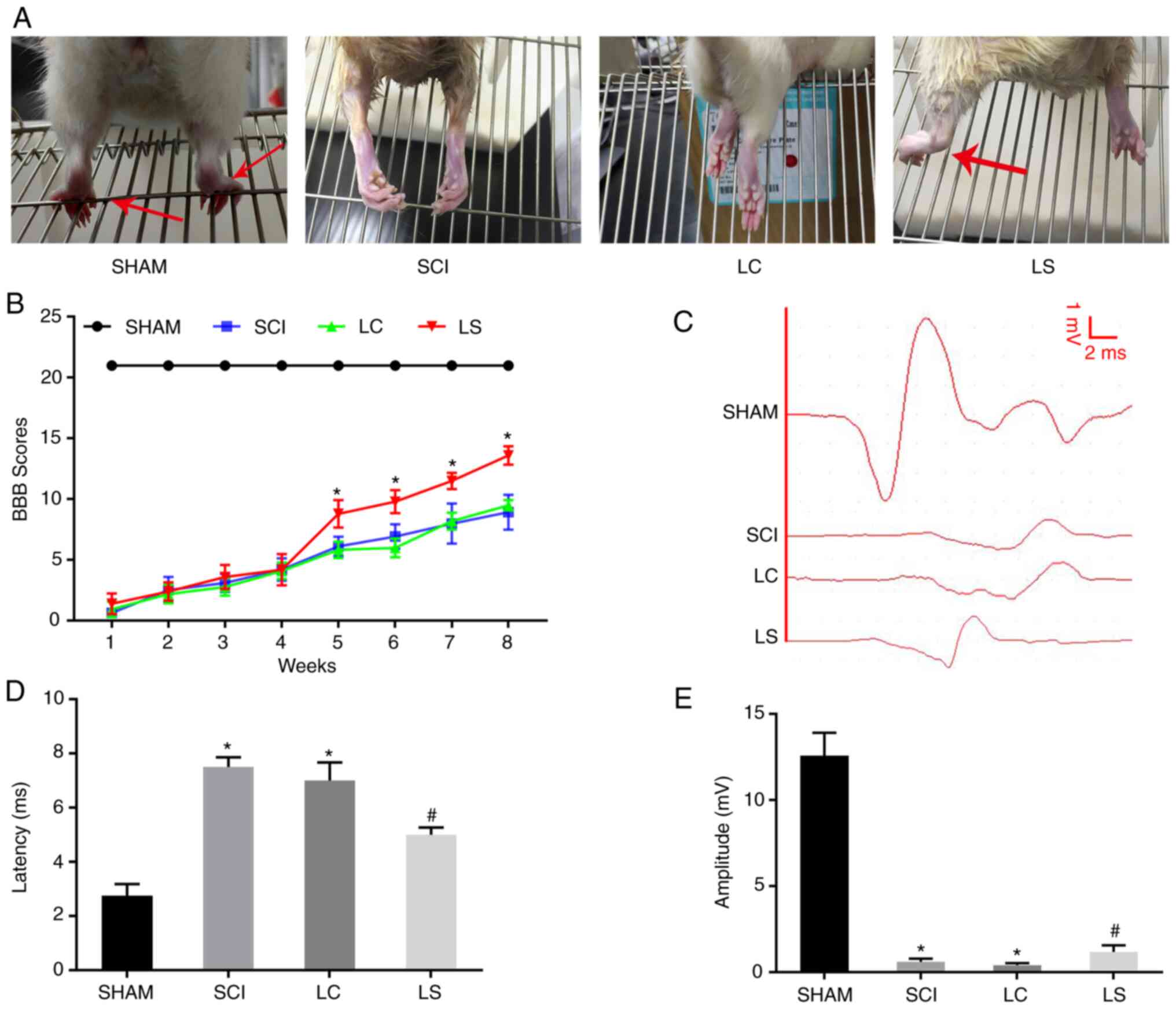 | Figure 3Transplantation of
STAT3-RNAi-transfected NSCs enhances functional recovery following
SCI. (A) Images showing hindlimb movements in the sham, SCI, LC and
LS groups. Red arrows indicate weight-supported stepping. (B) BBB
scores in the sham, SCI, LC and LS groups. (C) Electrophysiological
outcomes of the SCEP recordings at 8th week after SCI. Amplitude:
The magnitude of the voltage from each group of peaks to each group
of baselines. Latency: The period of time from the start of the
stimulation to the appearance of the peak. Quantification of the
(D) latency and (E) amplitude of the SCEP. *P<0.05
vs. SHAM; #P<0.05 vs. LC. RNAi, RNA interference;
NSCs, neural stem cells; SCI, spinal cord injury; BBB, Basso,
Beattie, Bresnahan; SCEP, spinal cord-evoked potential; SHAM, rats
without transfection of the spinal cord; SCI, rats with complete
transection of the spinal cord; LC, LV-control shRNA-transfected
NSCs were injected into rats with SCI; LS, LV-STAT3
shRNA-transfected NSCs were injected into rats with SCI. |
Transplantation of
STAT3-RNAi-transfected NSCs enhances tissue repair following
SCI
To further clarify if the transplantation of NSCs
transfected with STAT3 shRNA promotes the regeneration of damaged
tissues and nerves following SCI, H&E staining was performed to
investigate the degree of tissue repair in each group. The size of
the lesions was calculated in H&E-stained sections 8 weeks
after injury. In the sagittal plane, the LS group exhibited a
significantly smaller lesion area compared with that the LC group
(Fig. 4A and B). To further observe the survival of
neurons around the SCI area, Nissl staining was performed on the
spinal cord tissue of each group. At 3 mm from the center of the
SCI area, extensive loss of neurons was observed in the LC group.
