Introduction
Lower extremity arteriosclerosis obliterans (ASO)
refers to the deposition of lipids on the blood vessel walls of
lower limbs, which is rapidly relieved by interventional therapy
(1). However, the 1-year recurrence
rate of stenosis is 30-50% (2).
Therefore, it is crucial to identify the molecular mechanisms
underlying the ASO process to provide novel strategies for clinical
treatments. Excessive proliferation and migration of vascular
smooth muscle cells (VSMCs), mainly caused by the
monocyte/macrophage-induced inflammatory response, serve pivotal
roles in ASO progression and restenosis following treatment
(3). Therefore, the prevention and
relief of dysfunction of VSMCs are important for ASO treatment.
MicroRNAs (miRNAs/miRs) are involved in the
occurrence and progression of ASO. For example, miR-30b-5p is
involved in the differentiation of VSMCs through interaction with
muscleblind like splicing regulator 1(4). In addition, miR-143/145 inhibits the
proliferation of VSMCs by regulating the expression of mammalian
transcription factors and myocardin (5,6). These
findings confirm the importance of miRNAs in regulating the
dysfunction of VSMCs. The expression of miR-125b is notably altered
in ASO (7). For example, a previous
study demonstrated that miR-125b expression decreases in VSMCs and
miR-125b inhibits the proliferation and migration of VSMCs
(7). However, the underlying
molecular mechanism of miR-125b in ASO remains unclear. Therefore,
it is critical to investigate the potential promoting effects of
miR-125b on ASO progression to identify novel treatment strategies
for ASO.
The present study aimed to investigate the role of
miR-125b in ASO by simulating ASO with platelet-derived growth
factor-BB (PDGF-BB). The effect of PDGF-BB on miR-125b expression
was investigated. Subsequently, A10 cells were treated with PDGF-BB
to compare the effects of upregulated miR-125b expression on the
proliferative and migratory abilities of VSMCs. The potential
targets of miR-125b in VSMCs were also investigated.
Materials and methods
Cell culture
A10 cells from rat VSMCs were used in the present
study. The A10 cell line was purchased from The Cell Bank of Type
Culture Collection of The Chinese Academy of Sciences. Cells were
maintained in Dulbecco's modified Eagle's medium (DMEM; Gibco;
Thermo Fisher Scientific, Inc.) supplemented with 10% fetal calf
serum, 100 U/ml penicillin and 100 U/ml streptomycin (Thermo Fisher
Scientific, Inc.) at 37˚C with 5% CO2. PDGF-BB (20
ng/ml; PeproTech, Inc.), was used to investigate the effect of
PDGF-BB on miR-125b expression.
miR-125b mimic and inhibitor
transfection
A10 cells were seeded into 6-well plates
(3x105 cells per well) for 24 h and subsequently
transfected with miR-125b mimic or miR-125b inhibitor in parallel
with their respective control oligos (60 nmol/l; Shanghai
GenePharma Co., Ltd.) for 24 h at 37˚C using
Lipofectamine® 3000 reagent (Invitrogen; Thermo Fisher
Scientific, Inc.) and Opti-MEM (Thermo Fisher Scientific, Inc.),
according to the manufacturer's protocols. The sequences of
miR-125b mimic and inhibitor were as follows: miR-125b mimic
forward, 5'-UCCCUGAGACCCUAACUUGUGA-3' and reverse,
5'-UCACAAGUUAGGGUCUCAGGGA-3'; NC mimic forward,
5'-UUUGUACUACACAAAAGUACUG-3' and reverse,
5'-CAGUACUUUUGUGUAGUACAAA-3'; miR-125b inhibitor,
5'-UCACAAGUUAGGGUCUCAGGGA-3' and NC inhibitor,
5'-CAGUACUUUUGUGUAGUACAAA-3'. After RNA interference was performed,
the cells were cultured for an additional 48 h. Reverse
transcription-quantitative polymerase chain reaction (RT-qPCR) was
performed to assess the efficiency of miR-125b transfection.
RT-qPCR
Cells with different treatments were collected and
total RNA was extracted using TRIzol® reagent
(Invitrogen; Thermo Fisher Scientific, Inc.). Total RNA was reverse
transcribed into cDNA according to the manufacturer's instructions
of the PrimeScript RT Reagent kit (cat. no. RR047A; Takara
Biotechnology Co., Ltd.). mRNA expression was analyzed using a 7500
Fast™ system (Applied Biosystems; Thermo Fisher Scientific, Inc.)
and a Sensi Mix SYBR kit (QP10005; OriGene Technologies, Inc.). The
PCR conditions were as follows: 95˚C for 10 sec, 95˚C for 5 sec and
60˚C for 20 sec (35 cycles), followed by 95˚C for 15 sec, and 65˚C
for 10 sec. The following primer sequences were used for the qPCR:
miR-125b forward, 5'-GCGCTCCCTGAGACCCTAAC-3' and reverse,
5'-TGCAGGGTCCGAGGTAT-3'; and U6 forward, 5'-CTCGCTTCGGCAGCACA-3'
and reverse, 5'-AACGCTTCACGAATTTGCGT-3'. Relative expression levels
were calculated using the 2-∆∆Ct method (8) and normalized to the internal reference
gene U6.
