Introduction
Smoke inhalation injury is one of the primary causes
of acute lung injury (ALI) or acute respiratory distress syndrome
(ARDS), affecting >50,000 individuals annually (1,2). In
our previous study, it was demonstrated that subsets of
CD4+ T lymphocytes, including regulatory T cells (Tregs)
and T-helper (Th) 17 cells, served critical roles in the
development of smoke inhalation-induced ALI (3). Puerarin, a compound used widely in
Traditional Chinese Medicine, was found to exert protective effects
against gunpowder smog-induced ALI by inhibiting Th17 responses
(4). However, the molecular
mechanisms by which puerarin exerted its effects require further
study.
Angiotensin (Ang) II is a key member of the
renin-angiotensin system (RAS), and it is involved in the
regulation of inflammation, proliferation and fibrosis in ALI
(5). Notably, research on
autoimmune diseases has demonstrated that Ang II and its related
signaling pathways, such as the NF-κB pathway, may be involved in
the regulation of CD4+ T lymphocytes. Blocking the
interaction of Ang II with Ang II type 1 receptor (AT1-R) resulted
in upregulation of antigen-specific Tregs and inhibited
autoreactive Th17 cells (6). A
recent study on colonic inflammation demonstrated that the local
RAS in the colon is activated in colitis and promotes colonic
inflammation by inducing Th17-cell activation (7). Ang II can induce the secretion of
interleukin (IL)-6, which together with TGF-β can then promote the
differentiation of Th17 cells (8,9).
It has been demonstrated that puerarin suppressed
murine hemangioendothelioma cell proliferation during Ang
II-induced aortic aneurysm formation by affecting the rate of
apoptosis (10). In addition, a
previous study has shown that puerarin attenuated Ang II-induced
cardiac fibroblast proliferation by increasing catalase activity
and inhibiting hydrogen peroxide-dependent Rac-1 activation
(11).
As it was demonstrated that puerarin may attenuate
Ang II-induced inflammation in other diseases, the present study
was undertaken to determine whether puerarin may alleviate
inflammation via the RAS and the NF-κB pathway in gunpowder
smog-induced ALI.
Materials and methods
Animals and reagents
Male Wistar rats, aged 8-10 weeks and weighing
290-310 g, were supplied by the Laboratory Animal Center of the
People's Liberation Army General Hospital (Beijing, China), and
raised in accordance with the National Institutes of Health on
Animal Care and Ethical Guidelines (4). Rats were raised in
pathogen-free cages, maintained at a temperature of
20-25°C and a relative humidity of 50-70%, and received
food and water ad libitum. All experimental procedures were
approved by the Ethics Committee of the Medical College of the
People's Liberation Army [approval no. SCXK (Beijing) 2012-0001].
Puerarin (50 mg/ml) was obtained from Chengdu Tiantaishan
Pharmaceutical Co., Ltd. (4).
Animal model and experimental
procedures
The rat model was established based on our previous
study (3). Using a self-made smoke generator, smoke was
generated from 10 g gunpowder. The electromagnetic heater was
turned off as soon as the flame burnt out. A total of two rats at a
time were exposed to the smoke for 8 min(3). The situation
of rats was monitored every 2 min.
A total of 40 rats were randomly divided into four
groups as follows: Control group (termed Con; saline administration
plus ambient air inhalation), puerarin control group (termed Pue;
puerarin administration plus ambient air inhalation), ALI group
(termed ALI; saline administration plus smoke inhalation), and the
puerarin treatment group (termed ALI + Pue; puerarin administration
plus smoke inhalation). The rats in the Con and ALI groups were
administered with normal saline (NaCl, 0.9%) intraperitoneally. The
rats in the ALI + Pue group were injected with puerarin
intraperitoneally (100 mg/kg) 30 min after smoke inhalation. In the
Pue group, puerarin was administered intraperitoneally (100 mg/kg)
without smoke inhalation. The dose of puerarin used was based on a
previous study (4). A total of 24 h
after smoke exposure, all the rats were anesthetized with sodium
pentobarbital intraperitoneally (50 mg/kg; Sigma-Aldrich; Merck
KGaA), dissolved in normal saline at a concentration of 10 mg/ml.
Specific criteria (humane endpoints) were used to determine when
rats should be euthanized. While under anesthesia, all the rats
were euthanized by exsanguination: 10 ml blood was collected from
the abdominal aorta of the rats (weighing 290-310 g) for further
analysis. Death was confirmed based on the arrest of the cardiac
and respiratory function (≥5 min). After sacrifice, the left lung
was prepared for collection of bronchoalveolar lavage fluid (BALF).
The right upper lobe of the lung was used for immunohistochemistry
analysis and the right lower lobe of the lung was stored at
-80˚C.
Measurement of arterial blood
gases
Arterial blood was collected from the abdominal
aorta using a 10 ml syringe containing heparinized saline. The
blood samples were immediately injected into an ABL700 blood gas
analyzer (Radiometer) to measure the pH and the partial gas
pressures of oxygen (PaO2) and carbon dioxide
(PaCO2).
BALF analysis
The experimental protocols used were performed as
described previously (4). The
supernatant was prepared for the cytokine assays.
Ang II determination
The lung tissue samples (100 mg each) were
homogenized in normal saline and centrifuged for 20 min at 10,000 x
g at 4˚C (3). Blood samples were
collected from the abdominal aorta, and serum was obtained by
centrifugation (1,000 x g for 10 min). The concentration of Ang II
in the lung homogenates and serum were determined using ELISA kits,
according to the manufacturer's protocol (cat. no. 1158/5; R&D
Systems, Inc.).
Cytokine assays
The concentrations of the proinflammatory cytokines
IL-6, IL-1β, IL-17A and TNF-α in the BALF were determined using
ELISA kits (cat. nos. R6000B, RLB00, DY8410-05 and RTA00,
respectively; R&D Systems, Inc.), following the manufacturer's
protocol. The absorbance was measured at 450 nm using a microplate
reader (BioTek Instruments, Inc.).
Immunohistochemistry
The lung tissue was fixed with 4% paraformaldehyde
(cat. no. P1110; Beijing Solarbio Science & Technology Co.,
Ltd.) for 12 h at 4˚C. Paraffin-embedded 5-µm sections of lung
tissues were deparaffinized in xylene and hydrated in a decreasing
series of ethanol solutions. The sections were blocked using 10%
normal goat serum (cat. no. SL038; Beijing Solarbio Science &
Technology Co., Ltd.) for 1 h at room temperature. The sections
were probed with specific primary antibodies in a humidified
chamber at 4˚C overnight and secondary horseradish peroxidase
(HRP)-conjugated antibodies at room temperature for 1 h. The
primary antibodies used were: Monoclonal mouse
anti-angiotensin-converting enzyme (ACE; cat. no. ab270712; 1:100),
rabbit monoclonal anti-ACE2 (cat. no. ab108252; 1:100) and rabbit
polyclonal anti-AT1-R (cat. no. ab124505; 1:100) (all from Abcam).
