Introduction
The gut is the most important organ system for
metabolism and immunity, as well as a congenital barrier to prevent
pathogenic bacteria and toxins and other harmful substances from
passing through the intestinal tract into circulation and to ensure
homeostasis of the organism (1,2). At
present, several studies have confirmed that the development of a
variety of diseases is associated with impaired intestinal mucosal
barrier, such as severe pancreatitis, sepsis, inflammatory bowel
disease, diabetes, cardiovascular system diseases and acquired
immune deficiency syndrome (AIDS) (3-6).
Studies have demonstrated that increased intestinal epithelial cell
apoptosis, decreased expression of tight junction proteins,
increased intestinal permeability and other barrier injuries occur
in the chronic phase of human immunodeficiency virus (HIV)
infection (7-12).
AIDS is one of the most widespread and destructive
chronic infectious diseases to date (13-15).
Persistent intestinal barrier damage, which leads to an heterotopia
of microorganisms and induces a systemic inflammatory response, is
the main factor that promotes the progression of chronic HIV
infection to AIDS and leads to death (16). Therefore, it is important to perform
research on the mechanisms underlying intestinal barrier damage to
delay the progression of disease and prolong the life of patients.
Currently, no effective strategy is available to inhibit the
progression of gut barrier injury in patients with AIDS; thus, it
is urgent to identify potential therapeutic targets.
MicroRNAs (miRNAs/miRs) are noncoding,
single-stranded small RNA molecules of ~22 nucleotides that are
encoded by endogenous genes and are highly conserved in eukaryotic
cells (17). miRNAs serve an
important role in intestinal development, immune flora regulation,
and intestinal barrier damage and repair. Yang et al
(18) reported that miR-21 impaired
intestinal barrier integrity by downregulating the Ras homolog gene
family member B, and that miR-21 caused the disruption of tight
junctions in intestinal epithelial cells, resulting in reduced
transepithelial electrical resistance and increased mucosal
permeability. Zhao et al ((19) reported that miR-124 directly
targeted the aryl hydrocarbon receptor (AHR), which is inversely
correlated within inflamed intestinal epithelial cells. AHR
upregulation via the inhibition of miR-124 ameliorated experimental
colitis (19). Another study
demonstrated that inhibition of miR-146 attenuated
lipopolysaccharide (LPS)-induced intestinal mucosal cell apoptosis
(20). In addition, it was found
that miR-125b-5p contributed to HIV-1 latency in resting primary
CD4+ T lymphocytes (21). A
previous miRNA sequencing study from our group showed that multiple
miRNAs were abnormally expressed in the intestinal mucosa of
HIV/AIDS patients, including miR-125b-5p (22). However, the functions and mechanisms
of miR-125b-5p on intestinal mucosal cell injury remain
unclear.
Intestinal barrier damage is the main factor that
promotes the progression of AIDS. Glutamine transport in intestinal
mucosa is mainly dependent on the sodium-dependent amino acid
transport system, and alanine serine cysteine-preferring
transporter 2 (ASCT2) is a main factor involved in this system
(23). ASCT2 is one of the most
important glutamine transporters in organisms and regulates the
uptake of amino acids (24,25). Abnormal ASCT2 expression is
associated with multiple tumors, including breast cancer (26), colorectal cancer (27), hepatocellular carcinoma (28), gastric cancer (29), and ovarian cancer (30). Our previous work has demonstrated
that ASCT2 is abnormally low expressed in the damaged intestinal
mucosa of AIDS patients, suggesting that ASCT2 may be associated
with HIV-related intestinal mucosal barrier damage. However, the
mechanism by which ASCT2 may regulate intestinal barrier injury and
repair progression, especially HIV/AIDS progression, remains
unknown.
The present study predicted that ASCT2 may be a
candidate target of miR-125b-5p based on bioinformatics analysis.
It was then hypothesized that miR-125b-5p might target ASCT2 to
regulate intestinal barrier injury in patients with HIV/AIDS.
First, miR-125b-5p and ASCT2 expression was determined in the
intestinal mucosa of AIDS patients. Then, the potential role of
miR-125b-5p on the regulation of ASCT2 was evaluated in an
LPS-induced intestinal mucosa cell injury model in vitro. In
addition, the role of miR-125b-5p in the PI3K/AKT/mTOR pathway was
investigated, to further illuminate its regulatory mechanism.
Materials and methods
Clinical specimens and cell
culture
Colon biopsy samples from male patients with AIDS
and healthy subjects (both n=10; both aged 31-47 years) were
collected from the First Affiliated Hospital of Kunming Medical
University (Kunming, China) and used for RNA extraction. Patients
and healthy subjects were enrolled between June and December 2014.
Patients were diagnosed with AIDS according to the Guidelines for
the Diagnosis and Treatment of AIDS (31). The exclusion criteria for patients
were as follows: i) Patients with hepatitis C, hepatitis B or
syphilis; ii) patients with serious complications, such as
malignant fat tumor or cardiovascular disease and iii) patients
with systemic lupus erythematosus or rheumatoid arthritis. The
exclusion criteria for healthy subjects were as follows: i) Chronic
digestive disease and ii) abnormal routine physical and stool
examination. Human intestinal embryonic mucosa tissue-derived cells
(CCC-HIE-2) were purchased from the Cell Resource Center, Institute
of Basic Medicine, Chinese Academy of Medical Sciences. CCC-HIE-2
cells were cultured in DMEM/High Glucose medium (cat. no. 11965092;
Gibco; Thermo Fisher Scientific, Inc.) containing 10% FBS (cat. no.
S9020; Beijing Solarbio Science & Technology Co., Ltd.) at 37˚C
and 5% CO2. Cells were digested with 0.25% trypsin (cat.
no. 25200-072; Gibco; Thermo Fisher Scientific, Inc.) for 30 sec
and washed with 1X PBS (cat. no. P1020; Beijing Solarbio Science
& Technology Co., Ltd.). Then, 2x104 cells were
plated on a 10-cm2 cell dish with 10 ml medium and
cultured until the cells were in a good proliferation state.
LPS treatment of CCC-HIE-2 cells
CCC-HIE-2 cells (~5x105) were plated on a
10-cm2 cell dish with regular DMEM medium prior to
treatment. The next day, cells at 70% confluency were treated with
DMEM medium containing 20, 40, 80, 100 and 150 ng/ml LPS
(Sigma-Aldrich; Merck KGaA) to induce apoptosis. After 48 h, the
cells were collected, and untreated CCC-HIE-2 cells were used as a
control group. MTT assay and flow cytometry were performed to
investigate cell damage. Ultimately, 100 ng/ml was selected as the
optimum LPS concentration in subsequent experiments.
MTT assay
Induced or transfected cells were digested with
0.25% trypsin to create a single-cell suspension. Cells were
counted, and 2.5x104 cells were inoculated onto a
96-well plate for 24 h. The culture solution was aspirated with a
lance and carefully washed twice with PBS. A total of 100 µl of
fresh culture solution with a final concentration of 0.5 mg/ml MTT
(cat. no. M1020; Beijing Solarbio Science & Technology Co.,
Ltd.) were added to each well and cultured for 4 h. After the MTT
solution was discarded and 100 µl DMSO was added to each well, the
culture plate was placed on an oscillator and shaken for 10 min to
fully dissolve the purple crystals. The absorbance values of each
well were measured at 570 nm using a microplate reader (Bio-Rad
Laboratories, Inc.), and the cell survival rate was calculated.
Flow cytometry
An Annexin V-FITC Apoptosis Detection kit purchased
from Beyotime Institute of Biotechnology was used to analyze cell
apoptosis by flow cytometry. Briefly, 1x106 cells were
inoculated onto a 6-well plate and cultured for 24 h. Each sample
was prepared in triplicate. Following treatment with LPS for 48 h,
and transfections with ASCT2-overexpressing plasmid and/or
miR-125b-5p mimic/inhibitor for 24 h, the cells were harvested via
0.25% trypsin digestion, incubated with 10 µl Annexin V-FITC at 4˚C
for 30 min and then incubated with 5 µl propidium iodide (PI)
solution for 15 min in the dark. The stained cells were detected
using BD FACSCanto II flow cytometry (BD Biosciences) with
CellQuest software (BD Biosciences). The early apoptotic cells were
Annexin V-FITC-positive and PI-negative, and late apoptotic cells
were Annexin V-FITC-positive and PI-positive. The apoptotic rate of
cells is presented as the percentage of cells in early and late
apoptotic phases.
