Introduction
Articular cartilage has complex biomechanical
characteristics and high durability (1). However, due to its limited repair
activity, irreversible damage to its structure and function can
result from external injuries or natural degeneration. Currently,
clinical treatment methods for articular cartilage injury include
micro-fracture, autologous chondrocyte transplantation and
cartilage transplantation (2).
However, long-term therapeutic effects are not ideal due to
challenges with the application, such as difficulty in obtaining
materials and etc. (3). Bone marrow
mesenchymal stem cells (BMSCs) have multi-directional
differentiation potential and have been widely used as ideal seed
cells in bone tissue engineering (4,5).
Currently, studies on factors promoting differentiation mainly
involve cytokines, intermediate molecules of cartilage
differentiation signaling pathways and non-coding RNAs, with
satisfactory results being obtained regarding the differentiation
of BMSCs into chondrocytes (6,7).
Bcl-xL is a transmembrane molecule in the
mitochondria, which belongs to the Bcl-2 family. The classical
anti-apoptotic pathway of Bcl-xL plays a role in chondrogenesis and
differentiation (8). Nakagami et
al (9) applied angiotensin II
to promote cartilage healing in a fracture model and found that the
expression of Bcl-xL mRNA increased significantly. Moreover, Bcl-xL
is upregulated in chondrosarcoma with abnormal chondrocyte
proliferation (10). A previous
study revealed that in addition to apoptosis-related effects,
Bcl-xL also functions in an independent pro-metabolic process known
as the non-classical Bcl-xL pathway. In this pathway Bcl-xL does
not bind to Bax; rather, it directly dissociates from the
mitochondrial membrane, enters the nucleus and exerts its role in
mediating cellular metabolism by regulating corresponding cytokines
(11). Compared with the wild-type
Bcl-xL, the Bcl-xL mutant (where the GRI sequence is replaced by
ELN) has no anti-apoptotic effect; however, its other biological
functions remain unchanged (12).
Our previous study found that the expression of TGF-β3 in BMSCs
transfected with Bcl-xL mutant expression vectors was significantly
increased (13,14). TGF-β3 is a member of the TGF-β
superfamily and plays a significant role in promoting chondrogenic
differentiation (13,14). Members of the bone morphogenetic
protein (BMP) family also promote cartilage repair (15). BMP-2 enables for the migration and
aggregation of mesenchymal stem cells into clusters, maintains them
in a tight state, stimulates Smad phosphorylation, enhances Sox9
expression and promotes mesenchymal stem cell differentiation into
chondrocytes (16). BMP-7
phosphorylates SMAD1 and SMAD5, and induces the transcription of a
variety of osteoblastic and chondrogenic genes (17).
The present study investigated whether Bcl-xL
promotes chondrogenic differentiation of BMSCs in the
microenvironment of articular cartilage damage through its dual
roles in anti-apoptosis and TGF-β/BMP upregulation. The aim was to
provide a potential improvement to the application of bone tissue
engineering for the treatment of articular cartilage defects.
Materials and methods
Isolation and identification of
BMSCs
From January 1st 2018 to September 30th 2018, bone
marrow samples from 20 patients (healthy volunteers with lower limb
fractures; age, 18-60 years; 10 males and 10 females) were
collected from Pu Ai Hospital affiliated to Tongji Medical College,
Huazhong University of Science and Technology. All operations were
approved by the Ethics Review Committee of Pu'ai Hospital
affiliated with Tongji Medical College of Huazhong University of
Science and Technology, and all patients signed written informed
consent to participate in this study and for their samples to be
used for subsequent experiments. After skin preparation,
disinfection and toweling in the patient's crotch area, 15 ml of
bone marrow solution was obtained and treated with 15 ml of
lymphocyte separation solution (cat. no. XY08001T; Shanghai Xinyu
Biological Technology Co., Ltd.). The solution was centrifuged at
400 x g for 15 min at 4˚C, resuspended in PBS and centrifuged again
using the same conditions. After the supernatant was discarded, the
cells were suspended in a complete medium (DMEM/F12 (cat. no.
SH30023.01; HyClone; GE Healthcare Life Sciences) containing 10%
FBS (cat. no. 10270-106; Gibco; Thermo Fisher Scientific, Inc.) and
1% penicillin and streptomycin (cat. no. PAB180056; Bioswamp Wuhan
Beinle Biotechnology Co., Ltd.) and the cell concentration was
adjusted to 1x106 cells/ml. The cells were then cultured
in a polylysine-coated culture plate in an atmosphere containing 5%
CO2 and 95% air, at 37˚C for 48 h. The medium was
replaced every 72 h and the cells were subcultured or cryopreserved
when the confluence reached 80-90%. BMSCs were identified using
light microscopy (200x magnification) and flow cytometry (CD34,
CD45, CD73, CD90 and CD105) as previously described (18,19).
Vector construction and
transfection
The Bcl-xL sequence (NM 138578.1) was obtained from
the National Center for Biotechnology Information database. Since
cDNA cannot enter the cell directly without a corresponding vector
(20), the gene fragment was
introduced into BMSCs to make it stable and abundantly expressed.