The secondary HRP-conjugated antibodies used were goat anti-mouse
or anti-rabbit IgG (cat. nos. ZB-2305 and ZB-2301, respectively;
1:2,000; Zhongshan Bio-Tech Co, Ltd.). Specific staining was
detected using the two-step streptavidin-peroxidase method
(Non-Biotin HRP Direction System; Zhongshan Bio-Tech Co, Ltd.). The
signals were developed using 3,3'-diaminobenzidine. Subsequently,
the sections were counterstained using Mayer's hematoxylin,
dehydrated, cleared in xylene and imaged using a light microscope
(x400 magnification; Carl Zeiss AG).
Western blotting
Lung tissues were homogenized using liquid nitrogen.
RIPA lysis buffer (Beyotime Institute of Biotechnology) containing
PMSF (Beyotime Institute of Biotechnology) was used for the lysis
of the cells, followed by centrifugation at 4˚C (10,000 x g for 15
min). Protein concentrations in lung tissues were determined using
a BCA protein assay kit (Novagen; Merck KGaA). Protein samples (50
µg) were loaded on a 10% SDS-gel, resolved using SDS-PAGE and
transferred to a PVDF membrane. The membrane was blocked using 5%
skimmed milk in Tris-buffered saline (10 mM Tris, 150 mM NaCl,
pH7.4) with 0.1% Tween-20 (TBST) at room temperature for 1 h. The
membrane was probed using specific primary antibodies at 4˚C
overnight and secondary HRP-conjugated antibodies at room
temperature for 1 h. The primary antibodies used were: Mouse
monoclonal anti-ACE (cat. no. ab270712; 1:100; Abcam), rabbit
monoclonal anti-ACE2 (cat. no. ab108252; 1:1,000; Abcam), rabbit
polyclonal anti-AT1-R (cat. no. ab124505; 1:800; Abcam) anti-GAPDH
(cat. no. ab9485; 1:1,000; Abcam), mouse monoclonal anti-NF-κB p65
(cat. no. 6956; 1:1,000; Cell Signaling Technology, Inc.), mouse
monoclonal anti-phosphorylated (p-) NF-κB p65 (cat. no. 13346;
1:1,000; Cell Signaling Technology, Inc.) and mouse monoclonal
anti-NF-κB inhibitor α (IκB-α; cat. no. 4814; 1:1,000; Cell
Signaling Technology, Inc.). The secondary HRP-conjugated
antibodies used were goat anti-mouse and anti-rabbit IgG (cat. nos.
PV9002 and PV9001, respectively; 1:2,000; Zhongshan Bio-Tech Co,
Ltd.). After washing with TBST, the signals were developed using an
enhanced chemiluminescence detection kit (cat. no. P0018S; Beyotime
Institute of Biotechnology), imaged using a GelDoc XR automated gel
imaging system (Bio-Rad Laboratories, Inc.) and analyzed using
Image Lab version 3.0 (Bio-Rad Laboratories, Inc.).
Statistical analysis
All data are presented as the mean ± standard error
mean of at least three independent experiments. Differences in the
means of the groups were compared using the Bonferroni post hoc
test following Kruskal-Wallis in SPSS version 17.0 (SPSS Inc.).
Dunn's test was used for comparing two groups. P<0.05 was
considered to indicate a statistically significant difference.
Results
Analysis of arterial blood gases
Following smoke inhalation, arterial blood gases,
including PaO2 and PaCO2 were significantly
altered (Fig. 1E). Hypoxemia
occurred during the early stage of ALI/ARDS. A total of 24 h after
smoke exposure, the rats developed hypoxemia and hypercapnia with
decreased PaO2 and increased PaCO2 compared
with the control group (Fig. 1E).
Puerarin administration improved the hypoxemic symptoms by
increasing PaO2 (Fig.
1E). However, the PaCO2 was not significantly
decreased in the puerarin treatment group compared with the ALI
group (Fig. 1E).
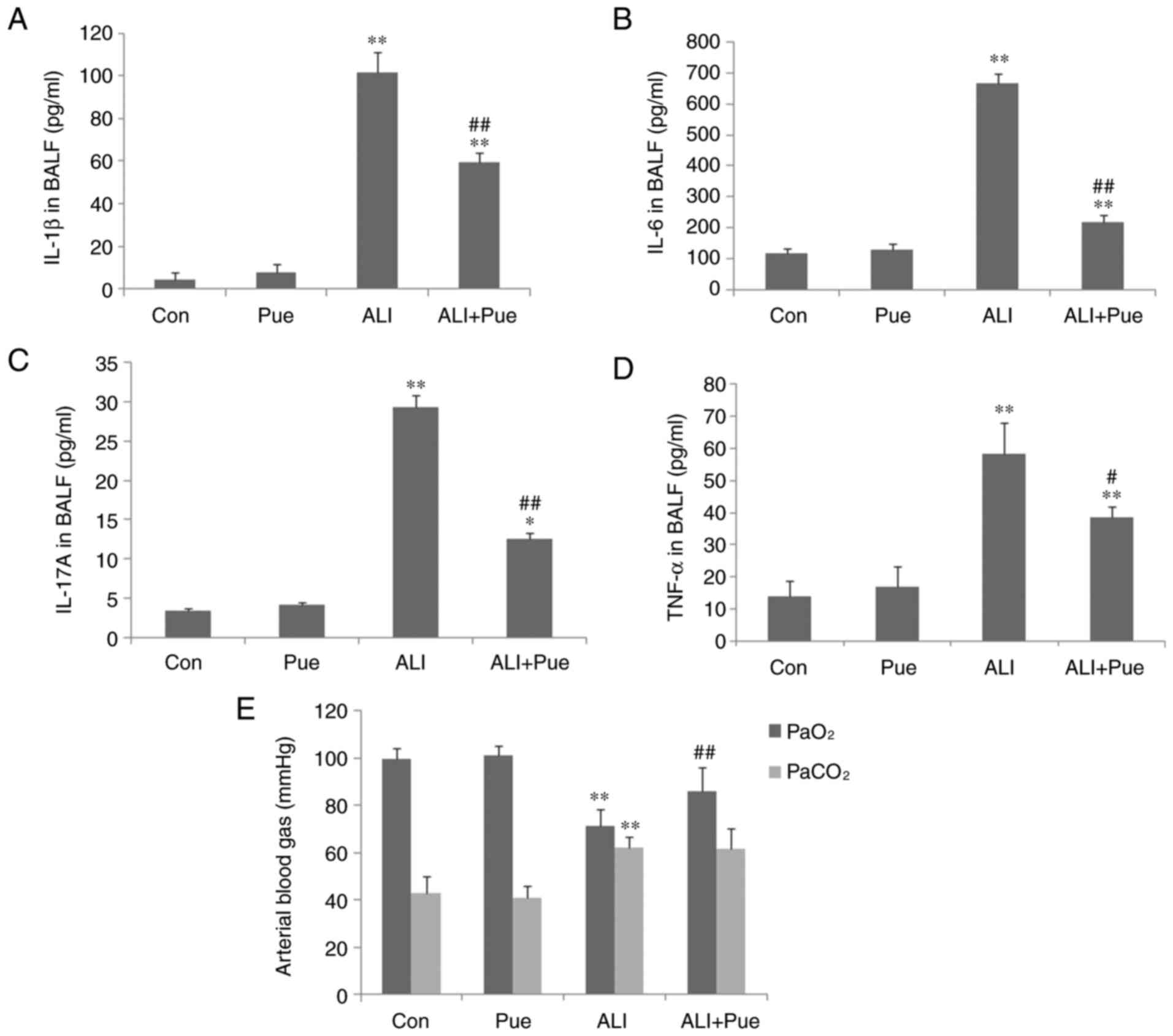 | Figure 1Cytokine levels in BALF and analysis
of arterial blood gases. (A) IL-1β, (B) IL-6, (C) IL-17A and (D)
TNF-α concentrations in BALF were determined using ELISA. (E)
Arterial blood gases, including PaO2 and
PaCO2 were analyzed. Data are presented as mean ±
standard error of the mean from three independent experiments (n=10
per group). *P<0.05, **P<0.01 vs. Con
group; #P<0.05, ##P<0.01 vs. ALI group.