Cell transfection
CCC-HIE-2 cells were digested with 0.25% trypsin,
and cell suspensions (200 cells/µl) were prepared (5x106
cells in a 6-well plate). When the cells reached 70% confluency,
they were treated with LPS for 48 h, and then transfected with
different concentrations of miR-125b-5p mimic or inhibitor (cat.
nos. MC10148 and MH10148, respectively; Thermo Fisher Scientific,
Inc.), or 4 µg ASCT2/pEGFP plasmid with 6 µl Lipofectamine 2000
(Invitrogen; Thermo Fisher Scientific, Inc.) for 24 h, according to
the instructions of the manufacturer. After 6 h, the transfection
media were replaced with fresh DMEM/High glucose (cat. no.
11965092; Gibco; Thermo Fisher Scientific, Inc.) containing 10%
fetal bovine serum (cat. no. S9020; Beijing Solarbio Science &
Technology Co., Ltd.). miRNA mimics and inhibitors are single-chain
oligonucleotides that are used to investigate gain-of-function or
loss-of-function effects, respectively. ASCT2 gene was inserted
into pEGFP plasmid with restriction enzyme cutting site KpnI
and XhoI. ASCT2/pEGFP plasmid was sequenced by Shanghai
Sangon Biotech Co., Ltd. The sequences of miR-125b-5p mimic and
inhibitor are as follows: hsa-miR-125b-5p mimic,
5'-UCCCUGAGACCCUAACUUGUCA-3'; has-miR-125b-5p inhibitor,
UCACAAGUUAGGGUCUCAGGGA-3'; mimic negative control (NC),
5'-UUCUCCGACGUGUCACGUTT-3'; and inhibitor NC,
5'-CAGUACUUUUGUGUAGUACAA-3'.
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was extracted from samples using TRIzol
reagent (cat. no. 15596018; Invitrogen; Thermo Fisher Scientific,
Inc.), and miRNA was extracted using the miRNeasy Mini kit (cat.
no. 217004; Qiagen GmbH). After quantifying the concentration and
purity of RNA with a DyNA Quant 200 nucleic acid concentration
tester (GE Healthcare), the RNA was reverse transcribed into cDNA
according to the instructions of the PrimeScript RT Reagent kit
(cat. no. RR036Q; Takara Biotechnology Co., Ltd.), and miRNA was
reverse transcribed using the miRNA First-Strand Synthesis kit
(cat. no. 638313; Clontech Laboratories, Inc.) according to the
manufacturer's instructions. RT-qPCR was performed using TB Green
Premix Ex Taq (cat. no. RR420A; Takara Biotechnology Co., Ltd.) or
miRNA qRT-PCR TB Green kit (cat. no. 638314; Clontech Laboratories,
Inc.) on a 7500 Real-time PCR system (Applied Biosystems; Thermo
Fisher Scientific, Inc.). RT-qPCR was performed as follows: Initial
denaturation, 95˚C for 10 min, followed by 40 cycles of 95˚C for 15
sec and 60˚C for 60 sec. The results were analyzed using the
2-ΔΔCq method (32).
β-actin or U6 served as the internal controls for the mRNA or miRNA
qPCR assays, respectively. The following primers were used in the
present study: ASCT2, forward 5'-TGTGTAGAGGAGAA GAATGG-3' and
reverse 5'-GATGGCAAGAGTGAGGAC-3'; β-actin, forward
5'-GGCCTCCAAGGAGTAAGACC-3' and reverse-5'-AGGGGTCTACATGGCAACTG-3';
miR-125b-5p, forward 5'-TCCCTGAGACCCTAACTTGTGA-3' and reverse
5'-AGTCTCAGGGTCCGAGGTATTC-3'; and U6, forward
5'-CTCGCTTCGGCAGCACA-3' and reverse 5'-AACGCTTC
ACGAATTTGCGT-3'.
Western blot analysis
The cell culture medium was discarded. Cells were
washed with PBS, collected and lysed using RIPA lysis buffer (cat.
no. P0013B; Beyotime Institute of Biotechnology) for protein
extraction. The protein concentration was quantified using a BCA
kit (cat. no. P0012; Beyotime Institute of Biotechnology). Equal
amounts (30 µg) of protein lysates were separated by 12% SDS-PAGE
(cat. no. P0012A; Beyotime Institute of Biotechnology) and then
transferred onto PVDF membranes (cat. no. FFP36; Beyotime Institute
of Biotechnology). The membranes were blocked with 5% nonfat milk
(cat. no. P0216; Beyotime Institute of Biotechnology) for 2 h at
4˚C and then incubated with primary antibodies against ASCT2
(1:1,000; cat. no. ab237704; Abcam), GAPDH (1:2,000; cat. no.
ab9485; Abcam), phosphorylated (p-) AKT (1:1,000; cat. no. ab81283;
Abcam), AKT (1:1,000; cat. no. ab8805; Abcam), p-PI3K (1:1,000;
cat. no. ab191606; Abcam), PI3K (1:1,000; cat. no. ab40776; Abcam),
p-mTOR (1:1,000; cat. no. ab109268; Abcam) and mTOR (1:1,000; cat.
no. ab134903; Abcam) overnight at 4˚C. After washing with 1X PBST
(cat. no. P1031; Beijing Solarbio Science & Technology Co.,
Ltd.), the membranes were probed with HRP-conjugated goat
anti-rabbit IgG secondary antibodies (1:5,000; cat. no. ab7090;
Abcam) for 2 h at room temperature. Then, protein bands were
observed using an ECL chemiluminescence kit (cat. no. P0018S;
Beyotime Institute of Biotechnology) on the Bio-best type 140E Gel
Imaging Analysis system (SIM International Group Co., Ltd.). GAPDH
served as an internal control. ImageJ 2.x software was used to
quantify the protein bands (National Institutes of Health).
Dual luciferase reporter assay
The relationship between miR-125b-5p and ASCT2 was
examined using a Dual-Luciferase Reporter Assay kit (cat. no.
E1910; Promega Corporation). A wild-type (WT) or mutant (MUT)
version of the binding site in the 3'-untranslated region (3'-UTR)
of ASCT2 was cloned into the pmiRGLO vector (Promega Corporation).
Cells were co-transfected with the miR NC or miR-125b-5p mimic and
with the pmiRGLO-ASCT2-WT or pmiRGLO-ASCT2-MUT reporter plasmids by
Lipofectamine 2000 (cat. no. 11668-027; Invitrogen; Thermo Fisher
Scientific, Inc.). After 48 h of culture, the luciferase reporter
system was used to analyze the luciferase activity. The relative
luciferase activity of CCC-HIE-2 cells was expressed as the ratio
of firefly luciferase activity relative to sea kidney luciferase
activity.
Statistical analysis
Data are expressed as the mean ± standard deviation.
Differences between two groups were compared using the Student's
t-test. Differences among three or more groups were compared using
one-way ANOVA with Tukey's post hoc test. Statistical analyses were
performed with GraphPad Prism 5 software (GraphPad Software Inc.).
P<0.05 was considered to indicate a statistically significant
difference.