Cell proliferation assay
Cell proliferation was assessed via Cell Counting
Kit-8 (CCK-8; Beyotime Institute of Biotechnology) and BrdU assays.
For the CCK-8 assay, In brief, 2x103 cells were seeded
into 96-well plates with 200 µl culture medium. Subsequently, 20 µl
CCK-8 regent was added into each well after 24 h incubation at
37˚C, and then were incubated at 37˚C in dark for another 4 h. The
optical density (OD) values were detected at 450 nm using a
microplate reader (Tecan Group, Ltd.). Cell proliferation was
quantified by standard curves, and a linear standard curve was
fitted for log [cell quantity] and OD. For the BrdU
assay, cells were seeded into 24-well plates for 24 h at 37˚C,
followed by different treatments. Cell proliferation was assessed
using the BeyoClick™ EDU-647 system (Beyotime Institute of
Biotechnology) at a wavelength of 450 nm.
Transwell assay
For the Transwell assay, 5x104 cells with
different treatments were plated into the upper chambers (24-well,
8.0 µm, Corning, Inc.) with serum free medium, and medium
supplemented with 10% FBS was plated into the lower chambers. After
48 h incubation at 37˚C with 5% CO2, the migratory cells
were stained with 0.5% crystal violet for 15 min at room
temperature and observed under a light microscope (magnification,
x100; Olympus Corporation).
Wound healing assay
For the wound healing assay, cells were seeded into
6-well plates until they reached 80% confluence. The cell
monolayers were scratched using 200 µl pipette tips, washed with
PBS to remove the detached cells and cultured in DMEM with 0.5% FBS
for 48 h. The wound distance was observed under a light microscope
(magnification, x200; Olympus Corporation).
Western blotting
Cells with different treatments were harvested with
a cytology brush, washed with ice-cold PBS and lysed using RIPA
lysis buffer (Sigma-Aldrich; Merck KGaA), containing a
phosphorylase and protease inhibitor mixture (Thermo Fisher
Scientific, Inc.). Total protein was quantified via the BCA assay.
Proteins (40 µg per lane) were separated by 10% SDS-PAGE, separated
proteins were transferred to PVDF membranes, which were
subsequently blocked with Tris-buffered saline with Tween solution
(0.1% Tween-20) and 5% skimmed milk at 4˚C overnight. The membranes
were incubated with primary antibodies against SRF (dilution,
1:1,000; cat. no. 16821-1-AP; ProteinTech Group, Inc.), AAMP
(dilution, 1:1,000; cat. no. 21220-1-AP; ProteinTech Group, Inc.)
and GAPDH (dilution, 1:10,000; cat. no. ab9385; Abcam) at 4˚C
overnight. Following incubating with the primary antibody,
membranes were incubated with secondary antibodies [horseradish
peroxidase-conjugated goat anti rabbit IgG (1:5,000; cat. no.
ab6721) or goat anti mouse IgG (1:2,000; cat. no. ab6789; Abcam)]
for 1.5 h at room temperature, which was performed in TBS-Tween
solution containing 5% skimmed milk. ECL chemiluminescence (Thermo
Fisher Scientific, Inc.) was followed by exposure and development.
ImageJ software (version 1.46; National Institutes of Health) was
used for analysis.
Immunofluorescence
In brief, A10 cells were seeded into 24-well plates
for 24 h and fixed with 4% paraformaldehyde at 4˚C for 30 min,
followed by permeabilization with 0.5% Triton X-100 and blocking
with 5% bovine serum albumin (Sigma-Aldrich; Merck KGaA) at room
temperature for 30 min. Samples were incubated with primary
antibodies against SRF (dilution, 1:200; cat. no. 16821-1-AP;
ProteinTech Group, Inc.) and AAMP (dilution, 1:200; cat. no.
21220-1-AP; ProteinTech Group, Inc.) overnight at 4˚C. Samples were
subsequently washed with PBS and incubated with secondary
antibodies for 1 h at room temperature, prior to re-washing. Nuclei
were stained with 15 µl DAPI (Sigma-Aldrich; Merck KGaA) for 2 min
at room temperature, prior to being observed under a fluorescence
microscope (magnification, x200; Carl Zeiss AG).