BALF, bronchoalveolar lavage fluid; PaO2, partial gas
pressure of oxygen; PaCO2, partial gas pressure of
carbon dioxide; Con, normal control group; Pue, puerarin control
group; ALI, smoke inhalation group; ALI + Pue, puerarin treatment
plus smoke inhalation group. |
Proinflammatory cytokine levels in the
BALF
As shown in Fig.
1A-D, the levels of IL-6, IL-1β, IL-17A and TNF-α in the BALF
in the control groups were low. The levels of IL-6, IL-1β, IL-17A
and TNF-α in the BALF significantly increased following exposure to
smoke (Fig. 1A-D). Puerarin
treatment significantly reduced the levels of IL-6, IL-1β, IL-17A
and TNF-α compared with the ALI group, particularly those of IL-6
and IL-17A (Fig. 1A-D). This
suggests that puerarin may reduce Th17 cell levels by decreasing
the levels of related cytokines, including IL-6 and IL-17A. The
concentration of IL-6, IL-1β, IL-17A and TNF-α in the BALF was not
significantly different between the Con and Pue groups (Fig. 1A-D).
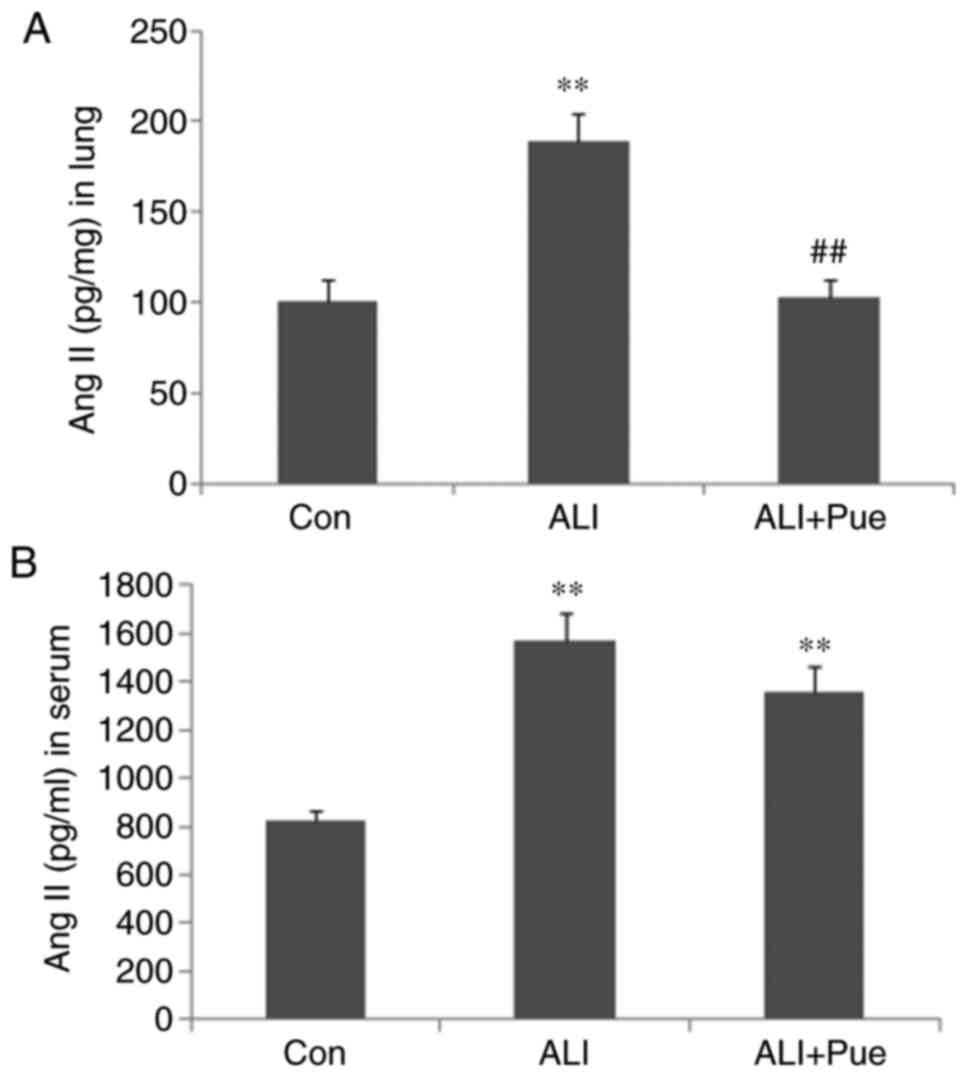
Ang II levels in the lung tissues and
circulation
To assess the effects of puerarin on Ang II
production, the Ang II levels in the lungs and circulation were
assayed. As shown in Fig. 2, the
levels of Ang II were significantly increased following smoke
inhalation in both the lung tissues and the peripheral blood of
rats. Puerarin treatment significantly reduced Ang II levels in the
lung tissues (Fig. 2A), but not in
the serum (Fig. 2B). Thus, the
suppression of activation of the local RAS in the lung may be more
important for the protective effects of puerarin, compared with the
RAS in the peripheral blood.