The cDNA of the human Bcl-xL gene was inserted into the
pcDNA3.1-Bcl-xL vector (Addgene, Inc.) with XhoI and
NheI restriction sites. The digested PCR gene fragments and
linearized vector were ligated at 16˚C overnight and the resulting
Bcl-xL overexpression vectors were transformed into competent DH5α
cells. Target plasmids were extracted from the bacterial liquid
according to the manufacturer's instructions. The transfection
efficiency was determined using reverse transcription-quantitative
PCR (RT-qPCR) as subsequently described.
The cDNA sequence corresponding to the GRI amino
acid sequence of Bcl-xL at positions 138-140 was replaced with a
sequence encoding ELN using the QuickChange Lighting Site-Directed
Mutagenesis kit (Agilent Technologies, Inc.). The pcDNA3.1-ΔBcl-xL
plasmid was constructed using pcDNA3.1-Bcl-xL as a template and
following the experimental procedures used in the construction of
the parent vector.
The following Bax and Bak primer sequences were
used: Bax forwards,
5'-GGATGCTCTGAGCAGATCATGAAGATTCAAGAGATCTTCATGATCTGCTCAGAGCTTTTTTCC-3'
and reverse,
5'-GAATAAAAAAGCTCTGAGCAGATCATGAAGATCTCTTGAATCTTCATGATCTGCTCAGAGCTC-3';
Bak forwards,
5'-GGATGGCAGAGAATGCCTATGAGTATTCAAGAGATACTCATAGGCATTCTCTGCCTTTTTTCC-3'
and reverse,
5'-GAATAAAAAAGGCAGAGAATGCCTATGAGTATCTCTTGAATACTCATAGGCATTCTCTGCCTC-3'.
The cDNA for the human Bax and Bak genes were inserted into the
pSuper-Bax and pSuper-Bak vectors (Addgene, Inc.), respectively,
using XhoI and BamHI restriction sites. The target
plasmids were extracted from the bacterial liquid and transfected
using the Lipofectamine 2000 reagent according to the
manufacturer's instructions.
The BMSCs were divided into four groups: Control
(not subjected to transfection), EV (empty pcDNA3.1-Bcl-xL vector),
OV (Bcl-xL overexpression) and ∆OV (∆Bcl-xL overexpression).
Saffron and toluidine blue
staining
The cells (7.5x105 per group) were
resuspended in a 15-ml centrifuge tube and centrifuged at 150 x g
for 5 min at 4˚C. The supernatant was aspirated and the cells were
resuspended in the complete cartilage differentiation induction
medium (RASMX-9004; Cyagen Biosciences, Inc.). The cells were
centrifuged at 150 x g for 5 min at 4˚C and incubated at 37˚C in 5%
CO2. After two weeks of continuous induction, the
resultant cartilage was fixed with 10% formalin at 4˚C for 48 h,
subjected to saffron O staining (cat. no. PAB180084; Wuhan Beinlai
Biotechnology Co., Ltd.) for 2 min at 60˚C and then observed under
a light microscope (200x magnification).
Cartilage slices were incubated at 65˚C for 1 h and
placed in xylene (15 min, 4˚C) and a concentration gradient of
alcohol (5 min, 4˚C). After two washes with double-distilled water
for 2 min each, the slices were placed in toluidine blue staining
solution (cat. no. G3668; Beijing Solarbio Science & Technology
Co., Ltd.) for 30 min. The cells were then washed with
double-distilled water, sealed with neutral gum and observed under
a light microscope (MD1000; Leica Microsystems, Inc.) to detect the
integrated optical density values in each group.
Flow cytometry
The cells (1x106 per group) were
resuspended in 100 µl of flow buffer (cat. no. PAB180076; Bioswamp
Wuhan Beinle Biotechnology Co., Ltd.) in an Eppendorf tube and 2 µl
of CD45-FITC (cat. no. 11-9459-42, eBioscience; Thermo Fisher
Scientific, Inc.), CD34-FITC (cat. no. CD34-581-01; Invitrogen;
Thermo Fisher Scientific, Inc.), CD73-FITC (cat. no. 11-0739-42r;
eBioscience; Thermo Fisher Scientific, Inc.), CD90-FITC (cat. no.
11-0903-82; eBioscience; Thermo Fisher Scientific, Inc.) or
CD105-FITC (cat. no. MA1-19594; Invitrogen; Thermo Fisher
Scientific, Inc.) was added. The cells were incubated in the dark
for 45 min at 4˚C, following which, 400 µl of flow cytometry dyeing
buffer (cat. no. PAB180076; Wuhan Beinlai Biotechnology Co., Ltd.)
was added to each tube. The cells were subjected to flow cytometry
(CytoFLEX S; Beckman Coulter, Inc.) and the results were analyzed
using the CYEXPERT software (CXP Analysis 2.0; Beckman Coulter,
Inc.).
Cells were cultured for 24 h at 37˚C, harvested,
treated with 1 ml of pre-cooled PBS and centrifuged at 1,000 x g
for 5 min at 4˚C. Subsequently, 10 µl of Annexin V-FITC and 10 µl
of PI were added. The cell samples were then analyzed using flow
cytometry as aforementioned. A one-step fluorescence compensation
strategy was used to eliminate interference with the FITC channel
(21).