Results
miR-125b-5p and ASCT2 expression
levels in the intestinal mucosa of patients with AIDS
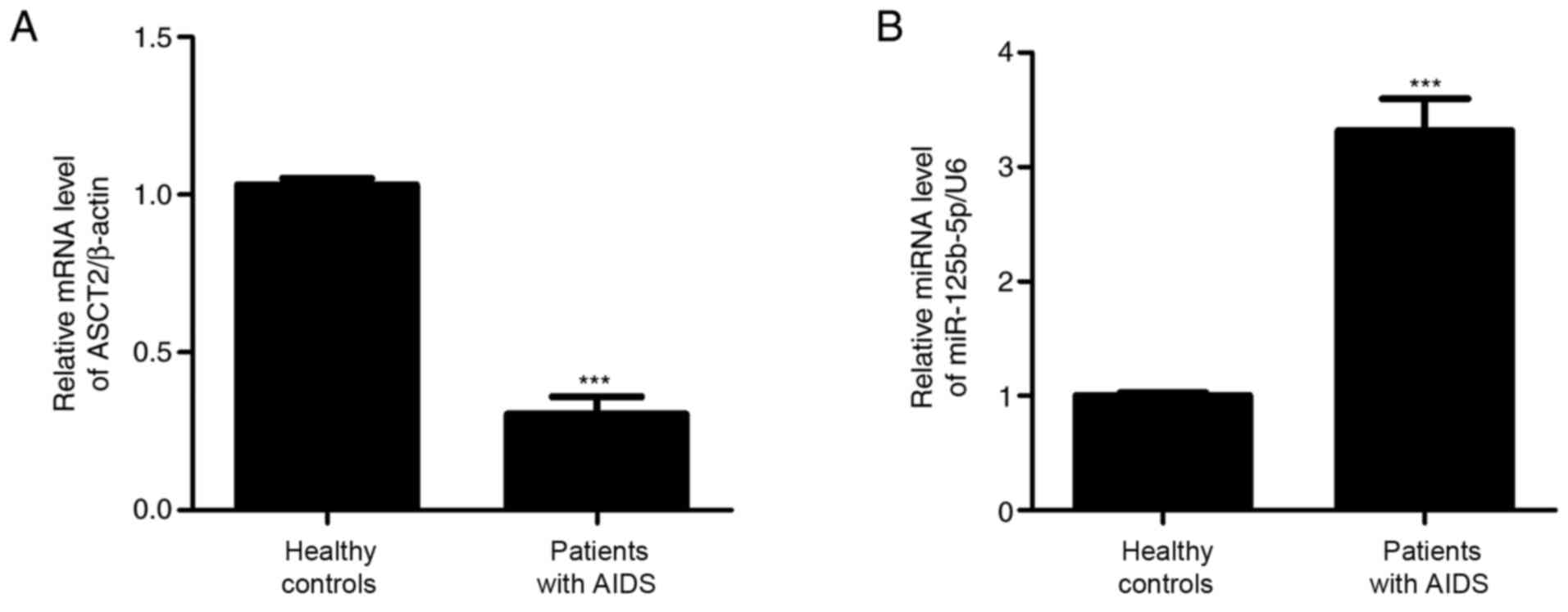
To measure miR-125b-5p and ASCT2 expression levels,
colon biopsy samples collected from 10 AIDS patients were examined
by RT-qPCR. As shown in Fig. 1,
miR-125b-5p expression levels were significantly increased, whereas
ASCT2 expression levels were significantly decreased in the AIDS
group compared with the control healthy group. These results
suggested that miR-125b-5p and ASCT2 may regulate intestinal
barrier injury in patients with AIDS.
miR-125b-5p and ASCT2 expression
levels in the LPS-induced intestinal mucosa cell injury model
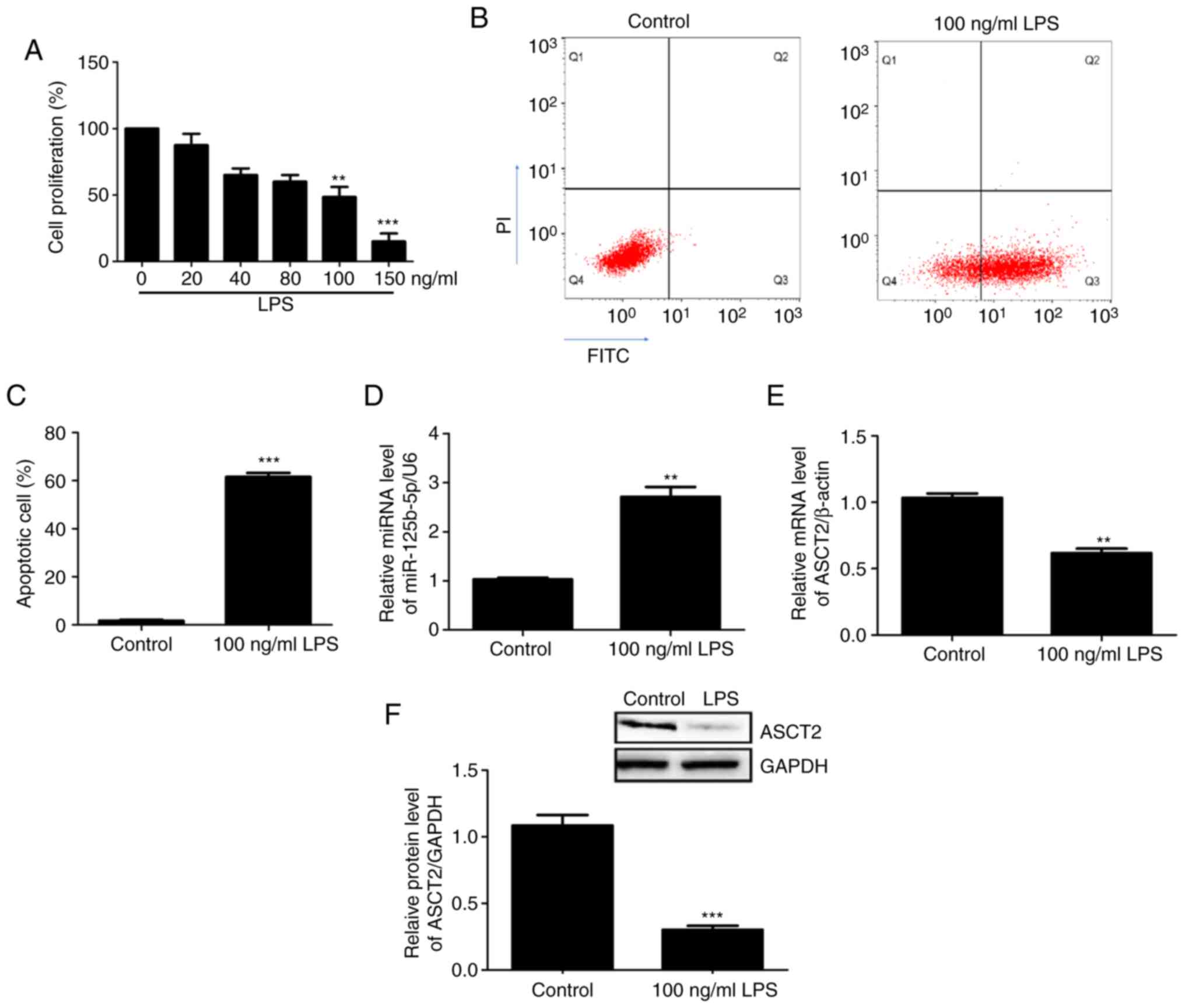
An in vitro intestinal mucosal cell injury
model was constructed using CCC-HIE-2 cells treated with different
concentrations of LPS. Cell proliferation was then measured using
the MTT assay. The results demonstrated that 100 ng/ml and 150
ng/ml LPS significantly inhibited CCC-HIE-2 cell proliferation
compared with 0 ng/ml LPS (Fig.
2A). Therefore, the 100 ng/ml LPS dose was used in subsequent
experiments. Apoptosis of the injured cells was detected by flow
cytometry. The results showed that cell apoptosis rates
significantly increased following treatment with LPS (100 ng/ml)
(Fig. 2B and C), suggesting that the in vitro
intestinal mucosa cell injury model was successfully established.