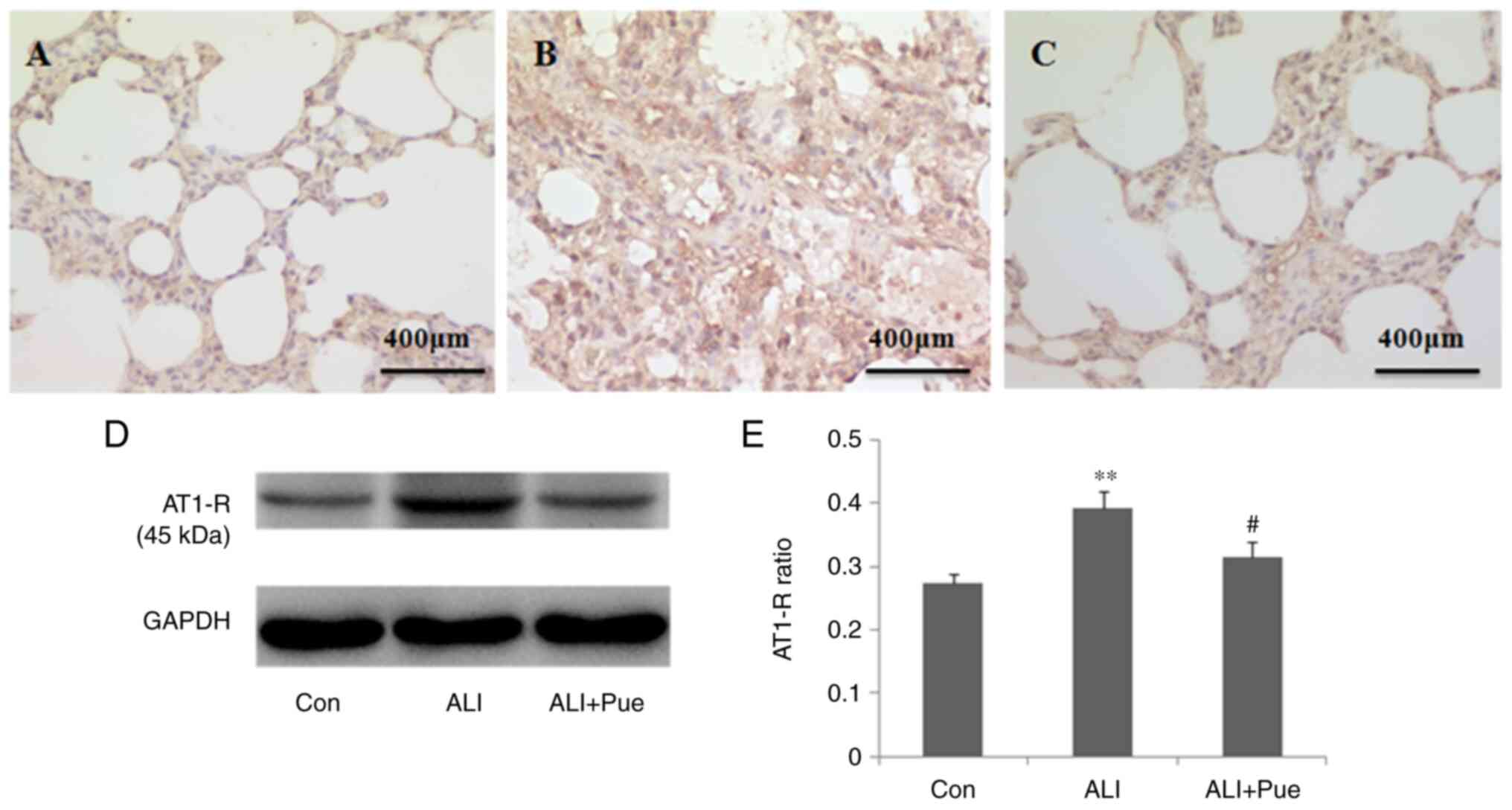
Effect of puerarin on the expression
of AT1-R
Immunohistochemical staining was used to determine
the localization of AT1-R in the lung tissue of rats. AT1-R was
expressed abundantly in inflammatory cells in the alveoli and
interstitial tissue (Fig.
3A-C).
The protein expression levels of AT1-R in the lung
tissues were quantified using western blotting. Fig. 3D shows representative images of the
immunoreactive bands (~45 kDa in weight). The protein expression
levels of AT1-R in the lungs were significantly increased 24 h
following smoke inhalation compared with those in the control group
(Fig. 3D and E). Of note, AT1-R protein expression
levels were decreased in the lungs following treatment with
puerarin compared with those in the ALI group (Fig. 3D and E).
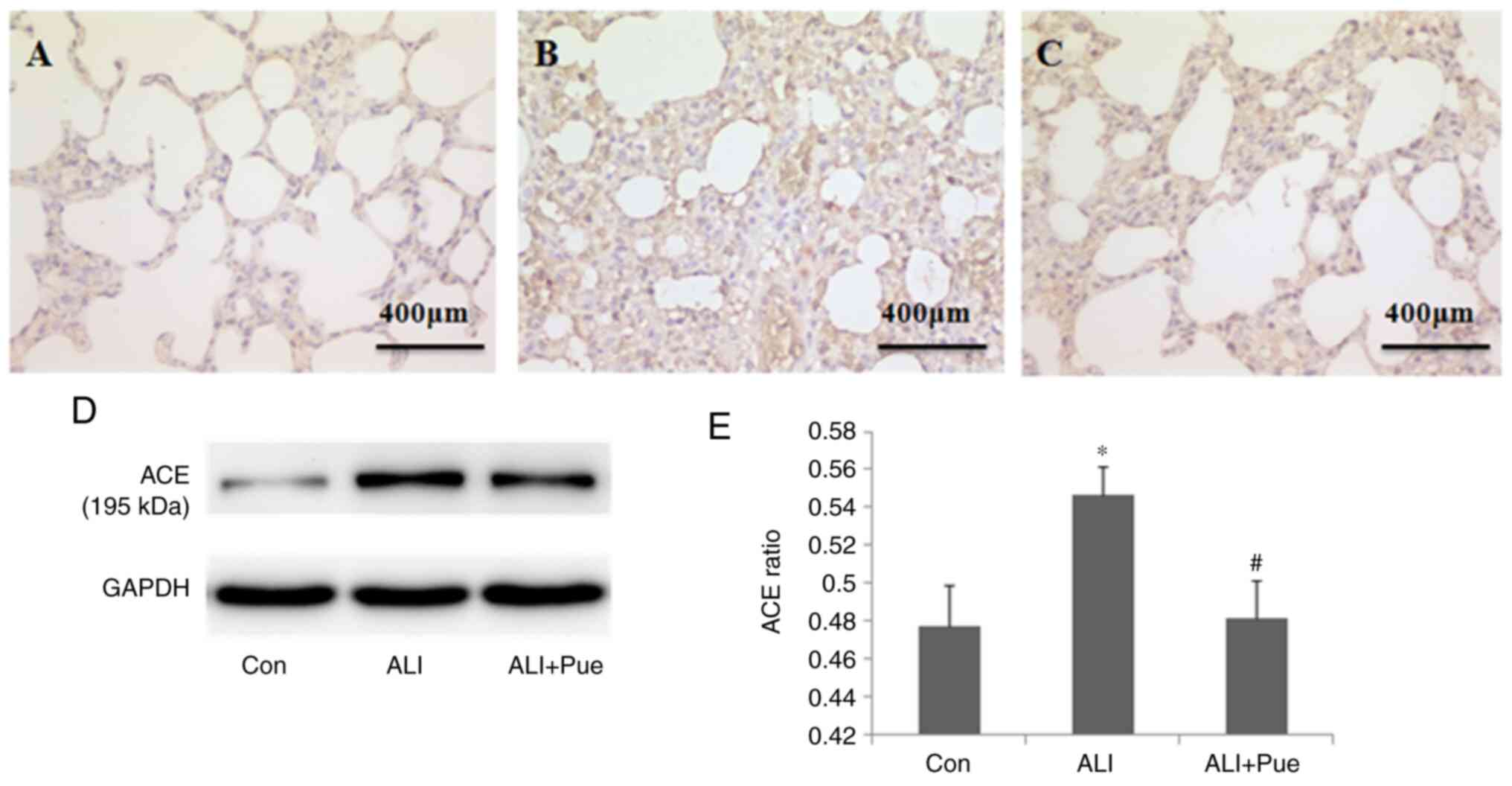
Effect of puerarin on the expression
of ACE
Immunohistochemical staining was used to determine
the localization of ACE in the lung tissues of rats. ACE expression
was evident in alveolar epithelial cells, as well as in endothelial
cells (Fig. 4A-C).