Statistical analysis
Statistical analysis was performed using SPSS 20.0
software (IBM Corp.). All experiments were performed for three
repeats excluding for Western blotting, which was only conducted
once. Data are presented as the mean ± standard deviation. Paired
Student's t-test was used to compare differences between two
groups, while one-way analysis of variance and the Tukey's post hoc
test were used to compare differences among multiple groups.
P<0.05 was considered to indicate a statistically significant
difference.
Results
PDGF-BB negatively regulates miR-125b
expression in VSMCs
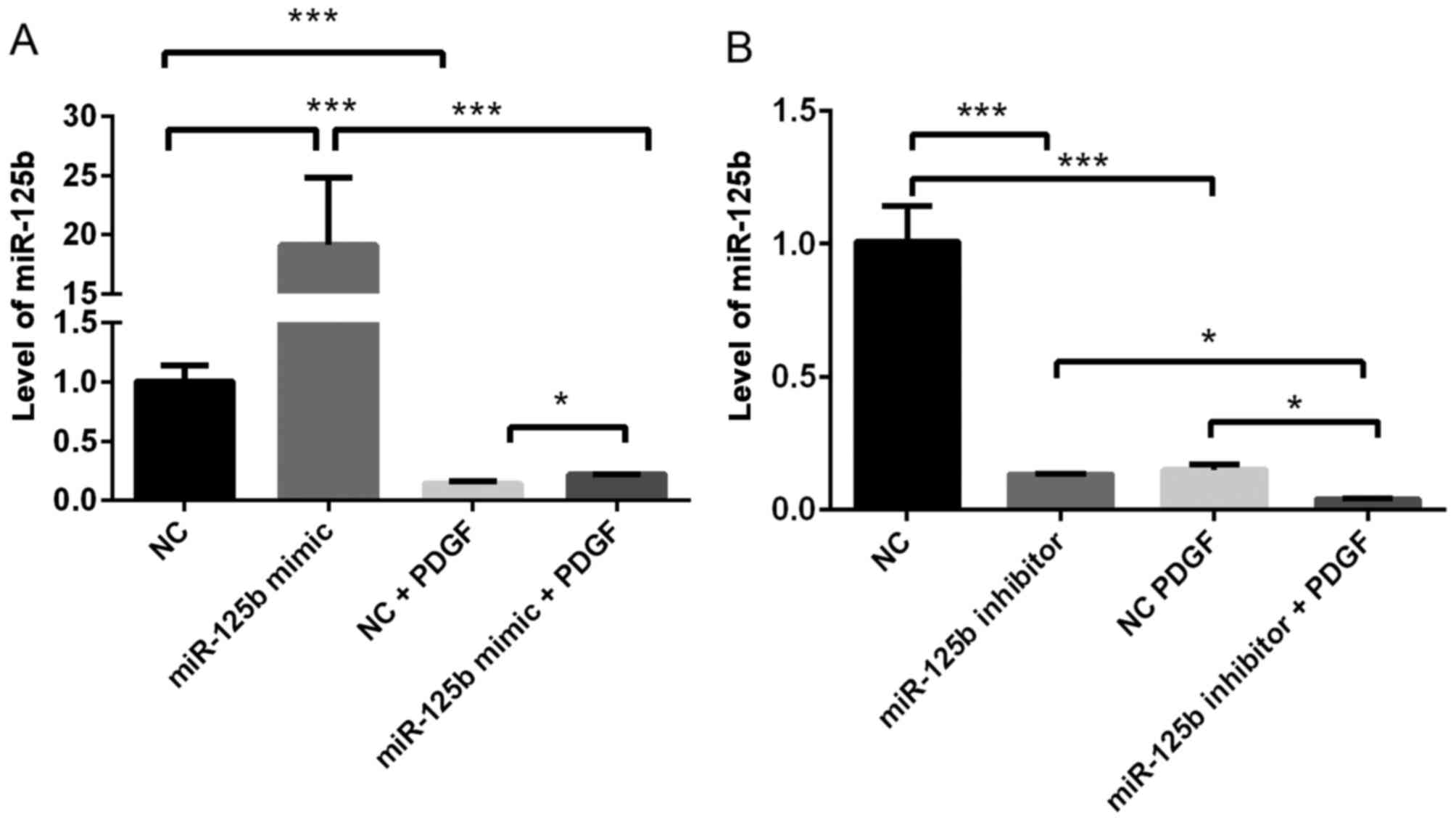
To determine the effect of PDGF-BB on miR-125b
expression, miR-125b expression levels in VSMCs with or without
PDGF-BB treatment were compared. As presented in Fig. 1, treatment with PDGF-BB
significantly decreased miR-125b expression in VSMCs (~80%;
P=0.0004). Notably, overexpression of miR-125b significantly
rescued miR-125b expression following treatment with PDGF-BB (~40%
upregulation; P=0.03).