RT-qPCR
Total RNA was extracted from 1x106 cells
using Trizol® reagent (according to the manufacturer's
procedures), and cDNA was synthesized using a Reverse Transcriptase
kit (Takara Bio, Inc.). qPCR was performed using a real-time PCR
system (Bio-Rad Laboratories, Inc.) using the SYBR Green PCR kit
(cat. no. KM4101; Kapa Biosystems; Roche Diagnostics). Each qPCR
reaction was performed in duplicate: 95˚C for 3 min; followed by 39
cycles of 95˚C for 5 sec, 56˚C for 10 sec, 72˚C for 25 sec; 65˚C
for 5 sec and 95˚C for 50 sec for final extension, using GAPDH as a
housekeeping gene. The results were analyzed using the
2-∆∆Cq method (22). The
primers were designed and configured by Nanjing Kingsy
Biotechnology Co., Ltd. (Table
I).
 | Table IPrimer sequences. |
Table I
Primer sequences.
| Primer | Sequence (5'→3') |
|---|
| Bcl-xL-F |
GCCACTTACCTGAATGACC |
| Bcl-xL-R | TGAGCCCAGCAGAACC |
| TGF-β-F |
ATTCCTGGCGATACCTCA |
| TGF-β-R | GGCGAAAGCCCTCAAT |
| BMP2-F |
TGACGAGGTCCTGAGCG |
| BMP2-R |
CCTGAGTGCCTGCGATA |
| GAPDH-F |
CCACTCCTCCACCTTTG |
| GAPDH-R |
CACCACCCTGTTGCTGT |
Western blotting
The protein (20 µg) extracts prepared by cell lysate
(cat. no. PAB180006; Bioswamp Wuhan Beinle Biotechnology Co., Ltd.)
from BMSCs which had been cultured for 24 h, and the concentration
was measured by BCA protein assay kit (cat. no. PAB180007; Bioswamp
Wuhan Beinle Biotechnology Co., Ltd.). Total protein was separated
by 12% sodium dodecyl sulfate-polyacrylamide gel electrophoresis
and transferred to PVDF membranes (EMD Millipore). The membranes
were blocked with 5% milk in Tris-buffered saline (pH 7.6)
containing 0.1% Tween-20 for 2 h at 24˚C. Subsequently, they were
incubated overnight at 4˚C with specific primary antibodies against
TGF-β1 (1:1,000; cat. no. ab92486; Abcam), BMP2 (1:1,000; cat. no.
ab14933; Abcam) and GAPDH (1:1,000; cat. no. 2118; Cell Signaling