ACE expression in the lungs was quantified using
western blotting, and Fig. 4D shows
representative images of the immunoreactive bands (~195 kDa). The
protein expression levels of ACE in the lungs increased 24 h
following smoke inhalation compared with those in the control group
(Fig. 4D and E). Notably, ACE levels significantly
decreased in the lungs following puerarin administration compared
with those in the lungs after smoke inhalation alone (Fig. 4D and E).
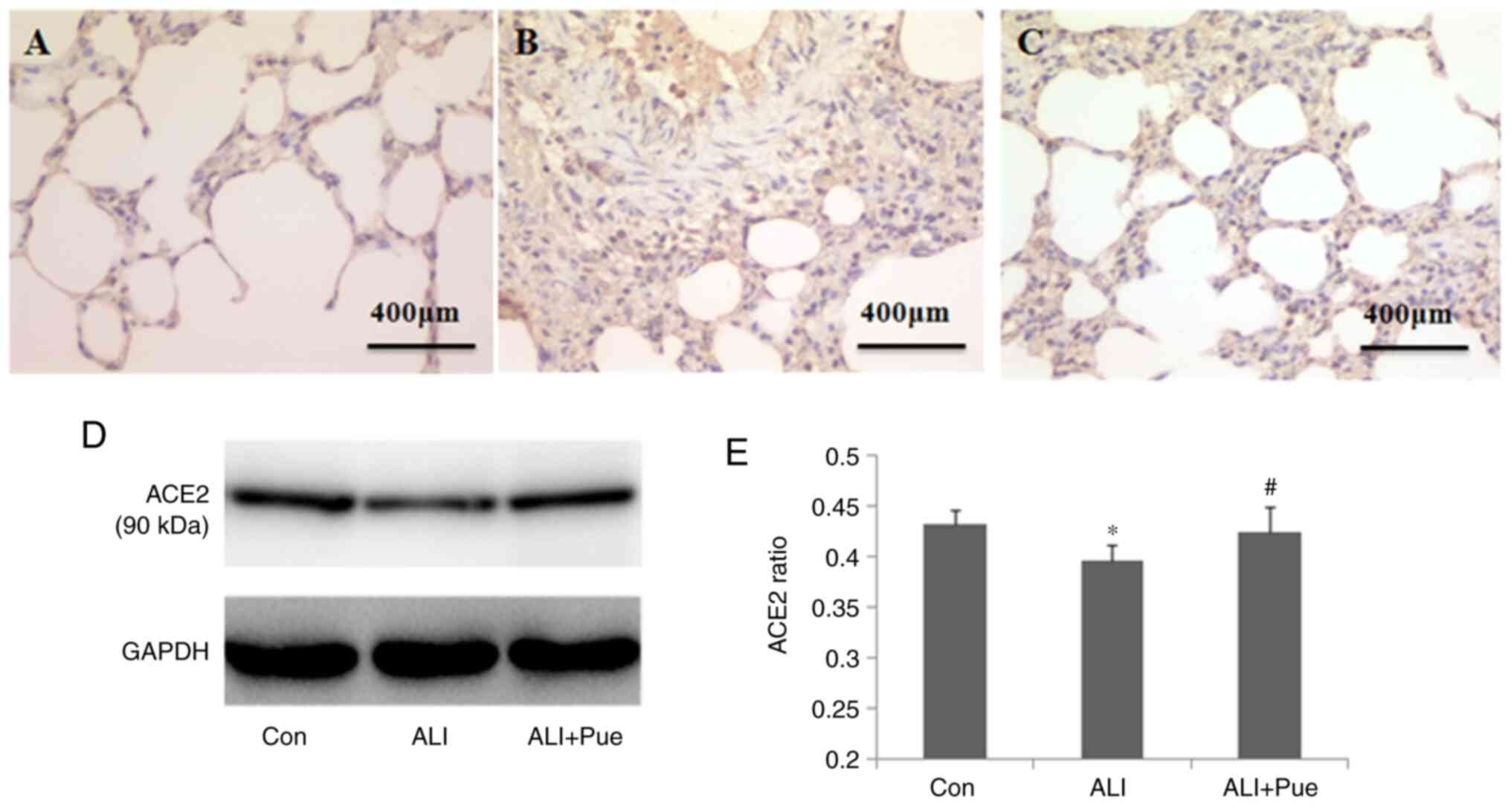
Effect of puerarin on the expression
of ACE2
Immuno-histochemical staining was used to determine
the localization of ACE2 in the lung tissues of rats. The major
sites of ACE2 expression were the alveolar epithelial and
endothelial cells (Fig. 5A-C).
The levels of ACE2 in the lungs were quantified
using western blotting, and Fig. 5D
shows representative images of the immunoreactive bands (~90 kDa).
The protein expression levels of ACE2 in the lungs decreased
following smoke inhalation compared with those in the control group
(Fig. 5D and E). Furthermore, the protein expression
levels of ACE2 increased in the lungs following administration of
puerarin compared with those in the lungs after smoke inhalation
(Fig. 5D and E).