miR-125b inhibits the proliferation of
VSMCs
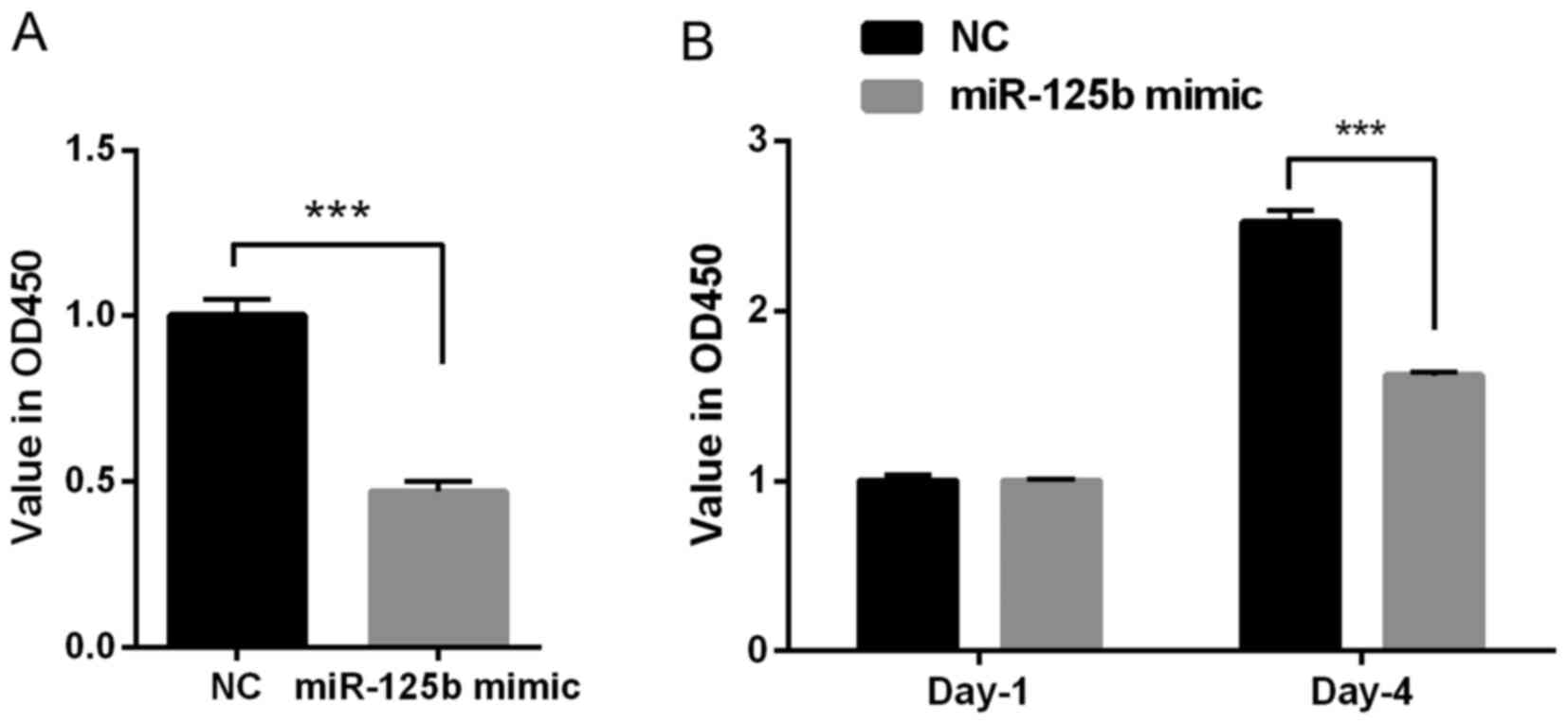
To investigate the role of miR-125b on the
proliferation of VSMCs, the CCK-8 and BrdU assays were performed.
The results of the CCK-8 assay indicated that the OD value was
decreased in the miR-125b mimic group compared with the NC group
(P<0.001; Fig. 2A); therefore,
the overexpression of miR-125b inhibited the proliferation of
VSMCs. Furthermore, the results of the BrdU assay demonstrated that
transfection with miR-125b mimic significantly decreased the OD
value at day 4 (~40%), compared with the NC group (P<0.001;
Fig. 2B). Taken together, these
results suggested that miR-125b inhibits cell proliferation.
