Introduction
Bronchopulmonary dysplasia (BPD), a serious
complication commonly occurring in immature infants, is
characterized by arrested alveolarization of the lungs, oxygen
toxicity and pulmonary injury (1,2).
Studies have indicated that BPD may lead to lung injury,
neurodevelopmental sequelae and even disability, resulting in a
high economic burden on families (3-5).
Hyperoxemia is the major cause of BPD, which is able to exacerbate
the damage of alveolar epithelium and endothelial cells (6). Due to perinatal infection and
inflammation, immature breathing and oxidative stress, preterm
infants are more susceptible to hyperoxia-induced BPD (7-9).
In rat models, hyperoxia induction may cause alveolar septation
suppression and terminal air space enhancement, which is similar to
observations in human BPD (10,11).
Hu et al (12) confirmed
that microRNA-29a (miRNA/miR-29a) inhibitor was able to relieve
hyperoxia-stimulated BPD in neonatal mice through the upregulation
of GRB2-associated binding protein 1. At present, effective
postnatal treatment strategies for BPD are limited and the major
therapeutic methods are mechanical ventilation and drug therapy. In
addition, the early identification by nursing staff and active
perinatal care in infants with low birth weight and BPD may reduce
patient deterioration and improve patient survival (13). Therefore, identifying and seeking
effective treatment for BPD is urgent.
Long non-coding RNAs (lncRNAs), a class of
non-coding RNA of >200 nucleotides in length, have been
indicated to be associated with various biological processes,
including cell proliferation, apoptosis and genomic stability
(14). RNA imprinted and
accumulated in nucleus (Rian) is an lncRNA with a parent-specific
expression, which is located on mouse chromosome 12 and includes
three long transcripts: Irm, Meg8 and AB063319 (15,16). A
large number of studies have revealed that Rian is a vital
regulator of multiple biological processes (15,17,18).
For instance, Yao et al (17) indicated that the upregulation of
lncRNA Rian suppressed cell apoptosis from cerebral
ischemia-reperfusion injury through Rian/miR-144-3p/GATA binding
protein 3 signaling.
miRNAs are short non-coding RNAs of 20-22
nucleotides in length, which control gene expression and may also
be involved in cancer progression (19). Considerable evidence has proven that
miRNAs have vital roles in the pathogenesis of diseases and are
considered potential targets for disease therapies. Dupont et
al (20) revealed the roles of
miRNAs in the transmission of obesity-related metabolic diseases
induced by paternal diet. However, the latent mechanisms of miRNAs
acting in human diseases remain to be further explored. miR-421 has
been confirmed to be upregulated in multiple diseases, including in
BPD in mice (21). miR-421 has been
indicated to promote the occurrence of osteosarcoma by regulating
the expression of MCP induced protein 1(22). Besides, previous studies have
revealed that miR-421 regulates cell growth, migration and
apoptosis in several types of cancer by sponging different genes,
including ATM serine/threonine kinase and forkhead box O4 (23,24).
Moreover, a study has shown that miR-421 is highly expressed in BPD
mice, and miR-421 inhibition decreases bronchopulmonary dysplasia
in a mouse model by targeting Fgf10(25). However, the exact functions of
miR-421 in BPD have not yet been fully elucidated.
In a bioinformatics analysis, a direct binding site
was identified between Rian and miR-421. It was therefore
hypothesized that Rian may have an important role in BPD by
regulating miR-421. The results of the present study suggested that
Rian was downregulated in lung tissues of a rat model of BPD and
that Rian-overexpression vector relieved BPD and reduced
inflammatory response in BPD rats. In addition, Rian-plasmid was
indicated to protect alveolar epithelial cells from
hyperoxia-induced damage. However, all of these effects were
reversed by miR-421 mimics. The present study suggested that
Rian/miR-421 may serve as a novel biomarker for the diagnosis of
BPD and as a target for treatment.
Materials and methods
Animals and BPD model
establishment
A total of 60 mice were provided by the Experimental
Animal Center of Shanghai and kept under standard conditions
(22±1˚C, 55±5% humidity, 12-h light/dark cycle) with free access to
food and water. All animal operations were in accordance with the
National Institutes of Health Guide for the Care and Use of
Laboratory Animals. In addition, the present study was approved by
the Animal Ethics Committee of the Experimental Animal Center of
The First People's Hospital of Lianyungang (Lianyungang,
China).
The establishment of the mouse model of BPD was
performed as previously described (26). In brief, the neonatal mice (age,
<12 h) were stimulated with hyperoxia (85% O2) or
kept in normal room air (21% O2) for 7 days. The mice
were randomly divided into six groups (n=10): The control, model,
model + control-plasmid (empty vector pcDNA3.1), model +
Rian-plasmid (Rian sequence was synthesized based on the Rian
sequence and then sub-cloned into the pcDNA3.1 vector; Shanghai
GeneChem Co., Ltd.), model + Rian-plasmid + mimics control
(5'-UUCUCCGAACGUGUCACGUTT-3'; RiboBio Co, Ltd.) and the model +
Rian-plasmid + miR-421 mimics (5'-AUCAACAGACAUUAAUUGGGCGC-3';
RiboBio Co, Ltd.) group. Mice in the control-plasmid, Rian-plasmid,
Rian-plasmid + mimics control and Rian-plasmid + miR-421 mimics
groups were intraperitoneally injected once every other day with
plasmid/mimics on postpartum days 2-14. Saline (0.9% NaCl) was
injected into control and model group mice. On postpartum day 14,
the mice were anesthetized with sodium pentobarbital (50 mg/kg;
i.p.) and then sacrificed by cervical dislocation (death was
verified by cardiac and respiratory arrest) for lung tissue and
blood collection.
Mean linear intercept (MLI) and radial alveolar
count (RAC) were calculated in reference to a previous study
(4). Furthermore, the lung
weight/body weight (LW/BW) ratio of mice was calculated as
previously described (4).
Cell culture and treatment
The MLE-12 cells were purchased from the American
Type Culture Collection and grown in Hites medium (Cytiva) with 15%
FBS (Gibco; Thermo Fisher Scientific, Inc.) and 100 U/ml penicillin
and streptomycin (Thermo Fisher Scientific, Inc.). The cells were
cultivated in an incubator with 5% CO2 at 37˚C. The
MLE-12 cells were then incubated in 12-well plates
(5x104 cells per well) for 24 h and stimulated with
Hites medium supplemented with 0.1% FBS for 6 h. Next, the MLE-12
cells were exposed to room air or hyperoxia (85% oxygen) for 6 h
and treated with or without plasmid/mimics for 24 h.
Reverse transcription-quantitative
(RT-q)PCR analysis
The isolation of RNA from the lung tissue of mice
with BPD and MLE-12 cells was performed using TRIzol®
reagent (Thermo Fisher Scientific, Inc.) following the
manufacturer's protocol. Total RNA was then reverse transcribed
into cDNA by using the HiScript™ II qRT SuperMix (Vazyme
Biotech Co., Ltd.). RT conditions were as follows: 25˚C for 5 min,
42˚C for 60 min and 80˚C for 2 min. The expression of Rian and
miR-421 was detected using SYBR Premix Ex-Taq (Takara Bio, Inc.).
The thermocycling conditions were as follows: Initial denaturation
at 95˚C for 5 min; 38 cycles of denaturation at 95˚C for 10 sec,
annealing at 60˚C for 20 sec and extension at 72˚C for 30 sec.
Primers were obtained from Sangon Biotech Co., Ltd. as follows:
GAPDH, forward, 5'-CTTTGGTATCGTGGAAGGACTC-3' and reverse,
5'-GTAGAGGCAGGGATGATGTTCT-3'; U6, forward,
5'-GCTTCGGCAGCACATATACTAAAAT-3' and reverse,
5'-CGCTTCACGAATTTGCGTGTCAT-3'; lncRNA Rian, forward,
5'-CTGTTGTGCCCTCCCTGGATG-3' and reverse,
5'-CCAGCTAGGCTGTGTAAATCATC-3'; miR-421, forward,
5'-GTCGCGCGGGUUAAUGCCTC-3' and reverse,
5'-GGACATUAGUUGUCUGUAAATAG-3'; TNF-α, forward,
5'-CCGGGAGAAGAGGGATAGCTT-3' and reverse,
5'-TCGGACAGTCACTCACCAAGT-3'; IL-6, forward:
5'-TAGTCCTTCCTACCCCAATTTCC-3' and reverse,
5'-TTGGTCCTTAGCCACTCCTTC-3'; IL-1β, forward:
5'-GAAATGCCACCTTTTGACAGTG-3' and reverse, CTGGATGCTCTCATCAGGACA-3'.
