Introduction
Liver injury after the onset of reperfusion is the
result of the interplay between different complex mechanisms, with
cellular damage occurring during both the ischemic and reperfusion
phases. Hepatic ischemia/reperfusion (I/R) causes major injury
following vascular occlusion during both liver surgery and liver
transplantation (1); however, its
pathogenesis is largely unknown. The destructive effects of I/R may
be triggered by the acute generation of reactive oxygen species
(ROS) and nitrogen species after reoxygenation, which cause a chain
reaction of cellular responses leading to inflammation, cell death
and, eventually, organ failure (2-4).
Several mediators, such as proinflammatory cytokines, ROS, adhesion
molecules, chemokines and excess nitric oxide (NO) contribute to
this injury (5). However, it was
previously demonstrated that reperfusion did not induce oxidative
stress, but sustained endoplasmic reticulum (ER) stress (ERS) in
the livers of rats subjected to traumatic-hemorrhagic shock
(6). The reason may be that the
model used in that study was produced by bleeding until
decompensation, followed by an inadequate or adequate reperfusion
phase.
The ER is an organelle with important signaling and
homeostatic functions, which is responsible for protein folding,
maturation and trafficking. ERS is a particular subcellular
pathological process involving an imbalance of homeostasis and ER
disorder. In the early stages of ERS, cells exhibit a protective
unfolded protein response that changes the cellular transcriptional
and translational programs to alleviate this process (7). ERS triggers an adaptive response
pathway, named the unfolded protein response (UPR) pathway, through
activation of pancreatic ER eukaryotic translation initiation
factor-2a kinase (PERK), inositol requiring enzyme-1 (IRE1) and
activating transcription factor-6 (ATF6). Several conditions,
including hypoxia, I/R injury, neurodegeneration, heart disease and
diabetes, may be associated with activation of ER-initiated cell
death pathways (8). In samples from
human livers subjected to I/R, UPR pathway activation was observed
(9). Therefore, ERS may play an
important role in hepatic I/R injury.
Endogenous hydrogen sulfide (H2S) is
increasingly being recognized as an important gaseous physiological
mediator synthesized from cysteine by cystathionine γ lyase (CSE)
and other naturally occurring enzymes. CSE is differentially
expressed in the cardiovascular, neuronal, immune, renal,
respiratory, gastrointestinal, reproductive, hepatic and endocrine
systems (10). H2S
serves important functions in cytoprotection, anti-inflammation,
nociceptive stimulation, regulation of insulin release and
longevity (11-13).
Acute administration of H2S, either prior to ischemia or
at reperfusion, ameliorates hepatic I/R injury through an
upregulation of intracellular antioxidant and antiapoptotic
signaling pathways (14). However,
whether ERS is involved in hepatic I/R injury remains unclear.
It was hypothesized that exogenous/endogenous
H2S inhibited ERS through limiting the extent of I/R
injury. The present study was undertaken to investigate the effects
of exogenous H2S on I/R injury using an in vivo rat
model. The aim was to determine whether exogenous H2S
can inhibit the expression of ERS- and cell apoptosis-related
genes, thereby leading to suppression of inflammatory reactions and
improvement of I/R damage, and whether inhibition of endogenous
H2S may result in aggravation of I/R injury.
Materials and methods
Animal model
In total, 48 Sprague-Dawley male rats, aged 10 weeks
(weighing 225-275 g) were supplied by The Animal Centre of Xining,
Qinghai Province and housed in groups of two to four in 40x60x30 cm
plastic cages with soft bedding under a 12-h day/night cycle. The
room temperature was maintained at 22±1˚C and relative humidity was
45-60%; water and food pellets were available ad libitum. Animal
studies were performed in compliance with the guidelines of animal
ethics of Qinghai University and were approved by the Ethics
Committee Qinghai University Affiliated Hospital (approval no.
20180152; Xining, China).
Experimental design and surgery
A total of 48 Sprague-Dawley rats were randomly
divided into four groups (n=12 group) as follows: Sham, I/R, I/R
preceded by NaHS (I/R-NaHS) and I/R preceded by
L-C-propargylglycine (PAG), a H2S inhibitor (I/R-PAG).
All surgeries were performed as previously described (15). All animals were anesthetized with 5%
isoflurane for induction and 2-3% isoflurane for maintenance in an
acrylic anesthesia chamber, and the liver was exposed through a
transverse abdominal incision. A 30-min period of hepatic warm
ischemia was introduced by cross-clamping the hepatoduodenal
ligament with a microvascular clamp (Pringle's maneuver) in the
rats, with the exception of those in the sham group. Subsequently,
the clamps were removed to allow reperfusion for 6 or 12 h before
sampling, in order to explore whether the I/R injury could be
reversed under the protective effect of H2S. The animals
of the I/R group were administered saline (1 ml/kg body weight)
intraperitoneally before closing the incision. The animals of the
I/R-NaHS group were administered a single dose (56 µmol/kg) of NaHS
intraperitoneally 30 min before closing the incision (the dosage
was set according to our preliminary experiment). To completely
inhibit endogenous H2S, the animals of the I/R-PAG group
were administered 50 mg/kg of PAG intraperitoneally 30 min before
and after closure of the incision. After the experiments were
completed, all the rats were euthanized with 8% isoflurane
inhalation for 30 min until breathing had stopped and their skin
became cyanotic.