miR-125b inhibits the migration of
VSMCs
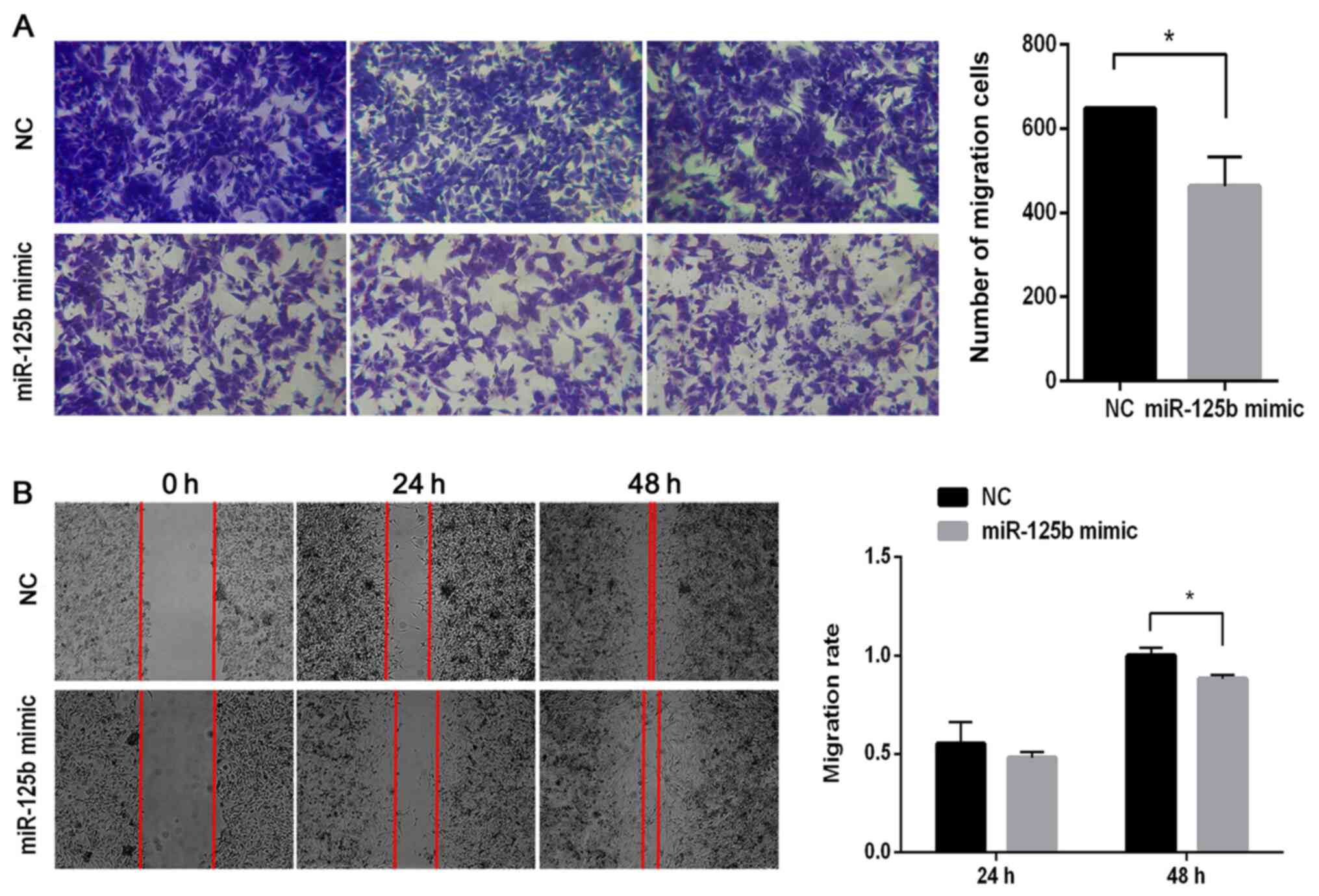
To investigate the role of miR-125b on the migration
of VSMCs, Transwell migration and wound healing assays were
performed. The results of the Transwell assay demonstrated that the
number of migratory cells significantly decreased in the miR-125b
mimic group compared with the NC group (P<0.05; Fig. 3A). As presented in Fig. 3B, there were few differences between
the NC and miR-125b mimic groups after 24 h. However, following 48
h of incubation, the cells had almost completely migrated into the
wound in the NC group, while the gaps remained visible in the
miR-125b mimic group. Following normalization, the migratory
ability in the miR-125b mimic group was ~88% of that in the NC
group (P<0.05).