By contrast, the LS group had significantly more neurons compared
with that in the LC group (Fig. 4A
and C). To explore the
interconnection of nerves in the injured area, a retrograde
tracking experiment with FG was performed. Tissues from the LS
group exhibited more neurons labeled by FG compared with those in
the LC group (Fig. 4A and D). Furthermore, at the T7 spinal cord
segment, FG-labeled neurons were more frequently observed in the LS
group compared with those in the LC group, indicating that the LS
group had more nerve connections on both ends of the lesion site
(Fig. 4A and D).
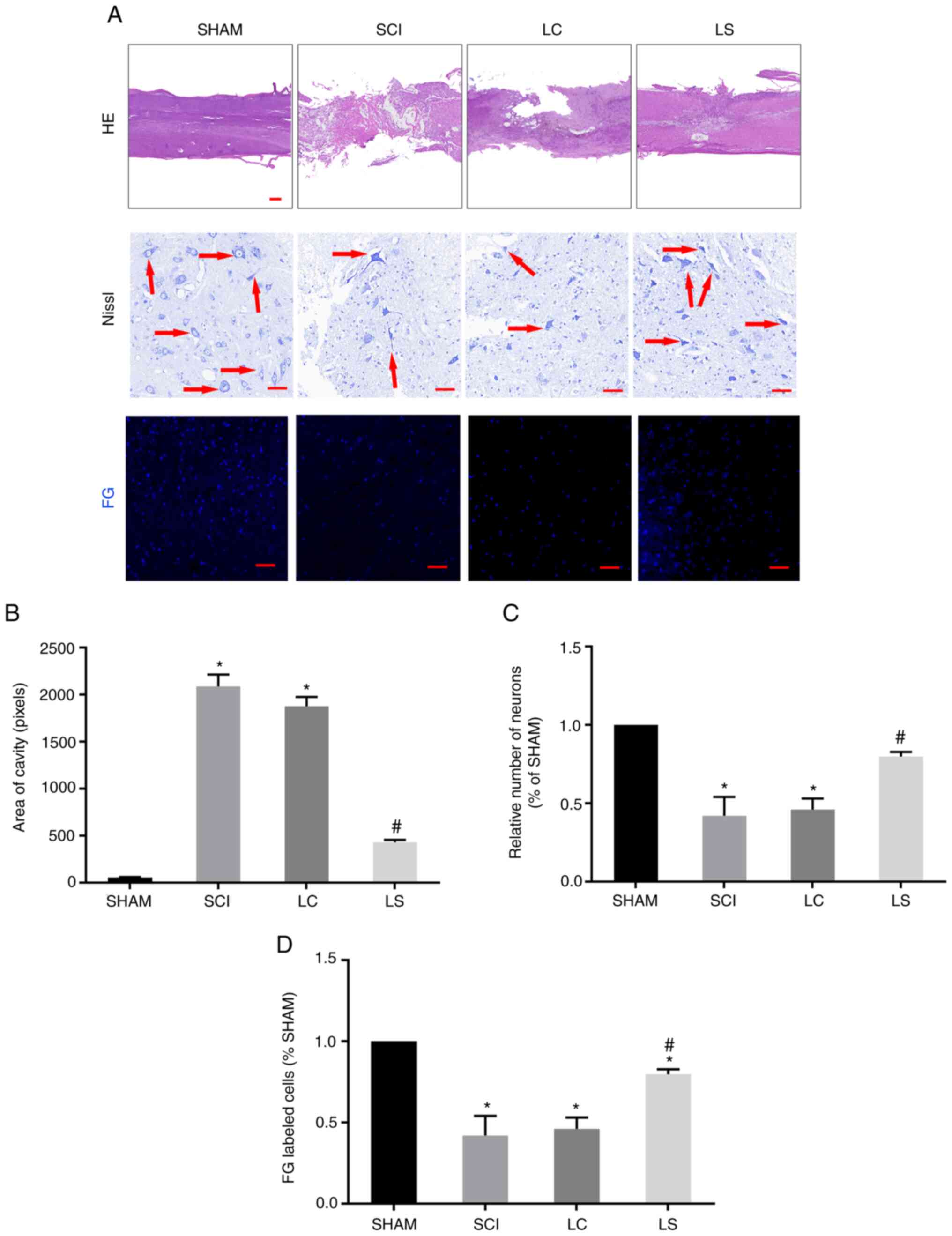 | Figure 4Transplantation of
STAT3-RNAi-transfected NSCs enhance tissue repair following SCI.