The expression levels of miR-125b-5p and ASCT2 in the LPS-injured
CCC-HIE-2 cells were detected by RT-qPCR and western blotting. The
RT-qPCR results revealed that miR-125b-5p expression levels were
significantly increased (Fig. 2D),
whereas ASCT2 expression levels were decreased (Fig. 2E) in LPS-injured CCC-HIE-2 cells
compared with control cells. Western blotting results showed that
the differences in ASCT2 protein levels were consistent with the
observed differences in mRNA levels (Fig. 2F). These results implied that the
in vitro intestinal mucosa cell injury model was
successfully established using LPS-induced CCC-HIE-2 cells.
Effect of miR-125b-5p on proliferation
and apoptosis of the LPS-induced intestinal mucosa cell injury
model
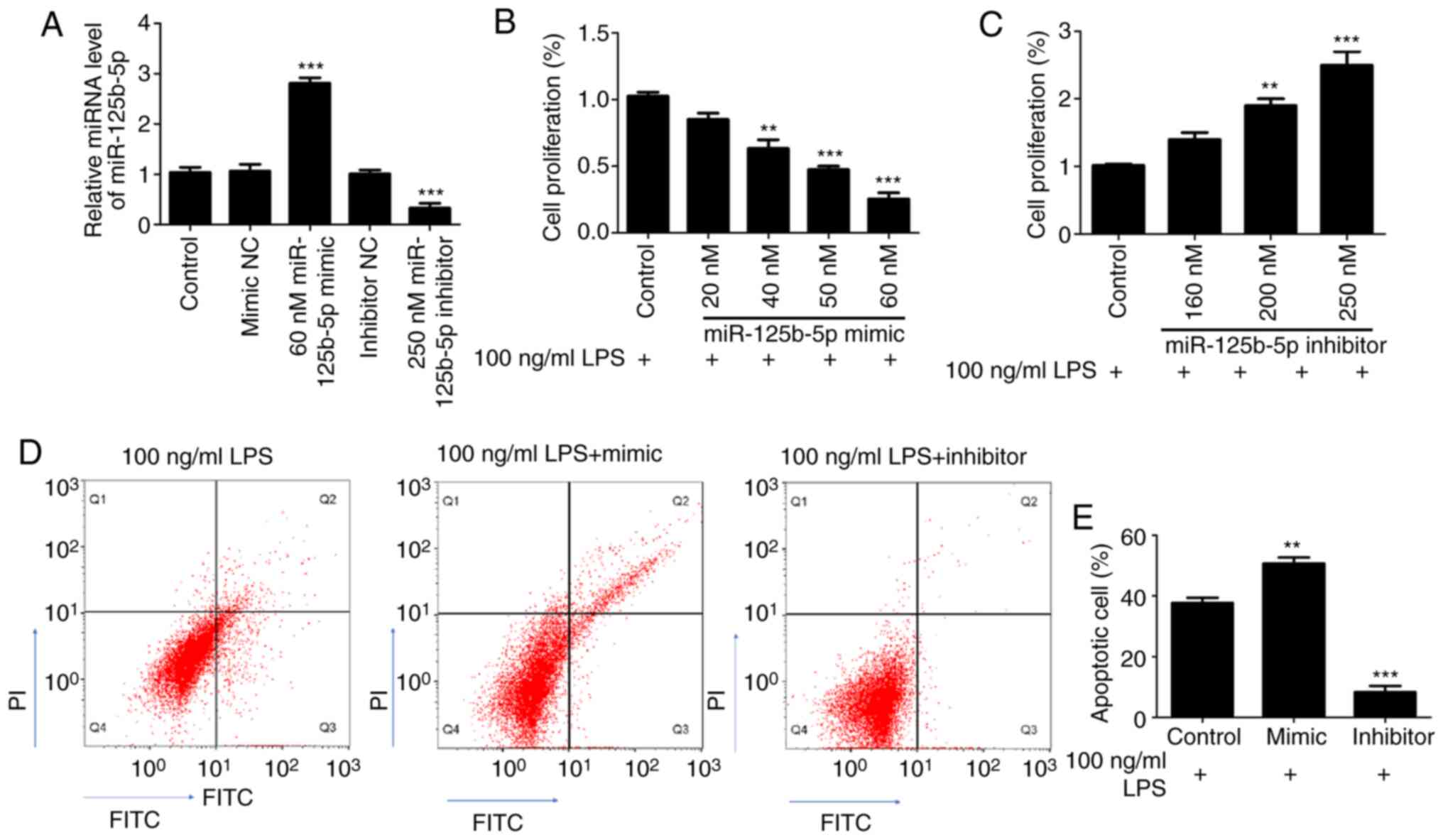
First, the transfection efficiency of miR-125b-5p
mimic and inhibitor was confirmed in CCC-HIE-2 cells. As shown in
the Fig. 3A, compared with their
respective control groups, the expression levels of miR-125b-5p
were significantly increased in the 60 nM miR-125b-5p mimic group,
and significantly decreased in the 250 nM miR-125b-5p inhibitor
group. There were no significant differences between the mimic NC
group, the inhibitor NC group and the untransfected cell control
group (Fig. 3A). Subsequently, to
investigate the effect of miR-125b-5p levels on LPS-induced
CCC-HIE-2 cell injury, LPS-treated CCC-HIE-2 cells were transfected
with the miR-125b-5p mimic or inhibitor, and MTT assays and flow
cytometry were performed to examine cell proliferation and
apoptosis. MTT assay results showed that the miR-125b-5p mimic
significantly inhibited LPS-treated CCC-HIE-2 cell proliferation in
a concentration-dependent manner (Fig.
3B). By contrast, the miR-125b-5p inhibitor significantly
increased LPS-treated CCC-HIE-2 cell proliferation in a
concentration-dependent manner (Fig.
3C). Therefore, the doses of 60 nM miR-125b-5p mimic and 250 nM
inhibitor were selected to perform the subsequent experiments. As
shown in Fig. 3D and E, transfection with the miR-125b-5p mimic
increased cell apoptosis, while transfection with the miR-125b-5p
inhibitor inhibited apoptosis, in the LPS-treated CCC-HIE-2 cell
model.