AAMP and SRF are targets of miR-125b
in VSMCs
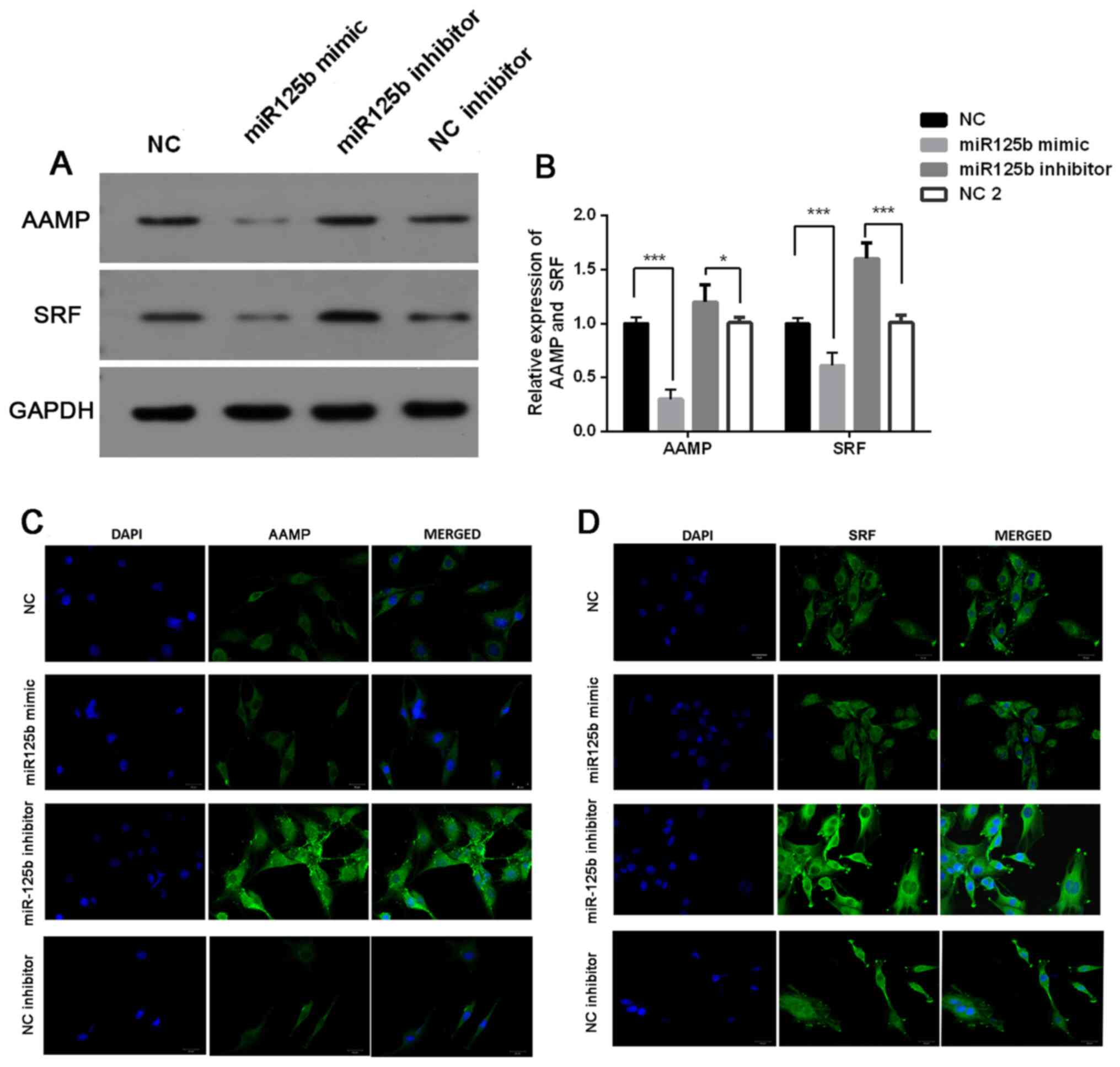
The present study investigated the association
between miR-125b expression and AAMP and SRF expression. The
results demonstrated that the protein expression levels of AAMP and
SRF were significantly downregulated following transfection with
miR-125b mimic, the effects of which were reversed following
transfection with miR-125b inhibitor (P<0.05; Fig. 4A and B). Immunofluorescence analysis
demonstrated that the miR-125b mimic-induced downregulation of AAMP
and SRF was very slight, while miR-125b inhibitor-induced
upregulation of AAMP and SRF expression levels was significant
(Fig. 4C and D).
Discussion
Lower extremity ASO is induced by atherosclerotic
thrombosis, which causes narrowing or occlusion of the lower
extremity arteries, and the persistent ischemia eventually leads to
tissue necrosis (9). The prevalence
of ASO increases with age, which affects ~8-10% of people >65
years and ~20% of people >80 years (9). ASO is initiated following the
accumulation of lipoproteins in the endothelium due to the abnormal
metabolism of glucose and lipid (1,10).