Serum biochemical analysis
At the end of the reperfusion period, 100-µl blood
samples were collected from the aorta under anesthesia using
heparinized syringes. The rats were euthanized after blood sample
collection and liver samples were harvested. After 300 x g
centrifugation at 4˚C for 30 min, plasma was separated and stored
at -20˚C until analysis. Serum concentrations of alanine
aminotransferase (ALT; cat. no. E-BC-K235-M, Elabscience
Biotechnology, Inc.) were measured at 37˚C using standard enzymatic
techniques (Ektachem 700 analyzer; Kodak).
TUNEL assay
The harvested liver samples were snap-frozen in
liquid nitrogen and stored in -80˚C. Subsequently, the frozen liver
samples were cut into 5-µm sections, and in situ apoptosis was
detected using a TUNEL assay kit (Promega Corporation): Proteinase
K was used to incubate the liver tissue samples for at 4˚C 20 min,
before 50 µl terminal deoxynucleotidyl transferase incubation
buffer was utilized to incubate the samples for 1 h at 37˚C in the
dark after the samples were washed for three times with PBS, 0.5
µg/ml DAPI (Beyotime Institute of Biotechnology) was used to stain
the nucleus for 15 min at room temperature. The sections were then
dehydrated in 100% ethanol, cleared with xylene and mounted
coverslips cell-side down on clean glass using a
fluorescence-compatible mounting medium (Vectashield; Vector
Laboratories, Inc.). Cells displaying brown staining within the
nucleus were counted as apoptotic. The number of apoptotic cells
was counted by an investigator who was blinded to the group
assignment by examining three non-overlapping microscopic fields of
view under high-power magnification (x400) via a Zeiss confocal
Axioskop 2 Plus microscope (Carl Zeiss AG) and expressed as
percentage (16).
Western blot analysis
Ischemic hepatic lobes were dissected and
homogenized in a buffer [2% SDS, 50 mM DTT, 62.5 mM Tris (pH 6.8),
10% glycerol] containing 1 mM PMSF (Amresco, LLC). The samples were
then boiled for 5 min and centrifuged at 7,500 x g for 10 min at
4˚C. Supernatants were collected and protein concentrations were
determined using a Bradford Protein Assay kit (Beyotime Institute
of Biotechnology). Protein samples (35 µg) were then loaded and
separated on a 12% SDS-PAGE. Following transfer onto a
nitrocellulose membrane, the membranes were incubated with 5%
skimmed milk to block non-specific antigens at room temperature for
2 h and probed with rabbit polyclonal antibodies (all 1:1,000)
against ATF6 (cat. no. ab203119, Abcam), PERK (cat. no. ab65142,
Abcam), glucose-regulated protein (GRP)78 (ab21685, Abcam),
TNF-receptor-associated factor (TRAF)-2 (cat. no. ab62488, Abcam),
C/EBP homologous protein (CHOP; cat. no. ab11419, Abcam),
caspase-12 (cat. no. ab62463, Abcam) and GAPDH (cat. no. ab9485,
Abcam) overnight at 4˚C. Following incubation with 1:1,000 diluted
goat anti-rabbit IgG (H&L) secondary antibody (cat. no.
G-21234, Themo Fisher Scientific) for 2 h at room temperature, the
protein bands were developed using an enhanced chemiluminescence
kit (Amersham; Cytiva).
Reverse transcription-quantitative
(RT-q) PCR analysis
Total RNA was isolated from liver tissue using
TRIzol® reagent (Thermo Fisher Scientific, Inc.)
according to the manufacturer's instructions and reversely
transcribed using PrimeScript™ RT reagent kit (Takara Bio, Inc.) at
37˚C for 45 min and then 95˚C for 5 min. cDNA was used as the
template for the qPCR reaction. The PCR conditions were as follows:
Step 1, pre-denaturing at 95˚C for 30 sec; step 2, 40 cycles at
94˚C for 5 sec, followed by 60˚C for 30 sec. qPCR was performed
with SYBR™ Green Master Mix kit (cat. no. 4344463; Thermo Fisher
Scientific, Inc.) using AB 2900HT fluorescence ration PCR
instrument. The primers for qPCR amplification are listed in
Table I. For determination of
relative quantification, the expression levels of the genes were
calculated and expressed based on the 2-ΔΔCq method
(17). GAPDH was selected as an
internal control gene.
 | Table IPrimers for quantitative PCR
amplification. |
Table I
Primers for quantitative PCR
amplification.