(A) Representative H&E staining micrographs showing cavity
formation in the sham, SCI, LC and LS groups 8 weeks after injury
(n=5). Images of surviving neurons 3 mm rostral to the injured
epicenter, as shown by Nissl staining. Red arrows point to
surviving neurons. FG-labeled neurons in the ventricolumna of the
T7 segment spinal cord. (B) Comparison of cavity area from part (A)
in each group. (C) Relative number of FG-labeled neurons in each
group, which was normalized to that in the sham group. (D)
Quantification analysis of the number of FG-labeled cells in each
group, which was normalized to that in the sham group.
*P<0.05 vs. sham, #P<0.05 vs. LC. Scale
bar, 100 µm. RNAi, RNA interference; NSCs, neural stem cells; SCI,
spinal cord injury; H&E, hematoxylin and eosin; FG, FluoroGold;
SHAM, rats without transfection of the spinal cord; SCI, rats with
complete transection of the spinal cord; LC, LV-control
shRNA-transfected NSCs were injected into rats with SCI; LS,
LV-STAT3 shRNA-transfected NSCs were injected into rats with
SCI. |
Targeted inhibition of STAT3 enhances
the neuronal differentiation of transplanted NSCs in spinal cord
lesions
The degree of differentiation in the grafted cells
was analyzed in the lesion site by immunofluorescence 8 weeks after
transplantation. In the LC and SCI group, the majority of cells
expressed astrocytic maker GFAP in the injured area, whilst only a
small number of cells expressed neuronal markers β3-tubulin and
MAP2 (Fig. 5A and B). However, in the LS group, the
percentage of β3-tubulin positive and MAP2 positive cells was
significantly higher compared with that in the LC and SCI groups,
whereas the percentage of GFAP positive cells was significantly
lower compared with that in the LC and SCI groups (Fig. 5A and B).
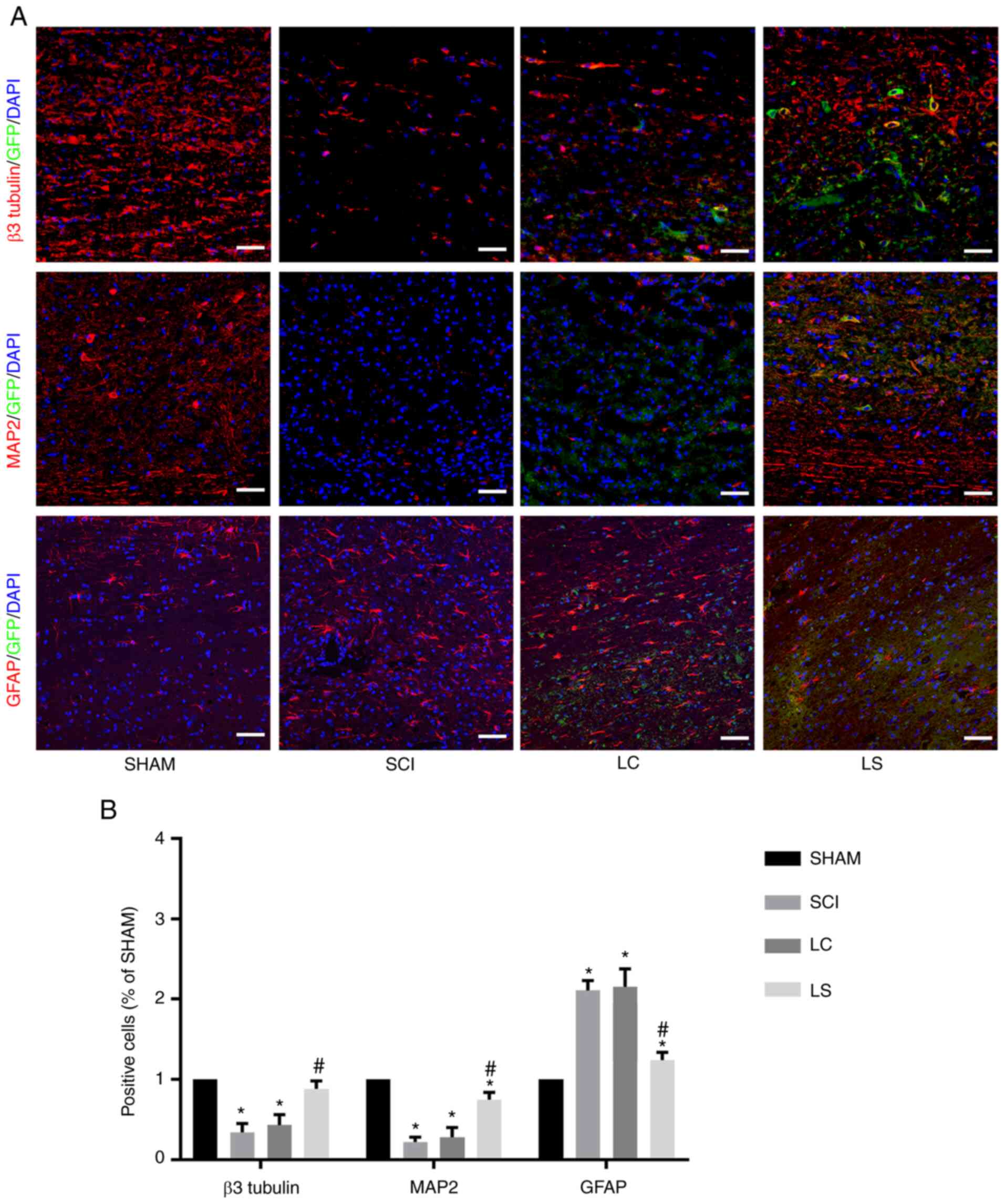 | Figure 5Targeted inhibition of STAT3 enhances
neuronal differentiation of transplanted NSCs in the spinal cord
lesions. (A) Immunostaining for β3-tubulin, MAP2 and GFAP (red),
GFP (green) and DAPI (blue) showing the differentiation of the
transplanted NSCs in the spinal cord lesions of each group. (B)
Quantification analysis of the number of β3-tubulin-, MAP2- and
GFAP-positive cells in each group, which was normalized to that in
the SHAM group. *P<0.05 vs. SHAM,
#P<0.05 vs. LC. Scale bar, 100 µm. NSCs, neural stem
cells; MAP2, microtubule-associated protein 2; GFAP, fibrillary
acidic protein; GFP, green fluorescent protein; SHAM, rats without
transfection of the spinal cord; SCI, rats with complete
transection of the spinal cord; LC, LV-control shRNA-transfected
NSCs were injected into rats with SCI; LS, LV-STAT3
shRNA-transfected NSCs were injected into rats with SCI. |
Discussion
It was shown in the present study that inhibiting
STAT3 not only promoted NSC differentiation into neurons, but also
inhibited their differentiation into astrocytes, potentially
through mTOR activation. In addition, it was found that the