This in turn results in a series of intracellular reactions, which
eventually induces vascular endothelial damage and the
monocyte/macrophage-induced inflammatory response (11). Inflammatory cells and factors
regulate the function of VSMCs, including cell proliferation,
migration, differentiation and matrix secretion (11). VSMCs are the major cellular
components that make up the middle layer of the arteries and are
involved in the process of vascular diseases. At present, it is
believed that the excessive proliferation and migration of VSMCs is
one of the main causes of ASO, and it is also the main cause of
restenosis following interventional treatment (10). Therefore, the prevention and relief
of dysfunction of VSMCs are important for ASO treatment.
PDGF-BB derived from platelets is considered as a
mediator of VSMC dysfunction and restenosis (12). PDGF-BB is a well-known
atherosclerosis promotional factor (13). In the present study, PDGF-BB was
used to imitate ASO. The effect of PDGF-BB treatment on miR-125b
expression in VSMCs was investigated. The results demonstrated that
miR-125b expression was significantly decreased following treatment
with PDGF-BB, the effects of which were reversed following
transfection with the miR-125b mimic. The present study used
PDGF-BB to treat cells to determine the effect of the miR-125b
mimic on the migratory and proliferative abilities of VSMCs. The
CCK-8 and BrdU assays were performed to assess cell proliferation
following treatment with PDGF-BB and transfection with miR-125b
mimic. The results demonstrated that miR-125b mimic significantly
inhibited cell proliferation. Furthermore, the results of the
Transwell and wound healing assays demonstrated that miR-125b mimic
decreased cell migration. Taken together, these results suggested
that miR-125b serves an important role in ASO, which is consistent
with previous findings (8).
Previous studies have demonstrated that miR-125b
expression is downregulated in a variety of tumor cells, including
hepatocellular carcinoma, breast cancer, ovarian tumors, bladder
cancer and leukemia (14-18).
Downregulation of miR-125b enhances the cell proliferative and
invasive abilities. In ASO, miR-125b is one of the miRNAs that is
significantly downregulated (7). It
has been suggested that miR-125b can inhibit the proliferation and
migration of VSMCs, and the molecular mechanisms are associated
with the regulation of SRF or myosin 1E, respectively (7,19).
However, miRNAs and their targets are complex and diverse (20). There is no clear evidence that
miR-125b affects AAMP. AAMP is highly expressed in cancer cells,
and its high expression is associated with the poor prognosis of
patients with cancer (21). In the
present study, miR-125b mimic and inhibitor were used to assess the
effect of miR-125b expression levels on AAMP and SRF expression.
Western blot analysis demonstrated that the protein expression
levels of AAMP and SRF were significantly decreased following
transfection with miR-125b mimic, the effects of which were
reversed following transfection with the miR-125b inhibitor. The
results of the immunofluorescence assay were consistent with these
findings.
The results of the present study demonstrated that
PDGF-BB was a negative factor of miR-125b expression in VSMCs.
Furthermore, transfection with miR-125b mimic increased miR-125b
expression in VSMCs, and promoted the cell proliferative and
migratory abilities following treatment with PDGF-BB. miR-125b
mimic and miR-125b inhibitor inhibited and induced AAMP and SRF
expression, respectively. However, further studies are required to
determine whether AAMP and SRF are target genes of miR-125b, and
whether miR-125b has other target genes. Taken together, the
results of the present study suggested that miR-125b is a potential
therapeutic strategy for preventing the progression of ASO.
Acknowledgements
Not applicable.
Funding
Funding: The present study was supported by the Major Natural
Science Project of Universities in Anhui Province (grant no.
KJ2016SD38), the Key Project of Natural Science Foundation of
Bengbu Medical College in Anhui Province (grant no. BYKY1862ZD) and
the Scientific and technological research project of Anhui Province
(grant no. 201904a07020020).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
XW and YG designed the study. XW, SC, CY and ZN, YS,
RL and ZG performed the experiments. SC, CY, ZN and YS performed
the statistical assessments. XW, SC and YG wrote the manuscript.
XW, SC and YG confirm the authenticity of all the raw data. All
authors read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Howard MD, Hood ED, Zern B, Shuvaev VV,
Grosser T and Muzykantov VR: Nanocarriers for vascular delivery of
anti-inflammatory agents. Annu Rev Pharmacol Toxicol. 54:205–226.
2014.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Diehm C, Allenberg JR, Pittrow D, Mahn M,
Tepohl G, Haberl RL, Darius H, Burghaus I and Trampisch HJ: German
Epidemiological Trial on Ankle Brachial Index Study Group.
Mortality and vascular morbidity in older adults with asymptomatic
versus symptomatic peripheral artery disease. Circulation.
120:2053–2061. 2009.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Doran AC, Meller N and McNamara CA: Role
of smooth muscle cells in the initiation and early progression of
atherosclerosis. Arterioscler Thromb Vasc Biol. 28:812–819.