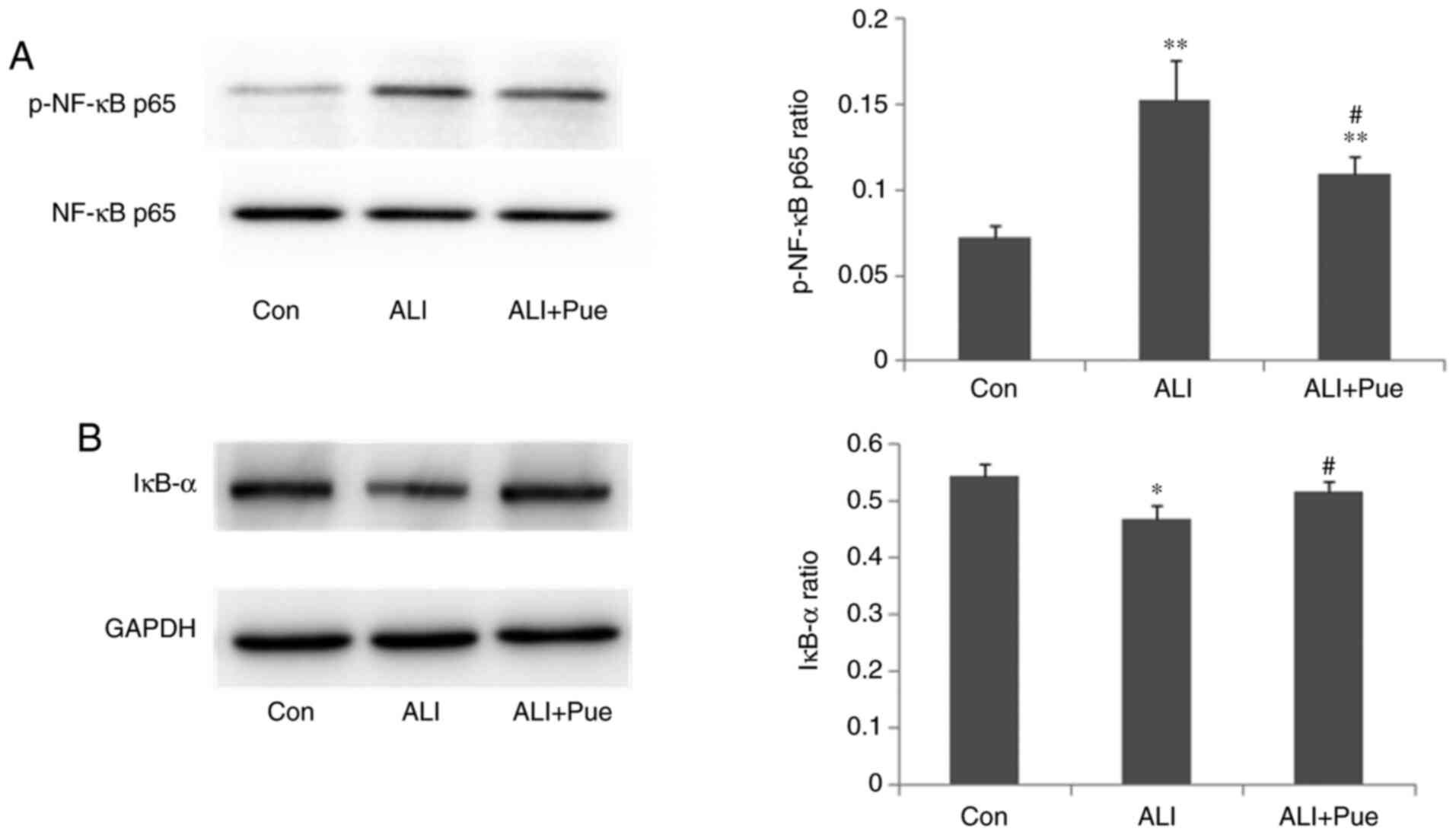
Effect of puerarin on the expression
of p-NF-κB p65 and IκB-α
The phosphorylation status of NF-κB p65 and the
expression levels of IκB-α in the lungs were quantified using
western blotting. The results revealed that the expression ratio of
p-NF-κB p65 relative to total NF-κB p65 in the lungs was
significantly increased 24 h following smoke inhalation compared
with that in the control group (Fig.
6A). Notably, NF-κB p65 phosphorylation was decreased in the
lungs following puerarin treatment compared with the ALI group
(Fig. 6A).
The protein expression levels of IκB-α in the lungs
decreased after smoke inhalation compared with those in the control
group (Fig. 6B). In addition, IκB-α
protein expression levels increased in the lungs following
administration of puerarin compared with that in the lungs after
smoke inhalation (Fig. 6B). Thus,
puerarin treatment inhibited the activation of the NF-κB pathway by
downregulating the phosphorylation of NF-κB p65 and upregulating
IκB-α expression.
Discussion
In our previous study, it was demonstrated that
puerarin attenuated inflammatory responses by inhibiting Th17
responses (4). However, the
underlying mechanisms involved were not determined. In autoimmune
diseases, it has been shown that the RAS and the NF-κB pathway may
contribute to the modulation of the levels of CD4+ T
lymphocytes (6). The RAS is
primarily composed of Ang II, AT1-R, ACE, ACE2 and Ang (1-7). ACE
and ACE2 share homology in their catalytic domains but have
different key functions in the RAS. ACE cleaves Ang I to generate
Ang II, whereas ACE2 is a negative regulator of the system, both
degrading the vasoconstrictor Ang II and producing the vasodilator
Ang (1-7) (12).
A previous study explored the effects of a
combination of felodipine + puerarin on the ACE2/Ang (1-7)/Mas
axis, to investigate the protective effects of this combination
against kidney damage in renovascular hypertensive rats. Compared
with the felodipine group, felodipine + puerarin significantly
attenuated fibrosis, decreased Ang II levels and increased Ang
(1-7) levels, upregulated the mRNA expression levels of ACE2 in the
bilateral kidneys, as well the levels of Mas in the ischemic
kidney, downregulated the levels of ACE in bilateral kidneys and
that of AT1-R in the ischemic kidney, and significantly decreased
the expression of TGF-β (13).
Therefore, it was hypothesized that puerarin may also exert its
anti-inflammatory effects by regulating the RAS and NF-κB pathways
in gunpowder smog-induced ALI.
In the present study, blood gas analysis
demonstrated that puerarin could reverse hypoxemia post-smoke
inhalation. However, levels of PaCO2 were unchanged by
puerarin treatment. Further studies should expand the experimental
scale to confirm the results and determine the underlying
mechanisms involved. In addition, puerarin significantly reduced
the levels of IL-6, IL-1β, IL-17A and TNF-α in the BALF, suggesting
that puerarin may attenuate cytokine-mediated inflammation.
However, the underlying mechanisms remain to be determined.
The RAS is intricately involved in maintaining blood
pressure homeostasis, as well as the balance of body fluids
(14). However, recent studies have
suggested that activation of the local RAS in the lungs may
contribute to the pathogenesis of ALI/ARDS via regulation of
inflammation, vascular permeability and fibroblast activity
(15). As a proinflammatory
mediator, Ang II is reported to influence the inflammatory process
(16-18).
Ang II is involved in the progression of lung injury via several
mechanisms, including activating inflammation in the lung, exerting
a proapoptotic effect on alveolar epithelial cells and promoting
fibrosis in ALI (19). The present
study revealed that puerarin significantly decreased Ang II levels
locally in the lung. However, the levels of Ang II in the
circulation were not notably reduced by puerarin. This result
suggested that the local RAS may have a pathogenic role in the
lungs, independently of the RAS in the circulation. Ang II can
promote inflammatory responses, including mononuclear cell
proliferation and chemotaxis, and the recruitment of
proinflammatory cells to the site of injury. Additionally,
inflammatory cells can produce Ang II, thus contributing to the
perpetuation of tissue damage (18). Ang II can induce IL-6 production,
which together with TGF-β can promote the differentiation of
IL-17A-producing Th17 cells (8,9).