transplantation of STAT3-RNAi-transfeced NSCs into rats following
SCI improved functional recovery, promoted axonal regeneration and
inhibited astrocyte differentiation after SCI.
Following SCI, the limited regenerative capacity of
the adult mammalian spinal cord has been attributed to the
formation of cavities and glial scars that interrupt the ascending
and descending pathways (26-28).
NSC transplantation is now considered to be a promising method for
treating SCI (29). However,
compared with NSCs in the brain, most NSCs in the spinal cord
differentiate into astrocytes, where no neurogenesis has been
observed following SCI (30). STAT3
is a member of the JAK-STAT signaling family (31), which transduces signals for a number
of cytokines and growth factors, including IL-6, ciliary
neurotrophic factor, leukemia inhibitory factor, EGF and
transforming growth factor α, which have been implicated as
triggers of reactive astrogliosis (32). STAT3 has also been found to be
expressed in NSCs, such that STAT3 is activated to promote NSC
proliferation but inhibit neurogenesis (33,34).
The binding of STAT3 to the GFAP promoter is essential for
astrocyte differentiation (35,36).
Blocking STAT3 has been reported to favor motor neurons whilst
inhibiting GFAP-positive astrocyte differentiation in NSCs in
vitro (14). In the present
study, STAT3 expression was inhibited in NSCs in vitro,
which led to increased neuronal differentiation and reduced
astrocyte differentiation at the same time. This result was
consistent with previous reports (14,33,34)
and once again demonstrated that inhibiting the expression of STAT3
promoted the differentiation of NSCs into neurons whilst
simultaneously inhibiting their differentiation into astrocytes. Of
note, increased activation of mTOR was observed when the STAT3
expression was suppressed. mTOR is a serine/threonine kinase of the
PI3K signaling pathway with two divergent complexes, mTOR complex 1
(mTORC1) and mTORC2(37). mTOR
serves a relevant role in the control of homeostasis of different
compartments containing stem cells, including NSCs (38). A number of studies have reported
that activation of the JAK/PI3K/Akt/mTOR signaling pathway promotes
neuronal differentiation in NSCs (39). However, one study also showed that
activating STAT3 and mTOR at the same time can promote
differentiation of NSC into glial cells (40). Easley et al (41) revealed that cells with high mTORC1
activity can severely alter dendrite formation and synaptic
integration. Although the downstream consequences of higher mTORC2
levels remain unclear, it is thought to be associated with
cytoskeletal remodeling (37). In
addition, the serine-threonine kinase FK506-binding
protein/mTOR/STAT3 pathway also serves a crucial role in glial
differentiation (42). Therefore,
the correlation between STAT3 and the mTOR signaling pathway during
NSC differentiation requires further investigation.