| Gene | Primer sequence
(5'→3') | Expected size
(bp) | GenBank accession
no. |
|---|
| ATF6 | Forward:
GGCTTCCTCCAGTTGTTC | 177 | NM_001107196.1 |
| | Reverse:
GTGACAGGCTTCTCTTCC | | |
| PERK | Forward:
TGTCTTGGTTGGGTCTGATG | 215 | NM_031599.2 |
| | Reverse:
CCTTCTTGCGGATGTTCTTG | | |
| GRP78 | Forward:
TAATCAGCCCACCGTAAC | 193 | NM_013083.2 |
| | Reverse:
GTTTCCTGTCCCTTTGTC | | |
| TRAF2 | Forward:
CTCTTCTTCGTGGTGATG | 101 | NM_001107815.2 |
| | Reverse:
TGCTCTCGGTTGTTATGG | | |
| CHOP | Forward:
GGAGAAGGAGCAGGAGAATG | 176 | NM_001109986.1 |
| | Reverse:
GAGACAGACAGGAGGTGATG | | |
| Caspase-12 | Forward:
ACTGTCCGAGTCTGAGAAAC | 104 | NM_130422.1 |
| | Reverse:
AGTGGCTATCCCTTTGCTTGTG | | |
| GAPDH | Forward:
TGCCACTCAGAAGACTGTGG | 85 | AF106860.2 |
| | Reverse:
GGATGCAGGGATGATGTTCT | | |
Statistical analysis
Data are expressed as mean ± SEM of three separate
repeated experiments. Two-way ANOVA followed by Bonferroni's post
hoc test was performed to assess variation among experimental
groups. P<0.05 was considered to indicate statistically
significant differences. All analyses were performed with SPSS 16.0
(SPSS Inc.).
Results
H2S restores liver function
following I/R and inhibition of endogenous H2S results
in aggravation of I/R damage
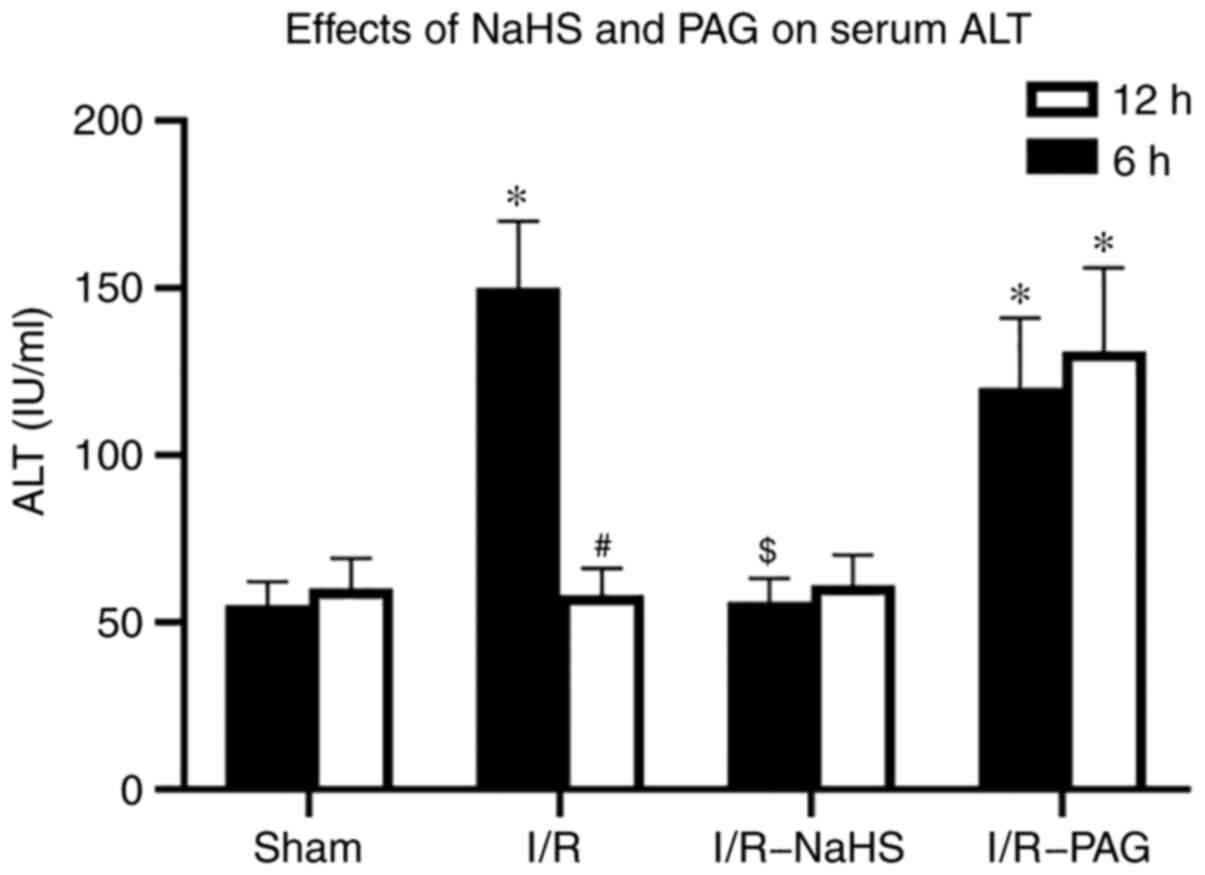
In order to assess hepatocellular damage due to I/R,
the serum ALT concentration was measured. A marked increase in the
ALT level was observed in the I/R group compared with that in the
Sham group after 6 h of reperfusion and the ALT level returned to
normal in the I/R group compared with that in the Sham group after
12 h reperfusion (Fig. 1).