The reactions were run in triplicate using the ABI PRISM 7900
sequence detection system (Thermo Fisher Scientific, Inc.). Target
gene expression was calculated with the 2-ΔΔCq method
(27).
Dual-luciferase reporter assay
Bioinformatics software (StarBase version 2.0;
http://starbase.sysu.edu.cn/starbase2/index.php) was
used to predict potential target genes of Rian using the
‘miRNA-lncRNA’ functions as described previously (28). The binding sites between miR-421 and
Rian were observed. The Rian-3'untranslated region (UTR), which
contains the miR-421 binding site, or the mutated (MUT) target
site, was synthesized via genomic PCR (29) from total RNA preps extracted from
MLE-12 cells and cloned into pMIR vectors (Thermo Fisher
Scientific, Inc.) to construct the reporter vector Rian-wild-type
(Rian-WT) or Rian-MUT-type (Rian-MUT). To point-mutate the miR-421
binding domain in the 3'UTR of Rian, a QuikChange Site-Directed
Mutagenesis kit (Stratagene; Agilent Technologies, Inc.) was used
according to the manufacturer's instructions. The 293T cells
(American Tissue Culture Collection) were transfected with Rian WT
or MUT plasmid combined with miR-421 mimics or mimics control using
Lipofectamine® 2000 (Thermo Fisher Scientific, Inc.)
following the manufacturer's protocol. The luciferase activity was
determined using a Dual-Luciferase Reporter Assay system (Promega
Corp.) and normalized to Renilla activity.
ELISA
Inflammatory factors (TNF-α, IL-6 and IL-1β) in the
supernatant of hyperoxia-induced MLE-12 cells were evaluated using
ELISA kits (cat. nos. PT512, PI326 and PI301; BioLegend, Inc.)
following the manufacturer's protocol. The optical density (OD)
value in each well was detected at 450 nm following the
manufacturer's protocol of the ELISA kits.
Cell transfection
The lncRNA sequence for Rian was synthesized based
on the Rian sequence and then sub-cloned into the pcDNA3.1 vector
(Shanghai GeneChem Co., Ltd.) to obtain Rian-plasmid. The empty
pcDNA3.1 vector was used as a control (control-plasmid; Shanghai
GeneChem Co., Ltd.). MLE-12 cells were cultured in room air or
under hyperoxia for 6 h. Subsequently, 100 ng control-plasmid, 100
ng Rian-plasmid, 100 nM mimics control
(5'-UUCUCCGAACGUGUCACGUTT-3'; RiboBio Co, Ltd.) or 100 nM miR-421
mimics (5'-AUCAACAGACAUUAAUUGGGCGC-3'; RiboBio Co, Ltd.) were
transfected into MLE-12 cells using Lipofectamine® 2000
(Thermo Fisher Scientific, Inc.) for 24 h, following the
manufacturer's protocol. At 24 h after cell transfection,
subsequent experimentation was performed and the overexpression
efficiency was determined using RT-qPCR.
MTT assay
Following transfection for 24 h, MLE-12 cells were
seeded into 96-well plates (1x104 cells/well; 100 µl
suspension/well) and incubated for 24 h at 37˚C. Cells were then
exposed to MTT (10 µl of 5 mg/ml added per well) for 4 h at 37˚C.
Following treatment, the solution was removed and 100 µl DMSO was
added to each well to dissolve the formazan product. Finally, the
OD at a wavelength of 570 nm was assessed using a multifunctional
plate reader (BioTek China) after 15 min of vibration mixing,
following the manufacturer's protocols.
Flow cytometry
Following transfection for 24 h, MLE-12 cell
apoptosis was determined with an Annexin V-FITC/PI apoptosis
detection kit (BD Biosciences), according to the manufacturer's
protocol. A flow cytometer (BD Biosciences) was used to quantify
cell apoptosis and the results were analyzed using CellQuest
software (version 5.1; BD Biosciences).
Detection of caspase-3 activity
The caspase-3 activity in MLE-12 cells was assessed
using a Caspase-3 Assay kit (cat. no. ab39401; Abcam), following
the manufacturer's instructions. In brief, MLE-12 cells were
disintegrated with buffer solution. Lysate was obtained and
centrifuged at 4˚C for 10 min at 1,500 x g. The supernatant was
then treated with caspase-3 reagent, followed by incubation at 37˚C
for 2 h. Following treatment, a microplate reader (BMG Labtech
GmbH) was used to measure the OD indicative of caspase-3 activity
at 405 nm.
Statistical analysis
Statistical analysis was performed using SPSS 20.0
(IBM, Corp.). Values are expressed as the mean ± standard deviation
from three independent experiments. Mean differences between two
groups were estimated with an unpaired Student's t-test and those
among multiple groups by one-way ANOVA followed by Tukey's post-hoc
test. P<0.05 was considered to indicate a statistically
significant difference.
Results
miR-421 directly interacts with
Rian
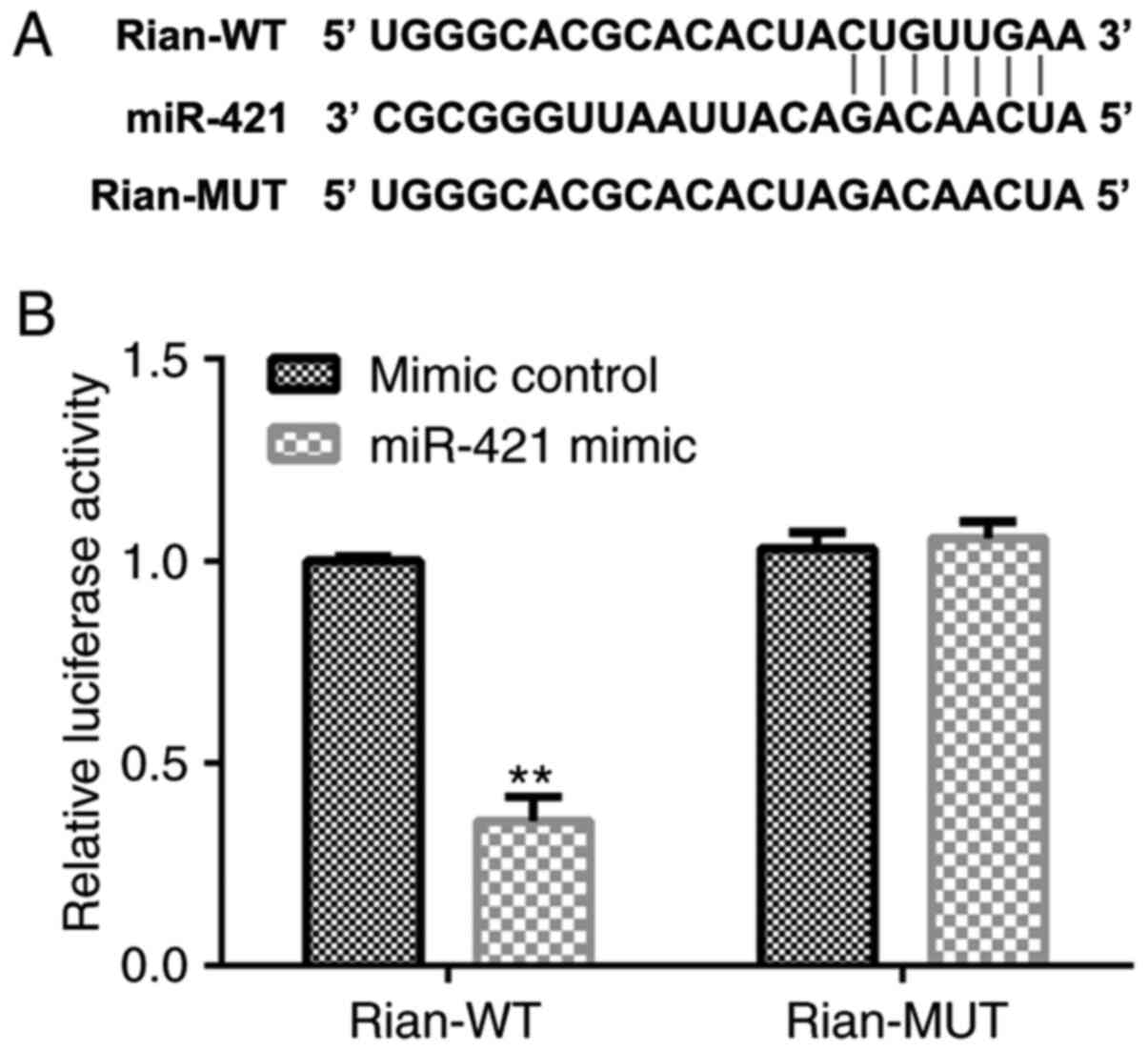
First, the potential association between miR-421 and
Rian was investigated. The prediction with StarBase indicated that
miR-421 was a latent target of Rian (Fig. 1A). Furthermore, the binding site of
miR-421 on Rian was confirmed via a dual-luciferase reporter assay.