ASCT2 is negatively regulated by
miR-125b-5p
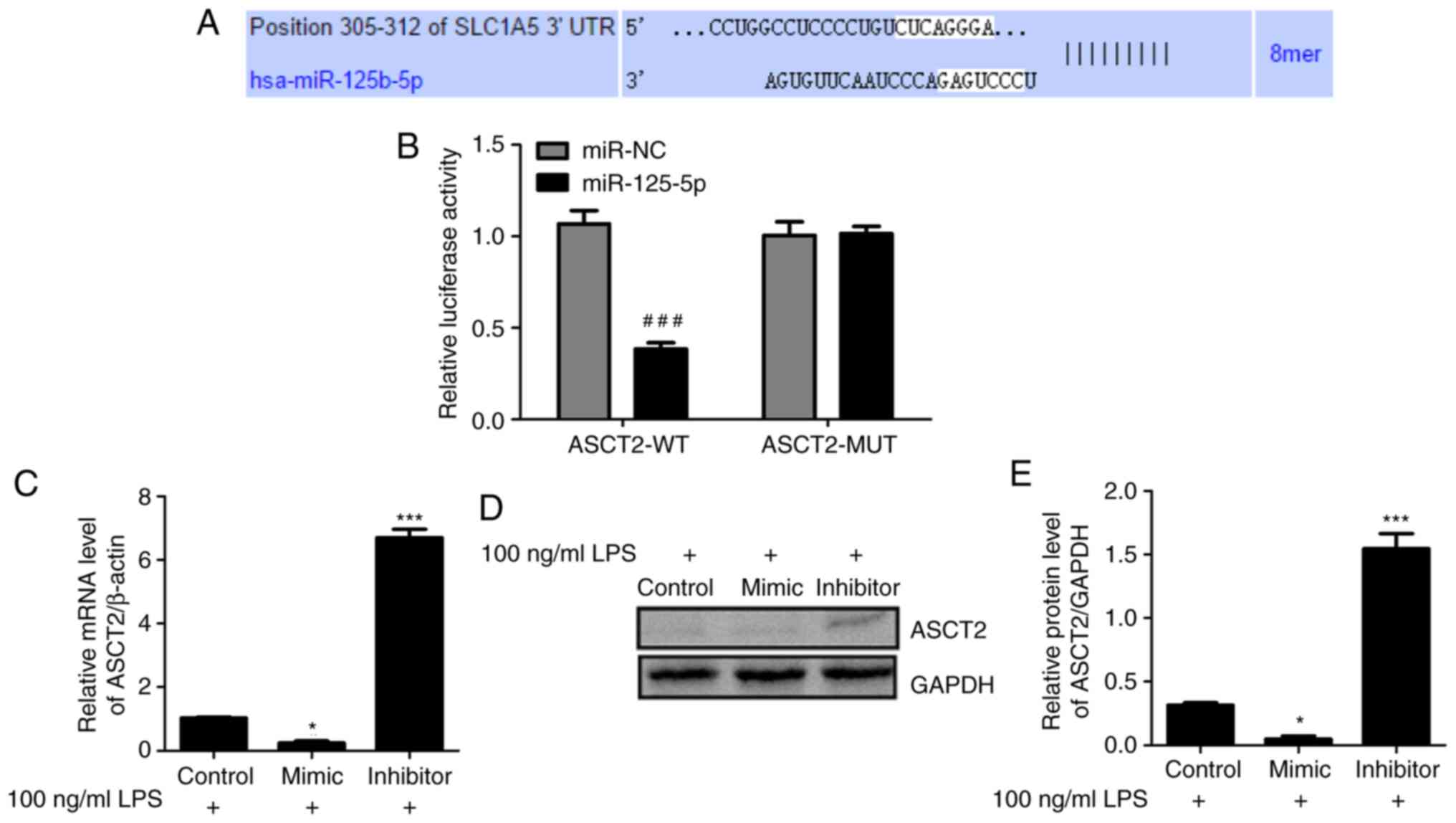
To explore the mechanism of miR-125b-5p in
LPS-induced intestinal mucosa cell injury, the potential targets of
miR-125b-5p were investigated using TargetScan (33). The TargetScan analysis predicted
that ASCT2 contains a sequence that is potentially complementary to
miR-125b-5p (Fig. 4A). To validate
this prediction, WT and MUT luciferase reporter vectors targeting
the 3'-UTR of ASCT2 were constructed and transfected into
LPS-treated CCC-HIE-2 cells. Dual luciferase reporter assays were
conducted to examine the relationship between ASCT2 and
miR-125b-5p. As shown in Fig. 4B,
luciferase activity was reduced by the miR-125b-5p mimic in cells
transfected with the ASCT2-WT reporter, whereas it was unchanged in
cells transfected with the ASCT2-MUT reporter. In addition, RT-qPCR
and western blot data indicated that the miR-125b-5p mimic
inhibited ASCT2 mRNA and protein expression levels, whereas the
miR-125b-5p inhibitor promoted ASCT2 mRNA and protein expression
levels in LPS-treated CCC-HIE-2 cells (Fig. 4C-E). These results revealed that
miR-125b-5p negatively regulated ASCT2 expression in the
LPS-induced intestinal mucosa cell injury model.
miR-125b-5p inhibits cell
proliferation and induces apoptosis in LPS-treated CCC-HIE-2 cells
by regulating ASCT2
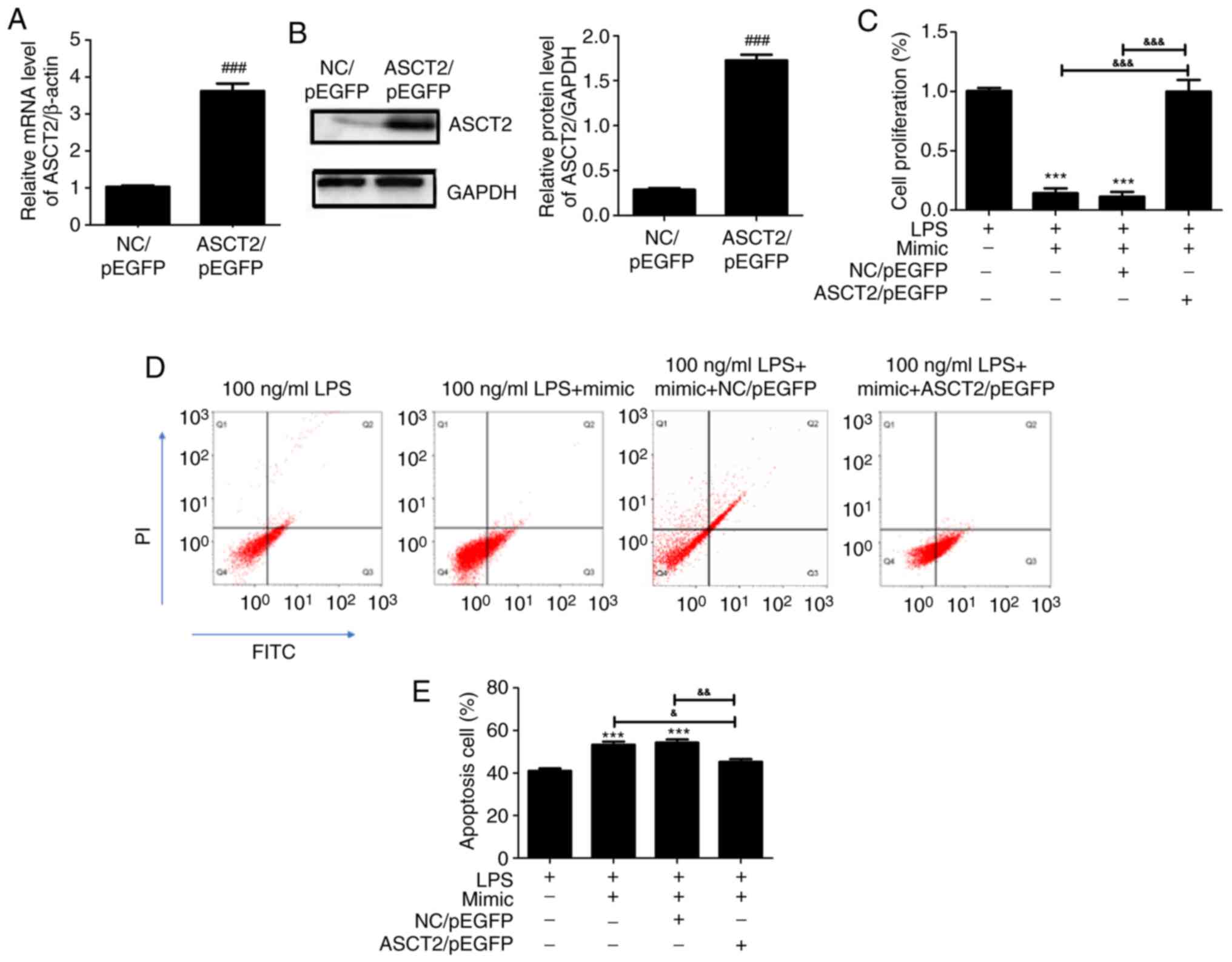
To clarify whether the upregulation of ASCT2 could
rescue the miR-125b-5p-mediated effects, an ASCT2/pEGFP plasmid was
transfected into LPS-treated CCC-HIE-2 cells. Compared with the
empty vector control (NC/pEGFP), the mRNA (Fig. 5A) and protein (Fig. 5B) expression levels of ASCT2 were
significantly increased following transfection with the ASCT2/pEGFP
plasmid. As presented in Fig. 5C,
the miR-125b-5p mimic inhibited LPS-treated CCC-HIE-2 cell
proliferation, and this effect was reversed by co-transfection with
the miR-125b-5p mimic and the ASCT2/pEGFP plasmid (Fig. 5C). Furthermore, cell apoptosis was
promoted by the miR-125b-5p mimic in LPS-treated CCC-HIE-2 cells,
but this was partially rescued by co-transfection with the
miR-125b-5p mimic and the ASCT2/pEGFP plasmid (Fig. 5D and E). These findings demonstrated that ASCT2
overexpression could alleviate LPS-induced intestinal mucosal cell
injury. The present study demonstrated a negative correlation
between ASCT2 and miR-125b-5p expression in LPS-induced intestinal
mucosa cell injury; of note, ASCT2 was involved in the
miR-125b-5p-mediated inhibition of LPS-treated CCC-HIE-2 cell
progression.
miR-125b-5p inhibits the PI3K/AKT/mTOR
signaling pathway in the LPS-induced intestinal mucosa cell injury
model
Previous studies have demonstrated that the mTOR
pathway has a mechanistic role in the development of intestinal
epithelial injury and repair (34,35).