Technologies, Inc.). After three washes with PBS/Tween 20, the
membranes were incubated with a horseradish peroxidase-conjugated
secondary goat anti-rabbit IgG (1:10,000; cat. no. PAB150011; Wuhan
Beinlai Biotechnology Co., Ltd.) for 2 h at 4˚C. Protein bands were
visualized using ECL color detection (Tanon-5200; Tanon Science and
Technology Co., Ltd.) and analyzed using the AlphaEase FC gel image
analysis software (version 4.2; Tanon Science and Technology Co.,
Ltd.).
Statistical analysis
Data are expressed as the mean ± SD (n=3). To
analyze the differences between groups, data comparisons were
performed using one-way ANOVAs and subsequent Tukey's post-hoc
tests. P<0.05 was considered a statistically significant
difference.
Results
Isolation and identification of
BMSCs
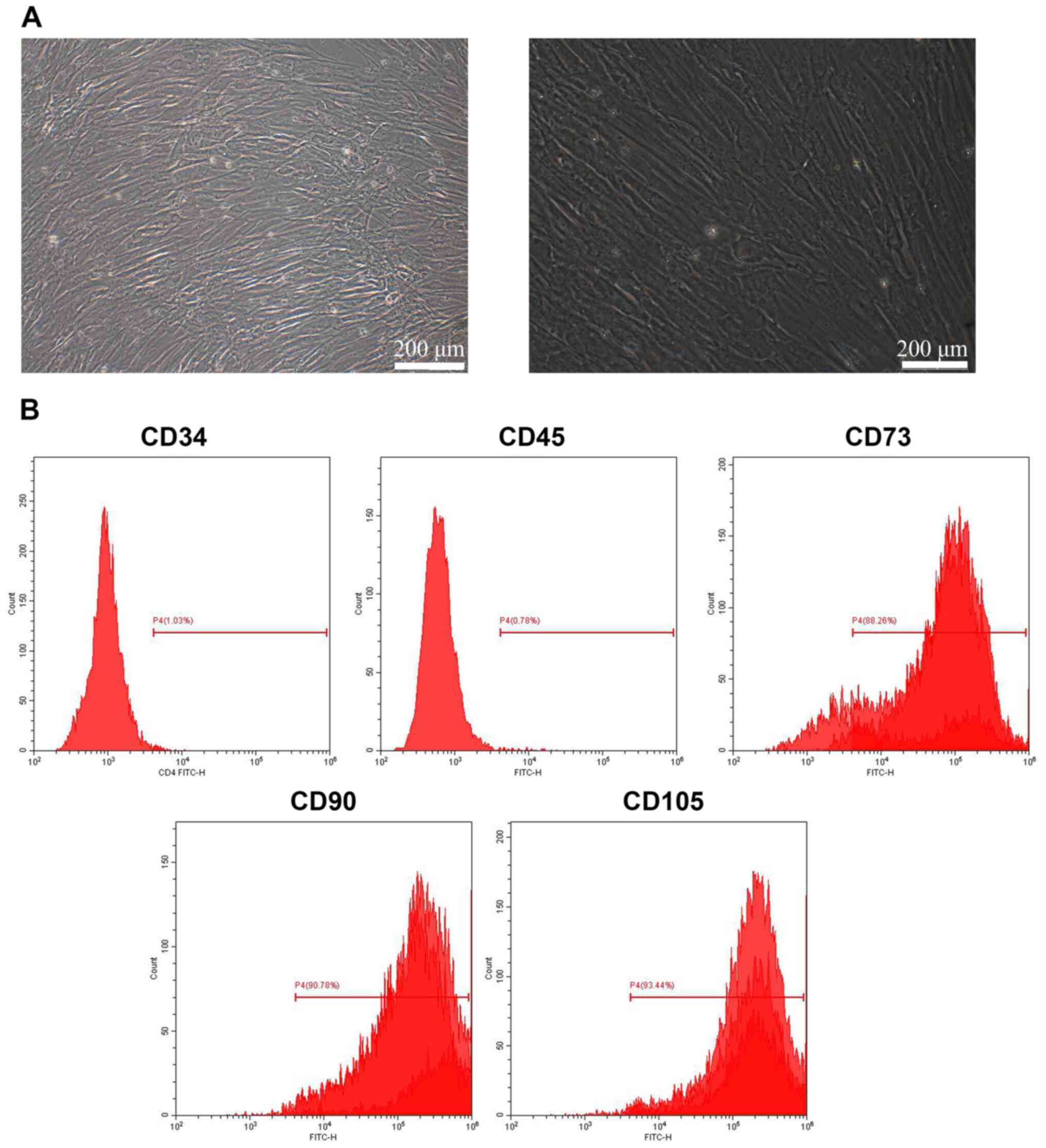
The morphology of BMSCs was observed under a
microscope. As shown in Fig. 1A,
BMSCs were morphologically consistent and arranged as ordered
fibroblast-like cells. The expression of BMSC surface markers was
further evaluated (Fig. 1B) and it
was found that CD34 and CD45 were negatively expressed (1.03 and
0.78%), while CD73, CD90 and CD105 showed positive expression
(88.26, 90.78 and 93.44%, respectively), suggesting that the BMSCs
were successfully isolated.
Transfection efficiency of the
overexpression and interference vector
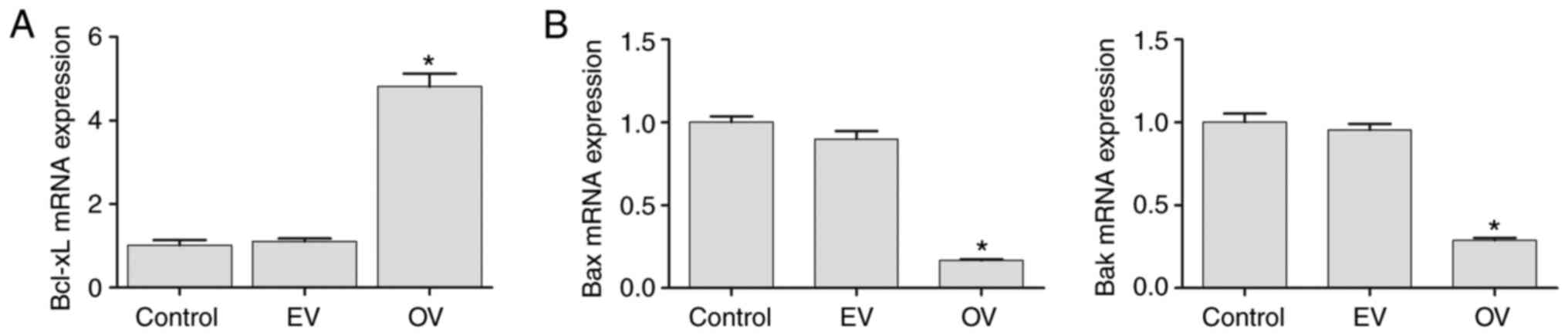
The expression of Bcl-xL mRNA in the control, EV and
OV groups was assessed to confirm the transfection efficiency of
the Bcl-xL overexpression vector. Compared with the control and EV
groups, expression of Bcl-xL in the OV group was significantly
increased (P<0.05) (Fig. 2A). As
shown in Fig. 2B, the expression of
Bax and Bak in the interference groups was significantly decreased
compared with the control group and EV group (P<0.05), implying
that the respective interference vectors were transfected
successfully (Fig. 2).