Pretreatment with NaHS prevented the elevations in ALT levels 6 h
after I/R compared with those in the I/R group. A significant
increase in the ALT level was observed in the I/R-PAG group after 6
and 12 h of reperfusion compared with those in the Sham group.
These results indicated that endogenous H2S may have a
protective function in liver I/R, and exogenous H2S
ameliorates hepatic I/R injury.
H2S ameliorates cell
apoptosis in I/R and inhibition of endogenous H2S
results in aggravation of cell apoptosis
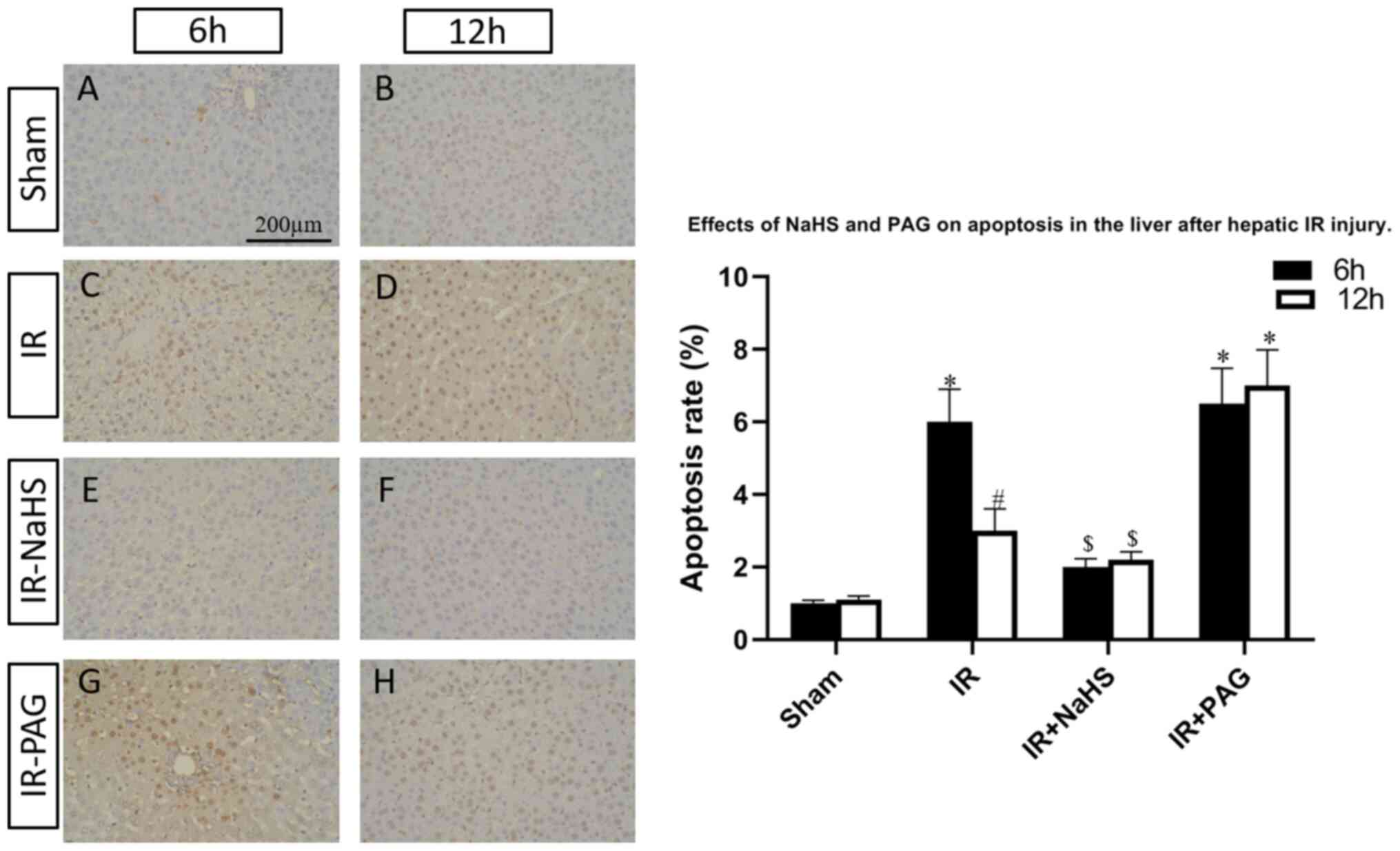
To investigate the functions of H2S
during liver I/R, the I/R-rats were pretreated with NaHS or the
endogenous H2S inhibitor, PAG. TUNEL assay was performed
to assess the percentage of apoptotic cells in the livers of the
rats of the four groups. As shown in Fig. 2, the hepatic cell apoptosis induced
by I/R was decreased by NaHS intervention; however, the apoptosis
rate was increased in the I/R-PAG group compared with the I/R group
after 6 and 12 h of reperfusion. Next, total protein and mRNA were
extracted from the livers and the expression levels of caspase-12,
which is an apoptosis-promoting protease, were detected in the four
groups. As shown in Figs. 3 and
4, caspase-12 was highly expressed
in the I/R and I/R-PAG groups, while no difference in caspase-12
expression was found between the I/R-PAG and I/R groups. More
importantly, NaHS intervention significantly alleviated I/R-induced
caspase-12 upregulation.