In the luciferase reporter assay, miR-421 mimics significantly
decreased the luciferase activity of Rian-WT reporter plasmid,
while no changes were observed in the Rian-MUT luciferase activity
(P<0.01; Fig. 1B). These
observations suggested that miR-421 directly binds to Rian.
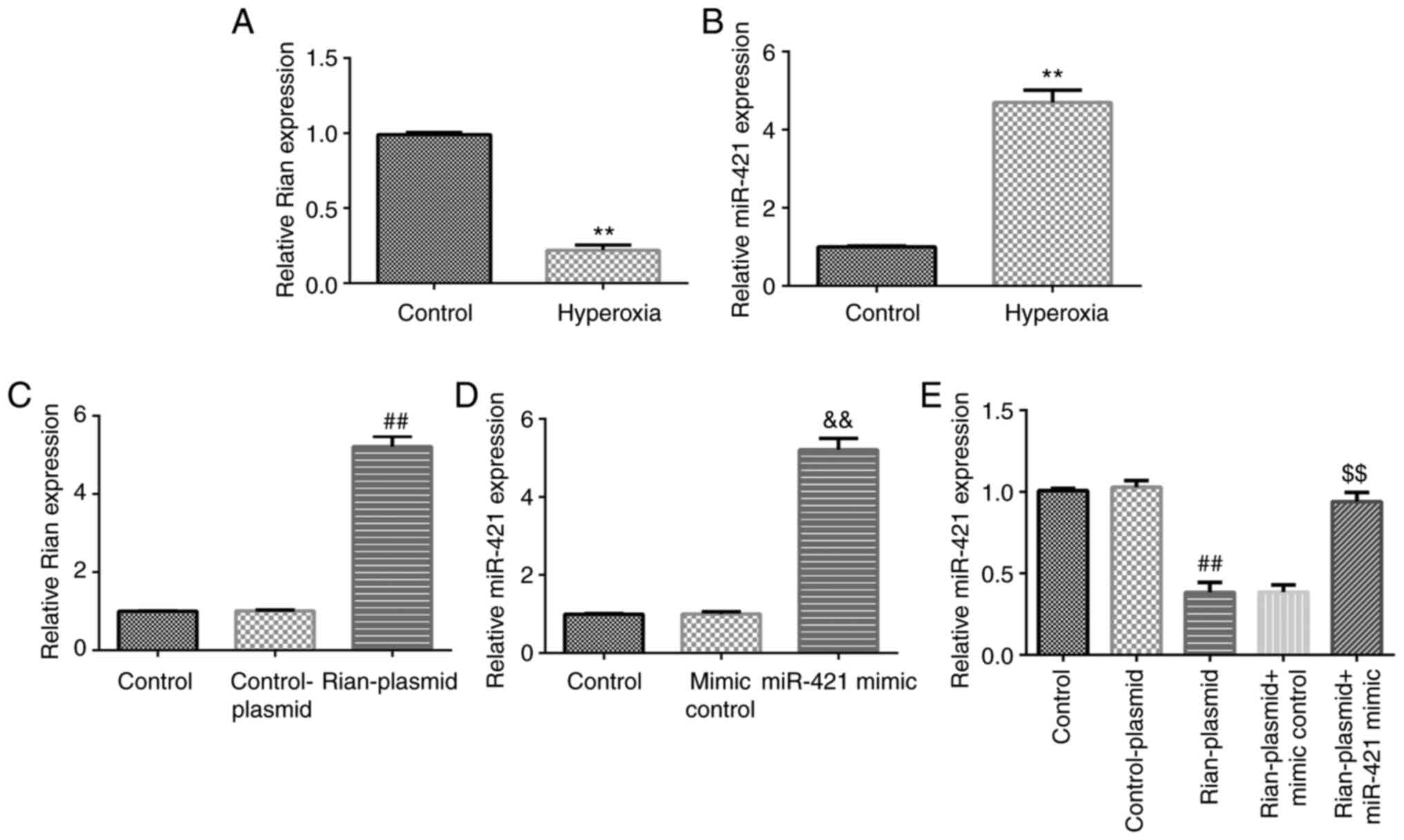
Expression of miR-421 and Rian in the
lung tissue of BPD mice
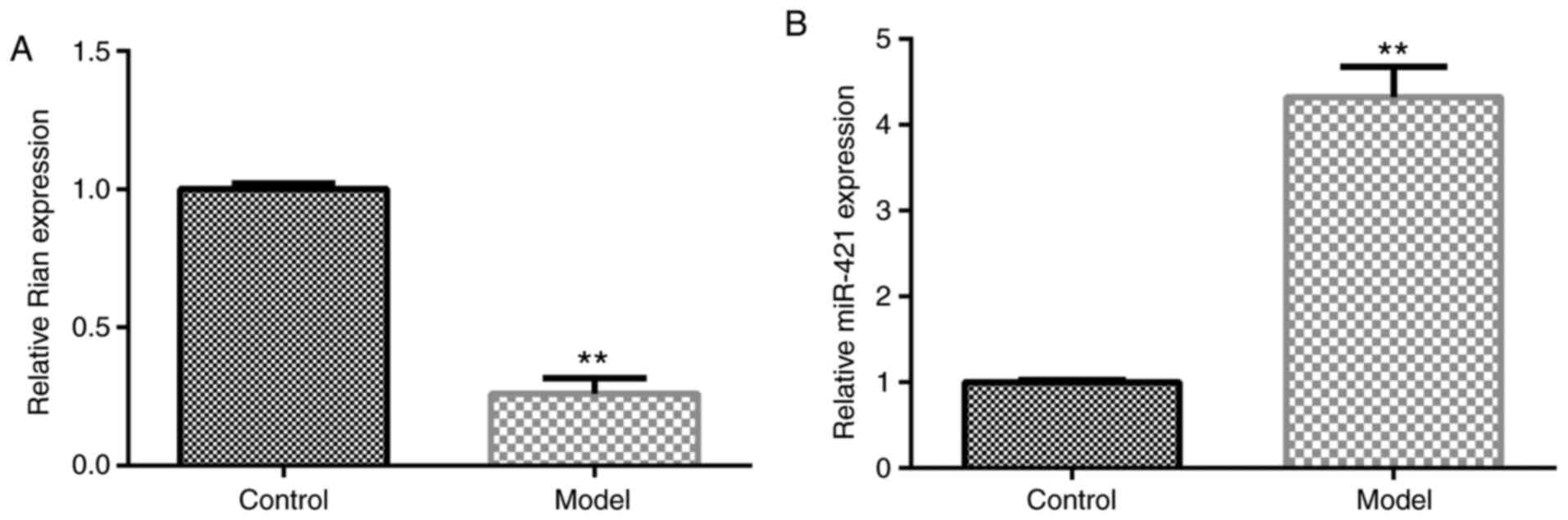
Next, the role of miR-421 or Rian in the progression
of BPD was evaluated using RT-qPCR. The results indicated that Rian
was notably downregulated in BPD mouse lung tissues as compared to
the control (P<0.01; Fig. 2A).