Accordingly, the present study demonstrated that puerarin reduced
the concentration of inflammatory cytokines, including IL-6, IL-1β,
IL-17A and TNF-α, and this may be attributed to the reduced levels
of Ang II by puerarin.
The majority of the biological functions of Ang II
are mediated by binding to its receptor, AT1-R (20). In the present study, puerarin
suppressed the expression of AT1-R in lung tissue. In addition, it
has previously been shown that the Ang-II/AT1-R pathway induces the
activation of the NF-κB pathway and contributes to apoptosis of
alveolar epithelial cells in seawater inhalation-induced lung
injury (21). p-NF-κB p65 levels
reflect the activation of NF-κB signaling. IκB-α is the primary
subset of IκB, and the major function of IκB-α is to regulate
activation of NF-κB. In the present study, administration of
puerarin downregulated smoke inhalation-induced expression of
p-NF-κB p65 and upregulated the smoke inhalation-suppressed
expression of IκB-α, suggesting that puerarin may inhibit the
activation of NF-κB. NF-κB pathways are involved in the regulation
of proinflammatory cytokines, such as IL-6 and TNF-α (22).
Suppression of the NF-κB pathway is consistent with the reduced
levels of IL-6, IL-1β and TNF-α following treatment with
puerarin.
The lungs are the major organs that express Ang II
and ACE in humans and mice. The expression of ACE/Ang-II is
markedly elevated in patients with ARDS (23). ACE has been reported to be
upregulated in ARDS, and this is induced by pneumonia, trauma,
aspiration, and pancreatitis (24-26).
These previous studies demonstrated that ACE, Ang II and AT1-R
promoted disease pathogenesis, induced lung edema and impaired lung
function. In addition, mice deficient for ACE exhibited significant
improvement in the symptoms. Furthermore, studies using ACE2
knockout mice have demonstrated that ACE2 protects murine lungs
from ARDS (14). The negative
regulation of Ang II accounts, in part, for the protective
functions of ACE2 in ARDS. In the present study, puerarin reduced
the expression of ACE, whilst enhancing the levels of ACE2. These
results were consistent with the decreased levels of Ang II
following puerarin treatment. Therefore, puerarin may attenuate
inflammatory responses in gunpowder smog-induced ALI, at least in
part by upregulating the expression of ACE2 and downregulating that
of ACE, Ang II and AT1-R.
Cumulatively, the present study is the first to
report that the possible immunoregulative mechanisms of puerarin in
gunpowder smog-induced ALI may partly be attributed to inhibition
of expression of Ang II, AT1-R and ACE and promotion of ACE2.
However, as results in rats do not always translate in humans,
future studies should confirm these findings in samples from
patients with smoke inhalation-induced acute lung injury.
Additionally, larger animal cohorts in vivo and multiple
cell lines in vitro may be beneficial to further confirm the
related mechanisms.
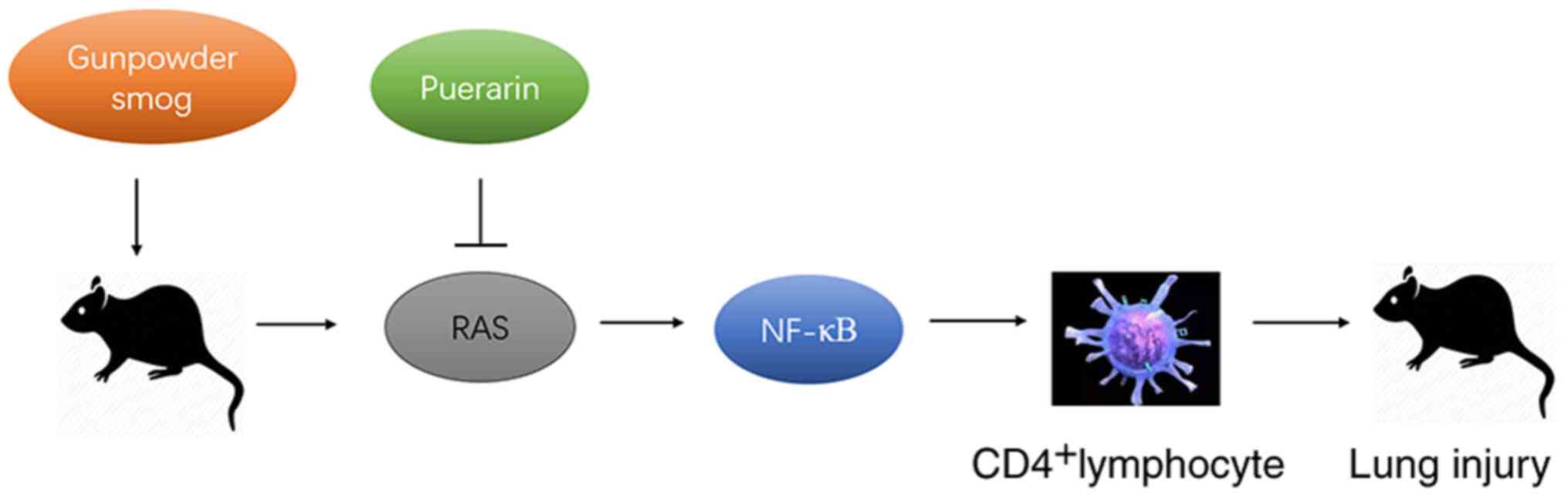
In conclusion, puerarin treatment suppressed the
smoke inhalation-induced expression of Ang II, AT1-R and ACE,
whilst enhancing the smoke inhalation-suppressed expression of
ACE2, together with the downregulation of NFκB. Therefore, puerarin
treatment significantly ameliorated the inflammation in gunpowder
smog-induced ALI, at least partly by regulating the RAS and NF-κB
pathways (Fig. 7). The present
findings provided evidence for its potential application in the
clinical management of smoke inhalation-induced acute lung injury
that accounts for extensive morbidity and mortality.
Acknowledgements
Not applicable.
Funding
Funding: The present study was supported by the National Natural
Scientific Foundation of China (grant no. 81700077), and the
Military Medical Scientific Research Project (grant no.
CWS12J021).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
FZ designed the project, wrote the manuscript and
performed the immunohistochemistry experiments. YW performed the
western blot analysis. PL and PD performed the measurement of
arterial blood gases. ML performed the cytokine assays. CW was
responsible for the design of the project, the revision of the
manuscript and performed some of the experiments. ML and CW are
responsible for confirming the authenticity of the raw data. All
authors read and approved the final manuscript.
Ethics approval and consent to
participate
All experimental procedures were approved by the
Ethics Committee of the Medical College of the People's Liberation
Army [approval no. SCXK (Beijing) 2012-0001].
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Fukuda S, Niimi Y, Andersen CR, Manyeza
ER, Rojas JD, Prough DS and Enkhbaatar P: Blood carboxyhemoglobin
elimination curve, half-lifetime, and arterial-venous differences
in acute phase of carbon monoxide poisoning in ovine smoke
inhalation injury model. Biochem Biophys Res Commun. 526:141–146.
2020.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Mokra D and Kosutova P: Biomarkers in
acute lung injury. Respir Physiol Neurobiol. 209:52–58.