Effect of the Bcl-xL mutant on BMSC
apoptosis and cartilage differentiation
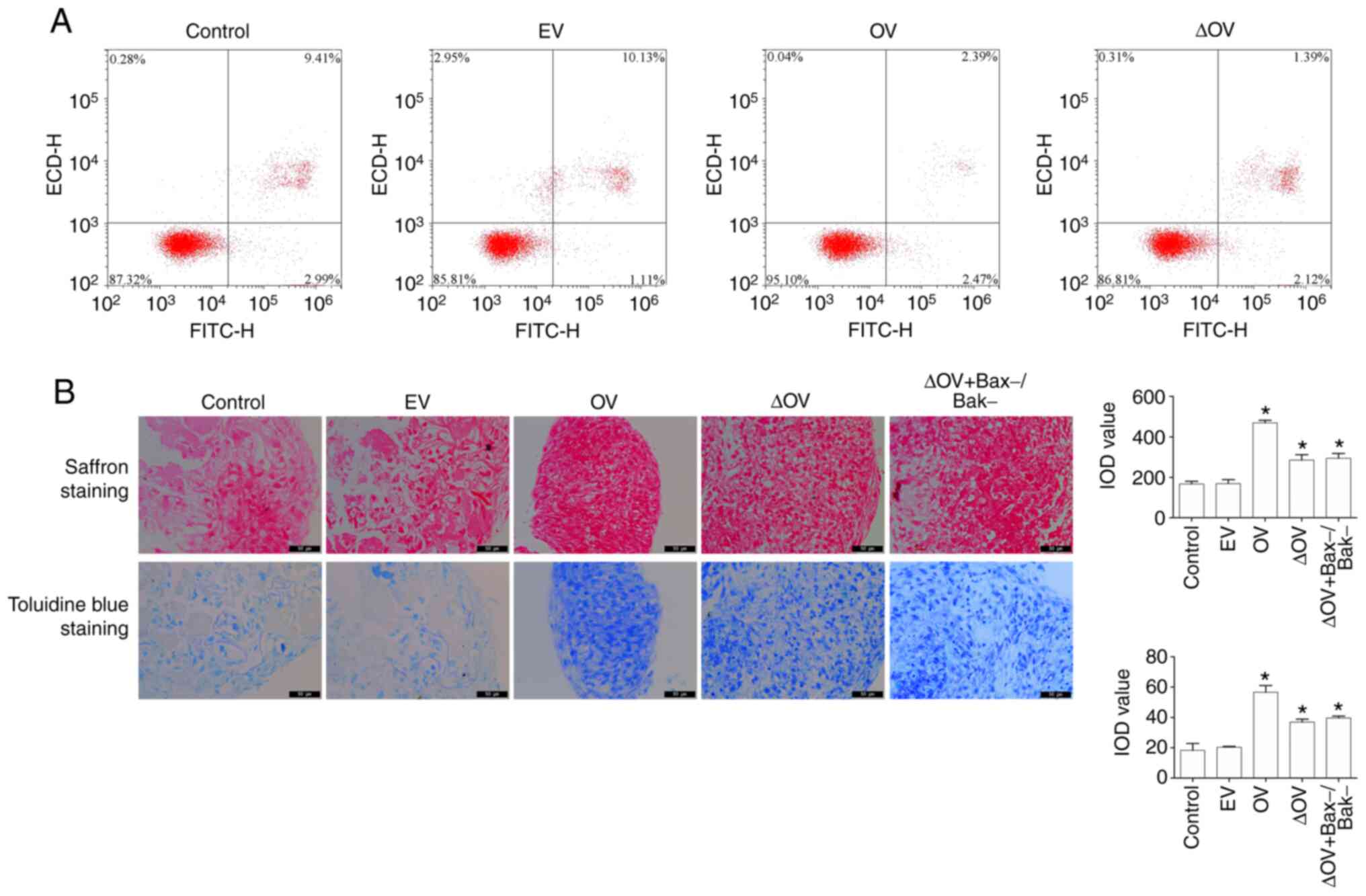
To investigate the effect of Bcl-xL mutant on
apoptosis and cartilage differentiation in BMSCs, flow cytometry,
toluidine blue staining and saffron staining were used to detect
the apoptotic rate. As shown in Fig.
3A, the rate of apoptosis in the OV group was significantly
decreased (P<0.05) compared with that in the control group,
while that of the ∆OV group did not change significantly
(P>0.05). Evaluation of cartilage differentiation showed that
the staining area of the OV and ∆OV groups increased (P<0.05)
compared with that of the control group (Fig. 3B). To further investigate whether
promotion of cartilage differentiation in the Bcl-xL mutant was
dependent on the expression of Bax/Bak, plasmids capable of
silencing Bax/Bak were constructed to create Bax-/Bak-cells.
Compared with that in the control group, the area of ∆OV and
∆OV+Bax-/Bak-staining increased (P<0.05), while there was no
significant difference in cartilage formation (indicated by saffron
and toluidine blue staining) between ∆OV and ∆OV+Bax-/Bak-
(P>0.05) (Fig. 3B). These
observations suggested that the Bcl-xL mutant promoted the
differentiation of BMSCs into cartilage without affecting BMSC
apoptosis and that the effect of promoting cartilage
differentiation of BMSCs is not dependent on Bax-/Bak-.