Furthermore, the level of miR-421 was markedly upregulated in the
lung tissue of BPD mice as compared to the control (P<0.01;
Fig. 2B). These results
demonstrated that miR-421/Rian may have a vital role in the
pathogenesis of BPD.
miR-421 mimics abolish the influence
of Rian-plasmid on BPD mice
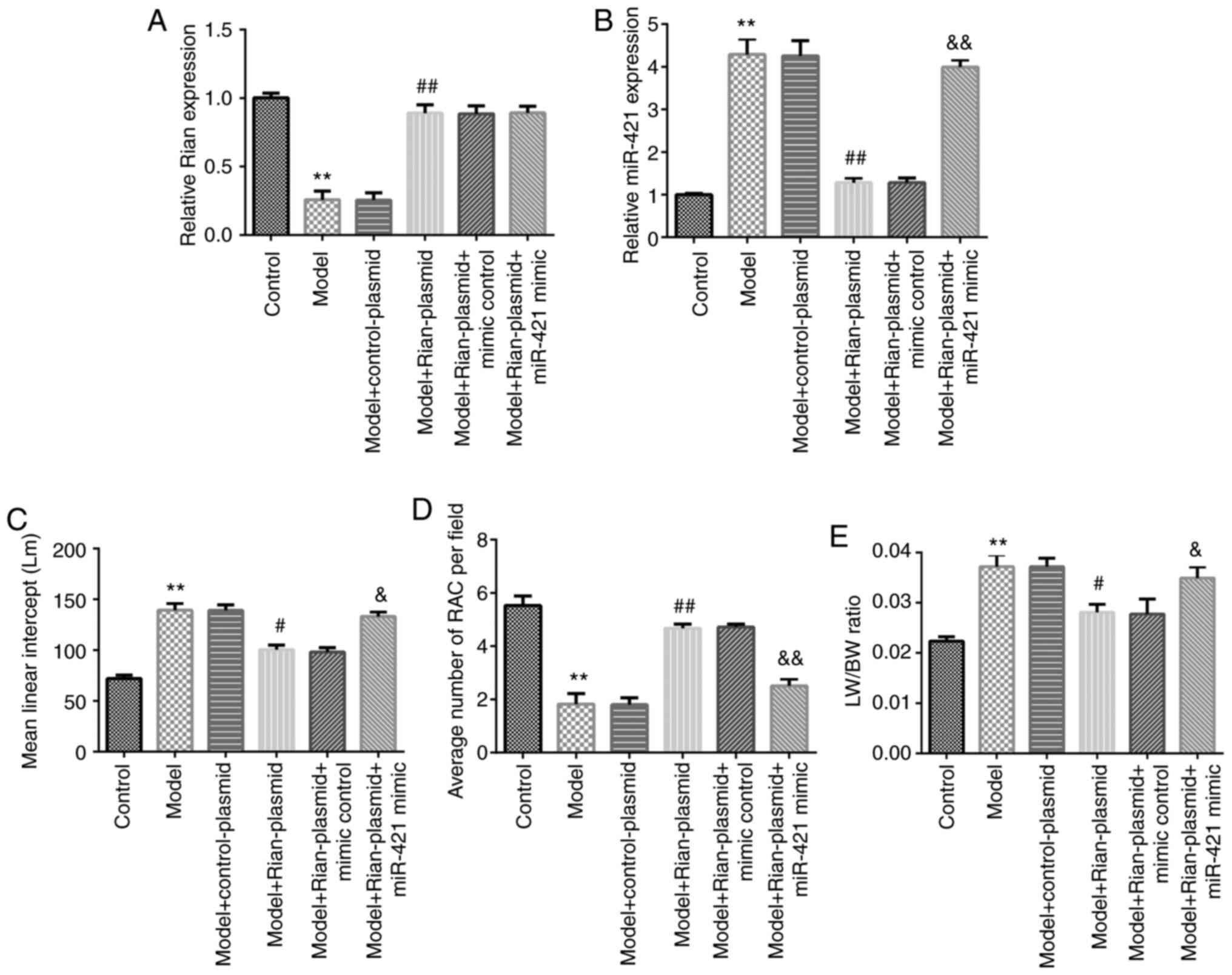
To further reveal the functions of Rian and miR-421
in the development of BPD, control-plasmid, Rian-plasmid, mimics
control or miR-421 mimics were injected into BPD mice and the
levels of Rian or miR-421 were evaluated using RT-qPCR. Rian was
indicated to be downregulated in BPD mouse lung tissues as compared
with those in the control group, while Rian-plasmid significantly
increased Rian expression compared with that in the model + control
plasmid group (P<0.01). No obvious differences in the expression
of Rian were observed among the model + Rian-plasmid, model +
Rian-plasmid + mimics control and model + Rian-plasmid + miR-421
mimics groups (Fig. 3A). In
addition, as presented in Fig. 3B,
Rian-plasmid markedly suppressed the level of miR-421 in BPD mouse
lung tissues, as compared with that in the control-plasmid group;
however, the effect of Rian-plasmid on miR-421 was reversed by
miR-421 mimics (P<0.01; Fig.
3B). Furthermore, quantitative analyses of the RAC and the MLI
of mice in different groups were performed to evaluate the
hyperoxia-induced lung damage and the LW/BW ratio was calculated as
an index of lung injury (4,30,31).
An increased MLI (P<0.01; Fig.
3C), suppressed RAC (P<0.01; Fig. 3D) and enhanced LW/BW ratio
(P<0.01; Fig. 3E) were observed
in model mice. As compared with the Model + control-plasmid group,
Rian-plasmid significantly decreased the MLI level (P<0.05),
enhanced RAC expression (P<0.01) and reduced the LW/BW ratio
(P<0.05), and these effects were reversed by miR-421 mimics. In
conclusion, the present results suggested that Rian exerted its
effect in BPD via miR-421.