Therefore, the present study investigated the expression levels of
PI3K/AKT/mTOR pathway-related proteins to further explore the
possible effects of miR-125b-5p on the LPS-induced intestinal
mucosa cell injury model. PI3K, p-PI3K, AKT, p-AKT, mTOR and p-mTOR
protein expression levels in LPS-treated CCC-HIE-2 cells were
detected by western blot. analysis Transfection with the
miR-125b-5p mimic significantly decreased PI3K, AKT and mTOR
phosphorylation levels (Fig. 6).
Notably, ASCT2 overexpression partially reversed this effect
(Fig. 6). Therefore, miR-125b-5p
inhibited the PI3K/AKT/mTOR signaling pathway in the LPS-induced
intestinal mucosa cell injury model. This finding suggested that
miR-125b-5p may regulate LPS-induced intestinal mucosa cell injury
through the PI3K/AKT/mTOR signaling pathway.
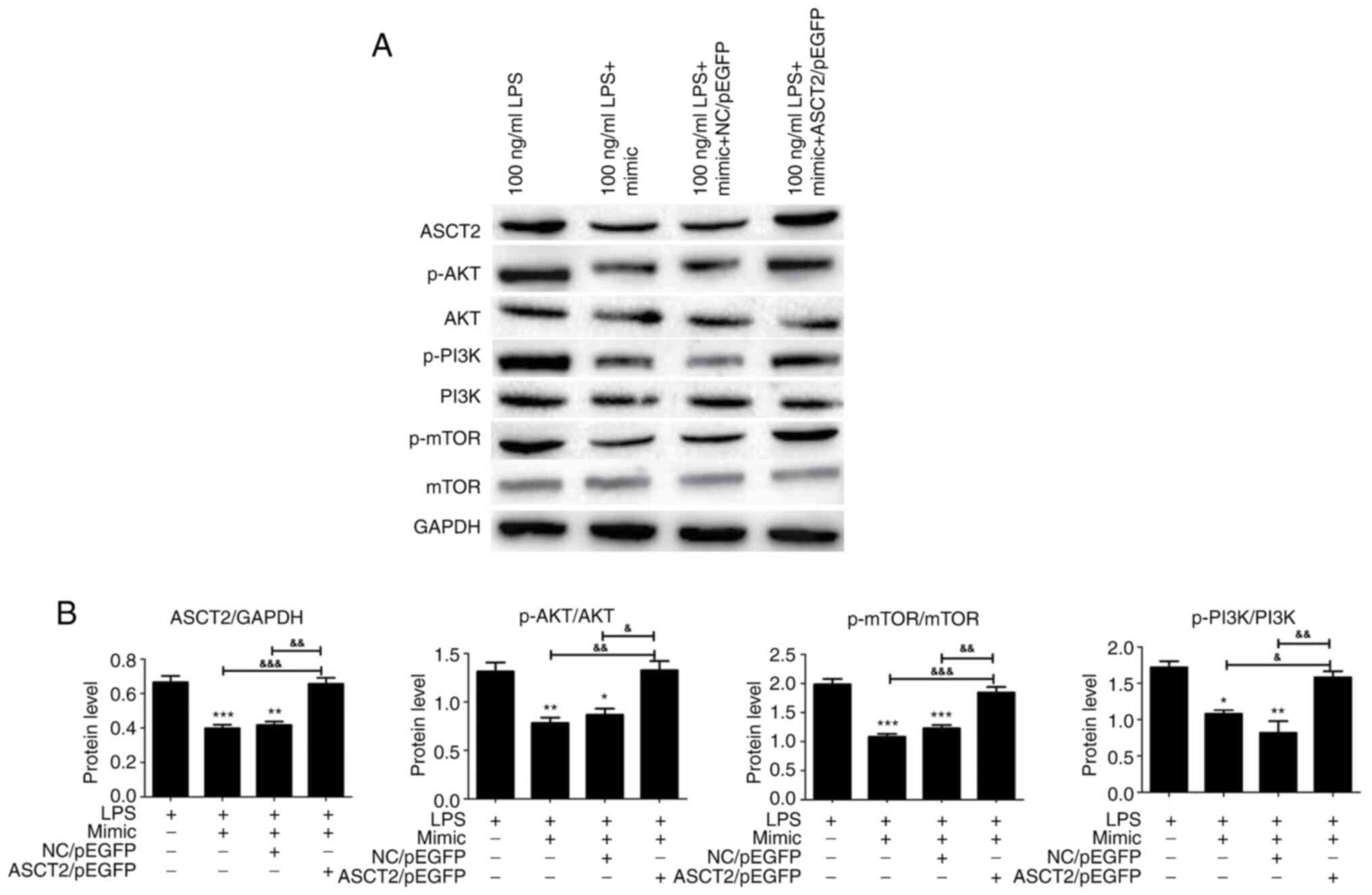 | Figure 6PI3K/AKT/mTOR pathway expression in
the miR-125b-5p and ASCT2-transfected LPS-induced intestinal mucosa
cell injury model. CCC-HIE-2 cells were transfected with the
miR-125b-5p mimic alone, the miR-125b-5p mimic + NC/pEGFP, or the
miR-125b-5p mimic + ASCT2/pEGFP for 24 h following 48 h treatment
with LPS, and the expression levels of proteins involved in the
PI3K/AKT/mTOR pathway were detected by western blot analysis. (A)
Representative blot images. (B) Quantitative western blot results.
Data are expressed as the mean ± SD (n=5). *P<0.05,
**P<0.01, ***P<0.001 vs. LPS-alone;
&P<0.05, &&P<0.01,
&&&P<0.001. miR, microRNA; ASCT2, alanine
serine cysteine-preferring transporter 2; LPS, lipopolysaccharide;
NC, negative control; p-, phosphorylated. |
Discussion
Serious injury to the intestinal barrier leads to
microbial heterotopia, which is common in patients with HIV/AIDS.
Microbial products, such as LPS and bacterial 16S RNA, have been
detected in the blood of AIDS patients since the early days of the
epidemic (36-38).
Blood LPS concentrations were detected in AIDS patients and healthy
controls in the present study and the results revealed that the
blood LPS concentration of AIDS patients was higher compared with
that of healthy controls (data not shown). Thus, an in vitro
intestinal mucosal cell injury model was established for the
present study with LPS treatment.
miRNAs are major regulators of cell function and
homeostasis (39). miRNAs are
involved in many physiological and pathological processes,
including cell differentiation, proliferation, signal transduction
and apoptosis, and their abnormal expression is closely related to
the occurrence and development of many diseases (40). miR-122a is implicated in the
regulation of intestinal epithelial tight junctions during
inflammation, and TNF-α induces the epithelial expression of
miR-122a, thereby causing the degradation of its target gene
occludin and resulting in increased mucosal permeability (41). miRNAs serve an important role in the
replication of HIV-infected viruses (42). The JunD gene inhibits proliferation
of intestinal epithelial cells by increasing miR-29b levels
transcriptionally and post-transcriptionally (43). Because miRNAs are highly conserved
and have good stability in the extracellular domain, miRNAs could
serve as markers for disease diagnosis. With the deepening of the
study on the related role of miRNAs in human physiological
diseases, the potential of miRNAs as a drug target has emerged and
become a research hotspot. However, studies on miRNAs in the repair
of intestinal barrier injury in HIV/AIDS patients are rarely
reported.