STAT3 is activated following SCI and plays a vital
role in the differentiation and organization of astrocytes
(43), which contribute to glial
scar formation (22,38,39).
In addition, according to previous studies, the microenvironment
surrounding the spinal cord lesion site following SCI result in the
differentiation of exogenous NSCs into astrocytes (28,44,45).
This leads to the formation of glial scars in the lesion site that
are widely regarded to inhibit axon regeneration and functional
recovery during the chronic phase of SCI (17,46,47).
Dai et al (24) reported
that STAT3-knockout mice exhibit less scar formation following SCI.
By contrast, STAT3 inhibitors have also been shown to reverse the
inhibitory effects of STAT3 on neuronal recovery (14). According to the present study, less
astrocytic differentiation was observed in STAT3-RNAi-transfected
NSCs, suggestive of reduced glial scar formation. In addition,
compared with that in control-RNAi-transfected NSCs, STAT3-RNAi-
transfected NSCs were more likely to differentiate into neurons and
exhibited more dendrite outgrowth. Neurons in the SCI area of the
LS group formed more connections with the upstream and downstream
neurons of the SCI area, which was demonstrated by the existence of
an increased number of FG-labeled neurons. Increased neuronal
connectivity is considered to be one of the conditions that promote
functional recovery following SCI (48). These results suggested that the
STAT3-RNAi-transfected NSCs produced highly developed neurons that
served a potentially useful role in neurogenesis, consistent with
the results of previous studies (14,23).
Wu et al (49) used
microRNA-15a to downregulate STAT3 in mice following SCI, following
which a superior functional recovery was observed. In the present
study, a more significant increase in the BBB scores of rats
transplanted with STAT3-RNAi- transfected NSCs was observed, with
an ~50% increase in the BBB scores in that group, compared with the
scores in the LC group 5 weeks following SCI. This result is
superior compared with that reported by previous studies that only
transplanted NSC (45,50,51).
Furthermore, rats transplanted with STAT3-RNAi-transfected NSCs
tended to have higher SCEP amplitudes compared with rats in the LC
and SCI groups, suggesting that motor and sensory axonal conduction
was reinforced following transplantation with STAT3-RNAi-
transfected NSCs (52). H&E
staining also confirmed that transplanting STAT3-RNAi-transfected
NSCs significantly reduced the lesion volume, potentially
contributing to functional recovery following SCI (53). However, although these results
indicated a potentially superior recovery following the
transplantation of STAT3-RNAi-transfected NSCs in SCI rats, the
specific mechanism underlying these phenomena require further
investigation. There are possible risks to this methodology,
including the low survival rate of transplanted cells (54) and the risk of the transplanted
undifferentiated stem cells developing into teratomas (55). Therefore, additional experiments are
required to further confirm and clarify the safety aspects before
it can be translated into other species.
In conclusion, in the present study, in vivo
and in vitro evidence of the role of STAT3 as a negative
regulator of neuronal differentiation in NSCs was provided. The
transplantation of STAT3-inhibited NSCs appears to be a promising
strategy for enhancing the benefit of NSC-mediated regenerative
cell therapy for SCI, where discovery of a prominent role of STAT3
in NSC biology can enhance the understanding of NSC
differentiation.
Acknowledgements
Not applicable.
Funding
Funding: The present study was supported by the Medical
Scientific Research Foundation of Guangdong Province of China
(grant no. A2017207), Science and Technology Promotion Program of
Air Force Medical Center, PLA (grant no. 2020KTA01) and Natural
Science Foundation of Guangdong Province (grant no.
2020A1515010306).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
LW, YW and SL designed and supervised the study. ZP
analyzed the data. TL, XZ and JD conducted the study, collected the
data and wrote the manuscript. SC and KZ collected and analyzed the
data. All authors have read and approved the final manuscript. LW
and TL confirm the authenticity of all the raw data.
Ethics approval and consent to
participate
All experimental animal procedures were approved by
the Care and Use Committee of Sun Yat-sen University (approval no.
SYXX2012-0081) and performed following the Guide to the Care and
Use of Experimental Animals provided by the National Research
Council (1996, USA).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Fan B, Wei Z, Yao X, Shi G, Cheng X, Zhou
X, Zhou H, Ning G, Kong X and Feng S: Microenvironment imbalance of
spinal cord injury. Cell Transplant. 27:853–866. 2018.PubMed/NCBI View Article : Google Scholar
|
|
2
|
McDonald JW and Sadowsky C: Spinal-Cord
injury. Lancet. 359:417–425. 2002.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Dominguez E, Rivat C, Pommier B, Mauborgne
A and Pohl M: JAK/STAT3 pathway is activated in spinal cord
microglia after peripheral nerve injury and contributes to
neuropathic pain development in rat. J Neurochem. 107:50–60.