miR-421 mimics reverse the effects of
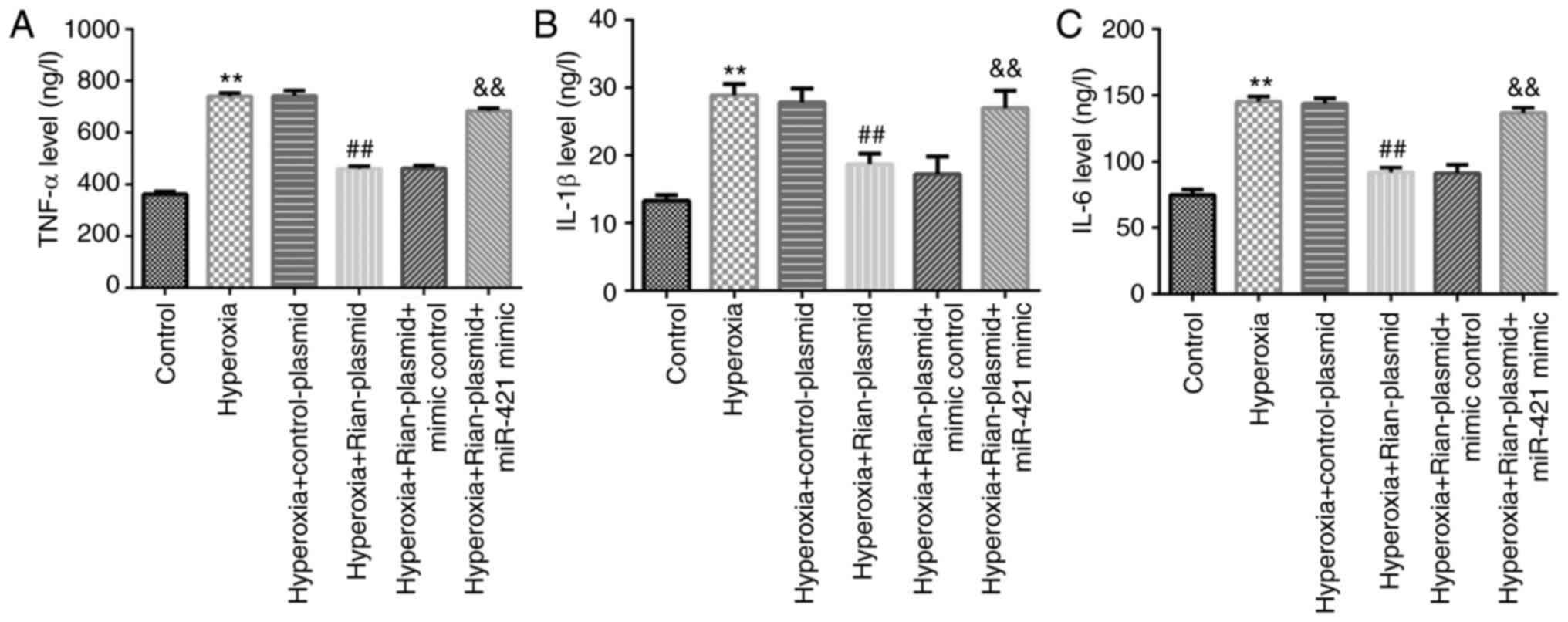
Rian-plasmid on inflammatory response in BPD mice
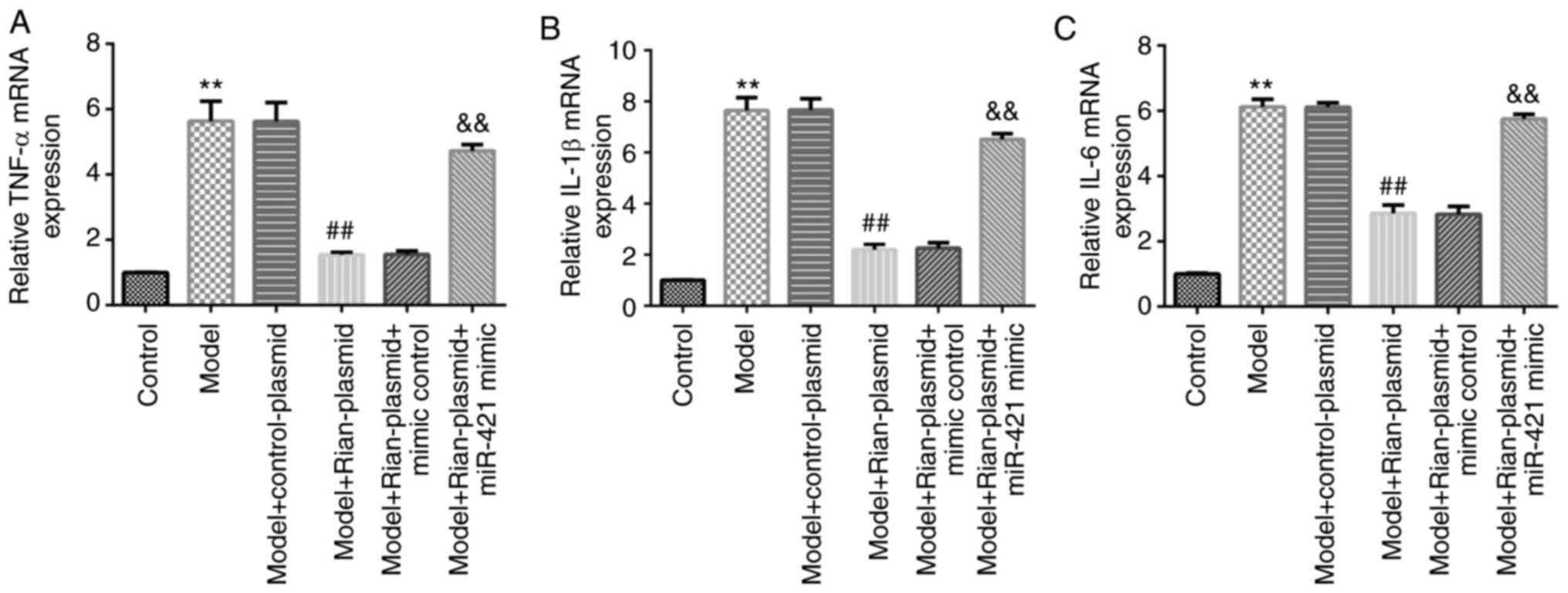
Given that immunoreaction is a vital marker in the
pathophysiological processes of BPD (7), RT-qPCR was performed to examine the
levels of inflammatory factors (TNF-α, IL-6 and IL-1β) in the lung
tissue of mice from different groups. The results revealed that
TNF-α (Fig. 4A), IL-6 (Fig. 4B) and IL-1β (Fig. 4C) levels were notably elevated in
model mouse lung tissues (P<0.01). In addition, in the Model +
Rian-plasmid group, the levels of TNF-α, IL-6 and IL-1β were
markedly inhibited as compared with those in the Model +
control-plasmid group (P<0.01). However, these inhibitions were
reversed following miR-421 mimics treatment.
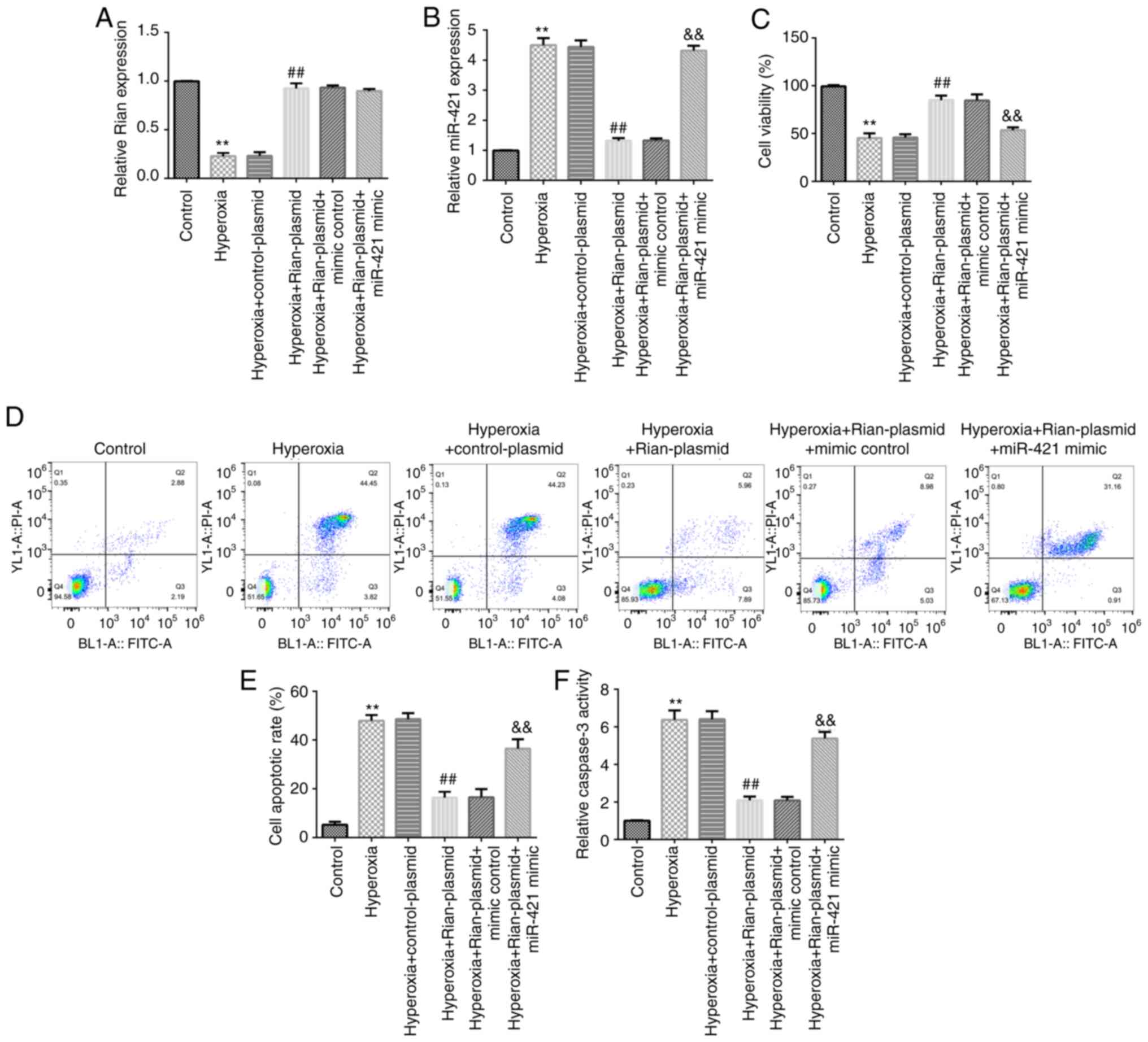
Effects of Rian-plasmid and miR-421
mimics on Rian or miR-421 expression in hyperoxia-induced lung
cells
Furthermore, the expression of Rian and miR-421 in
hyperoxia-induced lung cells were determined. RT-qPCR analysis
indicated that Rian was downregulated and miR-421 was upregulated
in hyperoxia-induced lung cells (Fig.