2015.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Zhang F, Li MY, Lan YT and Wang CB:
Imbalance of Th17/Tregs in rats with smoke inhalation-induced acute
lung injury. Sci Rep. 6(21348)2016.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Zhang F, Wang Z, Li M, Lan Y, Chen Y and
Wang C: Puerarin attenuates smoke inhalation injury by regulation
of Th1/Th2 expression and inhibition of Th17 cells in rats. Int
Immunopharmacol. 28:546–553. 2015.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Tan WS, Liao W, Zhou S, Mei D and Wong WF:
Targeting the renin-angiotensin system as novel therapeutic
strategy for pulmonary diseases. Curr Opin Pharmacol. 40:9–17.
2018.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Platten M, Youssef S, Hur EM, Ho PP, Han
MH, Lanz TV, Phillips LK, Goldstein MK, Bhat R, Raine CS, et al:
Blocking angiotensin-converting enzyme induces potent regulatory T
cells and modulates TH1- and TH17-mediated autoimmunity. Proc Nat
Acad Sci U S A. 106:14948–14953. 2009.PubMed/NCBI View Article : Google Scholar
|
|
7
|
He L, Du J, Chen Y, Liu C, Zhou M,
Adhikari S, Rubin DT, Pekow J and Li YC: Renin-angiotensin system
promotes colonic inflammation by inducing TH17 activation via
JAK2/STAT pathway. Am J Physiol Gastrointest Liver Physiol.
316:G774–G784. 2019.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Ju X, Ijaz T, Sun H, Ray S, Lejeune W, Lee
C, Recinos A 3rd, Guo DC, Milewicz DM, Tilton RG and Brasier AR:
Interleukin-6-signal transducer and activator of transcription-3
signaling mediates aortic dissections induced by angiotensin II via
the T-helper lymphocyte 17-interleukin 17 axis in C57BL/6 mice.
Arterioscler Thromb Vasc Biol. 33:1612–1621. 2013.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Bettelli E, Carrier Y, Gao W, Korn T,
Strom TB, Oukka M, Weiner HL and Kuchroo VK: Reciprocal
developmental pathways for the generation of pathogenic effector
TH17 and regulatory T cells. Nature. 441:235–238. 2006.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Yue J, Chang SW, Xiao ZX, Qi YF and He JX:
The protective effect of puerarin on angiotensin II-induced aortic
aneurysm formation by the inhibition of NADPH oxidase activation
and oxidative stress-triggered AP-1 signaling pathways. Oncol Lett.
16:3327–3332. 2018.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Chen G, Pan SF, Cui XL and Liu LH:
Puerarin attenuates angiotensin II-induced cardiac fibroblast
proliferation via the promotion of catalase activity and the
inhibition of hydrogen peroxide-dependent Rac-1 activation. Chin J
Nat Med. 16:41–52. 2018.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Tipnis SR, Hooper NM, Hyde R, Karran E,
Christie G and Turner AJ: A human homolog of angiotensin-converting
enzyme. Cloning and functional expression as a
captopril-insensitive carboxypeptidase. J Biol Chem.
275:33238–33243. 2000.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Bai S, Huang ZG, Chen L, Wang JT and Ding
BP: Effects of felodipine combined with puerarin on ACE2-Ang
(1-7)-Mas axis in renovascular hypertensive rat. Regul Pept.
184:54–61. 2013.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Imai Y, Kuba K and Penninger JM: The
discovery of angiotensin-converting enzyme 2 and its role in acute
lung injury in mice. Exp Physiol. 93:543–548. 2008.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Marshall RP, Webb S, Bellingan GJ,
Montgomery HE, Chaudhari B, McAnulty RJ, Humphries SE, Hill MR and
Laurent GJ: Angiotensin converting enzyme insertion/deletion
polymorphism is associated with susceptibility and outcome in acute
respiratory distress syndrome. Am J Respir Crit Care Med.
166:646–650. 2002.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Miyoshi M, Nagata K, Imoto T, Goto O,
Ishida A and Watanabe T: ANG II is involved in the LPS-induced
production of proinflammatory cytokines in dehydrated rats. Am J
Physiol Regul Integr Comp Physiol. 284:R1092–R1097. 2003.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Schieffer B, Schieffer E, Hilfiker-Kleiner
D, Hilfiker A, Kovanen PT, Kaartinen M, Nussberger J, Harringer W
and Drexler H: Expression of angiotensin II and interleukin 6 in
human coronary atherosclerotic plaques: Potential implications for
inflammation and plaque instability. Circulation. 101:1372–1378.
2000.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Ruiz-Ortega M, Lorenzo O, Suzuki Y,
Rupérez M and Egido J: Proinflammatory actions of angiotensins.
Curr Opin Nephrol Hypertens. 10:321–329. 2001.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Marshall RP, Puddicombe A, Cookson WO and
Laurent GJ: Adult familial cryptogenic fibrosing alveolitis in the
United Kingdom. Thorax. 55:143–146. 2000.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Wang D, Chai XQ, Magnussen CG, Zosky GR,
Shu SH, Wei X and Hu SS: Renin-angiotensin-system, a potential
pharmacological candidate, in acute respiratory distress syndrome
during mechanical ventilation. Pulm Pharmacol Ther.
58(101833)2019.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Wang Y, Zhu Y, Zhu Y, Lu Z and Xu F:
Regulation of the angiotensin II-p22phox-reactive oxygen species
signaling pathway, apoptosis and 8-oxoguanine-DNA glycosylase 1
retrieval in hyperoxia-induced lung injury and fibrosis in rats.
Exp Ther Med. 13:3397–3407. 2017.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Li Q and Verma IM: NF-kappaB regulation in
the immune system. Nat Rev Immunol. 2:725–734. 2002.PubMed/NCBI View
Article : Google Scholar
|
|
23
|
Wenz M, Steinau R, Gerlach H, Lange M and
Kaczmarczyk G: Inhaled nitric oxide does not change transpulmonary
angiotensin II formation in patients with acute respiratory
distress syndrome. Chest. 112:478–483. 1997.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Imai Y, Kuba K, Rao S, Huan Y, Guo F, Guan
B, Yang P, Sarao R, Wada T, Leong-Poi H, et al:
Angiotensin-converting enzyme 2 protects from severe acute lung
failure. Nature. 436:112–116. 2005.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Zou Z, Yan Y, Shu Y, Gao R, Sun Y, Li X,
Ju X, Liang Z, Liu Q, Zhao Y, et al: Angiotensin-converting enzyme
2 protects from lethal avian influenza A H5N1 infections. Nat
Commun. 5(3594)2014.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Podowski M, Calvi C, Metzger S, Misono K,
Poonyagariyagorn H, Lopez-Mercado A, Ku T, Lauer T, McGrath-Morrow
S, Berger A, et al: Angiotensin receptor blockade attenuates
cigarette smoke-induced lung injury and rescues lung architecture
in mice. J Clin Invest. 122:229–240. 2012.PubMed/NCBI View
Article : Google Scholar
|