2008.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Shibata N, Kakita A, Takahashi H, Ihara Y,
Nobukuni K, Fujimura H, Sakoda S, Sasaki S, Iwata M, Morikawa S, et
al: Activation of signal transducer and activator of
transcription-3 in the spinal cord of sporadic amyotrophic lateral
sclerosis patients. Neurodegener Dis. 6:118–126. 2009.PubMed/NCBI View Article : Google Scholar
|
|
5
|
LeComte MD, Shimada IS, Sherwin C and
Spees JL: Notch1-STAT3-ETBR signaling axis controls reactive
astrocyte proliferation after brain injury. Proc Natl Acad Sci USA.
112:8726–8731. 2015.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Okada S, Nakamura M, Katoh H, Miyao T,
Shimazaki T, Ishii K, Yamane J, Yoshimura A, Iwamoto Y, Toyama Y
and Okano H: Conditional ablation of stat3 or socs3 discloses a
dual role for reactive astrocytes after spinal cord injury. Nat
Med. 12:829–834. 2006.PubMed/NCBI View
Article : Google Scholar
|
|
7
|
Tsuda M, Kohro Y, Yano T, Tsujikawa T,
Kitano J, Tozaki-Saitoh H, Koyanagi S, Ohdo S, Ji RR, Salter MW and
Inoue K: JAK-STAT3 pathway regulates spinal astrocyte proliferation
and neuropathic pain maintenance in rats. Brain. 134:1127–1139.
2011.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Park KW, Lin CY, Benveniste EN and Lee YS:
Mitochondrial STAT3 is negatively regulated by SOCS3 and
upregulated after spinal cord injury. Exp Neurol. 284:98–105.
2016.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Gu F, Hata R, Ma YJ, Tanaka J, Mitsuda N,
Kumon Y, Hanakawa Y, Hashimoto K, Nakajima K and Sakanaka M:
Suppression of stat3 promotes neurogenesis in cultured neural stem
cells. J Neurosci Res. 81:163–171. 2005.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Cao F, Hata R, Zhu P, Nakashiro Ki and
Sakanaka M: Conditional deletion of stat3 promotes neurogenesis and
inhibits astrogliogenesis in neural stem cells. Biochem Biophys Res
Commun. 394:843–847. 2010.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Gong Z, Xia K, Xu A, Yu C, Wang C, Zhu J,
Huang X, Chen Q, Li F and Liang C: Stem cell transplantation: A
promising therapy for spinal cord injury. Curr Stem Cell Res Ther.
15:321–331. 2019.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Assinck P, Duncan GJ, Hilton BJ, Plemel JR
and Tetzlaff W: Cell transplantation therapy for spinal cord
injury. Nat Neurosci. 20:637–647. 2017.PubMed/NCBI View
Article : Google Scholar
|
|
13
|
Muheremu AJ, Peng J and Ao Q: Stem cell
based therapies for spinal cord injury. Tissue Cell. 48:328–333.
2016.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Natarajan R, Singal V, Benes R, Gao J,
Chan H, Chen H, Yu Y, Zhou J and Wu P: STAT3 modulation to enhance
motor neuron differentiation in human neural stem cells. PLoS One.
9(e100405)2014.PubMed/NCBI View Article : Google Scholar
|
|
15
|
White CW III, Fan X, Maynard JC, Wheatley
EG, Bieri G, Couthouis J, Burlingame AL and Villeda SA: Age-Related
loss of neural stem cell O-GlcNAc promotes a glial fate switch
through STAT3 activation. Proc Natl Acad Sci USA. 117:22214–22224.
2020.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Lizee G, Aerts JL, Gonzales MI, Chinnasamy
N, Morgan RA and Topalian SL: Real-Time quantitative reverse
transcriptase-polymerase chain reaction as a method for determining
lentiviral vector titers and measuring transgene expression. Hum
Gene Ther. 14:497–507. 2003.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Chen N, Cen JS, Wang J, Qin G, Long L,
Wang L, Wei F, Xiang Q, Deng DY and Wan Y: Targeted inhibition of
leucine-rich repeat and immunoglobulin domain-containing protein 1
in transplanted neural stem cells promotes neuronal differentiation
and functional recovery in rats subjected to spinal cord injury.
Crit Care Med. 44:e146–e157. 2016.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Bloomsmith MA, Perlman JE, Hutchinson E
and Sharpless M: Behavioral management programs to promote
laboratory animal welfare, In: Management of Animal Care and Use
Programs in Research, Education, and Testing. 2nd edition.