5A and B). Then, to determine
whether Rian could regulate miR-421 level, MLE-12 cells were
transfected with control-plasmid, Rian-plasmid, mimics control or
miR-421 mimics for 24 h. Results indicated that as compared with
that in the control-plasmid group, the level of Rian was clearly
increased in the Rian-plasmid group (P<0.01; Fig. 5C). miR-421 was determined to be
overexpressed in miR-421 mimics-transfected cells, as compared with
the mimics control group (P<0.01; Fig. 5D). Furthermore, Rian-plasmid
significantly decreased miR-421 levels and this inhibition was
reversed by miR-421 mimics (P<0.01; Fig. 5E). Based on these in vitro
results, it was hypothesized that Rian may be able to protect
against hyperoxia-induced lung damage, which was then further
assessed.
miR-421 mimics reverse the effects of
Rian-plasmid on MLE-12 cell viability and apoptosis
To further analyze the role of Rian in
hyperoxia-induced lung cells, MLE-12 cells were cultured under room
air (21% O2) or hyperoxia (85% O2) for 6 h.
Control-plasmid, Rian-plasmid, mimics control or miR-421 mimics
were then transfected into cells for 24 h. Subsequently, the
expression of Rian or miR-421 in the different groups was analyzed.
The results of the RT-qPCR analysis demonstrated that the level of
Rian was markedly decreased in the hyperoxia group in comparison to
the control group and markedly increased in the hyperoxia +
Rian-plasmid group in comparison to the hyperoxia + control plasmid
group. However, no obvious difference in the levels of Rian was
observed among the hyperoxia + Rian-plasmid, hyperoxia +
Rian-plasmid + mimics control and hyperoxia + Rian-plasmid +
miR-421 mimics groups (Fig. 6A).
Furthermore, miR-421 was upregulated in the hyperoxia-induced lung
cells in comparison with the control group and downregulated in the
hyperoxia + Rian-plasmid group as compared with that in the
hyperoxia + control plasmid group. However, in comparison with the
hyperoxia + Rian-plasmid + mimics control group, miR-421 mimics
increased the level of miR-421 in the hyperoxia-induced MLE-12
cells co-transfected with Rian-plasmid (Fig. 6B). Further analysis using the MTT
assay and flow cytometry revealed that in the hyperoxia group, cell
viability was inhibited (Fig. 6C)
and apoptosis was promoted (Fig. 6D
and E) in comparison to the control
group. Caspase-3 activity was also enhanced in the hyperoxia group
compared to the control group (Fig.
6F). Transfection with Rian-plasmid was able to reverse the
effects of hyperoxia, which was canceled out by miR-421 mimics.
Therefore, the above results indicated that in BPD, lncRNA Rian
protected against hyperoxia-induced lung cell injury through
targeting miR-421.
miR-421 mimics reverse the effects of
Rian-plasmid on inflammatory response in hyperoxia-induced lung
cells
Furthermore, in the present study, the molecular
mechanisms of action of lncRNA Rian in hyperoxia-induced lung cells
were examined. As presented in Fig.
7, the levels of TNF-α, IL-6 and IL-1β were markedly enhanced
in hyperoxia-induced lung cells. In addition, as compared with
those in the hyperoxia + control-plasmid groups, the TNF-α, IL-6
and IL-1β levels were markedly reduced in the hyperoxia +
Rian-plasmid group (P<0.01). However, these effects were
abolished by miR-421 mimics. The above results suggested that
lncRNA Rian is a crucial regulator in BPD development by sponging
miR-421.
Discussion
BPD is a serious lung disease with a high incidence
in newborns. Previous studies have indicated that newborns with BPD
frequently have respiratory and neurological diseases (32,33).
At present, the major treatments for BPD are mechanical
rehabilitation and drug therapy. Despite the fact that various
studies have explored therapeutic strategies of BPD (34,35),
the pathophysiological mechanisms and pathogenesis of BPD have
remained largely elusive. In addition, certain studies have
demonstrated that accurate diagnosis of BPD and interdisciplinary
care for affected pediatric patients were beneficial for the
treatment of BPD (36,37). Therefore, identifying effective
therapeutic strategies for BPD has become a key challenge in recent
years.
LncRNAs have been indicated to have crucial roles in
various diseases. A study by Cai et al (38) demonstrated that lncRNA gallbladder
cancer drug resistance-associated lncRNA1 (GBCDRlnc1) induces
chemoresistance of gallbladder carcinoma cells by activating
autophagy. Several studies have demonstrated that Rian is an
important regulator of biological processes (15,17-19).
However, the role of lncRNA Rian in BPD has remained to be fully
explored. Therefore, the present study was designed to examine the
functions of Rian and related mechanisms in lung tissues in
BPD.
Studies have confirmed that lncRNAs and miRNAs have
vital roles in the pathological mechanisms of BPD. Certain miRNAs
were confirmed to be vital regulators in disease development. They
may serve as oncogenes or suppressors in different malignancies.
For instance, miR-574-3p was reported to regulate adrenomedullin to
protect premature infants with BPD (39). A study by Wang et al
(40) revealed mRNA, lncRNA,
circular RNA and miRNA expression patterns in BPD mouse lung
tissues. Another study confirmed that miR-421 was upregulated in
BPD mice (25). Furthermore,
miR-421 upregulation or fibroblast growth factor 10 (FGF10)
silencing intensified the progression of BPD, while miR-421
inhibition alleviated bronchopulmonary dysplasia in a mouse model
by targeting FGF10(25). First,
based on a bioinformatics prediction with StarBase, a
dual-luciferase reporter assay confirmed that lncRNA Rian directly
regulated miR-421 by sponging its 3'UTR. Next, BPD mouse models
were generated and the miR-421 or Rian levels in the model and
control groups were evaluated by RT-qPCR. miR-421 was determined to
be upregulated, while Rian was downregulated in BPD mice as
compared to the control group, suggesting that miR-421 and Rian
were involved in the regulation of BPD. Recent studies have
confirmed that the altered expression of certain miRNAs is
associated with tumor diagnosis and prognosis (41,42).
It was speculated that an altered expression of Rian or miR-421 may
obstruct the development of BPD. To confirm the present hypothesis,
control-plasmid, Rian-plasmid, mimics control or miR-421 mimics
were injected in the peritoneum of the mice every other day between
days 2 and 14 after birth. The RT-qPCR results demonstrated that
Rian-plasmid prominently increased Rian expression in BPD lung
tissues and suppressed the miR-421 levels, while the effects of
Rian-plasmid were abolished by miR-421 mimics. In conclusion, the
present results suggested that miR-421 mimics interfered with the
functions of Rian in BPD mouse lung tissues.