Effect of the Bcl-xL mutant on the
expression of TGF-β and BMP
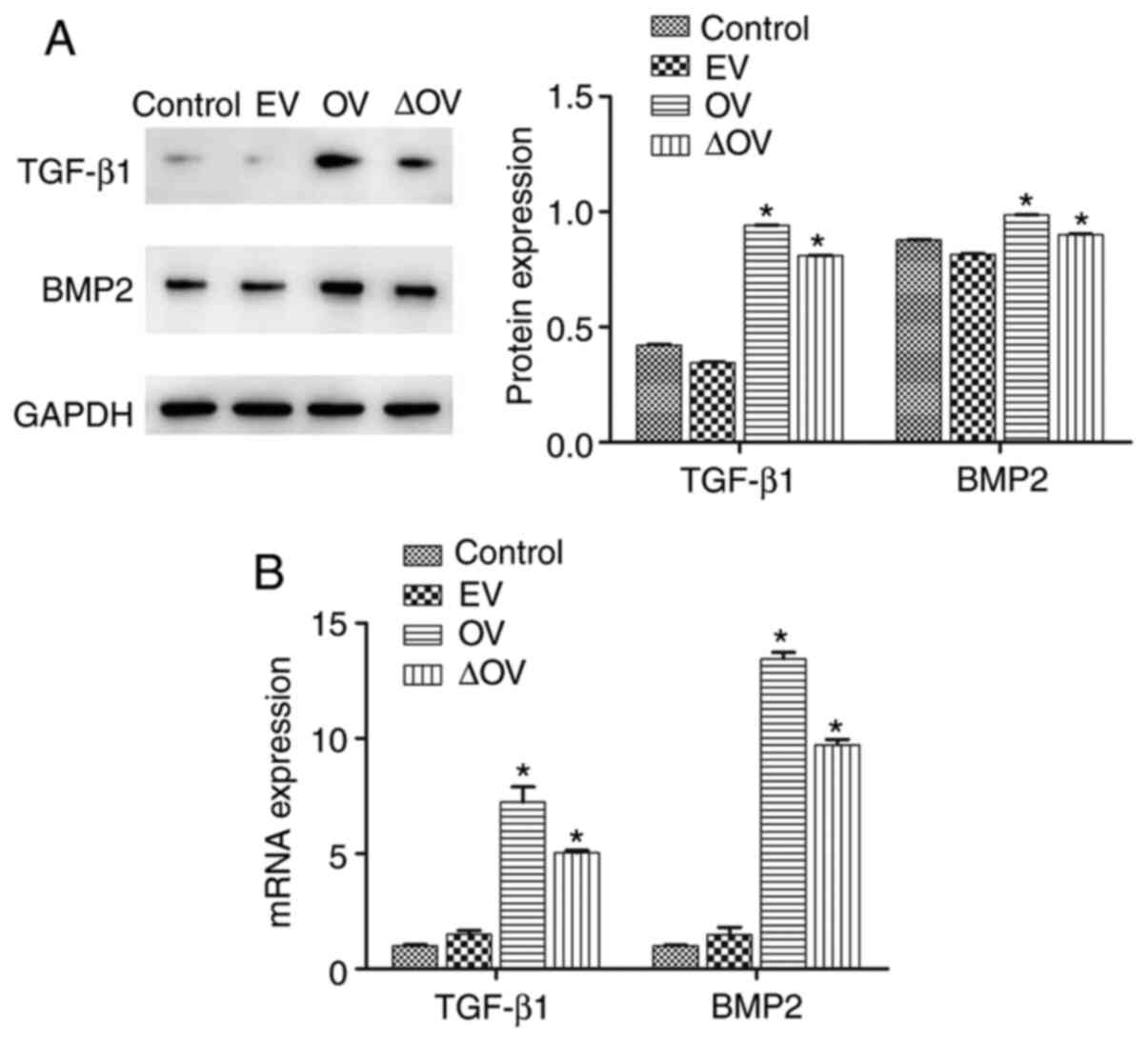
Western blotting and RT-qPCR were conducted to
assess the expression of TGF-β1 and BMP2. Fig. 4A and B show that OV and ∆OV induced higher
protein and mRNA expression levels of TGF-β1 and BMP2 than those
observed in the control and EV groups (P<0.05). Compared with
the OV group, the expression of TGF-β1 and BMP2 in ΔOV group was
significantly decreased (P<0.05), suggesting that Bcl-xL mutant
improved the expression of TGF-β1 and BMP2.
Discussion
Previous studies have shown that BMSCs are suitable
for clinical applications owing to their straightforward isolation
and differentiation from a variety of tissues (23,24).
In addition, BMSCs are considered to be good vectors for
cell-mediated gene therapy, as they are relatively easy to handle
in vitro (25). At present,
most clinical methods for repairing articular cartilage injury are
invasive; therefore, the development and application of new methods
employing more conservative approaches are urgently needed. Direct
intra-articular injection of BMSCs is a potentially conservative
cell therapy for the repair of cartilage defects (26). There are two primary methods for
applying BMSCs in treatment: One utilizes a suitable cell matrix as
a carrier for the implantation of an in vitro stem cell
scaffold, while the other involves the in vivo
differentiation of BMSCs by co-culturing with target cells. Jin
(27) demonstrated the feasibility
and high efficiency of autologous exostromal BMSC scaffolds in
cartilage tissue engineering, with both in vivo and in
vitro experiments. Hu (28)
also confirmed that BMSC and chondrocyte aggregation co-cultures
improved cartilage repair in a study on rabbit knee articular
cartilage defect. However, the targeted cartilage differentiation
of BMSCs is dependent on the local microenvironment and the
synergistic effects of various inducing factors (29). The effective induction of BMSC
differentiation into chondrocytes and initiation of cartilage
tissue formation has become a challenging process that needs to be
addressed.
Bcl-xL is an important anti-apoptotic molecule in
the Bcl-2 family that inhibits the pro-apoptotic molecule, Bax,
preventing the apoptosis caused by the release of cytochrome C
(30). Bcl-xL also blocks apoptosis
by inhibiting the binding of Apaf-1 and caspase-9 downstream of Bax
activation (31), as well as by
inhibiting apoptosis induced by the Fas-FasL pathway (32). Therefore, Bcl-xL exerts important
anti-apoptotic effects by blocking various apoptotic pathways
stimulated by pro-apoptotic factors, such as hypoxia and
inflammation, thereby demonstrating its role in promoting
chondrogenic differentiation (33).