H2S inhibits ERS and
inhibition of endogenous H2S results in aggravation of
ERS induced by I/R injury
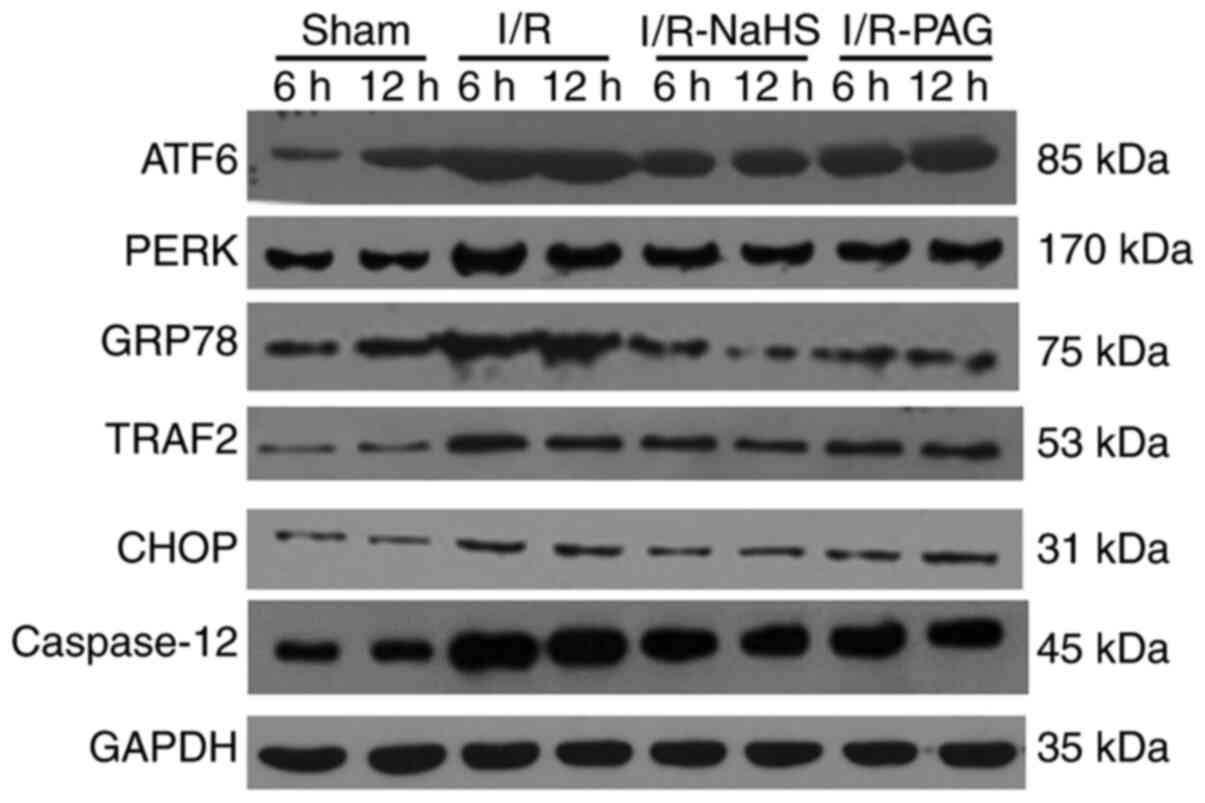
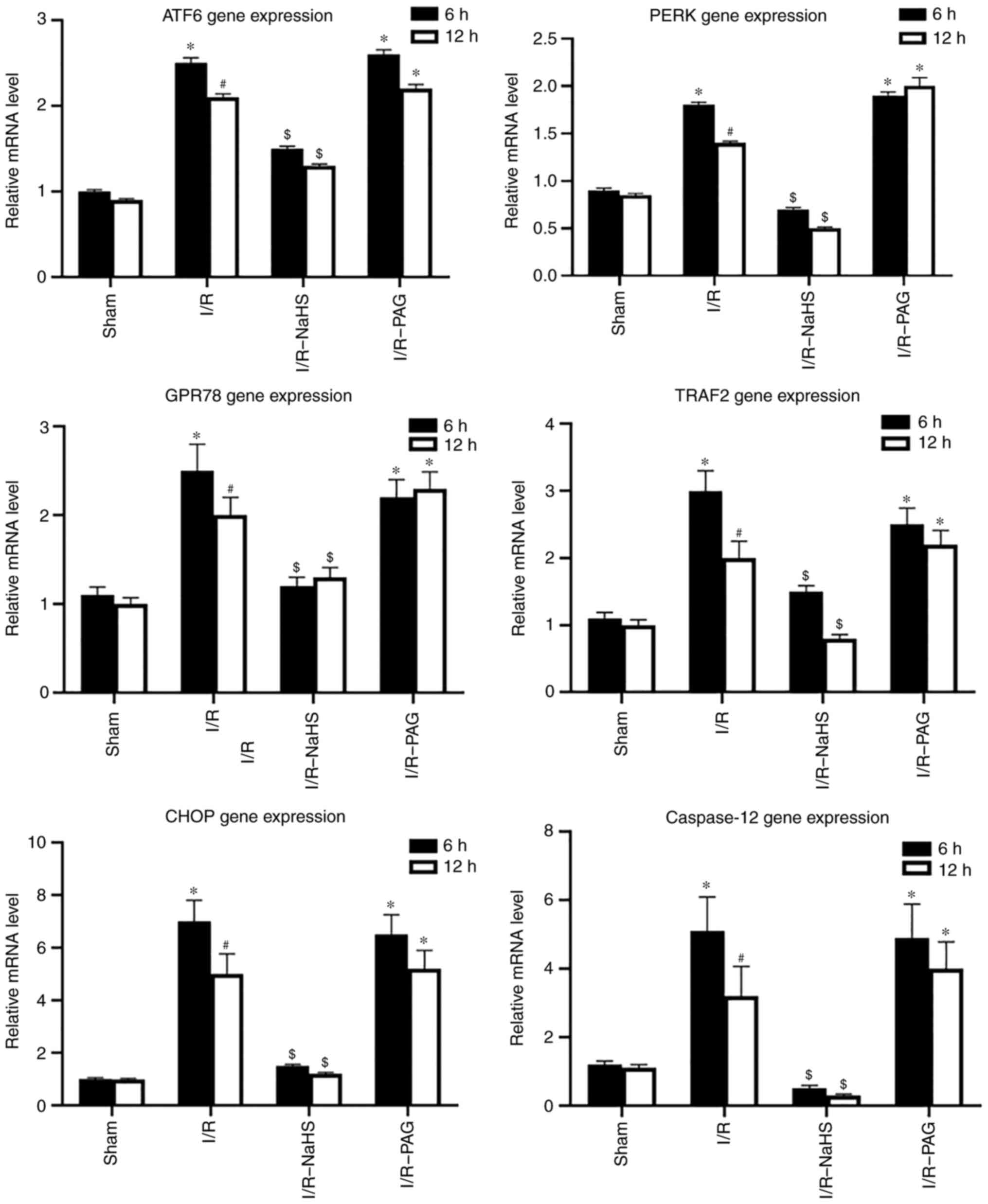
To investigate the molecular mechanism underlying
the protective effects of H2S, the protein and mRNA
levels of ERS-associated molecules were analyzed in liver tissue by
western blot and RT-qPCR assays. As shown in Figs. 3 and 4, the protein and mRNA levels of ATF6,
PERK, GRP78, TRAF2 and CHOP in the liver were markedly elevated by
I/R, whereas NaHS treatment attenuated the stimulatory effect of
I/R on ATF6, PERK, GRP78, TRAF2 and CHOP protein and mRNA levels.
PAG treatment exerted no obvious effect on the action of I/R on the
protein and mRNA levels of ATF6, PERK, GRP78, TRAF2 and CHOP
compared with the I/R group; therefore, it was inferred that
endogenous H2S may be involved in pathways other than
ERS-related pathways.
Discussion
The present study demonstrated that exogenous
H2S treatment, using NaHS as the H2S donor,
reduced I/R injury, whereas inhibition of endogenous H2S
by PAG treatment aggravated I/R injury in a hepatic I/R rat model.
In accordance with histological results, serum ALT measurement
demonstrated that the levels of ALT were elevated in the I/R and
I/R-PAG groups, indicating that H2S has a
hepatoprotective function against I/R. Similar with previous
findings, whereby Na2S was used as the H2S donor,
H2S attenuated the elevation of ALT after I/R (14).