The present study found that ASCT2 expression in
colon samples from patients with HIV/AIDS was downregulated and
negatively correlated with miR-125b-5p expression. Huang et
al (21) reported that
miR-125b-5p targets the 3' long terminal repeat region of the HIV
genome, leading to differential HIV-1 infectivity/resistance to
different cells in various developmental stages. The present study
established an LPS-induced CCC-HIE-2 cell injury model and examined
the effects of miR-125b-5p and ASCT2 on LPS-induced intestinal
mucosal injury in vitro. Dual luciferase reporter assays
were used to identify ASCT2 as a direct target of miR-125b-5p in
intestinal mucosa injury cells, suggesting that the suppression of
ASCT2 by miR-125b-5p may be a novel therapeutic approach for
intestinal mucosa injury in patients with HIV/AIDS.
In addition, the current results confirmed that the
effect of miR-125b-5p and ASCT2 on intestinal mucosa injury may be
related to the PI3K/AKT/mTOR pathway. A previous study indicated
that elevated ASCT2 expression promoted lung cell growth and
survival through mTOR signaling (44). Activation of the PI3K/AKT signaling
pathway inhibits cell apoptosis and facilitates the protective
effect of magnesium sulfate against intestinal ischemia-reperfusion
injury (45). Therefore, the
present study focused on the PI3K/AKT/mTOR signaling pathway when
assessing the effects of miR-125b-5p and ASCT2 in the intestinal
mucosa injury model. The results indicated that miR-125b-5p
upregulation decreased the expression levels of p-AKT, p-PI3K, and
p-mTOR in LPS-induced intestinal mucosa cells, whereas ASCT2
overexpression promoted p-AKT, p-PI3K, and p-mTOR expression. The
PI3K/AKT/mTOR pathway is involved in intestinal epithelial barrier
dysfunction following severe burn (46). The levels of p-PI3K, p-AKT, and
p-mTOR were decreased under miR-125b-5p treatment and increased
following ASCT2 overexpression, indicating that miR-125b-5p
downregulated ASCT2 expression by targeting the 3'-UTR of ASCT2 and
then promoted the injury of LPS-induced intestinal mucosa cells
through the PI3K/AKT/mTOR pathway.
In conclusion, the present study demonstrated that
miR-125b-5p overexpression inhibited growth in the LPS-induced
intestinal mucosal cell model by targeting ASCT2 expression and
inhibiting the PI3K/AKT/mTOR pathway. By contrast, the inhibition
of miR-125b-5p alleviated LPS-induced intestinal mucosal cell
injury by increasing ASCT2 expression and promoting the
PI3K/AKT/mTOR pathway. The present study was the first to
investigate the relationship between miR-125b-5p and ASCT2 in
LPS-induced intestinal mucosa cells and provided new insight into
the molecular mechanisms of intestinal barrier injury in HIV/AIDS
progression. These findings may indicate a path towards a novel
treatment for intestinal barrier injury in patients with
HIV/AIDS.
Acknowledgements
Not applicable.
Funding
Funding: This work was supported by the National Scientific
Foundation of China (grant no. 81870458), the Special Project of
Yunnan Science and Technology Department-Kunming Medical University
Applied Basic Research [grant no. 2019FE001(-196)], the Yunnan
Engineering Technology Center of Digestive Disease (grant no.
2018DH006), and the Project for Innovation Team of Yunnan
Provincial Science and Technology Department (grant no.
2018HC005).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
HG and KW conceived and designed the study. JG, YQ,
HW and JL performed the experiments. HG, QP and YZ processed data.
HG, JG and KW wrote the manuscript. HG and KW reviewed and edited
the manuscript. HG and KW confirm the authenticity of all the raw
data. All authors read and approved the manuscript.
Ethics approval and consent to
participate
All human tissue samples were collected at the First
Affiliated Hospital of Kunming Medical University, and written
informed consent was obtained from all patients. All methods were
approved by the Research Medical Ethics Committee of Kunming
Medical University (Kunming, China) and were performed in
accordance with the approved guidelines.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Druml W: Intestinaler Crosstalk. Med Klin
Intensivmed Notfmed. 113:470–477. 2018.PubMed/NCBI View Article : Google Scholar : (In German).
|
|
2
|
Groschwitz KR and Hogan SP: Intestinal
barrier function: Molecular regulation and disease pathogenesis. J
Allergy Clin Immunol. 124:3–20; quiz 21-22. 2009.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Hermiston ML and Gordon JI: Inflammatory
bowel disease and adenomas in mice expressing a dominant negative
N-cadherin. Science. 270:1203–1207. 1995.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Costantini TW, Peterson CY, Kroll L,
Loomis WH, Eliceiri BP, Baird A, Bansal V and Coimbra R: Role of
p38 MAPK in burn-induced intestinal barrier breakdown. J Surg Res.
156:64–69. 2009.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Yasuda T, Takeyama Y, Ueda T, Shinzeki M,
Sawa H, Nakajima T and Kuroda Y: Breakdown of intestinal mucosa via
accelerated apoptosis increases intestinal permeability in
experimental severe acute pancreatitis. J Surg Res. 135:18–26.
2006.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Sabatino A, Regolisti G, Cosola C,
Gesualdo L and Fiaccadori E: Intestinal microbiota in type 2
diabetes and chronic kidney disease. Curr Diab Rep.
17(16)2017.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Vyboh K, Jenabian MA, Mehraj V and Routy
JP: HIV and the gut microbiota, partners in crime: Breaking the
vicious cycle to unearth new therapeutic targets. J Immunol Res.
2015(614127)2015.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Nazli A, Chan O, Dobson-Belaire WN,
Ouellet M, Tremblay MJ, Gray-Owen SD, Arsenault AL and Kaushic C:
Exposure to HIV-1 directly impairs mucosal epithelial barrier
integrity allowing microbial translocation. PLoS Pathog.
6(e1000852)2010.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Kristoff J, Haret-Richter G, Ma D, Ribeiro
RM, Xu C, Cornell E, Stock JL, He T, Mobley AD, Ross S, et al:
Early microbial translocation blockade reduces SIV-mediated
inflammation and viral replication. J Clin Invest. 124:2802–2806.
2014.PubMed/NCBI View
Article : Google Scholar
|
|
10
|
Smith AJ, Schacker TW, Reilly CS and Haase
AT: A role for syndecan 1 and claudin 2 in microbial translocation
during HIV 1 infection. J Acquir Immune Defic Syndr. 55:306–315.
2010.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Cicala C, Arthos J and Fauci AS: Role of
T-cell trafficking in the pathogenesis of HIV disease. Curr Opin
HIV AIDS. 14:115–120. 2019.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Jenabian MA, El-Far M, Vyboh K, Kema I,
Costiniuk CT, Thomas R, Baril JG, LeBlanc R, Kanagaratham C,
Radzioch D, et al: Montreal Primary infection and Slow Progressor
Study Groups: Immunosuppressive tryptophan catabolism and gut
mucosal dysfunction following early HIV infection. J Infect Dis.
212:355–366. 2015.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Sharp PM and Hahn BH: Origins of HIV and
the AIDS pandemic. Cold Spring Harb Perspect Med.
1(a006841)2011.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Faria NR, Rambaut A, Suchard MA, Baele G,
Bedford T, Ward MJ, Tatem AJ, Sousa JD, Arinaminpathy N, Pépin J,
et al: HIV epidemiology. The early spread and epidemic ignition of
HIV-1 in human populations. Science. 346:56–61. 2014.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Worobey M, Watts TD, McKay RA, Suchard MA,
Granade T, Teuwen DE, Koblin BA, Heneine W, Lemey P and Jaffe HW:
1970s and ‘Patient 0’ HIV-1 genomes illuminate early HIV/AIDS
history in North America. Nature. 539:98–101. 2016.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Marchetti G, Tincati C and Silvestri G:
Microbial translocation in the pathogenesis of HIV infection and
AIDS. Clin Microbiol Rev. 26:2–18. 2013.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Lu TX and Rothenberg ME: MicroRNA. J
Allergy Clin Immunol. 141:1202–1207. 2018.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Yang Y, Ma Y, Shi C, Chen H, Zhang H, Chen
N, Zhang P, Wang F, Yang J, Yang J, et al: Overexpression of miR-21
in patients with ulcerative colitis impairs intestinal epithelial
barrier function through targeting the Rho GTPase RhoB. Biochem
Biophys Res Commun. 434:746–752. 2013.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Zhao Y, Ma T, Chen W, Chen Y, Li M, Ren L,
Chen J, Cao R, Feng Y, Zhang H, et al: MicroRNA-124 promotes
intestinal inflammation by targeting aryl hydrocarbon receptor in
Crohn's Disease. J Crohn's Colitis. 10:703–712. 2016.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Chassin C, Kocur M, Pott J, Duerr CU,
Gütle D, Lotz M and Hornef MW: miR-146a mediates protective innate
immune tolerance in the neonate intestine. Cell Host Microbe.