2018.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Woo CC, Liu W, Lin XY, Dorajoo R, Lee KW,
Richards AM, Lee CN, Wongsurawat T, Nookaew I and Sorokin V: The
interaction between 30b-5p miRNA and MBNL1 mRNA is involved in
vascular smooth muscle cell differentiation in patients with
coronary atherosclerosis. Int J Mol Sci. 21(11)2019.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Kumar S, Kim CW, Simmons RD and Jo H: Role
of flow-sensitive microRNAs in endothelial dysfunction and
atherosclerosis: Mechanosensitive athero-miRs. Arterioscler Thromb
Vasc Biol. 34:2206–2216. 2014.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Boucher JM, Peterson SM, Urs S, Zhang C
and Liaw L: The miR-143/145 cluster is a novel transcriptional
target of Jagged-1/Notch signaling in vascular smooth muscle cells.
J Biol Chem. 286:28312–28321. 2011.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Chen Z, Wang M, Huang K, He Q, Li H and
Chang G: MicroRNA-125b affects vascular smooth muscle cell function
by targeting serum response factor. Cell Physiol Biochem.
46:1566–1580. 2018.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Warnecke-Eberz U, Chon SH, Hölscher AH,
Drebber U and Bollschweiler E: Exosomal onco-miRs from serum of
patients with adenocarcinoma of the esophagus: Comparison of miRNA
profiles of exosomes and matching tumor. Tumour Biol. 36:4643–4653.
2015.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Johnson RC, Leopold JA and Loscalzo J:
Vascular calcification: Pathobiological mechanisms and clinical
implications. Circ Res. 99:1044–1059. 2006.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Furukawa K, Abumiya T, Sakai K, Hirano M,
Osanai T, Shichinohe H, Nakayama N, Kazumata K, Hida K and Houkin
K: Increased blood viscosity in ischemic stroke patients with small
artery occlusion measured by an electromagnetic spinning sphere
viscometer. J Stroke Cerebrovasc Dis. 25:2762–2769. 2016.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Togliatto G, Dentelli P, Rosso A, Lombardo
G, Gili M, Gallo S, Gai C, Solini A, Camussi G and Brizzi MF:
PDGF-BB carried by endothelial cell-derived extracellular vesicles
reduces vascular smooth muscle cell apoptosis in diabetes.
Diabetes. 67:704–716. 2018.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Bennett MR, Sinha S and Owens GK: Vascular
smooth muscle cells in atherosclerosis. Circ Res. 118:692–702.
2016.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Chapiro E, Russell LJ, Struski S, Cavé H,
Radford-Weiss I, Valle VD, Lachenaud J, Brousset P, Bernard OA,
Harrison CJ and Nguyen-Khac F: A new recurrent translocation
t(11;14)(q24;q32) involving IGH@ and miR-125b-1 in B-cell
progenitor acute lymphoblastic leukemia. Leukemia. 24:1362–1364.
2010.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Guan Y, Yao H, Zheng Z, Qiu G and Sun K:
miR-125b targets BCL3 and suppresses ovarian cancer proliferation.
Int J Cancer. 128:2274–2283. 2011.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Hirsch HA, Iliopoulos D, Joshi A, Zhang Y,
Jaeger SA, Bulyk M, Tsichlis PN, Liu XS and Struhl K: A
transcriptional signature and common gene networks link cancer with
lipid metabolism and diverse human diseases. Cancer Cell.
17:348–361. 2010.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Huang L, Luo J, Cai Q, Pan Q, Zeng H, Guo
Z, Dong W, Huang J and Lin T: MicroRNA-125b suppresses the
development of bladder cancer by targeting E2F3. Int J Cancer.
128:1758–1769. 2011.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Rajabi H, Jin C, Ahmad R, McClary C, Joshi
MD and Kufe D: MUCIN 1 oncoprotein expression is suppressed by the
miR-125b oncomir. Genes Cancer. 1:62–68. 2010.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Wang D, Gao B, Yue J, Liu F, Liu Y, Fu W
and Si Y: Exosomes from mesenchymal stem cells expressing miR-125b
inhibit neointimal hyperplasia via myosin IE. J Cell Mol Med.
23:1528–1540. 2019.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Liu B, Li J and Cairns MJ: Identifying
miRNAs, targets and functions. Brief Bioinform. 15:1–19.
2014.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Yao S, Shi F, Wang Y, Sun X, Sun W, Zhang
Y, Liu X, Liu X and Su L: Angio-associated migratory cell protein
interacts with epidermal growth factor receptor and enhances
proliferation and drug resistance in human non-small cell lung
cancer cells. Cell Signal. 61:10–19. 2019.PubMed/NCBI View Article : Google Scholar
|