The present study further demonstrated that the
protective effect of H2S was associated with its
attenuation of ERS induced by hepatic I/R injury, as the protein
and mRNA expression of ERS-associated molecules, including ATF6,
PERK, GRP78, TRAF2 and CHOP, were inhibited by NaHS treatment. A
previous study also demonstrated that H2S exerts
protective effects on the cardiovascular system, as it attenuates
cardiomyocytic ERS induced by hyperhomocysteinemia in rats
(18). ERS-induced apoptosis may
play a significant role in the pathophysiology of I/R injury
(19,20). ERS triggers the UPR pathway, which
has three branches, involving IRE1, PERK and ARF6(21). GRP78, an indicator of UPR pathway
activation also referred to as immunoglobulin heavy chain-binding
protein, is a central regulator of ER function due to its roles in
protein folding and assembly, targeting of misfolded proteins for
degradation, ER Ca2+ binding and controlling the
activation of transmembrane ERS sensors (21). Induction of GRP78 has been widely
used as a marker for ERS and the onset of the UPR. Due to its
antiapoptotic properties, stress induction of GRP78 represents an
important prosurvival component of the evolutionarily conserved UPR
(22). Active IRE1 recruits the
adaptor TRAF2, which activates apoptosis via the c-Jun N-terminal
kinase pathway (23,24). CHOP was originally identified in
response to DNA damage, and the CHOP level is a sensitive indicator
of ERS (23). Caspase-12 is an
enzyme that mediates ER-specific apoptosis and cytotoxicity
(25,26) and it was found to be highly
expressed in the I/R and I/R-PAG groups, while no difference in
caspase-12 expression was found between the I/R-PAG and I/R groups.
Therefore, caspase-12 may not play an important part in the process
of apoptosis that is caused by inhibition of endogenous
H2S. A previous study also demonstrated that
traumatic-hemorrhagic shock or reperfusion induces early and
persistent ERS of the liver or nitrosylative stress (6,27). The
present study demonstrated that NaHS treatment attenuated the
stimulatory action of I/R on ATF6, PERK, GRP78, TRAF2 and CHOP
protein and mRNA levels, which was consistent with the findings of
the aforementioned previous studies.
Endogenous H2S is a gaseous mediator
produced by CSE in numerous tissues, including the liver (28). The enzymatic activity of CSE was
found to be inhibited by PAG (29).
To determine the contribution of endogenous H2S to liver
protection against I/R, PAG was used to inhibit the production of
endogenous H2S. Low levels of H2S in the body
result in severe damage following I/R injury, with marked elevation
of ALT levels. However, the expression levels of ERS- and cell
apoptosis-related genes from I/R-PAG rats were similar with those
in I/R rats. Therefore, endogenous H2S may also be
involved in pathways other than ERS-related pathways.
In summary, the present study demonstrated that
exogenous H2S can effectively ameliorate I/R-induced
injury along with decreased protein and mRNA expression of
ERS-associated molecules, including ATF6, PERK, GRP78, TRAF2 and
CHOP. I/R was shown to induce ERS and activate ERS-related
apoptosis pathways, whereas inhibition of endogenous H2S
led to aggravation of damage after I/R and exogenous H2S
markedly reduced I/R injury. However, these conclusions were drawn
based on the results of the hepatic I/R rat model constructed in
the present study and they must be interpreted with caution, as the
underlying molecular mechanisms require further investigation.
Acknowledgements
Not applicable.
Funding
Funding: No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
HNF and XLW conceived the study. LC and KQM
participated in all the experiments; XLW and TSC analyzed the data;
LC drafted the manuscript. HNF, XLW and KQM have seen and confirm
the authenticity of the raw data. All authors have read and
approved the final version of this manuscript. All authors agree to
be accountable for all aspects of the work in ensuring that
questions related to the accuracy or integrity of any part of the
work are appropriately investigated and resolved.
Ethics approval and consent to
participate
Animal studies were performed in compliance with the
guidelines of animal ethics of Qinghai University and were approved
by the Animal Care Committee of Qinghai University (no.
20180152).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Jaeschke H and Woolbright BL: Current
strategies to minimize hepatic ischemia-reperfusion injury by
targeting reactive oxygen species. Transplant Rev (Orlando).
26:103–114. 2012.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Hines IN and Grisham MB: Divergent roles
of superoxide and nitric oxide in liver ischemia and reperfusion
injury. J Clin Biochem Nutr. 48:50–56. 2011.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Kupiec-Weglinski JW and Busuttil RW:
Ischemia and reperfusion injury in liver transplantation.
Transplant Proc. 37:1653–1656. 2005.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Urakami H, Abe Y and Grisham MB: Role of
reactive metabolites of oxygen and nitrogen in partial liver
transplantation: Lessons learned from reduced-size liver ischaemia
and reperfusion injury. Clin Exp Pharmacol Physiol. 34:912–919.
2007.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Abu-Amara M, Yang SY, Tapuria N, Fuller B,
Davidson B and Seifalian A: Liver ischemia/reperfusion injury:
Processes in inflammatory networks--a review. Liver Transpl.