8:358–368. 2010.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Huang J, Wang F, Argyris E, Chen K, Liang
Z, Tian H, Huang W, Squires K, Verlinghieri G and Zhang H: Cellular
microRNAs contribute to HIV-1 latency in resting primary CD4+ T
lymphocytes. Nat Med. 13:1241–1247. 2007.PubMed/NCBI View
Article : Google Scholar
|
|
22
|
Xu Y, Wang HW, Luo HY, Shu R, Liu J, Sun
L, Han XF, Lin N, Wang TH, Zeng YJ, et al: MicroRNA expression
profiling of intestinal mucosa tissue predicts multiple crucial
regulatory molecules and signaling pathways for gut barrier
dysfunction of AIDS patients. Mol Med Rep. 16:8854–8862.
2017.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Ducroc R, Sakar Y, Fanjul C, Barber A,
Bado A and Lostao MP: Luminal leptin inhibits L-glutamine transport
in rat small intestine: Involvement of ASCT2 and B0AT1. Am J
Physiol Gastrointest Liver Physiol. 299:G179–G185. 2010.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Akgün Şahin Z and Dayapoğlu N: Effect of
progressive relaxation exercises on fatigue and sleep quality in
patients with chronic obstructive lung disease (COPD). Complement
Ther Clin Pract. 21:277–281. 2015.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Kekuda R, Prasad PD, Fei YJ,
Torres-Zamorano V, Sinha S, Yang-Feng TL, Leibach FH and Ganapathy
V: Cloning of the sodium-dependent, broad-scope, neutral amino acid
transporter Bo from a human placental choriocarcinoma cell line. J
Biol Chem. 271:18657–18661. 1996.PubMed/NCBI View Article : Google Scholar
|
|
26
|
van Geldermalsen M, Wang Q, Nagarajah R,
Marshall AD, Thoeng A, Gao D, Ritchie W, Feng Y, Bailey CG, Deng N,
et al: ASCT2/SLC1A5 controls glutamine uptake and tumour growth in
triple-negative basal-like breast cancer. Oncogene. 35:3201–3208.
2016.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Cai C, Zeng B, Zeng J, Xin H and Tang C:
Effect of ASCT2 gene knock-down by shRNA on biological behaviors of
colorectal cancer cells. Zhonghua Wei Chang Wai Ke Za Zhi.
20:450–454. 2017.PubMed/NCBI(In Chinese).
|
|
28
|
Sun HW, Yu XJ, Wu WC, Chen J, Shi M, Zheng
L and Xu J: GLUT1 and ASCT2 as predictors for prognosis of
hepatocellular carcinoma. PLoS One. 11(e0168907)2016.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Kasai N, Sasakawa A, Hosomi K, Poh TW,
Chua BL, Yong WP, So J, Chan SL, Soong R, Kono K, et al: Anti-tumor
efficacy evaluation of a novel monoclonal antibody targeting
neutral amino acid transporter ASCT2 using patient-derived
xenograft mouse models of gastric cancer. Am J Transl Res.
9:3399–3410. 2017.PubMed/NCBI
|
|
30
|
Guo H, Xu Y, Wang F, Shen Z, Tuo X, Qian
H, Wang H and Wang K: Clinical associations between ASCT2 and p
mTOR in the pathogenesis and prognosis of epithelial ovarian
cancer. Oncol Rep. 40:3725–3733. 2018.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Aids group Soid. Chinese medical
association: Guideline of diagnosis and treatment for aids. Chin J
Infect Dis. 29:629–640. 2011.
|
|
32
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) Method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Agarwal V, Bell GW, Nam JW and Bartel DP:
Predicting effective microRNA target sites in mammalian mRNAs.
eLife. 4(4)2015.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Rhoads JM, Niu X, Odle J and Graves LM:
Role of mTOR signaling in intestinal cell migration. Am J Physiol
Gastrointest Liver Physiol. 291:G510–G517. 2006.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Sampson LL, Davis AK, Grogg MW and Zheng
Y: mTOR disruption causes intestinal epithelial cell defects and
intestinal atrophy postinjury in mice. FASEB J. 30:1263–1275.
2016.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Duprez DA, Kuller LH, Tracy R, Otvos J,
Cooper DA, Hoy J, Neuhaus J, Paton NI, Friis-Moller N, Lampe F, et
al: INSIGHT SMART Study Group: Lipoprotein particle subclasses,
cardiovascular disease and HIV infection. Atherosclerosis.
207:524–529. 2009.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Aqil M, Mallik S, Bandyopadhyay S, Maulik
U and Jameel S: Transcriptomic analysis of mRNAs in human monocytic
cells expressing the HIV-1 Nef protein and their exosomes. BioMed
Res Int. 2015(492395)2015.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Estes JD, Harris LD, Klatt NR, Tabb B,
Pittaluga S, Paiardini M, Barclay GR, Smedley J, Pung R, Oliveira
KM, et al: Damaged intestinal epithelial integrity linked to
microbial translocation in pathogenic simian immunodeficiency virus
infections. PLoS Pathog. 6(e1001052)2010.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Tili E, Michaille JJ, Piurowski V, Rigot B
and Croce CM: MicroRNAs in intestinal barrier function,
inflammatory bowel disease and related cancers-their effects and
therapeutic potentials. Curr Opin Pharmacol. 37:142–150.
2017.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Poltronieri P, Sun B and Mallardo M: RNA
Viruses: RNA roles in pathogenesis, coreplication and viral load.
Curr Genomics. 16:327–335. 2015.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Ye D, Guo S, Al-Sadi R and Ma TY: MicroRNA
regulation of intestinal epithelial tight junction permeability.
Gastroenterology. 141:1323–1333. 2011.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Vongrad V, Imig J, Mohammadi P, Kishore S,
Jaskiewicz L, Hall J, Günthard HF, Beerenwinkel N and Metzner KJ:
HIV-1 RNAs are not part of the Argonaute 2 associated RNA
interference pathway in macrophages. PLoS One.
10(e0132127)2015.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Zou T, Rao JN, Liu L, Xiao L, Chung HK, Li
Y, Chen G, Gorospe M and Wang JY: JunD enhances miR-29b levels
transcriptionally and posttranscriptionally to inhibit
proliferation of intestinal epithelial cells. Am J Physiol Cell
Physiol. 308:C813–C824. 2015.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Hassanein M, Hoeksema MD, Shiota M, Qian
J, Harris BK, Chen H, Clark JE, Alborn WE, Eisenberg R and Massion
PP: SLC1A5 mediates glutamine transport required for lung cancer
cell growth and survival. Clin Cancer Res. 19:560–570.
2013.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Chen SD, Chen YB, Peng Y, Xu J, Chen SS,
Zhang JL, Li ZZ and Tan Z: Role of PI3K/Akt signaling in the
protective effect of magnesium sulfate against ischemia-perfusion
injury of small intestine in rats. Chin Med J (Engl).
123:1447–1452. 2010.PubMed/NCBI
|
|
46
|
Huang Y, Feng Y, Wang Y, Wang P, Wang F
and Ren H: Severe burn-induced intestinal epithelial barrier
dysfunction is associated with endoplasmic reticulum stress and
autophagy in mice. Front Physiol. 9(441)2018.PubMed/NCBI View Article : Google Scholar
|