The effects of Rian-plasmid or miR-421 mimics on BPD
in mice were also evaluated in the present study. To evaluate the
hyperoxia-induced lung histological damage, quantitative analyses
of the RAC and the MLI were performed and the LW/BW ratio was
determined as an index of lung injury (4,30,31).
On day 14 from plasmid/mimics injection, the MLI, RAC and LW/BW
ratios were evaluated in different groups to assess the injury in
BPD mice. The results suggested that Rian-plasmid relieved mouse
BPD, as evidenced by enhanced RAC, reduced MLI and LW/BW ratio.
However, these effects on the MLI, RAC and LW/BW ratio were
reversed by miR-421 mimics. Supplemental oxygen provided to
newborns with BPD may cause oxidative stress and cytokine secretion
(43). It was previously verified
that increased levels of pro-inflammatory factors are associated
with BPD progression (44). To
further explore the functions of Rian in BPD, pro-inflammatory
cytokine secretion in BPD mice was detected after plasmid/mimics
treatment. It was observed that Rian-plasmid reduced the expression
of TNF-α, IL-6 and IL-1β in BPD mouse serum. Further analysis
suggested that these effects were clearly reversed by miR-421
mimics.
Prolonged hyperoxia exposure may result in lung
damage through the production of highly destructive oxygen radicals
(45). Next, the effect of Rian on
hyperoxia-induced lung cell injury in vitro was explored.
The RT-qPCR results indicated that miR-421 was significantly
overexpressed and Rian was clearly downregulated in
hyperoxia-induced MLE-12 cells, as compared to the control. In
addition, Rian-plasmid enhanced Rian levels and suppressed miR-421
expression in MLE-12 cells, while this inhibition was reversed by
miR-421 mimics. In addition, miR-421 was upregulated in miR-421
mimics-transfected MLE-12 cells. To further understand the
functional role of Rian in BPD, its effects on MLE-12-cell
viability, apoptosis and inflammatory factor secretion were
examined.
Following exposure to room air (21% O2)
or hyperoxia (85% O2) for 6 h, MLE-12 cells were
transfected with control-plasmid, Rian-plasmid, mimics control or
miR-421 mimics for 24 h. Previous studies have demonstrated the
vital roles of cell viability and apoptosis in the occurrence of
BPD (46). Consistent with previous
studies, it was indicated that Rian protects against
hyperoxia-induced lung injury, as evidenced by increased cell
viability and reduced apoptosis, and these effects were reversed by
miR-421 mimics. In addition, the activity of caspase-3 was markedly
depressed in Rian-plasmid-induced MLE-12 cells, and this inhibitory
effect was abolished by miR-421 mimics, demonstrating the
protective effects of Rian in BPD mice by targeting miR-421.
Furthermore, the secretion of inflammatory cytokines by MLE-12
cells was detected in different groups. The data revealed that
TNF-α, IL-6 and IL-1β were markedly upregulated in
hyperoxia-induced lung cells. Furthermore, as compared with those
in the hyperoxia + control-plasmid group, the TNF-α, IL-6 and IL-1β
levels were markedly decreased. However, these effects were
abolished by miR-421 mimics. The above findings suggested that Rian
has an important role in the development of BPD. However, there
were still certain limitations to the present study. For example,
in the animal experiment, the rodents were injected continuously
whilst being kept under continuous hyperoxia. However, in in
vitro experiments, the cells were first subjected to hyperoxia
and then transfected with Rian-overexpression vector/miR-421 mimic.
The effect of transfecting cells with Rian-overexpression
vector/miR-421 mimic under hyperoxia should be further
investigated. The mechanisms of BPD include additional pathways
apart from the miR-421-Rian axis, which remain to be investigated
in the future.
In conclusion, the results of the present study
demonstrated that Rian extenuated hyperoxia-induced lung injury in
BPD by inhibiting inflammatory response and preventing lung cell
apoptosis by targeting miR-421, suggesting that Rian may serve as a
potential diagnostic biomarker and therapeutic agent for patients
with BPD.
Acknowledgements
Not applicable.
Funding
Funding: No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
XT designed the study, in addition to performing all
experiments, analyzing the data and preparing the manuscript. YF
and CH contributed to performing the experiments and data
collection. All authors read and approved the final manuscript. XT
and YF confirm the authenticity of all the raw data.
Ethics approval and consent to
participate
The present study was approved by the Animal Ethics
Committee of the Experimental Animal Center of The First People's
Hospital of Lianyungang (Lianyungang, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Winsper C: The aetiology of borderline
personality disorder (BPD): Contemporary theories and putative
mechanisms. Curr Opin Psychol. 21:105–110. 2018.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Dumpa V and Bhandari V: Surfactant,
steroids and non-invasive ventilation in the prevention of BPD.
Semin Perinatol. 42:444–452. 2018.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Mir IN, Chalak LF, Brown LS, Johnson-Welch
S, Heyne R, Rosenfeld CR and Kapadia VS: Impact of multiple
placental pathologies on neonatal death, bronchopulmonary
dysplasia, and neurodevelopmental impairment in preterm infants.
Pediatr Res. 87:885–891. 2020.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Maturu P, Wei-Liang Y, Androutsopoulos VP,
Jiang W, Wang L, Tsatsakis AM and Couroucli XI: Quercetin
attenuates the hyperoxic lung injury in neonatal mice: Implications
for Bronchopulmonary dysplasia (BPD). Food Chem Toxicol. 114:23–33.
2018.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Bashir RA, Bhandari V, Vayalthrikkovil S,
Rabi Y, Soraisham A, Tang S, Al Awad E and Lodha A:
Chorioamnionitis at birth does not increase the risk of
neurodevelopmental disability in premature infants with
bronchopulmonary dysplasia. Acta Paediatr. 105:e506–e512.
2016.PubMed/NCBI View Article : Google Scholar
|
|
6
|
De Paepe ME, Mao Q, Chao Y, Powell JL,
Rubin LP and Sharma S: Hyperoxia-induced apoptosis and Fas/FasL
expression in lung epithelial cells. Am J Physiol Lung Cell Mol
Physiol. 289:L647–L659. 2005.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Kallapur GS and Jobe AH: Contribution of
inflammation to lung injury and development. Arch Dis Child Fetal
Neonatal Ed. 91:F132–F135. 2006.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Fiaturi N, Russo JW, Nielsen HC and
Castellot JJ Jr: CCN5 in alveolar epithelial proliferation and
differentiation during neonatal lung oxygen injury. J Cell Commun
Signal. 12:217–229. 2018.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Ratner V, Slinko S, Utkina-Sosunova I,
Starkov A, Polin RA and Ten VS: Hypoxic stress exacerbates
hyperoxia-induced lung injury in a neonatal mouse model of
bronchopulmonary dysplasia. Neonatology. 95:299–305.
2009.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Zhu Y, Fu J, Yang H, Pan Y, Yao L and Xue
X: Hyperoxia-induced methylation decreases RUNX3 in a newborn rat
model of bronchopulmonary dysplasia. Respir Res.
16(75)2015.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Mai LJ, Fu XX, He G, Zhao EN and Xue M:
Effect of asiaticoside on hyperoxia-induced bronchopulmonary
dysplasia in neonatal rats and related mechanism. Zhongguo Dang Dai
Er Ke Za Zhi. 22:71–76. 2020.PubMed/NCBI View Article : Google Scholar : (In Chinese).
|
|
12
|
Hu Y, Xie L, Yu J, Fu H, Zhou D and Liu H:
Inhibition of microRNA-29a alleviates hyperoxia-induced
bronchopulmonary dysplasia in neonatal mice via upregulation of
GAB1. Mol Med. 26(3)2019.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Zhang T, Chen J, Wu H, Pan W, Yang X, Li
Y, Liu M and Huang Y: Improved survival and survival without
bronchopulmonary dysplasia in very low birth weight infants after
active perinatal care. Niger J Clin Pract. 23:980–987.
2020.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Salviano-Silva A, Lobo-Alves SC, Almeida
RC, Malheiros D and Petzl-Erler ML: Besides pathology: Long
non-coding RNA in cell and tissue homeostasis. Noncoding RNA.