Wang et al (34) reported an
increase in cartilage formation during fracture repair in mice with
Bax gene deletion, suggesting that Bcl-xL continues to promote
cartilage production and repair during chondrogenesis, even in the
absence of the anti-apoptotic effect of Bcl-xL (presumably through
alternative pathways). Mutants are defined as individuals with
mutations showing phenotypes that differ from the wild type. A
previous study have shown that the Bcl-xL mutant, where the GRI
amino acid sequence at positions 138-140 is replaced by ELN, has no
anti-apoptotic effect, while its other biological functions remain
similar to those of wild-type Bcl-xL (12).
The present study examined the anti-apoptotic effect
of the Bcl-xL mutant on cartilage differentiation of BMSCs. it was
demonstrated that the Bcl-xL mutant promoted cartilage
differentiation of BMSCs without affecting BMSC apoptosis, whereby
this effect was not related to the expression of Bax and Bak. The
potential mechanism of action of the Bcl-xL mutant on promoting
cartilage differentiation of BMSCs was further examined. The
results showed that the expression of TGF-β and BMP increased
significantly after Bcl-xL and Bcl-xL mutant intervention,
revealing that the Bcl-xL mutant may promote cartilage
differentiation of BMSCs by upregulating TGF-β/BMP expression
levels.
A limitation of the present study was that the
differentiation experiments were performed after a long term
culture. BMSCs can differentiate to chondrocytes within two weeks.
As such, analyzing the effects of overexpression of Bcl-xL
longitudinally, examining the expression profiles of this protein
in the various groups to show the kinetics of expression, is
necessary. However, the main aim of the present study was to
observe the effect of the Bcl-xL mutant on cartilage
differentiation and the expression of TGF-β and BMP. As such,
further studies should examine the effects of Bcl-xL in the various
BMCS treatment groups in subsequent experiments.
In conclusion, the present study demonstrated that
Bcl-xL mutants promoted cartilage differentiation of BMSCs and
upregulated TGF-β/BMP expression levels, whereby this enhancement
of chondrogenic differentiation was not related to the expression
of Bax and Bak.
Acknowledgements
Not applicable.
Funding
Funding: This work was supported by financial grants from the
Wuhan Applied Foundational Frontier Project (grant no.
2020020601012309); Science and Technology Department of Hubei
Province (grant no. 2020CFB784); and Hubei Province health and
family planning scientific research project (grant no.
WJ2019Q006).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
KX, LY and WX contributed to the conception of the
study. KX and XG designed and performed the experiments, analyzed
the data and wrote the manuscript. RH and MX analyzed the data and
provided technical support. KX and LY confirm the authenticity of
all the raw data. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
All operations were approved by the Ethics Review
Committee of Pu'ai Hospital affiliated with Tongji Medical College
of Huazhong University of Science and Technology [approval no.
(2017)IEC(S118)]. Informed consent was obtained from all
patients.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Sandell LJ and Aigner T: Articular
cartilage and changes in arthritis: Cell biology of osteoarthritis.
Arthritis Res. 3:107–113. 2001.PubMed/NCBI View
Article : Google Scholar
|
|
2
|
Buckwalter JA and Mankin : Articular
cartilage: Tissue design and chondrocyte-matrix interactions. Instr
Course Lect. 47:477–486. 1998.PubMed/NCBI
|
|
3
|
Natoli RM and Athanasiou KA: Traumatic
loading of articular cartilage: Mechanical and biological responses
and post-injury treatment. Biorheology. 46:451–485. 2009.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Wang DH, Lin YF, Chen L, Mo YQ, Huang P
and Ma RX: Guided bone regeneration using a bone tissue engineering
complex consisting of a poly-dl-lactide membrane and bone
mesenchymal stem cells. Oncotarget. 9:16380–16388. 2017.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Dang M, Saunders L, Niu XF, Fan YB and Ma
PX: Biomimetic delivery of signals for bone tissue engineering.
Bone Res. 6(25)2018.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Ding WW, Huang JH and Xu WL: Research
progress on the differentiation of bone marrow mesenchymal stem
cells into chondrocytes. Med Rev. 12:2148–2151. 2015.
|
|
7
|
Yang K, Xie L, Zhu WM, Huang JH, Duan L
and Wang DP: Progress in the mechanism of microRNA regulating
chondrogenic differentiation of mesenchymal stem cells.
Orthopedics. 7:219–221. 2016.
|
|
8
|
Cheng EH, Wei MC, Weiler S, Flavell RA,
Mak TW, Lindsten T and Korsmeyer SJ: Bcl-2, Bcl-X(L) sequester BH3
domain-only molecules preventing Bax- and Bak-mediated
mitochondrial apoptosis. Mol Cell. 8:705–711. 2001.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Nakagami H, Kiomy Osako M, Shimizu H,
Hanayama R and Morishita R: Potential contribution of action of
renin angiotensin system to bone metabolism. Curr Hypertens Rev.
3:129–132. 2007.
|
|
10
|
Chen CB, Zhou H, Wei F, Jiang L, Liu XG,
Liu ZJ and Ma Q: Increased levels of hypoxia-inducible factor-1α
are associated with Bcl-xL expression, tumor apoptosis, and
clinical outcome in chondrosarcoma. J Orthop Res. 29:143–151.