Weichbrod RH, Thompson GAH and Norton JN (eds). CRC Press/Taylor
& Francis, Boca Raton, FL, pp63-82, 2018.
|
|
20
|
Peng Z, Li X, Fu M, Zhu K, Long L, Zhao X,
Chen Q, Deng DY and Wan Y: Inhibition of notch1 signaling promotes
neuronal differentiation and improves functional recovery in spinal
cord injury through suppressing the activation of ras homolog
family member A. J Neurochem. 150:709–722. 2019.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Basso DM, Beattie MS and Bresnahan JC: A
sensitive and reliable locomotor rating scale for open field
testing in rats. J Neurotrauma. 12:1–21. 1995.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Zhao X, Peng Z, Long L, Chen N, Zheng H,
Deng DY and Wan Y: Lentiviral vector delivery of short hairpin RNA
to NgR1 promotes nerve regeneration and locomotor recovery in
injured rat spinal cord. Sci Rep. 8(5447)2018.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Cheng X, Yeung PK, Zhong K, Zilundu PL,
Zhou L and Chung SK: Astrocytic endothelin-1 overexpression
promotes neural progenitor cells proliferation and differentiation
into astrocytes via the Jak2/Stat3 pathway after stroke. J
Neuroinflammation. 16(227)2019.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Dai J, Xu LJ, Han GD, Sun HL, Zhu GT,
Jiang HT, Yu GY and Tang XM: MicroRNA-125b promotes the
regeneration and repair of spinal cord injury through regulation of
JAK/STAT pathway. Eur Rev Med Pharmacol Sci. 22:582–589.
2018.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Lee DY: Roles of mTOR signaling in brain
development. Exp Neurobiol. 24:177–185. 2015.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Pfyffer D, Huber E, Sutter R, Curt A and
Freund P: Tissue bridges predict recovery after traumatic and
ischemic thoracic spinal cord injury. Neurology. 93:e1550–e1560.
2019.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Zhang T, Liu C and Chi L: Suppression of
miR-10a-5p in bone marrow mesenchymal stem cells enhances the
therapeutic effect on spinal cord injury via BDNF. Neurosci.
714(134562)2020.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Qian D, Li L, Rong Y, Liu W, Wang Q, Zhou
Z, Gu C, Huang Y, Zhao X, Chen J, et al: Blocking notch signal
pathway suppresses the activation of neurotoxic A1 astrocytes after
spinal cord injury. Cell Cycle. 18:3010–3029. 2019.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Mothe AJ and Tator CH: Review of
transplantation of neural stem/progenitor cells for spinal cord
injury. Int J Dev Neurosci. 31:701–713. 2013.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Namiki J and Tator CH: Cell proliferation
and nestin expression in the ependyma of the adult rat spinal cord
after injury. J Neuropathol Exp Neurol. 58:489–498. 1999.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Decker T and Kovarik P: Transcription
factor activity of STAT proteins: Structural requirements and
regulation by phosphorylation and interacting proteins. Cell Mol
Life Sci. 55:1535–1546. 1999.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Herrmann JE, Imura T, Song B, Qi J, Ao Y,
Nguyen TK, Korsak RA, Takeda K, Akira S and Sofroniew MV: STAT3 is
a critical regulator of astrogliosis and scar formation after
spinal cord injury. J Neurosci. 28:7231–7243. 2008.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Ohta S, Misawa A, Fukaya R, Inoue S,
Kanemura Y, Okano H, Kawakami Y and Toda M: Macrophage migration
inhibitory factor (MIF) promotes cell survival and proliferation of
neural stem/progenitor cells. J Cell Sci. 125:3210–3220.
2012.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Kong X, Gong Z, Zhang L, Sun X, Ou Z, Xu
B, Huang J, Long D, He X, Lin X, et al: JAK2/STAT3 signaling
mediates IL-6-inhibited neurogenesis of neural stem cells through
DNA demethylation/methylation. Brain Behav Immun. 79:159–173.
2019.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Cheng PY, Lin YP, Chen YL, Lee YC, Tai CC,
Wang YT, Chen YJ, Kao CF and Yu J: Interplay between SIN3A and
STAT3 mediates chromatin conformational changes and GFAP expression
during cellular differentiation. PLoS One. 6(e22018)2011.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Takizawa T, Nakashima K, Namihira M,
Ochiai W, Uemura A, Yanagisawa M, Fujita N, Nakao M and Taga T: DNA
methylation is a critical cell-intrinsic determinant of astrocyte
differentiation in the fetal brain. Dev Cell. 1:749–758.
2001.PubMed/NCBI View Article : Google Scholar
|
|
37
|
LiCausi F and Hartman NW: Role of mTOR
complexes in neurogenesis. Int J Mol Sci. 19(1544)2018.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Russell RC, Fang C and Guan KL: An
emerging role for TOR signaling in mammalian tissue and stem cell
physiology. Development. 138:3343–3356. 2011.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Lee JE, Lim MS, Park JH, Park CH and Koh
HC: S6K promotes dopaminergic neuronal differentiation through
PI3K/Akt/mTOR-dependent signaling pathways in human neural stem
cells. Mol Neurobiol. 53:3771–3782. 2016.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Wang B, Xiao Z, Chen B, Han J, Gao Y,
Zhang J, Zhao W, Wang X and Dai J: Nogo-66 promotes the
differentiation of neural progenitors into astroglial lineage cells
through mTOR-STAT3 pathway. PLoS One. 3(e1856)2008.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Easley CA IV, Ben-Yehudah A, Redinger CJ,
Oliver SL, Varum ST, Eisinger VM, Carlisle DL, Donovan PJ and
Schatten GP: mTOR-Mediated activation of p70 S6K induces
differentiation of pluripotent human embryonic stem cells. Cell
Reprogram. 12:263–273. 2010.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Rajan P, Panchision DM, Newell LF and
McKay RD: BMPs signal alternately through a SMAD or FRAP-STAT
pathway to regulate fate choice in CNS stem cells. J Cell Biol.