4(3)2018.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Bijkerk R, Au YW, Stam W, Duijs JMGJ,
Koudijs A, Lievers E, Rabelink TJ and van Zonneveld AJ: Long
non-coding RNAs Rian and Miat mediate myofibroblast formation in
kidney fibrosis. Front Pharmacol. 10(215)2019.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Gu T, He H, Xing Y, Liu Q, Gu N, Kenkichi
S, Jiang H and Wu Q: Expression of non-coding RNA AB063319 derived
from Rian gene during mouse development. J Mol Histol. 42:105–112.
2011.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Yao P, Li YL, Chen Y, Shen W, Wu KY and Xu
WH: Overexpression of long non-coding RNA Rian attenuates cell
apoptosis from cerebral ischemia-reperfusion injury via
Rian/miR-144-3p/GATA3 signaling. Gene. 737(144411)2020.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Zhong L, Jia J and Ye G:
Rian/miR-210-3p/Nfkb1 feedback loop promotes hypoxia-induced cell
apoptosis in myocardial infarction through deactivating the
PI3K/Akt signaling pathway. J Cardiovasc Pharmacol. 76:207–215.
2020.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Liu B, Li J and Cairns MJ: Identifying
miRNAs, targets and functions. Brief Bioinform. 15:1–19.
2014.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Dupont C, Kappeler L, Saget S, Grandjean V
and Levy R: Role of miRNA in the transmission of metabolic diseases
associated with paternal diet-induced obesity. Front Genet.
10(337)2019.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Chen J, Wu L, Sun Y, Yin Q, Chen X, Liang
S, Meng Q, Long H, Li F, Luo C and Xiao X: Mir-421 in plasma as a
potential diagnostic biomarker for precancerous gastric lesions and
early gastric cancer. PeerJ. 7(e7002)2019.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Ren Z, He M, Shen T, Wang K, Meng Q, Chen
X, Zhou L, Han Y, Ji C, Liu S and Fu Q: MiR-421 promotes the
development of osteosarcoma by regulating MCPIP1 expression. Cancer
Biol Ther. 21:231–240. 2020.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Yin Y, Xu L, Chang Y, Zeng T, Chen X, Wang
A, Groth J, Foo WC, Liang C, Hu H and Huang J: N-Myc promotes
therapeutic resistance development of neuroendocrine prostate
cancer by differentially regulating miR-421/ATM pathway. Mol
Cancer. 18(11)2019.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Chen L, Tang Y, Wang J, Yan Z and Xu R:
miR-421 induces cell proliferation and apoptosis resistance in
human nasopharyngeal carcinoma via downregulation of FOXO4. Biochem
Biophys Res Commun. 435:745–750. 2013.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Yuan HS, Xiong DQ, Huang F, Cui J and Luo
H: MicroRNA-421 inhibition alleviates bronchopulmonary dysplasia in
a mouse model via targeting Fgf10. J Cell Biochem. 120:16876–16887.
2019.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Park MS, Rieger-Fackeldey E, Schanbacher
BL, Cook AC, Bauer JA, Rogers LK, Hansen TN, Welty SE and Smith CV:
Altered expressions of fibroblast growth factor receptors and
alveolarization in neonatal mice exposed to 85% oxygen. Pediatr
Res. 62:652–657. 2007.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Li JH, Liu S, Zhou H, Qu LH and Yang JH:
starBase v2.0: Decoding miRNA-ceRNA, miRNA-ncRNA and protein-RNA
interaction networks from large-scale CLIP-Seq data. Nucleic Acids
Res. 42 (Database Issue):D92–D97. 2014.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Clément T, Salone V and Rederstorff M:
Dual luciferase gene reporter assays to study miRNA function.
Methods Mol Biol. 1296:187–198. 2015.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Chen X, Zhang X and Pan J: Effect of
montelukast on bronchopulmonary dysplasia (BPD) and related
mechanisms. Med Sci Monit. 25:1886–1893. 2019.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Chen X, Peng W, Zhou R, Zhang Z and Xu J:
Montelukast improves bronchopulmonary dysplasia by inhibiting
epithelial-mesenchymal transition via inactivating the TGF-β1/Smads
signaling pathway. Mol Med Rep. 22:2564–2572. 2020.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Stark A, Dammann C, Nielsen HC and Volpe
MV: A pathogenic relationship of bronchopulmonary dysplasia and
retinopathy of prematurity? A review of angiogenic mediators in
both diseases. Front Pediatr. 6(125)2018.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Kalikkot TR, Guaman MC and Shivanna B:
Bronchopulmonary dysplasia: A review of pathogenesis and
pathophysiology. Respir Med. 132:170–177. 2017.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Principi N, Di Pietro GM and Esposito S:
Bronchopulmonary dysplasia: Clinical aspects and preventive and
therapeutic strategies. J Transl Med. 16(36)2018.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Hwang JS and Rehan VK: Recent advances in
bronchopulmonary dysplasia: Pathophysiology, prevention, and
treatment. Lung. 196:129–138. 2018.PubMed/NCBI View Article : Google Scholar
|
|
36
|
van Rossem MC, van de Loo M, Laan BJ, de
Sonnaville ES, Tamminga P, van Kaam AH and Onland W: Accuracy of
the diagnosis of bronchopulmonary dysplasia in a referral-based
health care system. J Pediatr. 167:540–544.e1. 2015.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Abman SH, Collaco JM, Shepherd EG, Keszler
M, Cuevas-Guaman M, Welty SE, Truog WE, McGrath-Morrow SA, Moore
PE, Rhein LM, et al: Interdisciplinary care of children with severe
bronchopulmonary dysplasia. J Pediatr. 181:12–28. 2017.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Cai Q, Wang S, Jin L, Weng M, Zhou D, Wang
J, Tang Z and Quan Z: Long non-coding RNA GBCDRlnc1 induces
chemoresistance of gallbladder cancer cells by activating
autophagy. Mol Cancer. 18(82)2019.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Gong X, Qiu J, Qiu G and Cai C:
Adrenomedullin regulated by miRNA-574-3p protects premature infants
with bronchopulmonary dysplasia. Biosci Rep.
40(BSR20191879)2020.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Wang J, Yin J, Wang X, Liu H, Hu Y, Yan X,
Zhuang B, Yu Z and Han S: Changing expression profiles of mRNA,
lncRNA, circRNA and miRNA in lung tissue reveal the
pathophysiological of bronchopulmonary dysplasia (BPD) in mouse
model. J Cell Biochem. 120:9369–9380. 2019.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Sun Z, Shi K, Yang S, Liu J, Zhou Q, Wang
G, Song J, Li Z, Zhang Z and Yuan W: Effect of exosomal miRNA on
cancer biology and clinical applications. Mol Cancer.
17(147)2018.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Xu B, Liu J, Xiang X, Liu S, Zhong P, Xie
F, Mou T and Lai L: Expression of miRNA-143 in pancreatic cancer
and its clinical significance. Cancer Biother Radiopharm.
33:373–379. 2018.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Hanna Y, Laliberté C, Ben Fadel N, Lemyre
B, Thébaud B, Barrowman N, Bijelic V, Hoey L and Katz SL: Effect of
oxygen saturation targets on the incidence of bronchopulmonary
dysplasia and duration of respiratory supports in extremely preterm
infants. Paediatr Child Health. 25:173–9. 2020.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Kaneko M, Sato M, Ogasawara K, Imamura T,
Hashimoto K, Momoi N and Hosoya M: Serum cytokine concentrations,
chorioamnionitis and the onset of bronchopulmonary dysplasia in
premature infants. J Neonatal Perinatal Med. 10:147–155.
2017.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Dias-Freitas F, Metelo-Coimbra C and
Roncon-Albuquerque RJ: Molecular mechanisms underlying hyperoxia
acute lung injury. Respir Med. 119:23–28. 2016.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Yin R, Yuan L, Ping L and Hu L: Neonatal
bronchopulmonary dysplasia increases neuronal apoptosis in the
hippocampus through the HIF-1alpha and p53 pathways. Respir Physiol
Neurobiol. 220:81–87. 2016.PubMed/NCBI View Article : Google Scholar
|