2011.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Ming LH, Wang P, Bank A, Yu J and Zhang L:
PUMA dissociates Bax and Bcl-xl to induce apoptosis in colon cancer
cells. J Biol Chem. 281:16034–16042. 2006.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Cong YM, Liu P, Jin D, Guang WJ and Ma YH:
Relationship between Bcl-2 family and apoptosis. Chin J Med Res.
2008.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Lee HL, Yu B, Deng P, Wang CY and Hong C:
Transforming growth factor-β-induced KDM4B promotes chondrogenic
differentiation of human mesenchymal stem cells. Stem Cells.
34:711–719. 2016.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Jin EJ, Park JH, Lee SY, Chun JS, Bang OS
and Kang SS: Wnt-5a is involved in TGF-beta3-stimulated
chondrogenic differentiation of chick wing bud mesenchymal cells.
Int J Biochem Cell Biol. 38:183–195. 2006.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Rueger DC and Chubinskaya S: Bone
morphogenetic proteins in articular cartilage repair. Prog Inflamm
Res. 109–132. 2004.
|
|
16
|
Zhao L, Li G and Zhou GQ: SOX9 directly
binds CREB as a novel synergism with the PKA pathway in
BMP-2-induced osteochondrogenic differentiation. J Bone Miner Res.
24:826–836. 2009.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Retting KN, Song B, Yoon BS and Lyons KM:
BMP canonical Smad signaling through Smad1 and Smad5 is required
for endochondral bone formation. Development. 136:1093–1104.
2009.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Mahmood SC, Michael B, Angela M, John P
and David H: Cryopreservation of whole adipose tissue for future
use in regenerative medicine. J Surg Res. 187:24–35.
2014.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Sierra-Sánchez Á, Ordóñez-Luque A,
Espinosa-Ibáñez O, Ruiz-García A and Arias-Santiago S: Epithelial
in vitro differentiation of mesenchymal stem cells. Curr Stem Cell
Res Ther. 13:409–422. 2018.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Feng Z, Chen Q, Ren MQ, Tian ZG and Gong
YP: CD40L inhibits cell growth of THP-1 cells by suppressing the
PI3K/Akt pathway. Onco Targets Ther. 12:3011–3017. 2019.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Bayer J, Grunwald D, Lambert C, Mayol JF
and Maynadié M: Thematic workshop on fluorescence compensation
settings in multicolor flow cytometry. Cytometry B Clin Cytom.
72:8–13. 2007.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Tsuchiya A, Kojima Y, Ikarashi S, Seino S,
Watanabe Y, Kawata Y and Terai S: Clinical trials using mesenchymal
stem cells in liver diseases and inflammatory bowel diseases.
Inflamm Regen. 37(16)2017.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Antoniou KM, Karagiannis K, Tsitoura E,
Bibaki E, Lasithiotaki I, Proklou A, Spandidos DA and Tzanakis N:
Clinical applications of mesenchymal stem cells in chronic lung
diseases. Biomed Rep. 8:314–318. 2018.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Gafni Y, Turgeman G, Liebergal M, Pelled
G, Gazit Z and Gazit D: Stem cells as vehicles for orthopedic gene
therapy. Gene Ther. 11:417–426. 2006.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Djouad F, Mrugala D, Noël D and Jorgensen
C: Engineered mesenchymal stem cells for cartilage repair. Regen
Med. 1:529–537. 2006.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Jin CZ: Application of autologous bone
marrow mesenchymal stem cell matrix scaffold in cartilage tissue
engineering. CRTER. 33:7864–7866. 2015.
|
|
28
|
Hu B: Bone marrow mesenchymal stem cells
and chondrocytes aggregation and co-culture to repair osteochondral
defects in rabbit knee joint. SCU 2016.
|
|
29
|
Novak T, Seelbinder B, Twitchell CM,
Voytik-Harbin Prof SL and Prof CPN: Dissociated and reconstituted
cartilage microparticles in densified collagen induce local hMSC
differentiation. Adv Funct Mater. 26:5427–5436. 2016.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Eskes R, Desagher S, Antonsson B and
Martinou JC: Bid induces the oligomerization and insertion of Bax
into the outer mitochondrial membrane. Mol Cell Biol. 20:929–935.
2000.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Rosśe T, Olivier R, Monney L, Rager M,
Conus S, Fellay I, Jansen B and Borner C: Bcl-2 prolongs cell
survival after Bax-induced release of cytochrome C. Nature.
391:496–499. 1998.PubMed/NCBI View
Article : Google Scholar
|
|
32
|
Biswas RS, Cha HJ, Hardwick JM and
Srivastava RK: Inhibition of drug-induced Fas ligand transcription
and apoptosis by Bcl-XL. Mol Cell Biochem. 225:7–20.
2001.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Zhou W, Wu SM, Chen HJ and Pu SX:
Expression of Bcl-x mRNA after hypoxic-ischemic brain injury in
neonatal rats. J Clin Neurol. 13:7–9. 2000.
|
|
34
|
Wang CY, Chen LL, Kuo PY, Chang JL, Wang
YJ and Hung SC: Apoptosis in chondrogenesis of human mesenchymal
stem cells: Effect of serum and medium supplements. Apoptosis.
15:439–449. 2010.PubMed/NCBI View Article : Google Scholar
|