161:911–921. 2003.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Wanner IB, Anderson MA, Song B, Levine J,
Fernandez A, Gray-Thompson Z, Ao Y and Sofroniew MV: Glial scar
borders are formed by newly proliferated, elongated astrocytes that
interact to corral inflammatory and fibrotic cells via
STAT3-dependent mechanisms after spinal cord injury. J Neurosci.
33:12870–12886. 2013.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Kim C, Kim HJ, Lee H, Lee H, Lee SJ, Lee
ST, Yang SR and Chung CK: Mesenchymal stem cell transplantation
promotes functional recovery through MMP2/STAT3 related
astrogliosis after spinal cord injury. Int J Stem Cells.
12:331–339. 2019.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Hackett AR, Lee DH, Dawood A, Rodriguez M,
Funk L, Tsoulfas P and Lee JK: STAT3 and SOCS3 regulate NG2 cell
proliferation and differentiation after contusive spinal cord
injury. Neurobiol Dis. 89:10–22. 2016.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Hosseini SM, Sani M, Haider KH, Dorvash M,
Ziaee SM, Karimi A and Namavar MR: Concomitant use of mesenchymal
stem cells and neural stem cells for treatment of spinal cord
injury: A combo cell therapy approach. Neurosci Lett. 668:138–146.
2018.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Lopez-Serrano C, Torres-Espín A, Hernández
J, Alvarez-Palomo AB, Requena J, Gasull X, Edel MJ and Navarro X:
Effects of the post-spinal cord injury microenvironment on the
differentiation capacity of human neural stem cells derived from
induced pluripotent stem cells. Cell Transplant. 25:1833–1852.
2016.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Squair JW, West CR, Popok D, Assinck P,
Liu J, Tetzlaff W and Krassioukov AV: High thoracic contusion model
for the investigation of cardiovascular function after spinal cord
injury. J Neurotrauma. 34:671–684. 2017.PubMed/NCBI View Article : Google Scholar
|
|
49
|
Wu WD, Wang LH, Wei NX, Kong DH, Shao G,
Zhang SR and Du YS: MicroRNA-15a inhibits inflammatory response and
apoptosis after spinal cord injury via targeting STAT3. Eur Rev Med
Pharmacol Sci. 23:9189–9198. 2019.PubMed/NCBI View Article : Google Scholar
|
|
50
|
van Gorp S, Leerink M, Kakinohana O,
Platoshyn O, Santucci C, Galik J, Joosten EA, Hruska-Plochan M,
Goldberg D, Marsala S, et al: Amelioration of motor/sensory
dysfunction and spasticity in a rat model of acute lumbar spinal
cord injury by human neural stem cell transplantation. Stem Cell
Res Ther. 4(57)2013.PubMed/NCBI View Article : Google Scholar
|
|
51
|
Zhao XM, He XY, Liu J, Xu Y, Xu FF, Tan
YX, Zhang ZB and Wang TH: Neural stem cell transplantation improves
locomotor function in spinal cord transection rats associated with
nerve regeneration and IGF-1 R expression. Cell Transplant.
28:1197–1211. 2019.PubMed/NCBI View Article : Google Scholar
|
|
52
|
Winkler T, Sharma HS, Gordh T, Badgaiyan
RD, Stålberg E and Westman J: Topical application of dynorphin A
(1-17) antiserum attenuates trauma induced alterations in spinal
cord evoked potentials, microvascular permeability disturbances,
edema formation and cell injury: An experimental study in the rat
using electrophysiological and morphological approaches. Amino
Acids. 23:273–281. 2002.PubMed/NCBI View Article : Google Scholar
|
|
53
|
Ramadan WS, Abdel-Hamid GA, Al-Karim S and
Abbas AT: Histological, immunohistochemical and ultrastructural
study of secondary compressed spinal cord injury in a rat model.
Folia Histochem Cytobiol. 55:11–20. 2017.PubMed/NCBI View Article : Google Scholar
|
|
54
|
Pereira IM, Marote A, Salgado AJ and Silva
NA: Filling the gap: Neural stem cells as a promising therapy for
spinal cord injury. Pharmaceuticals (Basel). 29(65)2019.PubMed/NCBI View Article : Google Scholar
|
|
55
|
Trounson A and McDonald C: Stem cell
therapies in clinical trials: Progress and challenges. Cell Stem
Cell. 17:11–22. 2015.PubMed/NCBI View Article : Google Scholar
|