16:1016–1032. 2010.PubMed/NCBI View
Article : Google Scholar
|
|
6
|
Duvigneau JC, Kozlov AV, Zifko C, Postl A,
Hartl RT, Miller I, Gille L, Staniek K, Moldzio R, Gregor W, et al:
Reperfusion does not induce oxidative stress but sustained
endoplasmic reticulum stress in livers of rats subjected to
traumatic-hemorrhagic shock. Shock. 33:289–298. 2010.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Deng Y, Srivastava R and Howell SH:
Endoplasmic reticulum (ER) stress response and its physiological
roles in plants. Int J Mol Sci. 14:8188–8212. 2013.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Xu C, Bailly-Maitre B and Reed JC:
Endoplasmic reticulum stress: Cell life and death decisions. J Clin
Invest. 115:2656–2664. 2005.PubMed/NCBI View
Article : Google Scholar
|
|
9
|
Emadali A, Nguyên DT, Rochon C, Tzimas GN,
Metrakos PP and Chevet E: Distinct endoplasmic reticulum stress
responses are triggered during human liver transplantation. J
Pathol. 207:111–118. 2005.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Wang R: Physiological implications of
hydrogen sulfide: A whiff exploration that blossomed. Physiol Rev.
92:791–896. 2012.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Kimura H: Hydrogen sulfide: From brain to
gut. Antioxid Redox Signal. 12:1111–1123. 2010.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Calvert JW, Coetzee WA and Lefer DJ: Novel
insights into hydrogen sulfide--mediated cytoprotection. Antioxid
Redox Signal. 12:1203–1217. 2010.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Kabil O and Banerjee R: Redox biochemistry
of hydrogen sulfide. J Biol Chem. 285:21903–21907. 2010.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Jha S, Calvert JW, Duranski MR,
Ramachandran A and Lefer DJ: Hydrogen sulfide attenuates hepatic
ischemia-reperfusion injury: Role of antioxidant and antiapoptotic
signaling. Am J Physiol Heart Circ Physiol. 295:H801–H806.
2008.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Tan EK, Shuh M, Francois-Vaughan H,
Sanders JA and Cohen AJ: Negligible oval cell proliferation
following ischemia-reperfusion injury with and without partial
hepatectomy. Ochsner J. 17:31–37. 2017.PubMed/NCBI
|
|
16
|
Zhang T, Ma Y, Xu KQ and Huang WQ:
Pretreatment of parecoxib attenuates hepatic ischemia/reperfusion
injury in rats. BMC Anesthesiol. 15(165)2015.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) Method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Wei H, Zhang R, Jin H, Liu D, Tang X, Tang
C and Du J: Hydrogen sulfide attenuates
hyperhomocysteinemia-induced cardiomyocytic endoplasmic reticulum
stress in rats. Antioxid Redox Signal. 12:1079–1091.
2010.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Georgiev P, Dahm F, Graf R and Clavien PA:
Blocking the path to death: Anti-apoptotic molecules in
ischemia/reperfusion injury of the liver. Curr Pharm Des.
12:2911–2921. 2006.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Jaeschke H and Lemasters JJ: Apoptosis
versus oncotic necrosis in hepatic ischemia/reperfusion injury.
Gastroenterology. 125:1246–1257. 2003.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Shore GC, Papa FR and Oakes SA: Signaling
cell death from the endoplasmic reticulum stress response. Curr
Opin Cell Biol. 23:143–149. 2011.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Lee AS: The glucose-regulated proteins:
Stress induction and clinical applications. Trends Biochem Sci.
26:504–510. 2001.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Szegezdi E, Logue SE, Gorman AM and Samali
A: Mediators of endoplasmic reticulum stress-induced apoptosis.
EMBO Rep. 7:880–885. 2006.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Tabas I and Ron D: Integrating the
mechanisms of apoptosis induced by endoplasmic reticulum stress.
Nat Cell Biol. 13:184–190. 2011.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Nakagawa T, Zhu H, Morishima N, Li E, Xu
J, Yankner BA and Yuan J: Caspase-12 mediates
endoplasmic-reticulum-specific apoptosis and cytotoxicity by
amyloid-beta. Nature. 403:98–103. 2000.PubMed/NCBI View
Article : Google Scholar
|
|
26
|
Szegezdi E, Fitzgerald U and Samali A:
Caspase-12 and ER-stress-mediated apoptosis: The story so far. Ann
N Y Acad Sci. 1010:186–194. 2003.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Obert DP, Wolpert AK and Korff S:
Modulation of endoplasmic reticulum stress influences
ischemia-reperfusion injury after hemorrhagic shock. Shock.
52:e76–e84. 2019.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Emerson B, Dransfield I, Morton NM and
Gray GA: Expression and localisation of H2S metabolising
enzymes in the murine myocardium and liver. Nitric Oxide.
27:S29–S30. 2012.
|
|
29
|
Han SJ, Kim JI, Park JW and Park KM:
Hydrogen sulfide accelerates the recovery of kidney tubules after
renal ischemia/reperfusion injury. Nephrol Dial Transplant.
30:1497–1506. 2015.PubMed/NCBI View Article : Google Scholar
|